Oxidized LDL Modify the Human Adipocyte Phenotype to an Insulin Resistant, Proinflamatory and Proapoptotic Profile

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Laboratory Measurements
2.3. Isolation of Stromal Vascular Fraction and Mature Adipocytes
2.4. In Vitro Differentiated Adipocyte Culture
2.5. Analysis of the Culture Medium
2.6. RNA Extraction and Real-Time Quantitative PCR
2.7. Immunohistochemical Staining
2.8. Oil Red-O Staining
2.9. Cellular Cholesterol Quantification
2.10. Glucose Uptake
2.11. Statistical Analysis
3. Results
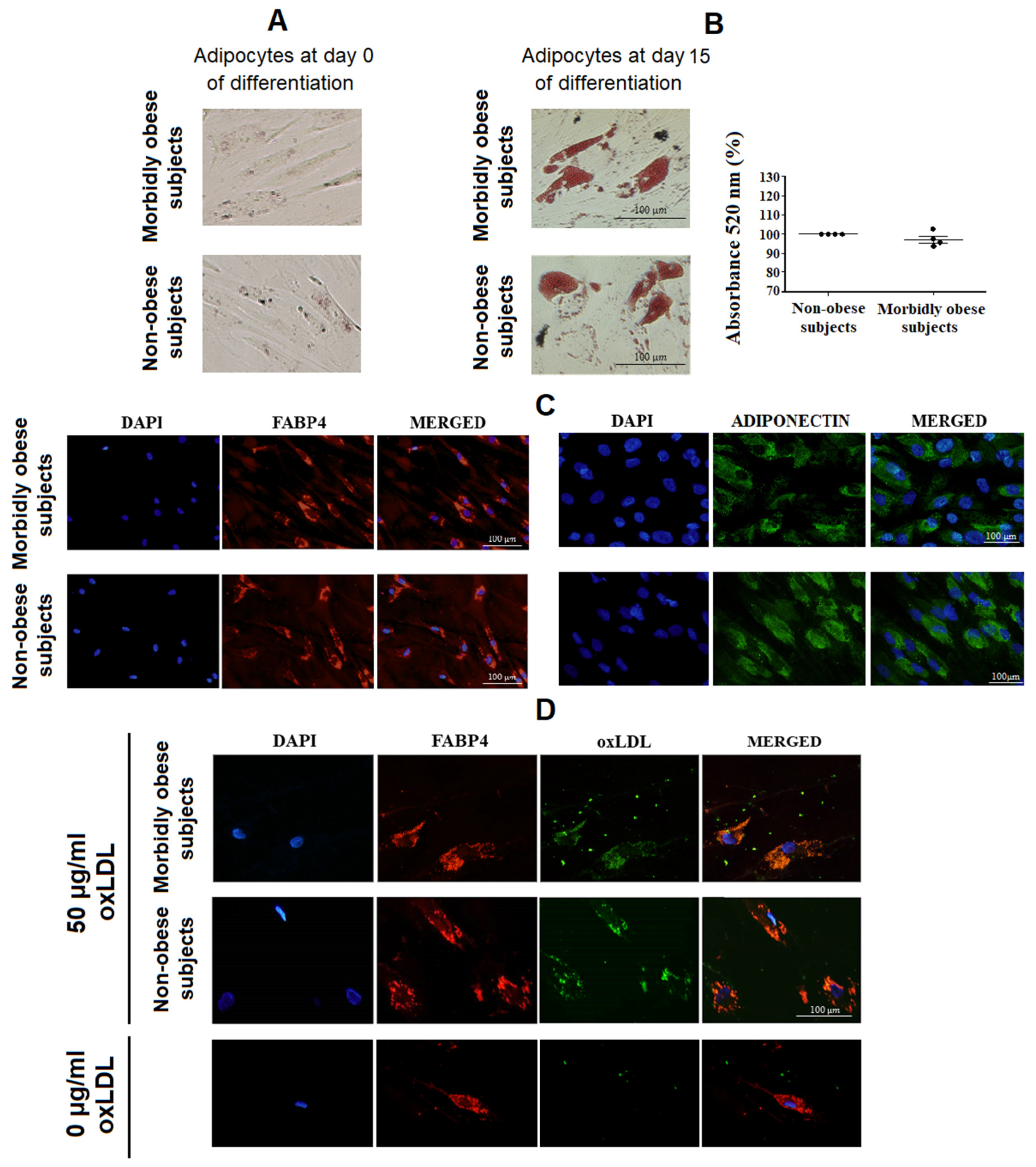
3.1. OxLDL Uptake by Visceral In Vitro Differentiated Adipocytes
3.2. Esterified Cholesterol within Visceral In Vitro Differentiated Adipocytes Increases with OxLDL
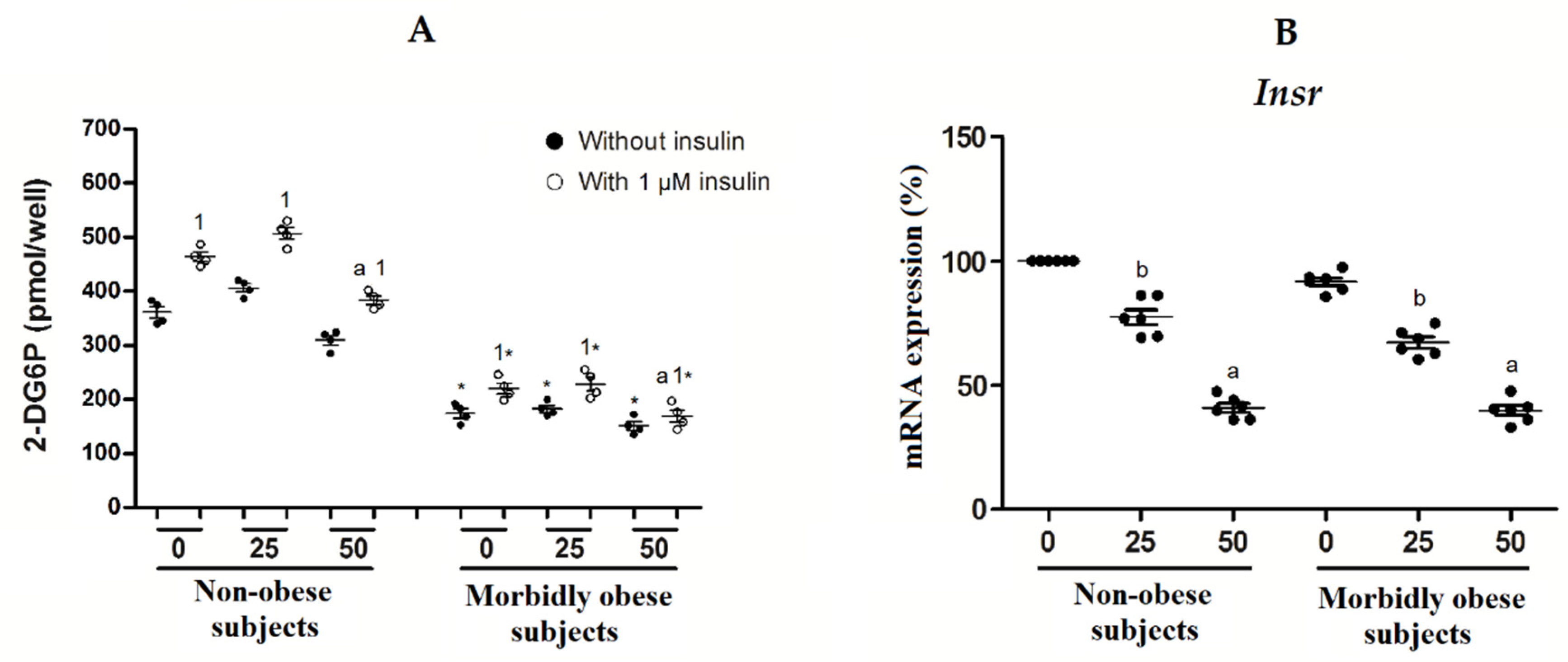
3.3. OxLDL Produces an Increase in Insulin Resistance of Visceral In Vitro Differentiated Adipocytes
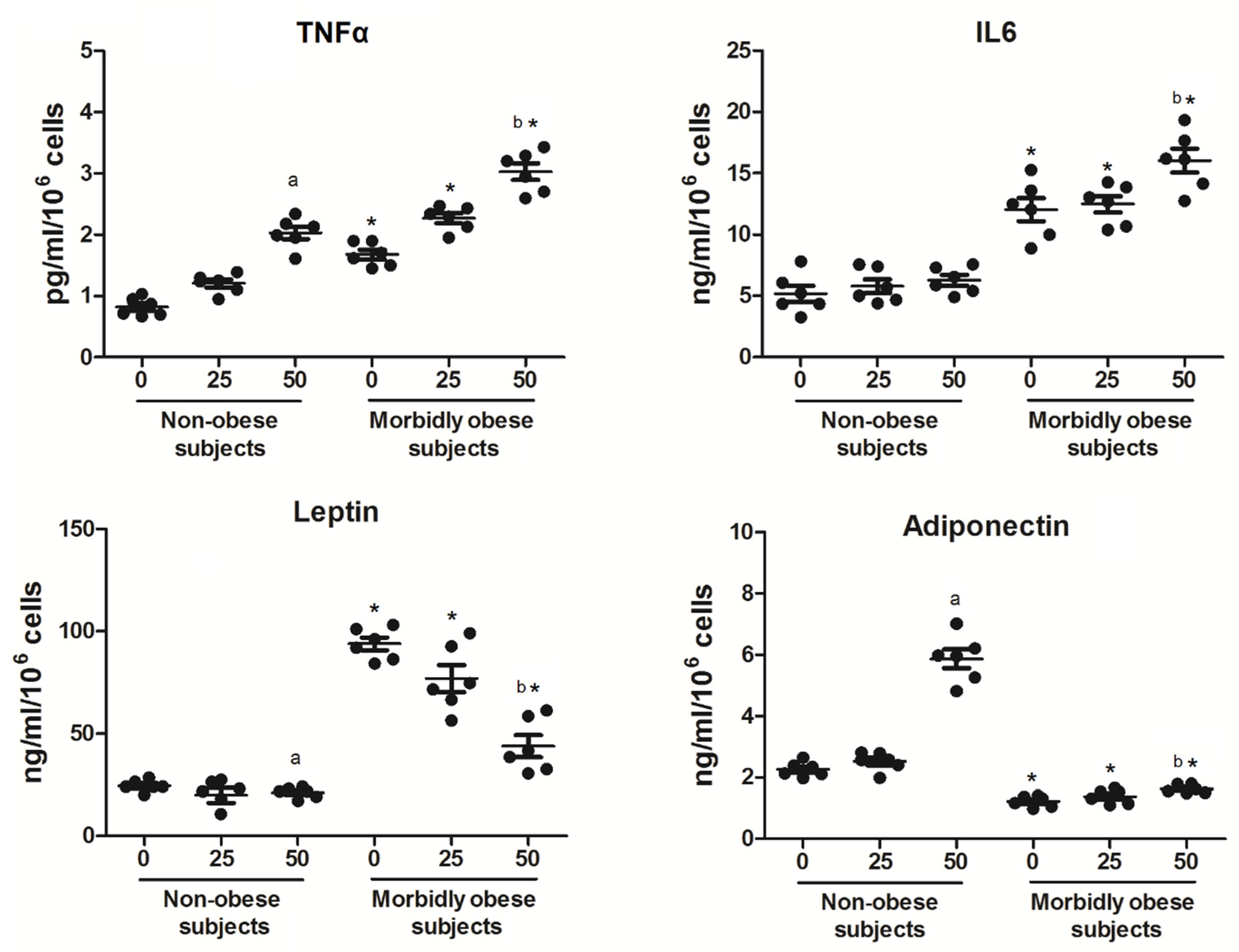
3.4. OxLDL Modifies the Secretion of Adipocytokines
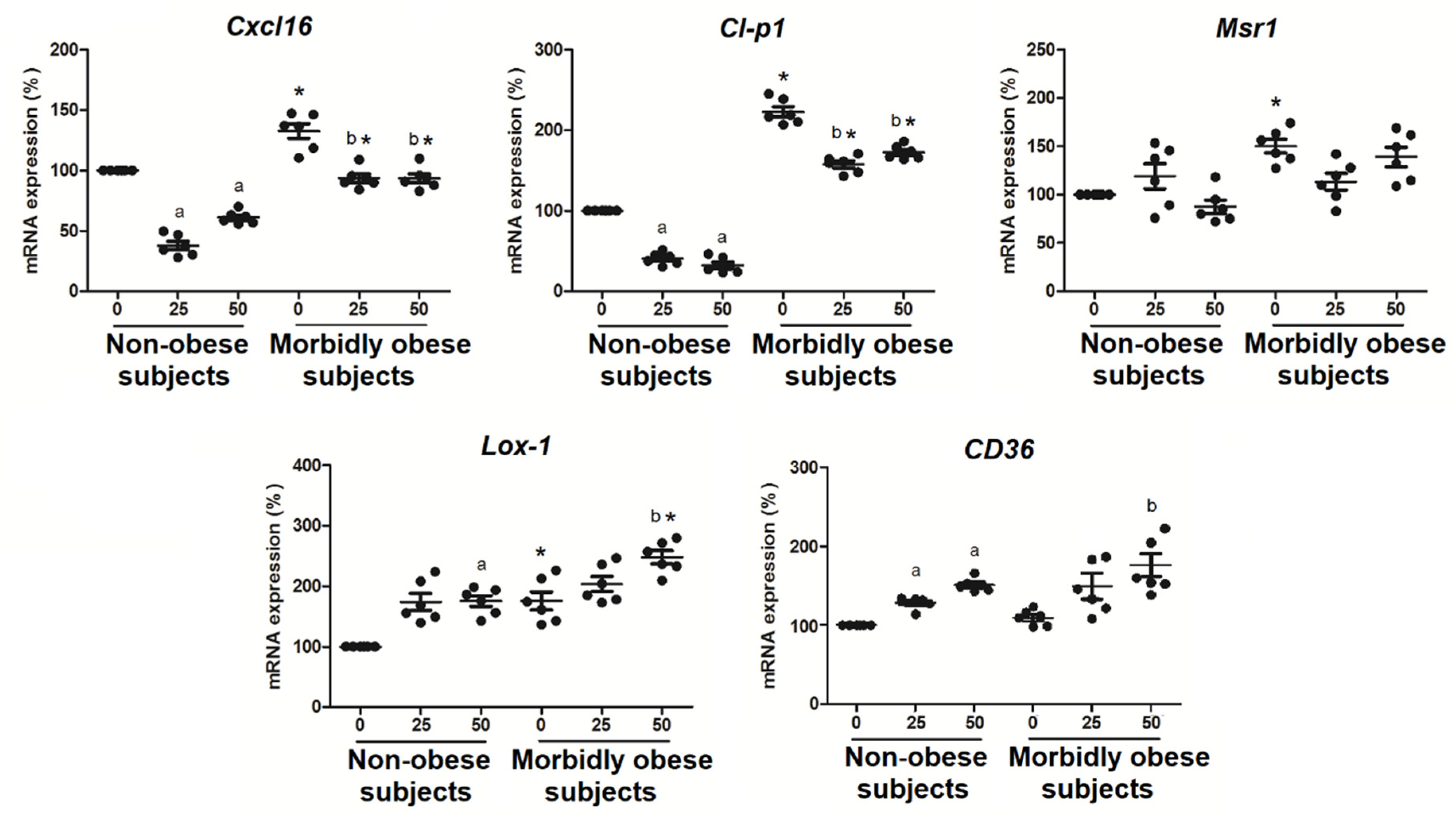
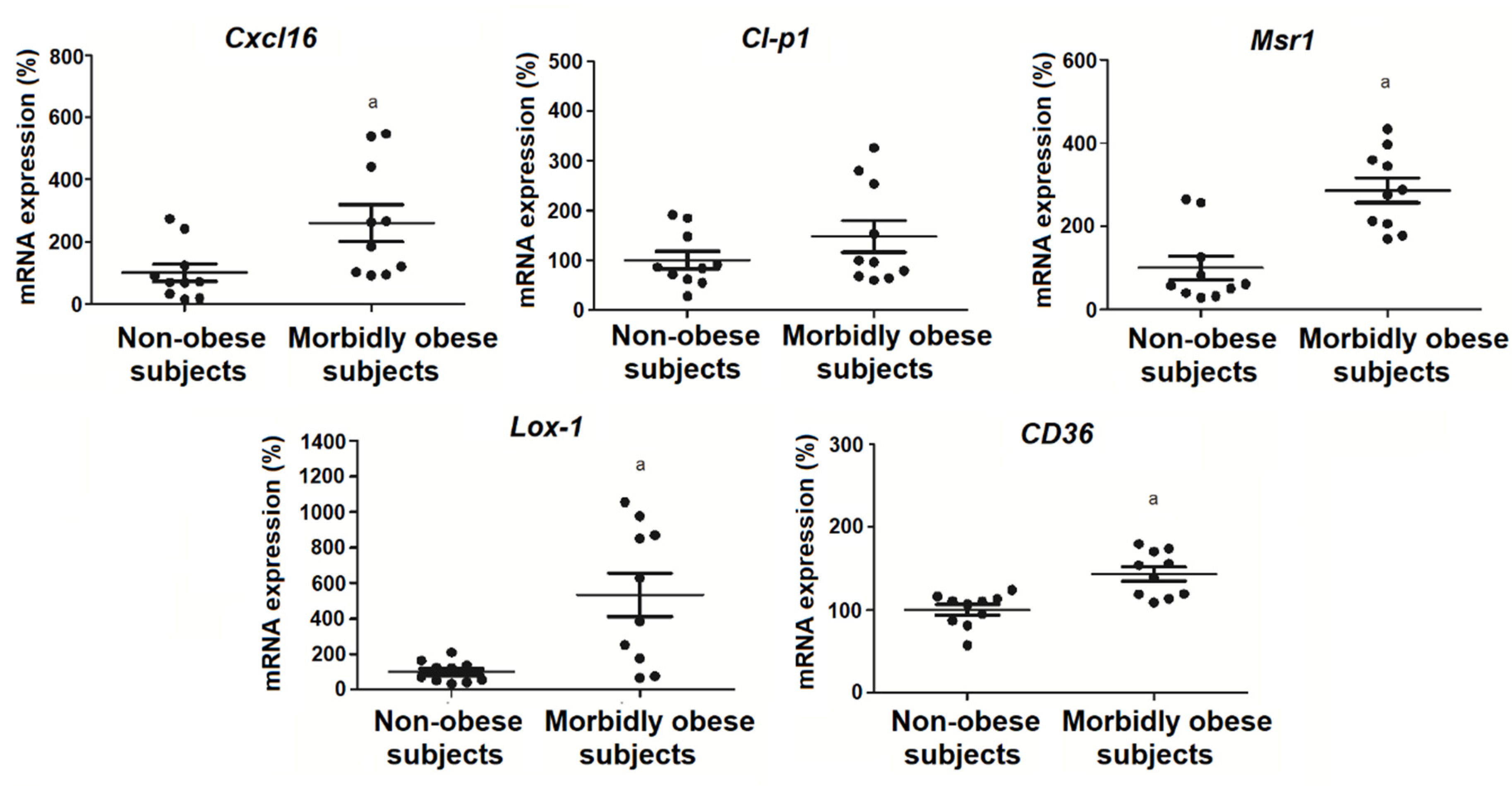
3.5. OxLDL Increases the Expression of SRs in Visceral In Vitro Differentiated Adipocytes
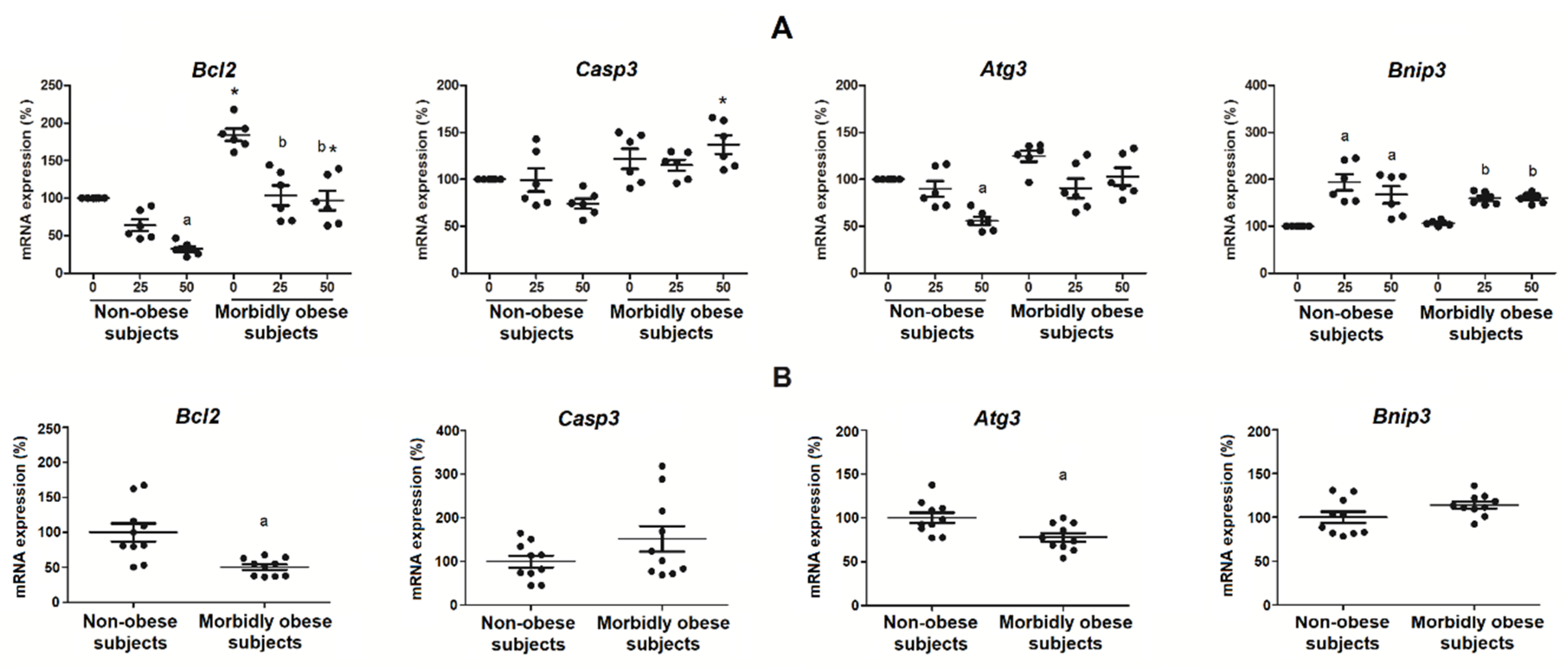
3.6. Oxldl Modifies the Expression of Apoptosis, Necrosis, and Autophagy Markers in Visceral In Vitro Differentiated Adipocytes
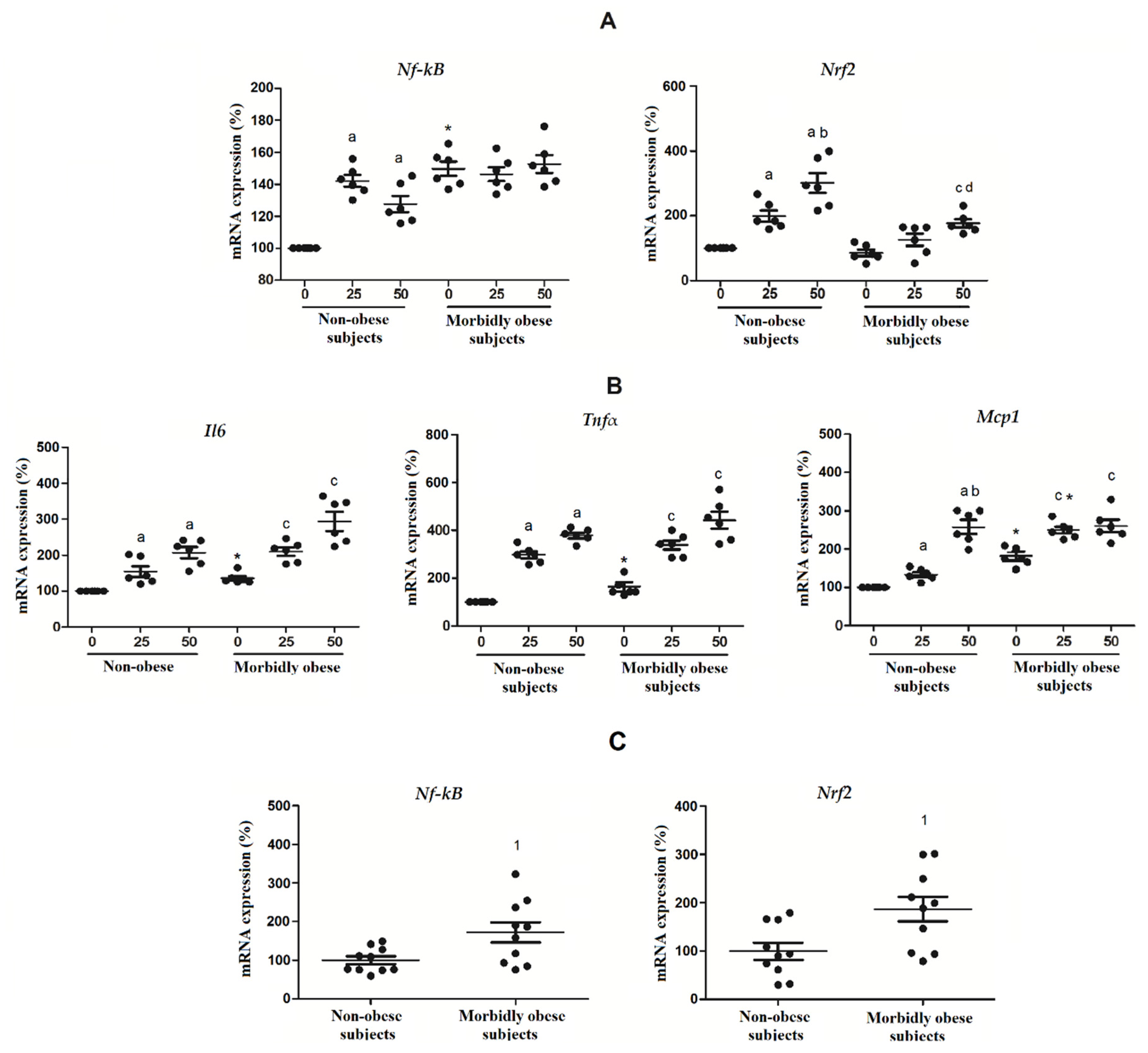
3.7. OxLDL Modifies the Expression of Nrf2 and NF-kB in Visceral In Vitro Differentiated Adipocytes
4. Discussion
4.1. Effects of oxLDL on Intracellular Cholesterol and Insulin Resistance
4.2. Effects of OxLDL on Adipocytokine Secretion
4.3. Effects of OxLDL on Scavenger Receptors
4.4. Effects of OxLDL on Apoptosis, Necrosis, and Autophagy Markers
4.5. Effects of OxLDL on Transcription Factors
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, J. Adipose tissue macrophages in the development of obesity-induced inflammation, insulin resistance and type 2 Diabetes. Arch. Pharm. Res. 2013, 36, 208–222. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Chen, M.; Wu, Z.; Zhao, S. Ox-LDL Induces ER Stress and Promotes the adipokines Secretion in 3T3-L1 Adipocytes. PLoS ONE 2013, 8, e81379. [Google Scholar] [CrossRef] [Green Version]
- Weinbrenner, T.; Schröder, H.; Escurriol, V.; Fito, M.; Elosua, R.; Vila, J.; Marrugat, J.; Covas, M.I. Circulating oxidized LDL is associated with increased waist circumference independent of body mass index in men and women. Am. J. Clin. Nutr. 2006, 83, 30–35. [Google Scholar] [CrossRef] [Green Version]
- Holvoet, P.; Lee, D.H.; Steffes, M.; Gross, M.; Jacobs, D.R. Association between circulating oxidized low-density lipoprotein and incidence of the metabolic syndrome. JAMA 2008, 299, 2287–2293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holvoet, P.; Jenny, N.S.; Schreiner, P.J.; Tracy, R.P.; Jacobs, D.R. The relationship between oxidized LDL and other cardiovascular risk factors and subclinical CVD in different ethnic groups: The Multi-Ethnic Study of Atherosclerosis (MESA). Atherosclerosis 2007, 194, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Ishigaki, Y.; Oka, Y.; Katagiri, H. Circulating oxidized LDL: A biomarker and a pathogenic factor. Curr. Opin. Lipidol. 2009, 20, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Miller, Y.I.; Chang, M.K.; Binder, C.J.; Shaw, P.X.; Witztum, J.L. Oxidized low density lipoprotein and innate immune receptors. Curr. Opin. Lipidol. 2003, 14, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Kunjathoor, V.V.; Febbraio, M.; Podrez, E.A.; Moore, K.J.; Andersson, L.; Koehn, S.; Rhee, J.S.; Silverstein, R.; Hoff, H.F.; Freeman, M.W. Scavenger receptors class A-I/II and CD36 are the principal receptors responsible for the uptake of modified low density lipoprotein leading to lipid loading in macrophages. J. Biol. Chem. 2002, 277, 49982–49988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Archivio, M.; Scazzocchio, B.; Filesi, C.; Varì, R.; Maggiorella, M.T.; Sernicola, L.; Santangelo, C.; Giovannini, C.; Masella, R. Oxidised LDL up-regulate CD36 expression by the Nrf2 pathway in 3T3-L1 preadipocytes. FEBS Lett. 2008, 582, 2291–2298. [Google Scholar] [CrossRef] [Green Version]
- Cuffe, H.; Liu, M.; Key, C.C.; Boudyguina, E.; Sawyer, J.K.; Weckerle, A.; Bashore, A.; Fried, S.K.; Chung, S.; Parks, J.S. Targeted Deletion of Adipocyte Abca1 (ATP-Binding Cassette Transporter A1) Impairs Diet-Induced Obesity. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 733–743. [Google Scholar] [CrossRef] [Green Version]
- Salvayre, R.; Auge, N.; Benoist, H.; Negre-Salvayre, A. Oxidized low-density lipoprotein-induced apoptosis. Biochim. Biophys. Acta 2002, 1585, 213–221. [Google Scholar] [CrossRef]
- Chisolm, G.M.; Chai, Y.C. Regulation of cell growth by oxidized LDL. Free Radic. Biol. Med. 2000, 28, 1697–1707. [Google Scholar] [PubMed]
- Wintergerst, E.S.; Jelk, J.; Rahner, C.; Asmis, R. Apoptosis induced by oxidized low density lipoprotein in human monocyte-derived macrophages involves CD36 and activation of caspase-3. Eur. J. Biochem. 2000, 267, 6050–6059. [Google Scholar] [CrossRef] [PubMed]
- Kuniyasu, A.; Hayashi, S.; Nakayama, H. Adipocytes recognize and degrade oxidized low density lipoprotein through CD36. Biochem. Biophys. Res. Commun. 2002, 295, 319–323. [Google Scholar] [CrossRef]
- Garrido-Sánchez, L.; Vendrell, J.; Fernández-García, D.; Ceperuelo-Mallafré, V.; Chacón, M.R.; Ocaña-Wilhelmi, L.; Alcaide, J.; Tinahones, F.J.; García-Fuentes, E. De Novo Lipogenesis in Adipose Tissue Is Associated with Course of Morbid Obesity after Bariatric Surgery. PLoS ONE 2012, 7, e31280. [Google Scholar] [CrossRef] [Green Version]
- Soriguer, F.; Rojo-Martínez, G.; Almaraz, M.C.; Esteva, I.; Ruiz de Adana, M.S.; Morcillo, S.; Valdés, S.; García-Fuentes, E.; García-Escobar, E.; Cardona, I.; et al. Incidence of type 2 diabetes in southern Spain (Pizarra Study). Eur. J. Clin. Invest. 2008, 38, 126–133. [Google Scholar] [CrossRef]
- Rodriguez-Pacheco, F.; Gutierrez-Repiso, C.; Garcia-Serrano, S.; Alaminos-Castillo, M.A.; Ho-Plagaro, A.; Valdes, S.; Garcia-Arnes, J.; Gonzalo, M.; Andrade, R.J.; Moreno-Ruiz, F.J.; et al. The pro-/anti-inflammatory effects of different fatty acids on visceral adipocytes are partially mediated by GPR120. Eur. J. Nutr. 2016, 56, 1743–1752. [Google Scholar] [CrossRef]
- Pachón-Peña, G.; Serena, C.; Ejarque, M.; Petriz, J.; Duran, X.; Oliva-Olivera, W.; Simó, R.; Tinahones, F.J.; Fernández-Veledo, S.; Vendrell, J. Obesity Determines the Immunophenotypic Profile and Functional Characteristics of Human Mesenchymal Stem Cells From Adipose Tissue. Stem Cells Transl. Med. 2016, 5, 464–475. [Google Scholar] [CrossRef]
- Shan, T.; Liu, W.; Kuang, S. Fatty acid binding protein 4 expression marks a population of adipocyte progenitors in white and brown adipose tissues. FASEB J. 2013, 27, 277–287. [Google Scholar] [CrossRef] [Green Version]
- Masella, R.; Varì, R.; D’Archivio, M.; Santangelo, C.; Scazzocchio, B.; Maggiorella, M.T.; Sernicola, L.; Titti, F.; Sanchez, M.; Di Mario, U.; et al. Oxidised LDL modulate adipogenesis in 3T3-L1 preadipocytes by affecting the balance between cell proliferation and differentiation. FEBS Lett. 2006, 580, 2421–2429. [Google Scholar] [CrossRef] [Green Version]
- Lin, F.Y.; Tsao, N.W.; Shih, C.M.; Lin, Y.W.; Yeh, J.S.; Chen, J.W.; Nakagami, H.; Morishita, R.; Sawamura, T.; Huang, C.Y. The biphasic effects of oxidized-low density lipoprotein on the vasculogenic function of endothelial progenitor cells. PLoS ONE 2015, 10, e0123971. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Pacheco, F.; Gutierrez-Repiso, C.; García-Serrano, S.; Ho-Plagaro, A.; Gómez-Zumaquero, J.M.; Valdes, S.; Gonzalo, M.; Rivas-Becerra, J.; Montiel-Casado, C.; Rojo-Martínez, G.; et al. Growth hormone-releasing hormone is produced by adipocytes and regulates lipolysis through growth hormone receptor. Int. J. Obes. 2017, 41, 1547–1555. [Google Scholar] [CrossRef] [PubMed]
- Castillo, M.; Hortal, J.H.; García-Fuentes, E.; Zafra, M.F.; García-Peregrín, E. Coconut oil affects lipoprotein composition and structure of neonatal chicks. J. Biochem. 1996, 119, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Kelly, A.; Jacobs, D.J.; Sinaiko, A.; Moran, A.; Steffen, L.; Steinberger, J. Relation of circulating oxidized LDL to obesity and insulin resistance in children. Pediatr. Diabetes 2010, 11, 552–555. [Google Scholar] [CrossRef] [Green Version]
- Demirel, F.; Bideci, A.; Cinaz, P.; Yesilkaya, E.; Hasano, A. Serum leptin, oxidized low density lipoprotein and plasma asymmetric dimethylarginine levels and their relationship with dyslipidaemia in adolescent girls with polycystic ovary syndrome. Clin. Endocrinol. 2007, 67, 129–134. [Google Scholar] [CrossRef]
- Sjogren, P.; Basu, S.; Rosell, M.; Silveira, A.; De Faire, U.; Vessby, B.; Hamsten, A.; Hellenius, M.L.; Fisher, R.M. Measures of oxidized low-density lipoprotein and oxidative stress are not related and not elevated in otherwise healthy men with the metabolic syndrome. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 2580–2586. [Google Scholar] [CrossRef] [Green Version]
- Garrido-Sánchez, L.; García-Almeida, J.M.; García-Serrano, S.; Cardona, I.; García-Arnes, J.; Soriguer, F.; Tinahones, F.J.; García-Fuentes, E. Improved carbohydrate metabolism after bariatric surgery raises antioxidized LDL antibody levels in morbidly obese patients. Diabetes Care 2008, 31, 2258–2264. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.P.; Wu, J.; Zhang, D.Q.; Ye, H.J.; Liu, L.; Li, J.Q. Fenofibrate enhances CD36 mediated endocytic uptake and degradation of oxidized low density lipoprotein in adipocytes from hypercholesterolemia rabbit. Atherosclerosis 2004, 177, 255–262. [Google Scholar] [CrossRef]
- Yu, B.L.; Zhao, S.P.; Hu, J.R. Cholesterol imbalance in adipocytes: A posible mechanism of adipocytes dysfunction in obesity. Obes. Rev. 2010, 11, 560–567. [Google Scholar] [CrossRef]
- Voet, D.; Voet, J.G. Biochemistry, 4th ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2011; p. 449. [Google Scholar]
- Zhang, Y.; McGillicuddy, F.C.; Hinkle, C.C.; O’Neill, S.; Glick, J.M.; Rothblat, G.H.; Reilly, M.P. Adipocyte modulation of high-density lipoprotein cholesterol. Circulation 2010, 121, 1347–1355. [Google Scholar] [CrossRef] [Green Version]
- Rosenson, R.S.; Brewer, H.B., Jr.; Davidson, W.S.; Fayad, Z.A.; Fuster, V.; Goldstein, J.; Hellerstein, M.; Jiang, X.C.; Phillips, M.C.; Rader, D.J.; et al. Cholesterol efflux and atheroprotection: Advancing the concept of reverse cholesterol transport. Circulation 2012, 125, 1905–1919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scazzocchio, B.; Varì, R.; D’Archivio, M.; Santangelo, C.; Filesi, C.; Giovannini, C.; Masella, R. Oxidized LDL impair adipocyte response to insulin by activating serine/threonine kinases. J. Lipid Res. 2009, 50, 832–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagishita, Y.; Uruno, A.; Fukutomi, T.; Saito, R.; Saigusa, D.; Pi, J.; Fukamizu, A.; Sugiyama, F.; Takahashi, S.; Yamamoto, M. Nrf2 Improves Leptin and Insulin Resistance Provoked by Hypothalamic Oxidative Stress. Cell Rep. 2017, 18, 2030–2044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Kulkarni, S.R.; Donepudi, A.C.; More, V.R.; Slitt, A.L. Enhanced Nrf2 activity worsens insulin resistance, impairs lipid accumulation in adipose tissue, and increases hepatic steatosis in leptin-deficient mice. Diabetes 2012, 61, 3208–3218. [Google Scholar] [CrossRef] [Green Version]
- Chartoumpekis, D.V.; Palliyaguru, D.L.; Wakabayashi, N.; Fazzari, M.; Khoo, N.K.H.; Schopfer, F.J.; Sipula, I.; Yagishita, Y.; Michalopoulos, G.K.; O’Doherty, R.M.; et al. Nrf2 deletion from adipocytes, but not hepatocytes, potentiates systemic metabolic dysfunction after long-term high-fat diet-induced obesity in mice. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E180–E195. [Google Scholar] [CrossRef] [Green Version]
- Unno, Y.; Sakai, M.; Sakamoto, Y.I.; Kuniyasu, A.; Nakayama, H.; Nagai, R.; Horiuchi, S. Advanced glycation end products-modified proteins and oxidized LDL mediate down-regulation of leptin in mouse adipocytes via CD36. Biochem. Biophys. Res. Commun. 2004, 325, 151–156. [Google Scholar] [CrossRef]
- Mazière, C.; Mazière, J.C. Activation of transcription factors and gene expression by oxidized low-density lipoprotein. Free Radic. Biol. Med. 2009, 46, 127–137. [Google Scholar] [CrossRef]
- Liu, M.; Liu, F. Transcriptional and post-translational regulation of adiponectin. Biochem. J. 2009, 425, 41–52. [Google Scholar] [CrossRef]
- Francisco, V.; Pino, J.; Campos-Cabaleiro, V.; Ruiz-Fernández, C.; Mera, A.; Gonzalez-Gay, M.A.; Gómez, R.; Gualillo, O. Obesity, fat mass and immune system: Role for leptin. Front. Physiol. 2018, 9, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Ghigliotti, G.; Barisione, C.; Garibaldi, S.; Fabbi, P.; Brunelli, C.; Spallarossa, P.; Altieri, P.; Rosa, G.; Spinella, G.; Palombo, D.; et al. Adipose tissue immune response: Novel triggers and consequences for chronic inflammatory conditions. Inflammation 2014, 37, 1337–1353. [Google Scholar] [CrossRef] [Green Version]
- Kaur, S.; Zilmer, K.; Leping, V.; Zilmer, M. The levels of adiponectin and leptin and their relation to other markers of cardiovascular risk in patients with psoriasis. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 1328–1333. [Google Scholar] [CrossRef]
- Porreca, E.; Di Febbo, C.; Moretta, V.; Angelini, A.; Guglielmi, M.D.; Di Nisio, M.; Cuccurullo, F. Circulating leptin is associated with oxidized LDL in postmenopausal women. Atherosclerosis 2004, 175, 139–143. [Google Scholar] [CrossRef]
- Jovinge, S.; Ares, M.P.; Kallin, B.N.J. Human monocytes/macrophages release TNF-alpha in response to Ox-LDL. Arter. Thromb. Vasc. Biol. 1996, 16, 1573–1579. [Google Scholar] [CrossRef]
- Lappas, M.; Yee, K.; Permezel, M.; Rice, G.E. Sulfasalazine and BAY 11-7082 interfere with the nuclear factor-kappa B and I kappa B kinase pathway to regulate the release of proinflammatory cytokines from human adipose tissue and skeletal muscle in vitro. Endocrinology 2005, 146, 1491–1497. [Google Scholar] [CrossRef]
- Hamilton, T.A.; Ma, G.P.; Chisolm, G.M. Oxidized low density lipoprotein suppresses the expression of tumor necrosis factor-alpha mRNA in stimulated murine peritoneal macrophages. J. Immunol. 1990, 144, 2343–2350. [Google Scholar] [PubMed]
- Neuenschwander, P.F.; Morrissey, J.H. Deletion of the membrane anchoring region of tissue factor abolishes autoactivation of factor VII but not cofactor function: Analysis of a mutant with a selective deficiency in activity. J. Biol. Chem. 1992, 267, 14477–14482. [Google Scholar] [PubMed]
- Santiago-Fernandez, C.; Gutierrez-Repiso, C.; Garcia-Almeida, J.M.; Alcaide, J.; Ocaña-Wilhelmi, L.; Rivas-Becerra, J.; Rodriguez-Morata, A.; Tome, M.; Oliva-Olivera, W.; Tatzber, F.; et al. Effect of hypoxia on scavenger receptors and inflammation in adipocytes. Atherosclerosis 2017, 263, e251–e252. [Google Scholar] [CrossRef]
- Mehta, J.L.; Li, D.Y. Identification and autoregulation of receptor for ox-LDL in cultured human coronary artery endothelial cells. Biochem. Biophys. Res. Commun. 1998, 248, 511–514. [Google Scholar] [CrossRef] [PubMed]
- Khaidakov, M.; Mitra, S.; Kang, B.Y.; Wang, X.; Kadlubar, S.; Novelli, G.; Raj, V.; Winters, M.; Carter, W.C.; Mehta, J.L. Oxidized LDL receptor 1 (OLR1) as a possible link between obesity, dyslipidemia and cancer. PLoS ONE 2011, 6, e20277. [Google Scholar] [CrossRef]
- Izquierdo, M.C.; Martin-Cleary, C.; Fernandez-Fernandez, B.; Elewa, U.; Sanchez-Niño, M.D.; Carrero, J.J.; Ortiz, A. CXCL16 in kidney and cardiovascular injury. Cytokine Growth Factor Rev. 2014, 25, 317–325. [Google Scholar] [CrossRef]
- Bordicchia, M.; Spannella, F.; Ferretti, G.; Bacchetti, T.; Vignini, A.; Di Pentima, C.; Mazzanti, L.; Sarzani, R. PCSK9 is Expressed in Human Visceral Adipose Tissue and Regulated by Insulin and Cardiac Natriuretic Peptides. Int. J. Mol. Sci. 2019, 20, 245. [Google Scholar] [CrossRef] [Green Version]
- Demers, A.; Samami, S.; Lauzier, B.; Des Rosiers, C.; Ngo Sock, E.T.; Ong, H.; Mayer, G. PCSK9 Induces CD36 Degradation and Affects Long-Chain Fatty Acid Uptake and Triglyceride Metabolism in Adipocytes and in Mouse Liver. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 2517–2525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kataoka, H.; Kume, N.; Miyamoto, S.; Minami, M.; Morimoto, M.; Hayashida, K.; Hashimoto, N.; Kita, T. Oxidized LDL Oxidized LDL modulates Bax/Bcl-2 through the lectinlike Ox-LDL receptor-1 in vascular smooth muscle cells. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 955–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Mehta, J.L.; Haider, N.; Zhang, X.; Narula, J.; Li, D. Role of Caspases in Ox-LDL-Induced Apoptotic Cascade in Human Coronary Artery Endothelial Cells. Circ. Res. 2004, 94, 370–376. [Google Scholar]
- Xu, X.; Shao, N.; Duan, X.; Zhang, X.; Zhang, Y. Tacrolimus alleviates Ox-LDL damage through inducing vascular endothelial autophagy. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3199–3206. [Google Scholar] [PubMed]
- Mollace, V.; Gliozzi, M.; Musolino, V.; Carresi, C.; Muscoli, S.; Mollace, R.; Tavernese, A.; Gratteri, S.; Palma, E.; Morabito, C.; et al. Oxidized LDL attenuates protective autophagy and induces apoptotic cell death of endothelial cells: Role of oxidative stress and LOX-1 receptor expression. Int. J. Cardiol. 2015, 184, 152–158. [Google Scholar] [CrossRef]
- Tinahones, F.J.; Coín Aragüez, L.; Murri, M.; Oliva Olivera, W.; Mayas Torres, M.D.; Barbarroja, N.; Gomez Huelgas, R.; Malagón, M.M.; El Bekay, R. Caspase induction and BCL2 inhibition in human adipose tissue: A potential relationship with insulin alteration. Diabetes Care 2013, 36, 513–521. [Google Scholar]
- Hsieh, C.C.; Yen, M.H.; Yen, C.H.; Lau, Y.T. Oxidized low density lipoprotein induces apoptosis via generation of reactive oxygen species in vascular smooth muscle cells. Cardiovasc. Res. 2001, 49, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Ray, R.; Chen, G.; Velde, C.V.; Cizeau, J.; Park, J.H.; Reed, J.C.; Daniel Gietz, R.; Greenberg, A.H. BNIP3 heterodimerizes with Bcl-2/Bcl-X(L) and induces cell death independent of a Bcl-2 homology 3 (BH3) domain at both mitochondrial and nonmitochondrial sites. J. Biol. Chem. 2000, 275, 1439–1448. [Google Scholar] [CrossRef] [Green Version]
- Hamacher-Brady, A.; Brady, N.R.; Logue, S.E.; Sayen, M.R.; Jinno, M.; Kirshenbaum, L.A.; Gottlieb, R.A.; Gustafsson, Å.B. Response to myocardial ischemia/reperfusion injury involves Bnip3 and autophagy. Cell Death Differ. 2007, 14, 146–157. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Massen, S.; Terenzio, M.; Lang, V.; Chen-Lindner, S.; Eils, R.; Novak, I.; Dikic, I.; Hamacher-Brady, A.; Brady, N.R. Modulation of serines 17 and 24 in the LC3-interacting region of Bnip3 determines pro-survival mitophagy versus apoptosis. J. Biol. Chem. 2013, 288, 1099–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wardyn, J.D.; Ponsford, A.H.; Sanderson, C.M. Dissecting molecular cross-talk between Nrf2 and NF-κB response pathways. Biochem. Soc. Trans. 2015, 43, 621–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brasier, A.R. The nuclear factor-kappaB-interleukin-6 signalling pathway mediating vascular inflammation. Cardiovasc. Res. 2010, 86, 211–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, H.; Tao, T.; Wang, X.; Liu, M.; Song, D.; Liu, X.; Shi, D. Zedoarondiol Attenuates Endothelial Cells Injury Induced by Oxidized Low-Density Lipoprotein via Nrf2 Activation. Cell Physiol. Biochem. 2018, 48, 1468–1479. [Google Scholar] [CrossRef]
- Ishii, T.; Itoh, K.; Ruiz, E.; Leake, D.S.; Unoki, H.; Yamamoto, M.; Mann, G.E. Role of Nrf2 in the regulation of CD36 and stress protein expression in murine macrophages. Circ. Res. 2004, 94, 609–616. [Google Scholar] [CrossRef] [Green Version]
- Ruotsalainen, A.K.; Inkala, M.; Partanen, M.E.; Lappalainen, J.P.; Kansanen, E.; Mäkinen, P.I.; Heinonen, S.E.; Laitinen, H.M.; Heikkilä, J.; Vatanen, T.; et al. The absence of macrophage Nrf2 promotes early atherogenesis. Cardiovasc. Res. 2013, 98, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Murri, M.; García-Fuentes, E.; García-Almeida, J.M.; Garrido-Sánchez, L.; Mayas, M.D.; Bernal, R.; Tinahones, F.J. Changes in oxidative stress and insulin resistance in morbidly obese patients after bariatric surgery. Obes. Surg. 2010, 20, 363–368. [Google Scholar] [CrossRef]
- Huang, Y.; Li, W.; Su, Z.Y.; Kong, A.N. The complexity of the Nrf2 pathway: Beyond the antioxidant response. J. Nutr. Biochem. 2015, 26, 1401–1413. [Google Scholar] [CrossRef]
- Knerr, I.; Herzog, D.; Rauh, M.; Rascher, W.; Horbach, T. Leptin and ghrelin expression in adipose tissues and serum levels in gastric banding patients. Eur. J. Clin. Invest. 2006, 36, 389–394. [Google Scholar] [CrossRef]
- Barbarroja, N.; López-Pedrera, R.; Mayas, M.D.; García-Fuentes, E.; Garrido-Sánchez, L.; Macías-González, M.; El Bekay, R.; Vidal-Puig, A.; Tinahones, F.J. The obese healthy paradox: Is inflammation the answer? Biochem. J. 2010, 430, 141–149. [Google Scholar] [CrossRef] [Green Version]
- Hammes, T.O.; Costa Cdos, S.; Rohden, F.; Margis, R.; de Almeida, J.C.; Padoin, A.V.; Mottin, C.C.; Guaragna, R.M. Parallel down-regulation of FOXO1, PPARγ and adiponectin mRNA expression in visceral adipose tissue of class III obese individuals. Obes. Facts 2012, 5, 452–459. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Non-Obese Subjects (n = 10) | Morbidly Obese Subjects (n = 10) |
|---|---|---|
| Sex (male/female) | 4/6 | 4/6 |
| Age (years) | 42.18 ± 15.1 | 40.79 ± 9.02 |
| Weight (Kg) | 74.3 ± 12.5 | 131.8 ± 20.1 ¶ |
| BMI (kg/m2) | 27.0 ± 3.4 | 49.3 ± 7.8 ¶ |
| Waist (cm) | 98.2 ± 11.3 | 135.5 ± 15.7 ¶ |
| Hip (cm) | 101.0 ± 9.1 | 149.1 ± 15.3 ¶ |
| Waist/hip ratio | 0.97 ± 0.12 | 0.91 ± 0.10 |
| SBP | 127.4 ± 19.7 | 149.1 ± 15.3 ∗ |
| DBP | 80.6 ± 10.2 | 82.7 ± 12.5 |
| Glucose (mg/dL) | 96.7 ± 17.5 | 91.3 ± 16.1 |
| Cholesterol (mg/dL) | 201.1 ± 35.3 | 185.0 ± 37.7 |
| Triglycerides (mg/dL) | 135.3 ± 61.1 | 116.1 ± 51.7 |
| HDL (mg/dL) | 48.5 ± 8.5 | 48.8 ± 11.2 |
| LDL (mg/dL) | 126.8 ± 32.4 | 113.6 ± 31.9 |
| Insulin (μIU/mL) | 9.2 ± 6.3 | 13.6 ± 4.2 |
| HOMA-IR | 2.7 ± 1.6 | 4.4 ± 3.5 |
| Oxidized-LDL (mU/L) | 60562.5 ± 10687.7 | 70658.1 ± 13975.4 ∗ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santiago-Fernández, C.; Martin-Reyes, F.; Tome, M.; Ocaña-Wilhelmi, L.; Rivas-Becerra, J.; Tatzber, F.; Pursch, E.; Tinahones, F.J.; García-Fuentes, E.; Garrido-Sánchez, L. Oxidized LDL Modify the Human Adipocyte Phenotype to an Insulin Resistant, Proinflamatory and Proapoptotic Profile. Biomolecules 2020, 10, 534. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10040534
Santiago-Fernández C, Martin-Reyes F, Tome M, Ocaña-Wilhelmi L, Rivas-Becerra J, Tatzber F, Pursch E, Tinahones FJ, García-Fuentes E, Garrido-Sánchez L. Oxidized LDL Modify the Human Adipocyte Phenotype to an Insulin Resistant, Proinflamatory and Proapoptotic Profile. Biomolecules. 2020; 10(4):534. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10040534
Chicago/Turabian StyleSantiago-Fernández, Concepción, Flores Martin-Reyes, Mónica Tome, Luis Ocaña-Wilhelmi, Jose Rivas-Becerra, Franz Tatzber, Edith Pursch, Francisco J. Tinahones, Eduardo García-Fuentes, and Lourdes Garrido-Sánchez. 2020. "Oxidized LDL Modify the Human Adipocyte Phenotype to an Insulin Resistant, Proinflamatory and Proapoptotic Profile" Biomolecules 10, no. 4: 534. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10040534