Structural and Quantitative Characterization of Mucin-Type O-Glycans and the Identification of O-Glycosylation Sites in Bovine Submaxillary Mucin

Abstract
:1. Introduction
2. Materials and Methods
2.1. Purity
2.2. O-Glycan Preparation
2.3. O-Glycan Profile
2.4. Structure and Quantification of O-Glycans
2.5. Glycopeptide Preparation
2.6. O-Glycosylation Site Analysis
3. Results
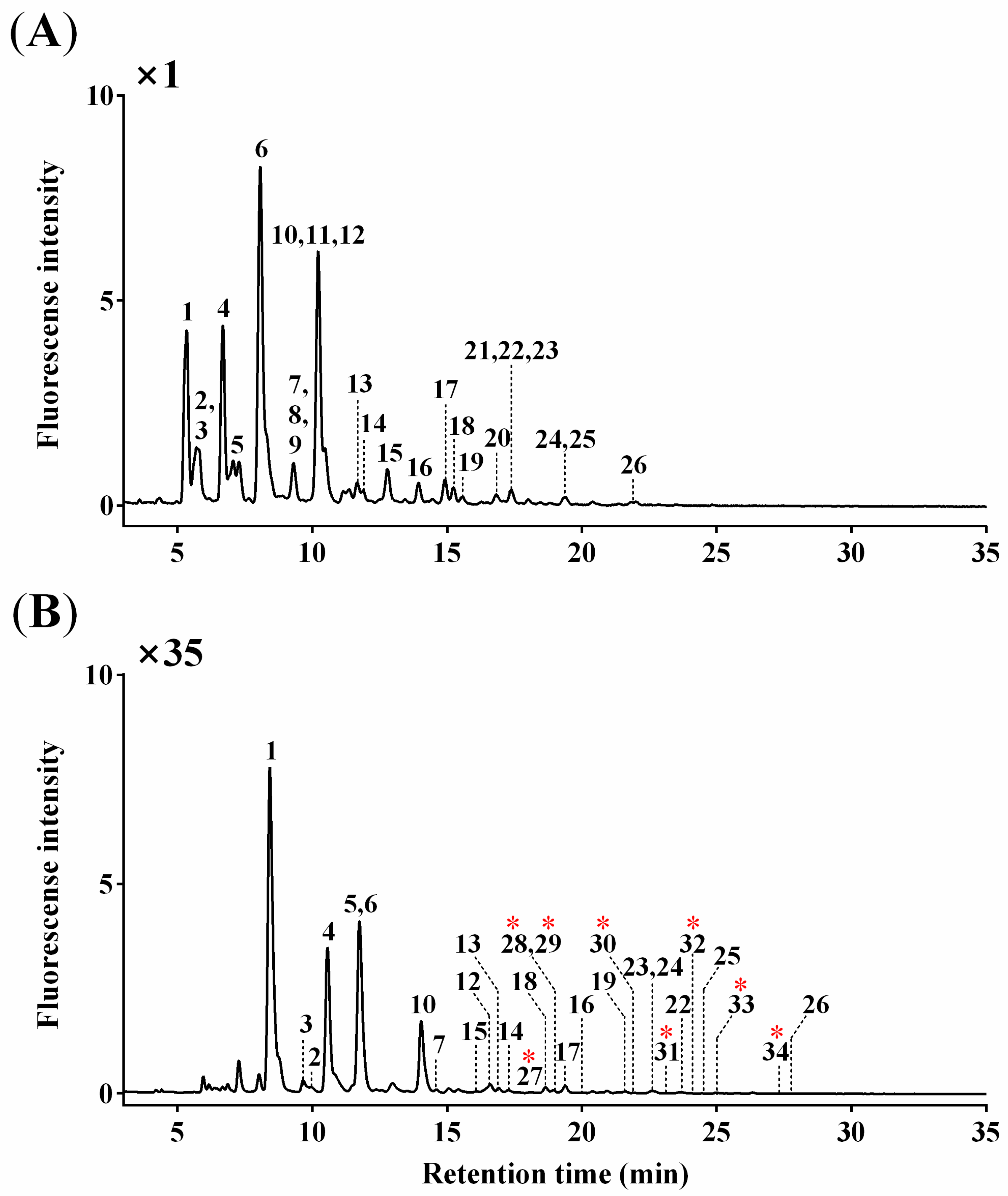
3.1. O-Glycan Profiles
3.2. O-Glycan Structures
3.3. Quantitative Characterization of O-Glycans
3.4. Glycopeptides Enrichment
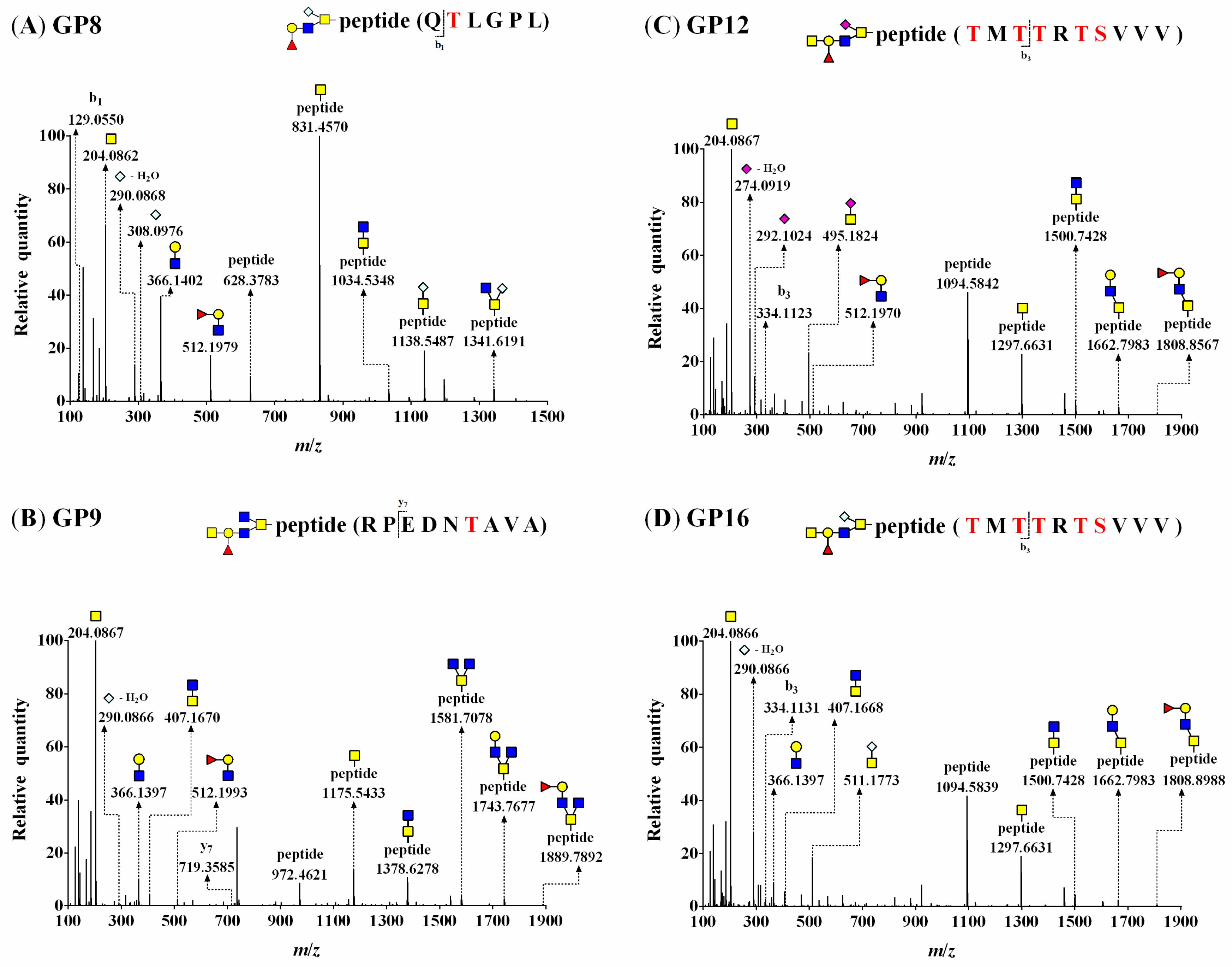
3.5. O-Glycopeptide Structures
3.6. O-Glycosylation Sites at Four BSM Domains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | amino acid |
| AB | 2-aminobenzamide |
| BSM | bovine submaxillary mucin |
| EIC | extracted ion chromatogram |
| ESI | electrospray ionization |
| Fuc | fucose |
| Gal | galactose |
| GalNAc | N-acetylgalactosamine |
| GlcNAc | N-acetylglucosamine |
| HCD | high-energy collisional dissociation |
| Hex | hexose |
| HexNAc | hexosamine |
| LC | liquid chromatography |
| MALDI | matrix-assisted laser desorption/ionization |
| MS | mass spectrometry |
| MS/MS | tandem mass spectrometry |
| Neu5Ac | N-acetylneuraminic acid |
| Neu5Gc | N-glycolylneuraminic acid |
| O-glycan | Ser/Thr-linked glycan |
| PMP | 1-phenyl-3-methyl-5-pyrazolone |
| ProA | procainamide |
| SA | sialic acid |
| SDS-PAGE | sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| TIC | total ion chromatogram |
| TOF | time-of-flight |
| UPLC | ultra-performance liquid chromatography |
References
- Bansil, R.; Stanley, E.; LaMont, J.T. Mucin biophysics. Annu. Rev. Physiol. 1995, 57, 635–657. [Google Scholar] [CrossRef] [PubMed]
- Jensen, P.H.; Kolarich, D.; Packer, N.H. Mucin-type O-glycosylation—Putting the pieces together. FEBS J. 2010, 277, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Petrou, G.; Crouzier, T. Mucins as multifunctional building blocks of biomaterials. Biomater. Sci. 2018, 6, 2282–2297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergstrom, K.S.B.; Xia, L. Mucin-type O-glycans and their roles in intestinal homeostasis. Glycobiology 2013, 23, 1026–1037. [Google Scholar] [CrossRef] [PubMed]
- Guzman-Aranguez, A.; Argüeso, P. Structure and biological roles of mucin-type O-glycans at the ocular surface. Ocul. Surf. 2010, 8, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Authimoolam, S.P.; Dziubla, T.D. Biopolymeric mucin and synthetic polymer analogs: Their structure, function and role in biomedical applications. Polymers 2016, 8, 71. [Google Scholar] [CrossRef] [Green Version]
- Sandberg, T.; Blom, H.; Caldwell, K.D. Potential use of mucins as biomaterial coatings. I. Fractionation, characterization, and model adsorption of bovine, porcine, and human mucins. J. Biomed. Mater. Res. A 2009, 91, 762–772. [Google Scholar] [CrossRef]
- Bansil, R.; Celli, J.P.; Hardcastle, J.M.; Turner, B.S. The influence of mucus microstructure and rheology in Helicobacter pylori infection. Front. Immunol. 2013, 4, 310. [Google Scholar] [CrossRef] [Green Version]
- Makarovsky, D.; Fadeev, L.; Salam, B.B.; Zelinger, E.; Matan, O.; Inbar, J.; Jurkevitch, E.; Gozin, M.; Burdman, S. Silver nanoparticles complexed with bovine submaxillary mucin possess strong antibacterial activity and protect against seedling infection. Appl. Environ. Microbiol. 2018, 84, e02212–e02217. [Google Scholar] [CrossRef] [Green Version]
- Bansil, R.; Turner, B.S. Mucin structure, aggregation, physiological functions and biomedical applications. Curr. Opin. Colloid Interface Sci. 2006, 11, 164–170. [Google Scholar] [CrossRef]
- Madsen, J.B.; Sotres, J.; Pakkanen, K.I.; Efler, P.; Svensson, B.; Hachem, M.A.; Arnebrant, T.; Lee, S. Structural and mechanical properties of thin films of bovine submaxillary mucin versus porcine gastric mucin on a hydrophobic surface in aqueous solutions. Langmuir 2016, 32, 9687–9696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Woitach, J.T.; Keil, R.L.; Bhavanandan, V.P. Bovine submaxillary mucin contains multiple domains and tandemly repeated non-identical sequences. Biochem. J. 1998, 331, 193–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banazadeh, A.; Veillon, L.; Wooding, K.M.; Zabet, M.; Mechref, Y. Recent advances in mass spectrometric analysis of glycoproteins. Electrophoresis 2017, 38, 162–189. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Mazumdar, S. Electrospray ionization mass spectrometry: A technique to access the information beyond the molecular weight of the analyte. Int. J. Anal. Chem. 2012, 2012, 282574. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Tolić, N.; Qu, Y.; Meng, D.; Zhao, R.; Zhang, Q.; Moore, R.J.; Zink, E.M.; Lipton, M.S.; Paša-Tolić, L.; et al. Characterization of intact N- and O-linked glycopeptides using higher energy collisional dissociation. Anal. Biochem. 2014, 452, 96–102. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Packer, N.H.; Lawson, M.A.; Jardine, D.R.; Redmond, J.W. A general approach to desalting oligosaccharides released from glycoproteins. Glycoconj. J. 1998, 15, 737–747. [Google Scholar] [CrossRef]
- Kim, J.; Lee, J.; Jang, Y.; Ha, J.; Kim, D.; Ji, M.; Lee, Y.K.; Kim, W.; You, S.; Do, J.; et al. N-glycans of bovine submaxillary mucin contain core-fucosylated and sulfated glycans but not sialylated glycans. Int. J. Biol. Macromol. 2019, 138, 1072–1078. [Google Scholar] [CrossRef]
- Park, H.; Kim, J.; Lee, Y.K.; Kim, W.; You, S.K.; Do, J.; Jang, Y.; Oh, D.B.; Kim, J.I.; Kim, H.H. Four unreported types of glycans containing mannose-6-phosphate are heterogeneously attached at three sites (including newly found Asn 233) to recombinant human acid alpha-glucosidase that is the only approved treatment for Pompe disease. Biochem. Biophys. Res. Commun. 2018, 495, 2418–2424. [Google Scholar] [CrossRef]
- Segu, Z.M.; Mechref, Y. Characterizing protein glycosylation sites through higher-energy C-trap dissociation. Rapid Commun. Mass Spectrom. 2010, 24, 1217–1225. [Google Scholar] [CrossRef]
- Kozak, R.P.; Royle, L.; Gardner, R.A.; Fernandes, D.L.; Wuhrer, M. Suppression of peeling during the release of O-glycans by hydrazinolysis. Anal. Biochem. 2012, 423, 119–128. [Google Scholar] [CrossRef]
- Keser, T.; Pavić, T.; Lauc, G.; Gornik, O. Comparison of 2-aminobenzamide, procainamide and RapiFluor-MS as derivatizing agents for high-throughput HILIC-UPLC-FLR-MS N-glycan analysis. Front. Chem. 2018, 6, 324. [Google Scholar] [CrossRef]
- Klapoetke, S.; Zhang, J.; Becht, S.; Gu, X.; Ding, X. The evaluation of a novel approach for the profiling and identification of N-linked glycan with a procainamide tag by HPLC with fluorescent and mass spectrometric detection. J. Pharm. Biomed. Anal. 2010, 53, 315–324. [Google Scholar] [CrossRef]
- Li, Z.; Chai, W. Mucin O-glycan microarrays. Curr. Opin. Struct. Biol. 2019, 56, 187–197. [Google Scholar] [CrossRef]
- Ruhaak, L.R.; Zauner, G.; Huhn, C.; Bruggink, C.; Deelder, A.M.; Wuhrer, M. Glycan labeling strategies and their use in identification and quantification. Anal. Bioanal. Chem. 2010, 397, 3457–3481. [Google Scholar] [CrossRef] [Green Version]
- Madsen, J.B.; Svensson, B.; Hachem, M.A.; Lee, S. Proteolytic degradation of bovine submaxillary mucin (BSM) and its impact on adsorption and lubrication at a hydrophobic surface. Langmuir 2015, 31, 8303–8309. [Google Scholar] [CrossRef]
- Kotsias, M.; Kozak, R.P.; Gardner, R.A.; Wuhrer, M.; Spencer, D.I.R. Improved and semi-automated reductive β-elimination workflow for higher throughput protein O-glycosylation analysis. PLoS ONE 2019, 14, e0210759. [Google Scholar] [CrossRef] [Green Version]
- Reiding, K.R.; Blank, D.; Kuijper, D.M.; Deelder, A.M.; Wuhrer, M. High-throughput profiling of protein N-glycosylation by MALDI-TOF-MS employing linkage-specific sialic acid esterification. Anal. Chem. 2014, 86, 5784–5793. [Google Scholar] [CrossRef]
- Reusch, D.; Haberger, M.; Falck, D.; Peter, B.; Maier, B.; Gassner, J.; Hook, M.; Wagner, K.; Bonnington, L.; Bulau, P.; et al. Comparison of methods for the analysis of therapeutic immunoglobulin G Fc-glycosylation profiles-part 2: Mass spectrometric methods. MAbs 2015, 7, 732–742. [Google Scholar] [CrossRef]
- Zauner, G.; Koeleman, C.A.M.; Deelder, A.M.; Wuhrer, M. Mass spectrometric O-glycan analysis after combined O-glycan release by beta-elimination and 1-phenyl-3-methyl-5-pyrazolone labeling. Biochim. Biophys. Acta. 2012, 1820, 1420–1428. [Google Scholar] [CrossRef]
- Kim, W.; Kim, J.; You, S.; Do, J.; Jang, Y.; Kim, D.; Lee, J.; Ha, J.; Kim, H.H. Qualitative and quantitative characterization of sialylated N-glycans using three fluorophores, two columns, and two instrumentations. Anal. Biochem. 2019, 571, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.R.; Gao, W.N.; Grimm, R.; Jiang, S.; Liang, Y.; Ye, H.; Li, Z.G.; Yau, L.F.; Huang, H.; Liu, J.; et al. A method to identify trace sulfated IgG N-glycans as biomarkers for rheumatoid arthritis. Nat. Commun. 2017, 8, 631. [Google Scholar] [CrossRef] [PubMed]
- Van Den Steen, P.; Rudd, P.M.; Dwek, R.A.; Opdenakker, G. Concepts and principles of O-linked glycosylation. Crit. Rev. Biochem. Mol. Biol. 1998, 33, 151–208. [Google Scholar] [CrossRef] [PubMed]
- Hart, G.W.; Akimoto, Y. Essentials of Glycobiology, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2009; p. 800. [Google Scholar]
- Hussain, M.R.M.; Din, N.; Hassan, M.; Razaq, A.; Iqbal, Z. Physiological significance of Fuc and Sialic acid containing glycans in the body. Arab. J. Chem. 2016, 9, S9–S20. [Google Scholar] [CrossRef] [Green Version]
- Mattu, T.S.; Royle, L.; Langridge, J.; Wormald, M.R.; Van den Steen, P.E.; Van Damme, J.; Opdenakker, G.; Harvey, D.J.; Dwek, R.A.; Rudd, P.M. O-glycan analysis of natural human neutrophil gelatinase B using a combination of normal phase-HPLC and online tandem mass spectrometry: Implications for the domain organization of the enzyme. Biochemistry 2000, 39, 15695–15704. [Google Scholar] [CrossRef]
- Bergstrom, K.; Fu, J.; Xia, L. Glycoscience: Biology and Medicine; Springer Japan: Tokyo, Japan, 2015; pp. 1073–1080. [Google Scholar]
- Zaretsky, J.Z.; Wreschner, D.H. Gel-Forming and Soluble Mucins; Bentham Science: Sharjah, UAE, 2013; p. 647. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No a | Proposed Structure b | Relative Quantity (%) c (Category) d | Mass (m/z) e | Mass Error (ppm) f | Peak No a | Proposed Structure b | Relative Quantity (%) c (Category) d | Mass (m/z) e | Mass Error (ppm) f | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Theoretical [M + H]+ | Observed [M + H]+ | Theoretical [M + H]+ | Observed [M + H]+ | ||||||||
| 1 |  | 9.3 (Ma, 2, S) | 633.2614 | 633.2611 | 0.4 | 18 |  | 0.7 (Mi, 4, S, G, C3) | 1014.3885 | 1014.3885 | 0.0 |
| 2 |  | 1.1 (Ma, 3, N, C1) | 707.2982 | 707.2978 | 0.5 | 19 |  | 0.5 (Mi, 6, N, F, C4) | 1259.5148 | 1259.5148 | 0.0 |
| 3 |  | 0.7 (Mi, 3, N, C4) | 748.3247 | 748.3245 | 0.3 | 20 |  | 0.1 (Mi, 6, N, F, C4) | 1202.4933 | 1202.4937 | −0.3 |
| 4 |  | 11.0 (Ma, 2, S) | 649.2563 | 649.2560 | 0.4 | 21 |  | 0.6(Mi, 5, S, F, C3) | 1160.4464 | 1160.4464 | 0.0 |
| 5 |  | 4.6 (Ma, 4, N, F, C2) | 853.3561 | 853.3558 | 0.3 | 22 |  | 0.4 (Mi, 7, N, F, C4) | 1364.5462 | 1364.5465 | −0.2 |
| 6 |  | 32.6 (Ma, 3, S, C3) | 836.3408 | 836.3399 | 1.0 | 23 |  | 0.1 (Mi, 6, S, F, C3) | 1306.5043 | 1306.5033 | 0.8 |
| 7 |  | 1.7 (Ma, 4, N, C2) | 910.3775 | 910.3770 | 0.6 | 24 |  | 0.3 (Mi, 6, S, F, C3) | 1347.5309 | 1347.5308 | 0.1 |
| 8 |  | 1.5 (Ma, 3, S, C1) | 795.3142 | 795.3137 | 0.6 | 25 |  | 0.1 (Mi, 7, N, F, C2) | 1307.5247 | 1307.5247 | 0.0 |
| 9 |  | 0.4 (Mi, 4, N, C4) | 951.4041 | 951.4039 | 0.2 | 26 |  | 0.1 (Mi, 8, N, F, C2) | 1567.6255 | 1567.6254 | 0.1 |
| 10 |  | 20.5 (Ma, 3, S, C5) | 852.3357 | 852.3349 | 1.0 | 27 |  | - (T, 5, N, F, C3) | 1098.5188 | 1098.5190 | −0.2 |
| 11 |  | 1.0 (Ma, 4, S, F, C1) | 941.3721 | 941.3721 | 0.0 | 28 |  | - (T, 5, N, G, C2) | 1171.5352 | 1171.5353 | −0.1 |
| 12 |  | 0.7 (Mi, 4, N, G, C2) | 869.3510 | 869.3505 | 0.6 | 29 |  | - (T, 5, N, G, C4) | 1212.5617 | 1212.5617 | 0.0 |
| 13 |  | 3.1 (Ma, 5, N, F, C4) | 1056.4354 | 1056.4353 | 0.1 | 30 |  | - (T, 6, N, F, C2) | 1317.5931 | 1317.5933 | −0.2 |
| 14 |  | 2.2 (Ma, 5, N, F, G, C2) | 1015.4089 | 1015.4089 | 0.0 | 31 |  | - (T, 6, N, F, G, C2) | 1276.5665 | 1276.5665 | 0.0 |
| 15 |  | 2.3 (Ma, 4, S, C2) | 998.3936 | 998.3931 | 0.5 | 32 |  | - (T, 6, S, F, C3) | 1389.6142 | 1389.6145 | 0.0 |
| 16 |  | 2.4 (Ma, 6, N, F, C2) | 1161.4668 | 1161.4667 | 0.1 | 33 |  | - (T, 6, S, F, C3) | 1462.6306 | 1462.6305 | 0.0 |
| 17 |  | 2.0 (Ma, 5, S, F, C3) | 1144.4515 | 1144.4514 | 0.1 | 34 |  | - (T, 7, N, F, C2) | 1504.6775 | 1504.6773 | 0.2 |
| Name | O-Glycopeptide a | Peptide Mass (m/z) | O-Glycopeptide Mass (m/z) b | Attached O-Glycan d | Glycan No e | |||
|---|---|---|---|---|---|---|---|---|
| Theoretical [M + H]+ | Observed [M + H]+ | Theoretical [M + H]+ | Observed [M + H]+ | Error (ppm)c | ||||
| GP1 | 307RPSYGAL | 763.4097 | 763.4146 | 730.8356 | 730.8362 | 0.8 |  | G6 |
| GP2 | 625QTLGPL | 628.3665 | 628.3776 | 663.3140 | 663.3190 | 7.7 |  | G6 |
| GP3 | 1080RPEDNTAVA | 972.4745 | 972.4632 | 835.3680 | 835.3622 | −6.9 |  | G6 |
| GP4 | 307RPSYGAL | 763.4097 | 763.4107 | 738.8330 | 738.8342 | 1.5 |  | G10 |
| GP5 | 625QTLGPL | 628.3665 | 628.3776 | 671.3114 | 671.3165 | 7.6 |  | G10 |
| GP6 | 1080RPEDNTAVA | 972.4745 | 972.4631 | 843.3654 | 843.3608 | −5.5 |  | G10 |
| GP7 | 625QTLGPL | 628.3665 | 628.3773 | 561.7743 | 561.7794 | 9.1 |  | G1 |
| GP8 | 625QTLGPL | 628.3665 | 628.3783 | 825.3668 | 825.3721 | 6.5 |  | G21 |
| GP9 | 1080RPEDNTAVA | 972.4745 | 972.4621 | 1046.9550 | 1046.9504 | −4.4 |  | G19 |
| GP10 | 56SGETRTSVI | 949.4949 | 949.4953 | 722.3385 | 722.3384 | −0.1 |  | G1 |
| GP11 | 272GSPSSVSSAEQI | 1148.5430 | 1148.5795 | 923.4022 | 923.4198 | 19.0 |  | G6 |
| GP12 | 728TMTTRTSVVV | 1094.5874 | 1094.5842 | 1152.0195 | 1152.0195 | 0.0 |  | G24 |
| GP13 | 259SHSSSGRSRTI | 1174.5923 | 1174.6086 | 1017.4533 | 1017.4581 | 4.7 |  | G15 |
| GP14 | 56SGETRTSVI | 949.4949 | 949.4957 | 730.3359 | 730.3360 | 0.1 |  | G4 |
| GP15 | 272GSPSSVSSAEQI | 1148.5430 | 1148.5803 | 931.3997 | 931.4170 | 18.6 |  | G10 |
| GP16 | 728TMTTRTSVVV | 1094.5874 | 1094.5839 | 1160.0169 | 1160.0167 | −0.2 |  | G33 |
| GP17 | 259SHSSSGRSRTI | 1174.5923 | 1174.6099 | 1025.4508 | 1025.4579 | 7.0 |  | G18 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.; Ryu, C.; Ha, J.; Lee, J.; Kim, D.; Ji, M.; Park, C.S.; Lee, J.; Kim, D.K.; Kim, H.H. Structural and Quantitative Characterization of Mucin-Type O-Glycans and the Identification of O-Glycosylation Sites in Bovine Submaxillary Mucin. Biomolecules 2020, 10, 636. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10040636
Kim J, Ryu C, Ha J, Lee J, Kim D, Ji M, Park CS, Lee J, Kim DK, Kim HH. Structural and Quantitative Characterization of Mucin-Type O-Glycans and the Identification of O-Glycosylation Sites in Bovine Submaxillary Mucin. Biomolecules. 2020; 10(4):636. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10040636
Chicago/Turabian StyleKim, Jihye, Changsoo Ryu, Jongkwan Ha, Junmyoung Lee, Donghwi Kim, Minkyoo Ji, Chi Soo Park, Jaeryong Lee, Dae Kyong Kim, and Ha Hyung Kim. 2020. "Structural and Quantitative Characterization of Mucin-Type O-Glycans and the Identification of O-Glycosylation Sites in Bovine Submaxillary Mucin" Biomolecules 10, no. 4: 636. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10040636