Origanum majorana Essential Oil Triggers p38 MAPK-Mediated Protective Autophagy, Apoptosis, and Caspase-Dependent Cleavage of P70S6K in Colorectal Cancer Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Origanum majorana Essential Oil
2.2. Cell Culture, Chemicals, and Antibodies
2.3. Measurement of Cellular Viability
2.4. HPLC-MS
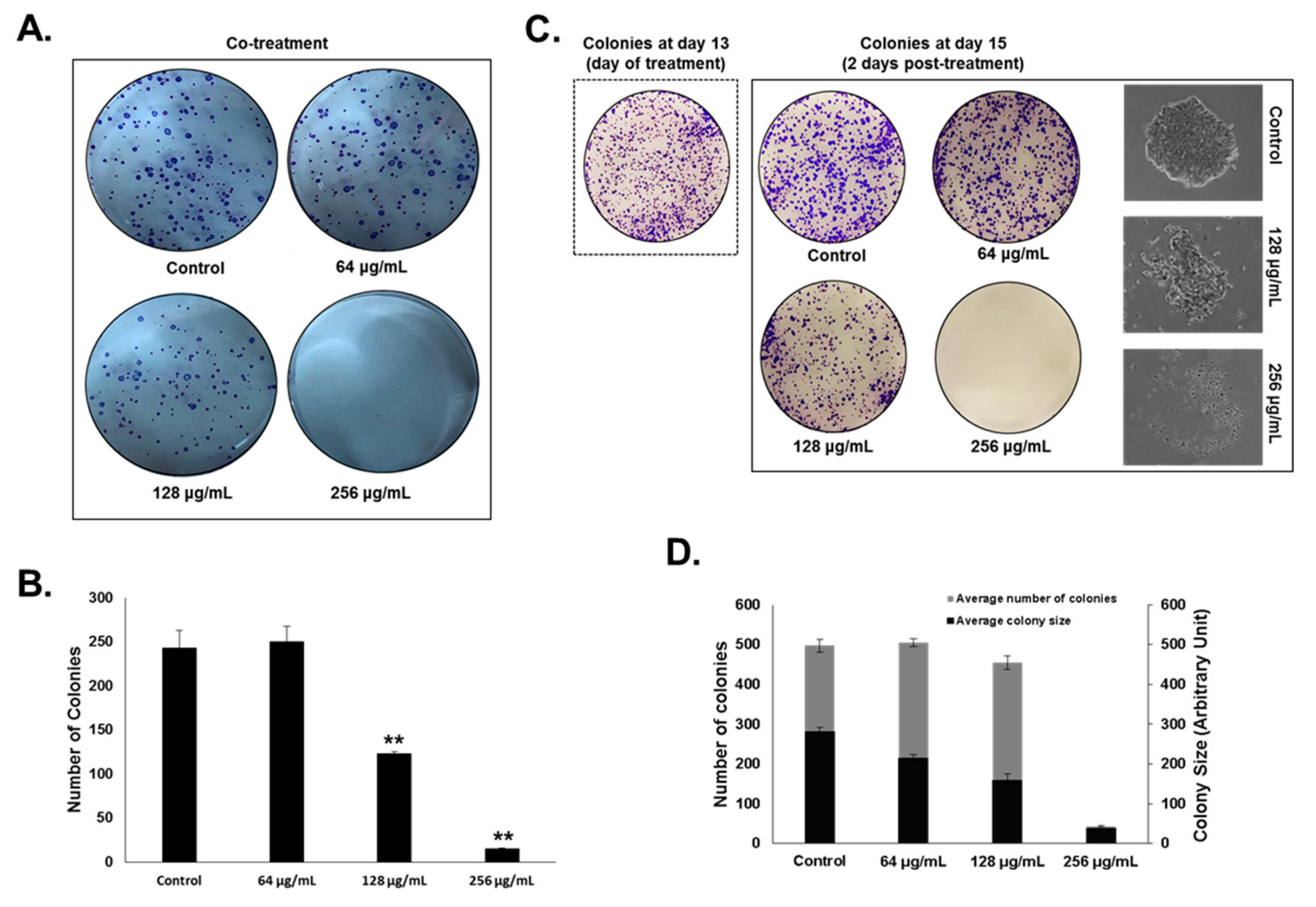
2.5. Colony Formation Assay
2.6. Whole Cell Extract and Western Blot Analysis
2.7. Statistical Analysis
3. Results
3.1. HPLC-MS Identification of Constituents in Origanum majorana Essential Oil
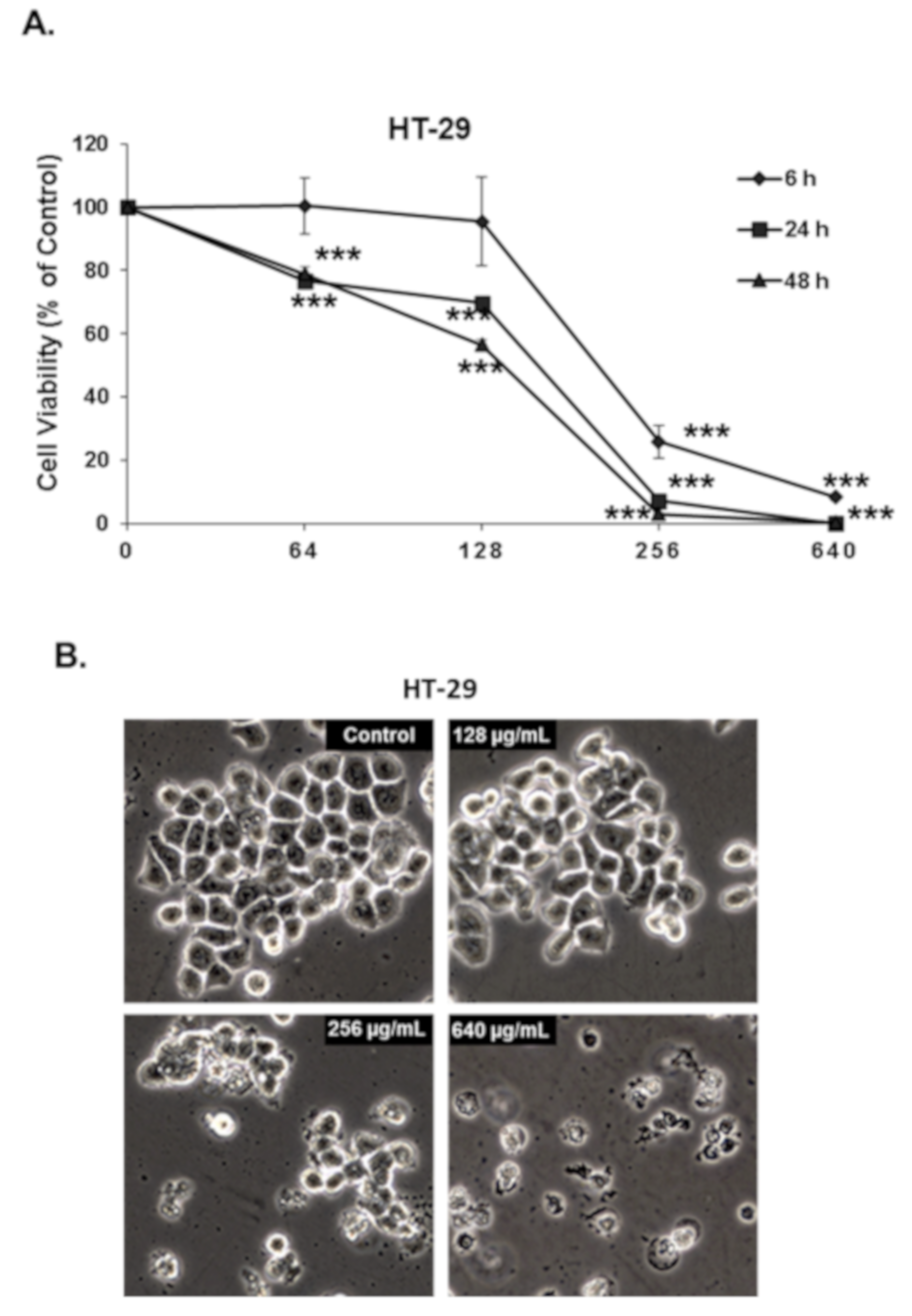
3.2. Origanum majorana Essential Oil Inhibits the Cellular Viability and Colony Growth of Human Colorectal Cancer Cells
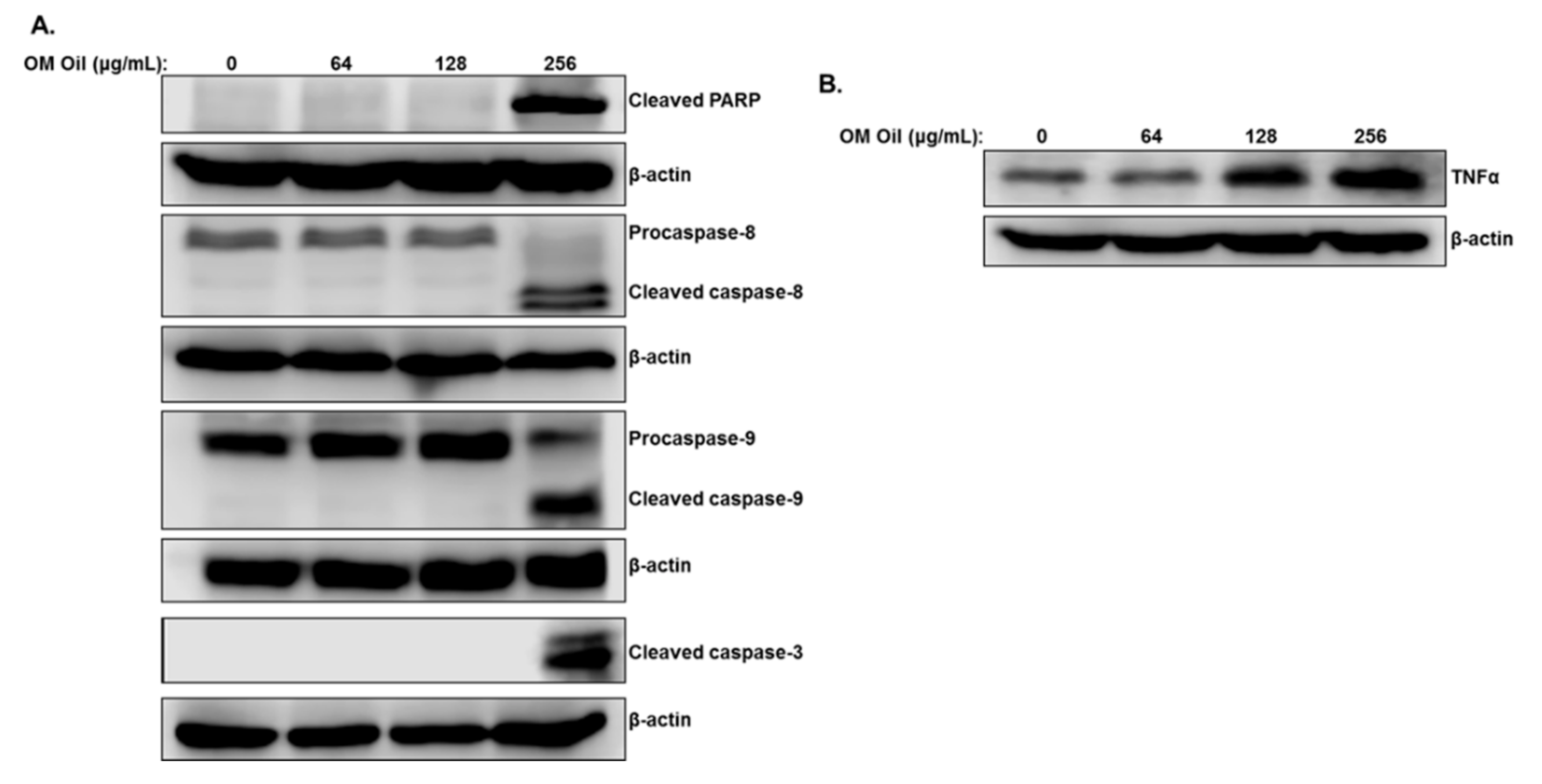
3.3. Origanum majorana Essential Oil Induces Apoptotic Cell Death of Colon Cancer Cells through Activation of the Extrinsic and Intrinsic Pathways
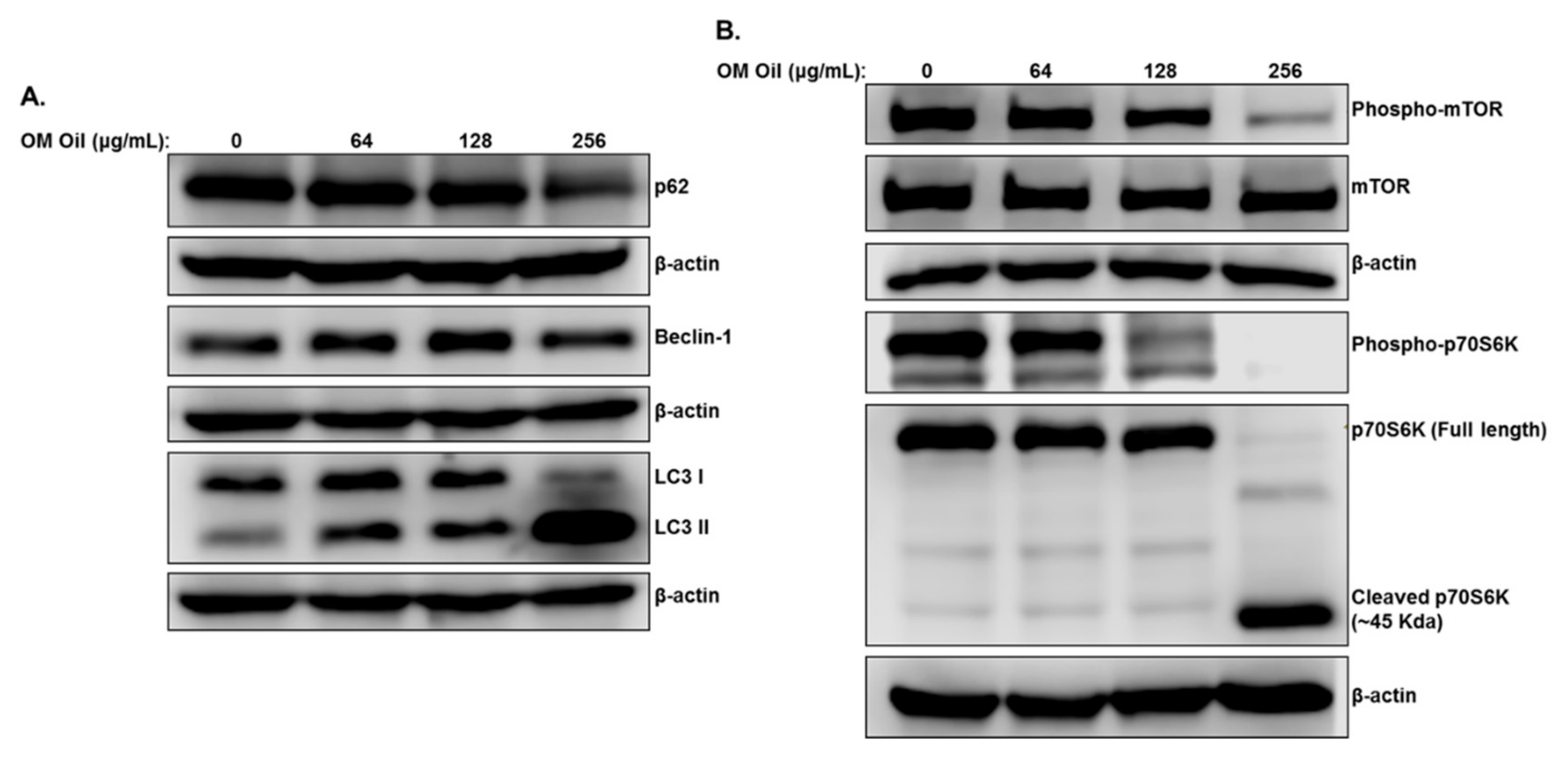
3.4. OMEO Triggers Autophagy through Downregulation of the mTOR/p70S6K Signaling Pathway
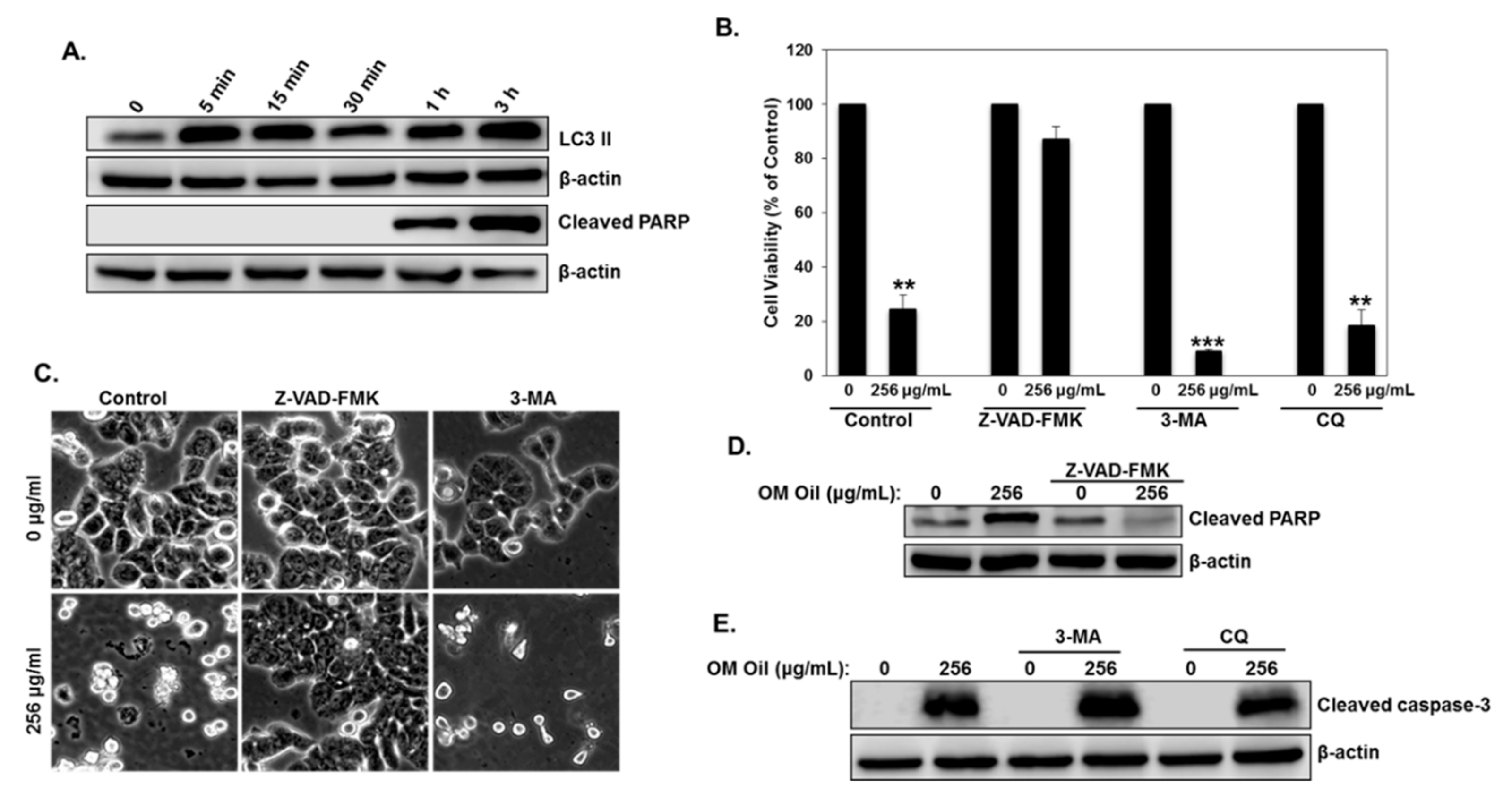
3.5. Origanum majorana Essential Oil Induces Protective Autophagy and Apoptotic Cell Death
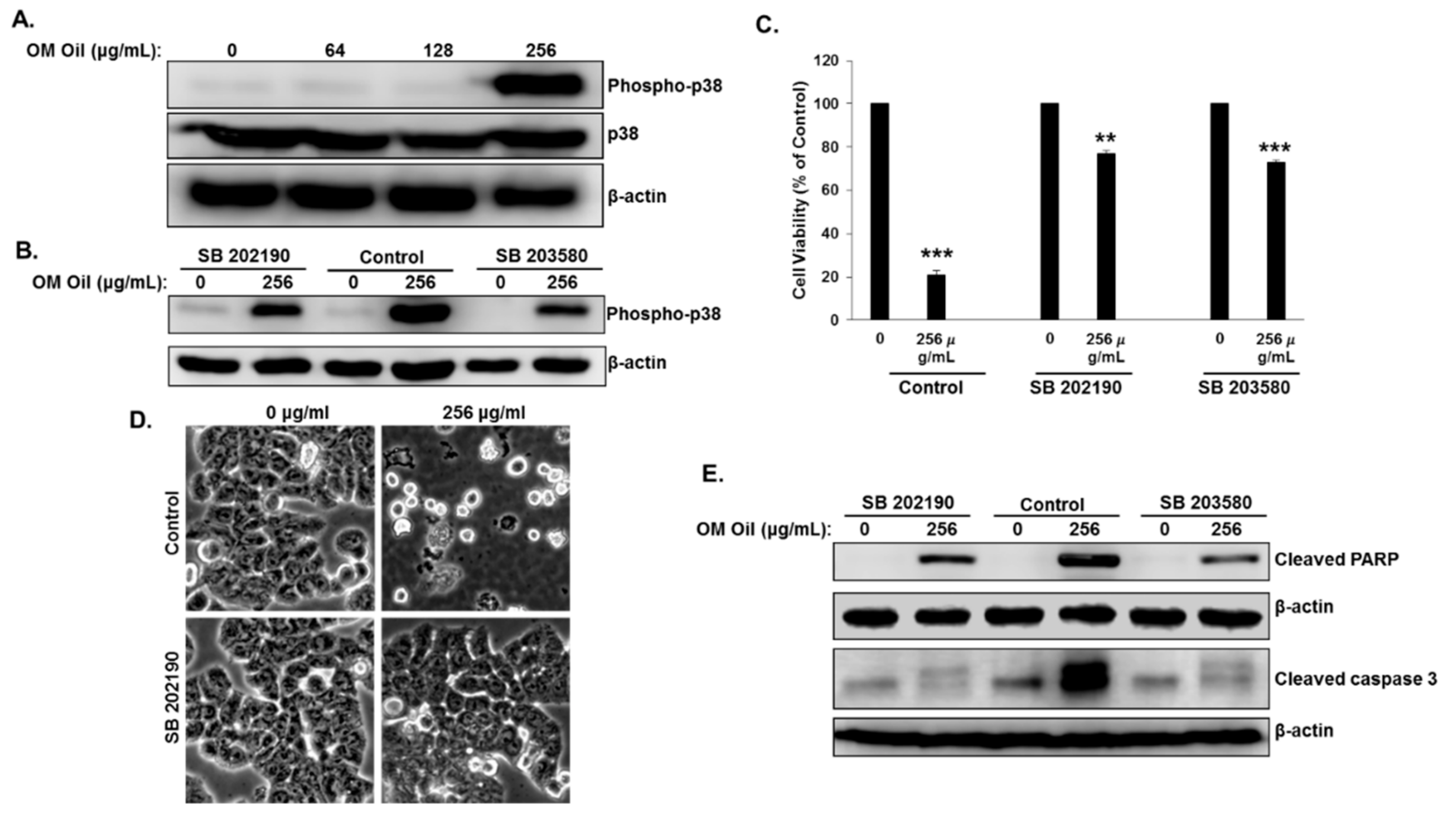
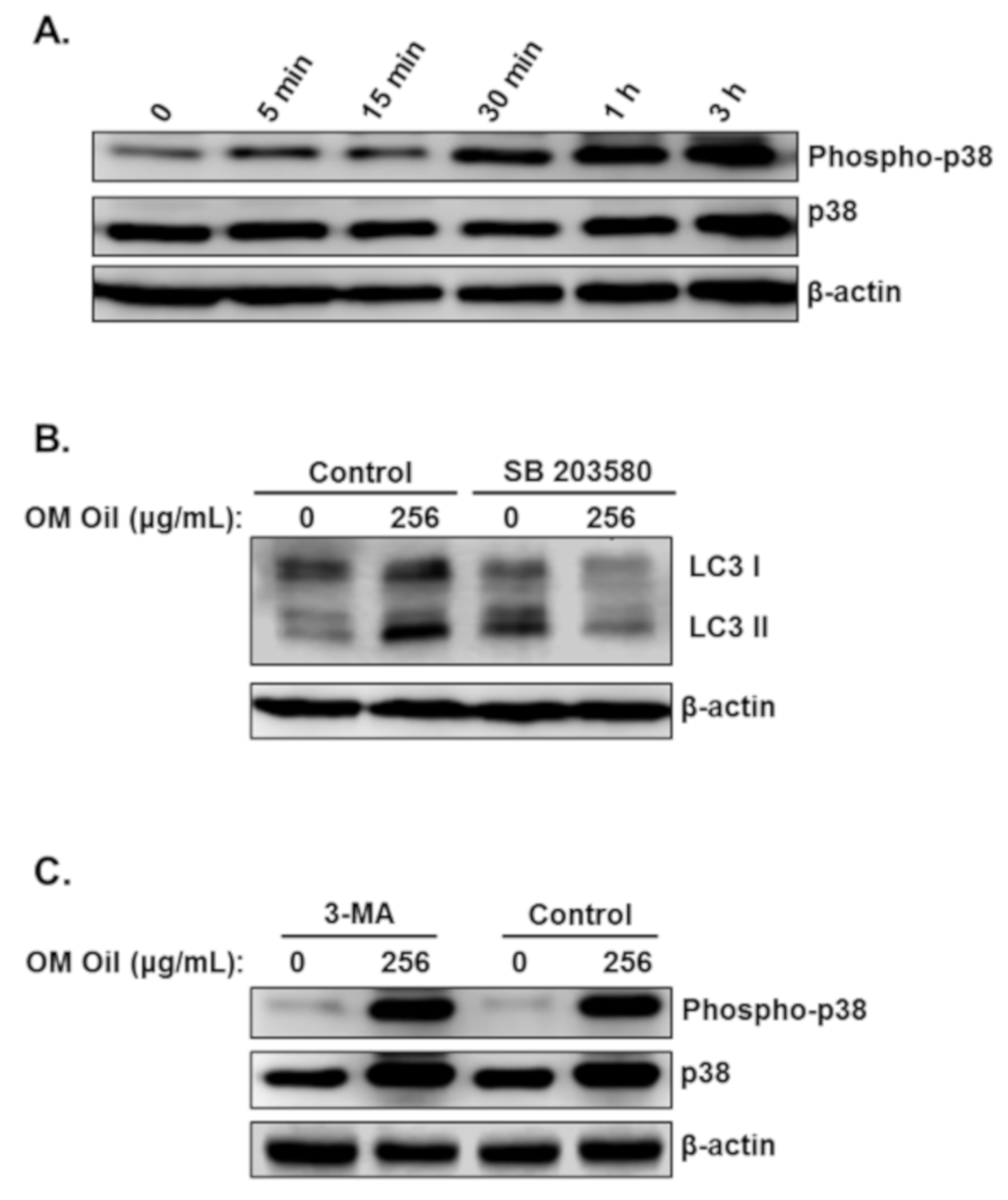
3.6. OMEO Induces p38 MAPK-Mediated Protective Autophagy and Apoptosis
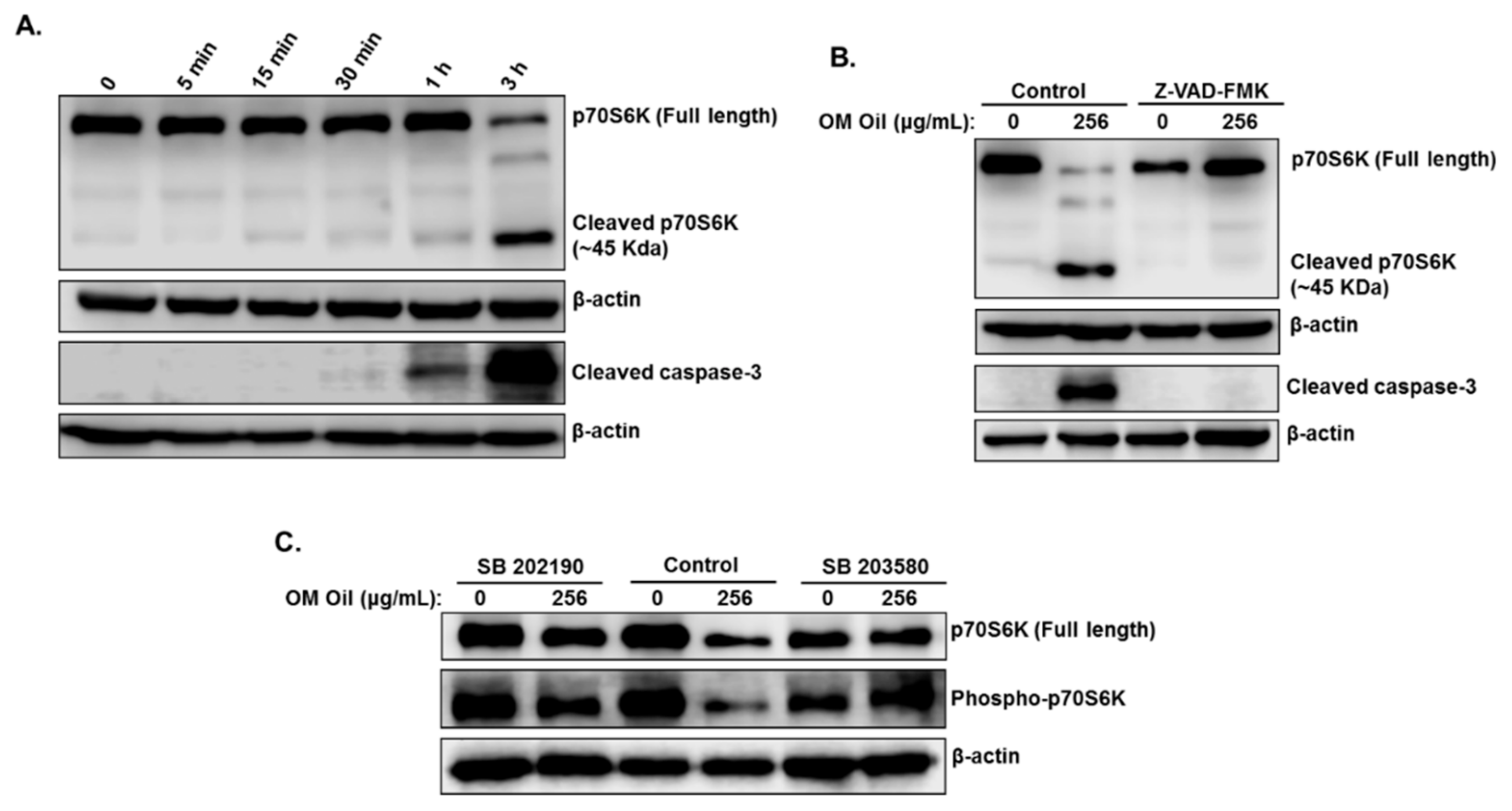
3.7. OMEO Induces p38 MAPK-Mediated Caspase-Dependent Cleavage of p70S6K.
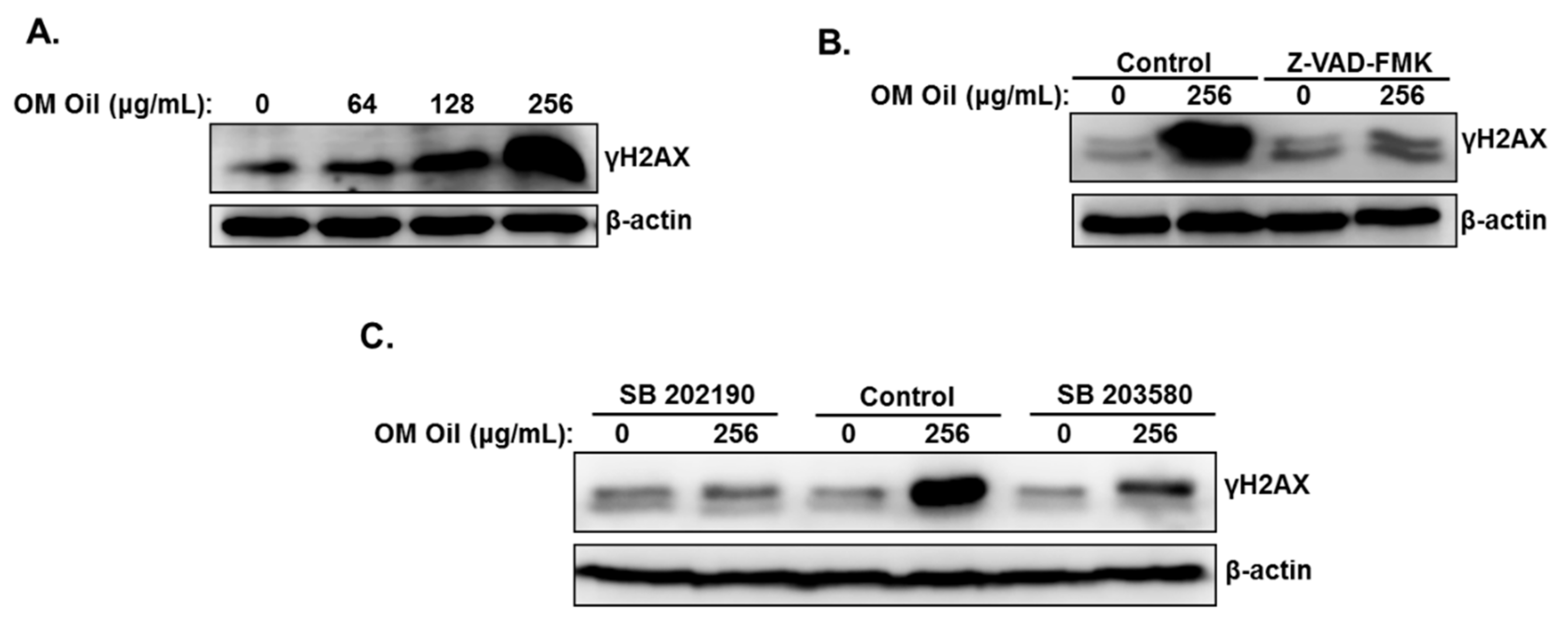
3.8. DNA Damage-Independent Activation of p38 MAPK
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Kuipers, E.J.; Grady, W.M.; Lieberman, D.; Seufferlein, T.; Sung, J.J.; Boelens, P.G.; van de Velde, C.J.H.; Watanabe, T. Colorectal cancer. Nat. Rev. Dis. Primers 2015, 1, 15065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peluso, G.; Incollingo, P.; Calogero, A.; Tammaro, V.; Rupealta, N.; Chiacchio, G.; Sandoval Sotelo, M.L.; Minieri, G.; Pisani, A.; Riccio, E.; et al. Current Tissue Molecular Markers in Colorectal Cancer: A Literature Review. Biomed. Res. Int. 2017, 2017, 2605628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balunas, M.J.; Kinghorn, A.D. Drug discovery from medicinal plants. Life Sci. 2005, 78, 431–441. [Google Scholar] [CrossRef]
- Nobili, S.; Lippi, D.; Witort, E.; Donnini, M.; Bausi, L.; Mini, E.; Capaccioli, S. Natural compounds for cancer treatment and prevention. Pharmacol. Res. Off. J. Ital. Pharmacol. Soc. 2009, 59, 365–378. [Google Scholar] [CrossRef]
- Petrovska, B. Historical review of medicinal plants’ usage. Pharmacogn. Rev. 2012, 6, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.; Dwivedi, J.; Jain, P.K.; Satpathy, S.; Patra, A. Medicinal Plants for Treatment of Cancer: A Brief Review. Pharmacogn. J. 2015, 8, 87–102. [Google Scholar] [CrossRef] [Green Version]
- Raut, J.S.; Karuppayil, S.M. A status review on the medicinal properties of essential oils. Ind. Crop. Prod. 2014, 62, 250–264. [Google Scholar] [CrossRef]
- Adorjan, B.; Buchbauer, G. Biological properties of essential oils: An updated review. Flavour Fragr. J. 2010, 25, 407–426. [Google Scholar] [CrossRef]
- Tasdemir, D.; Kaiser, M.; Demirci, B.; Demirci, F.; Baser, K.H.C. Antiprotozoal Activity of Turkish Origanum onites Essential Oil and Its Components. Molecules 2019, 24, 4421. [Google Scholar] [CrossRef] [Green Version]
- Rajkumar, S.; Jebanesan, A. Repellent activity of selected plant essential oils the malarial fever mosquito Anopheles stephensi. Trop. Biomed. 2008, 24, 71–75. [Google Scholar]
- Goel, P.; Vasudeva, N. Origanum majorana L. -Phyto-pharmacological review. J. Essent. Oil Bear. Plants 2015, 6, 261–267. [Google Scholar]
- Duletic, S.; Alimpić Aradski, A.; Kolarevic, S.; Vuković-Gačić, B.; Oaldje, M.; Živković, J.; Šavikin, K.; Marin, P. Antineurodegenerative, antioxidant and antibacterial activities and phenolic components of Origanum majorana L. (Lamiaceae) extracts. J. Appl. Bot. Food Qual. 2018, 91. [Google Scholar]
- Leeja, L.; Thoppil, J. Antimicrobial activity of methanol extract of Origanum majorana L. (Sweet marjoram). J. Environ. Biol. Acad. Environ. Biol. India 2007, 28, 145–146. [Google Scholar]
- Amor, G.; Caputo, L.; La Storia, A.; De Feo, V.; Mauriello, G.; Fechtali, T. Chemical Composition and Antimicrobial Activity of Artemisia herba-alba and Origanum majorana Essential Oils from Morocco. Molecules 2019, 24, 4021. [Google Scholar] [CrossRef] [Green Version]
- Della Pepa, T.; Elshafie, H.S.; Capasso, R.; De Feo, V.; Camele, I.; Nazzaro, F.; Scognamiglio, M.R.; Caputo, L. Antimicrobial and Phytotoxic Activity of Origanum heracleoticum and O. majorana Essential Oils Growing in Cilento (Southern Italy). Molecules 2019, 24, 2576. [Google Scholar] [CrossRef] [Green Version]
- Al Dhaheri, Y.; Attoub, S.; Arafat, K.; Abuqamar, S.; Viallet, J.; Saleh, A.; Al Agha, H.; Eid, A.; Iratni, R. Anti-metastatic and anti-tumor growth effects of Origanum majorana on highly metastatic human breast cancer cells: Inhibition of NFκB signaling and reduction of nitric oxide production. PLoS ONE 2013, 8, e68808. [Google Scholar] [CrossRef] [Green Version]
- Dhaheri, Y.; Eid, A.; AbuQamar, S.; Attoub, S.; Khasawneh, M.; Aiche, G.; Hisaindee, S.; Iratni, R. Mitotic Arrest and Apoptosis in Breast Cancer Cells Induced by Origanum majorana Extract: Upregulation of TNF-α and Downregulation of Survivin and Mutant p53. PLoS ONE 2013, 8, e56649. [Google Scholar] [CrossRef]
- Benhalilou, N.; Alsamri, H.; Alneyadi, A.; Athamneh, K.; Alrashedi, A.; Altamimi, N.; Al Dhaheri, Y.; Eid, A.H.; Iratni, R. Origanum majorana Ethanolic Extract Promotes Colorectal Cancer Cell Death by Triggering Abortive Autophagy and Activation of the Extrinsic Apoptotic Pathway. Front Oncol. 2019, 9, 795. [Google Scholar] [CrossRef]
- Yazdanparast, R.; Shahriyary, L. Comparative effects of Artemisia dracunculus, Satureja hortensis and Origanum majorana on inhibition of blood platelet adhesion, aggregation and secretion. Vasc. Pharmacol. 2008, 48, 32–37. [Google Scholar] [CrossRef]
- Soliman, A.M.; Desouky, S.; Marzouk, M.; Sayed, A.A. Origanum majorana Attenuates Nephrotoxicity of Cisplatin Anticancer Drug through Ameliorating Oxidative Stress. Nutrients 2016, 8, 264. [Google Scholar] [CrossRef] [PubMed]
- Al-Howiriny, T.; Alsheikh, A.; Alqasoumi, S.; Al-Yahya, M.; ElTahir, K.; Rafatullah, S. Protective Effect of Origanum majorana L. “Marjoram” on Various Models of Gastric Mucosal Injury in Rats. Am. J. Chin. Med. 2009, 37, 531–545. [Google Scholar] [CrossRef] [PubMed]
- Makrane, H.; Aziz, M.; Berrabah, M.; Mekhfi, H.; Ziyyat, A.; Bnouham, M.; Legssyer, A.; Elombo, F.K.; Gressier, B.; Eto, B. Myorelaxant Activity of essential oil from Origanum majorana L. on rat and rabbit. J. Ethnopharmacol. 2019, 228, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Mossa, A.-T.H.; Refaie, A.A.; Ramadan, A.; Bouajila, J. Amelioration of Prallethrin-Induced Oxidative Stress and Hepatotoxicity in Rat by the Administration of Origanum majorana Essential Oil. Biomed. Res. Int. 2013, 2013, 859085. [Google Scholar] [CrossRef] [Green Version]
- Dantas, A.D.S.; Klein-Júnior, L.C.; Machado, M.S.; Guecheva, T.N.; Santos, L.D.D.; Zanette, R.A.; Mello, F.B.D.; Pêgas Henriques, J.A.; Mello, J.R.B.D. Origanum majorana Essential Oil Lacks Mutagenic Activity in the Salmonella/Microsome and Micronucleus Assays. Sci. World J. 2016, 2016, 3694901. [Google Scholar]
- Nurzyńska-Wierdak, R.; Zawiślak, G.; Kowalski, R. The Content and Composition of Essential Oil of Origanum majorana L. Grown in Poland Depending on Harvest Tme and Method of Raw Material Preparation. J. Essent. Oil Bear. Plants 2015, 18, 1482–1489. [Google Scholar] [CrossRef]
- Ramos, S.; Rojas, L.; Lucena, M.; Meccia, G.; Alfredo, U. Chemical Composition and Antibacterial Activity of Origanum majorana L. Essential Oil from the Venezuelan Andes. J. Essent. Oil Res. J. Essent Oil Res 2011, 23, 45–49. [Google Scholar] [CrossRef]
- Vági, E.; Simándi, B.; Suhajda, Á.; Héthelyi, É. Essential oil composition and antimicrobial activity of Origanum majorana L. extracts obtained with ethyl alcohol and supercritical carbon dioxide. Food Res. Int. 2005, 38, 51–57. [Google Scholar] [CrossRef]
- Tabanca, N.; Özek, T.; Baser, K.H.C.; Tümen, G. Comparison of the Essential Oils of Origanum majorana L. and Origanum x majoricum Cambess. J. Essent. Oil Res. 2004, 16, 248–252. [Google Scholar] [CrossRef]
- Gulhati, P.; Cai, Q.; Li, J.; Liu, J.; Rychahou, P.G.; Qiu, S.; Lee, E.Y.; Silva, S.R.; Bowen, K.A.; Gao, T.; et al. Targeted inhibition of mammalian target of rapamycin signaling inhibits tumorigenesis of colorectal cancer. Clin. Cancer Res. 2009, 15, 7207–7216. [Google Scholar] [CrossRef] [Green Version]
- Dolado, I.; Nebreda, A. Regulation of Tumorigenesis by p38α MAP Kinase. In Stress Activated Protein Kinases Top Curr Genet; Springer: Berlin/Heidelberg, Germany, 2007; Volume 20, pp. 99–128. [Google Scholar]
- Dhar, R.; Persaud, S.D.; Mireles, J.R.; Basu, A. Proteolytic cleavage of p70 ribosomal S6 kinase by caspase-3 during DNA damage-induced apoptosis. Biochemistry 2009, 48, 1474–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.; Sun, P. The pathways to tumor suppression via route p38. Trends Biochem. Sci. 2007, 32, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Fathy, S.A.; Emam, M.; Agwa, S.; Zahra, F.A.; Youssef, F.; Sami, R.M. The antiproliferative effect of Origanum majorana on human hepatocarcinoma cell line: Suppression of NF-κB. Cell. Mol. Biol. (Noisy-Le-Grandfrance) 2016, 62, 80–84. [Google Scholar]
- Rezzoug, M.; Bakchiche, B.; Gherib, A.; Roberta, A.; FlaminiGuido; Kilinçarslan, Ö.; Mammadov, R.; Bardaweel, S.K. Chemical composition and bioactivity of essential oils and Ethanolic extracts of Ocimum basilicum L. and Thymus algeriensis Boiss. & Reut. from the Algerian Saharan Atlas. BMC Complement. Altern. Med. 2019, 19, 146. [Google Scholar] [PubMed]
- Kawase, K.; Mothé, C.; Furtado, F.; Coelho, G. Changes in essential oil of Origanum vulgare L. affected by different extraction methods. Int. J. Recent Res. Appl. Stud. 2013, 14, 238. [Google Scholar]
- Eisenberg-Lerner, A.; Bialik, S.; Simon, H.-U.; Kimchi, A. Life and death partners: Apoptosis, autophagy and the cross-talk between them. Cell Death Differ. 2009, 16, 966–975. [Google Scholar] [CrossRef]
- Xiao, X.; Wang, W.; Li, Y.; Yang, D.; Li, X.; Shen, C.; Liu, Y.; Ke, X.; Guo, S.; Guo, Z. HSP90AA1-mediated autophagy promotes drug resistance in osteosarcoma. J. Exp. Clin. Cancer Res. 2018, 37, 201. [Google Scholar] [CrossRef]
- Liu, M.; Zhao, G.; Zhang, D.; An, W.; Lai, H.; Li, X.; Cao, S.; Lin, X. Active fraction of clove induces apoptosis via PI3K/Akt/mTOR-mediated autophagy in human colorectal cancer HCT-116 cells. Int. J. Oncol. 2018, 53. [Google Scholar] [CrossRef]
- Ge, J.; Liu, Y.; Li, Q.; Guo, X.; Gu, L.; Ma, Z.G.; Zhu, Y.P. Resveratrol Induces Apoptosis and Autophagy in T-cell Acute Lymphoblastic Leukemia Cells by Inhibiting Akt/mTOR and Activating p38-MAPK. Biomed. Environ. Sci. 2013, 26, 902–911. [Google Scholar]
- Benbrook, D.; Long, A. Integration of autophagy, proteasomal degradation, unfolded protein response and apoptosis. Exp. Oncol. 2012, 34, 286–297. [Google Scholar]
- Rubinstein, A.D.; Kimchi, A. Life in the balance—A mechanistic view of the crosstalk between autophagy and apoptosis. J. Cell Sci. 2012, 125, 5259–5268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Q.; Li, F.; Shi, K.; Wu, P.; An, J.; Yang, Y.; Xu, C. Involvement of p38 in signal switching from autophagy to apoptosis via the PERK/eIF2??/ATF4 axis in selenite-treated NB4 cells. Cell Death Dis. 2014, 5, e1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.-Y.; Park, K.-I.; Kim, S.-H.; Yu, S.-N.; Park, S.-G.; Kim, Y.; Seo, Y.-K.; Ma, J.-Y.; Ahn, S.C. Inhibition of Autophagy Promotes Salinomycin-Induced Apoptosis via Reactive Oxygen Species-Mediated PI3K/AKT/mTOR and ERK/p38 MAPK-Dependent Signaling in Human Prostate Cancer Cells. Int. J. Mol. Sci. 2017, 18, 1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenton, T.R.; Gout, I.T. Functions and regulation of the 70kDa ribosomal S6 kinases. Int. J. Biochem. Cell Biol. 2011, 43, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Wang, J.; Yu, G.; Guo, T.; Hu, C.; Ren, P. Expression and clinical significance of mammalian target of rapamycin/P70 ribosomal protein S6 kinase signaling pathway in human colorectal carcinoma tissue. Oncol. Lett. 2015, 10, 277–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.D.; Zhang, W.J.; Zhang, M.Y.; Yuan, L.J.; Cha, Y.L.; Ying, X.F.; Wu, G.; Wang, H.Y. Overexpression of RPS6KB1 predicts worse prognosis in primary HCC patients. Med. Oncol. 2012, 29, 3070–3076. [Google Scholar] [CrossRef]
- Yamnik, R.L.; Digilova, A.; Davis, D.C.; Brodt, Z.N.; Murphy, C.J.; Holz, M.K. S6 Kinase 1 Regulates Estrogen Receptor α in Control of Breast Cancer Cell Proliferation. J. Biol. Chem. 2009, 284, 6361–6369. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, J.L.; Garcia, E.; Pieper, R.O. S6K1 plays a key role in glial transformation. Cancer Res. 2008, 68, 6516–6523. [Google Scholar] [CrossRef] [Green Version]
- Grasso, S.; Tristante, E.; Saceda, M.; Carbonell, P.; Mayor-López, L.; Carballo-Santana, M.; Carrasco-García, E.; Rocamora-Reverte, L.; García-Morales, P.; Carballo, F.; et al. Resistance to Selumetinib (AZD6244) in colorectal cancer cell lines is mediated by p70S6K and RPS6 activation. Neoplasia 2014, 16, 845–860. [Google Scholar] [CrossRef] [Green Version]
- Piedfer, M.; Bouchet, S.; Tang, R.; Billard, C.; Dauzonne, D.; Bauvois, B. p70S6 kinase is a target of the novel proteasome inhibitor 3,3′-diamino-4′-methoxyflavone during apoptosis in human myeloid tumor cells. Biochim. Et Biophys. Acta (Bba) Mol. Cell Res. 2013, 1833, 1316–1328. [Google Scholar] [CrossRef] [Green Version]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Athamneh, K.; Alneyadi, A.; Alsamri, H.; Alrashedi, A.; Palakott, A.; El-Tarabily, K.A.; Eid, A.H.; Al Dhaheri, Y.; Iratni, R. Origanum majorana Essential Oil Triggers p38 MAPK-Mediated Protective Autophagy, Apoptosis, and Caspase-Dependent Cleavage of P70S6K in Colorectal Cancer Cells. Biomolecules 2020, 10, 412. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10030412
Athamneh K, Alneyadi A, Alsamri H, Alrashedi A, Palakott A, El-Tarabily KA, Eid AH, Al Dhaheri Y, Iratni R. Origanum majorana Essential Oil Triggers p38 MAPK-Mediated Protective Autophagy, Apoptosis, and Caspase-Dependent Cleavage of P70S6K in Colorectal Cancer Cells. Biomolecules. 2020; 10(3):412. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10030412
Chicago/Turabian StyleAthamneh, Khawlah, Aysha Alneyadi, Halima Alsamri, Asma Alrashedi, Abdulrasheed Palakott, Khaled A. El-Tarabily, Ali H. Eid, Yusra Al Dhaheri, and Rabah Iratni. 2020. "Origanum majorana Essential Oil Triggers p38 MAPK-Mediated Protective Autophagy, Apoptosis, and Caspase-Dependent Cleavage of P70S6K in Colorectal Cancer Cells" Biomolecules 10, no. 3: 412. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10030412