In Vitro Digestion of Grape Seed Oil Inhibits Phospholipid-Regulating Effects of Oxidized Lipids

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. Grape Seed Oil Samples
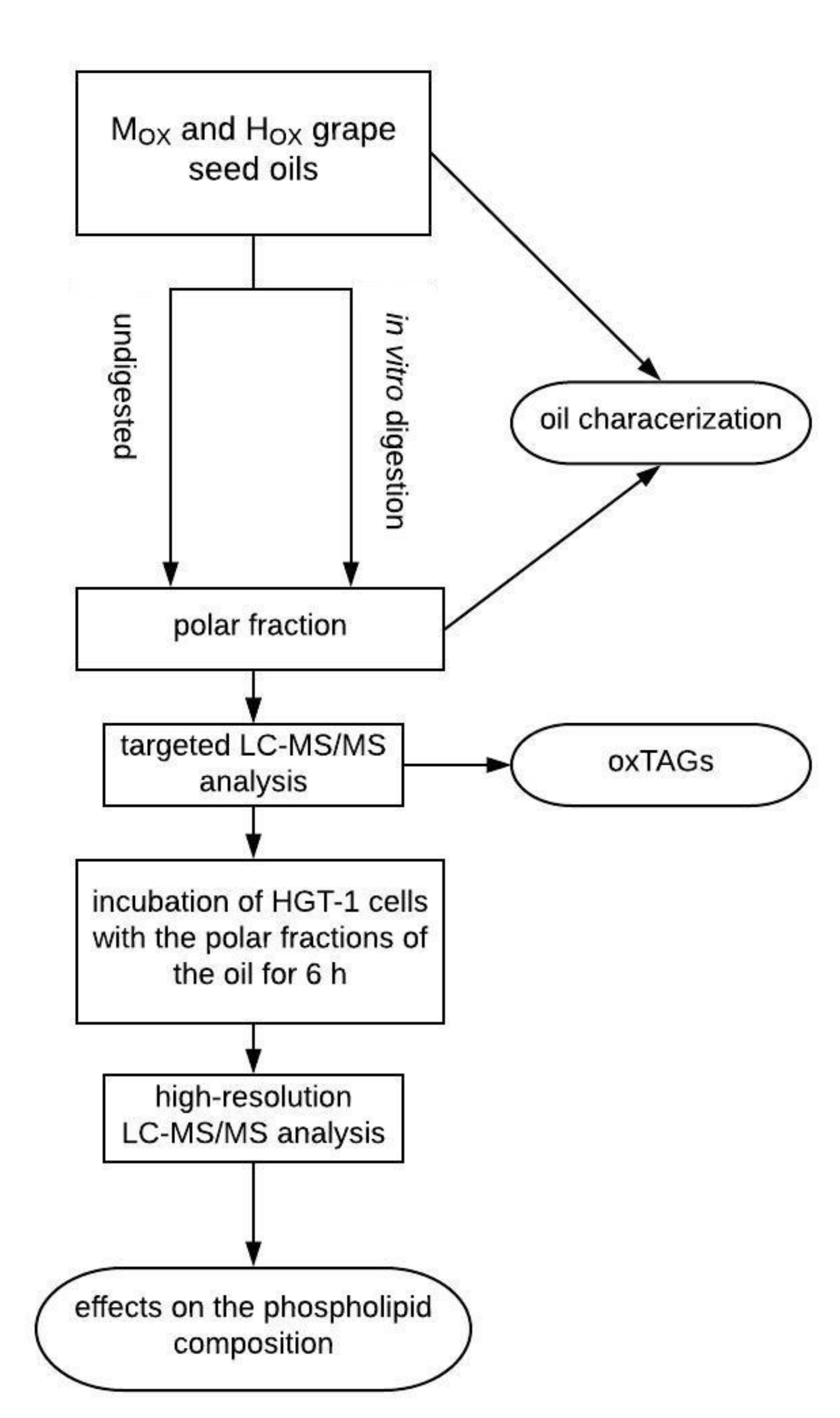
2.3. Study Design
2.3.1. Gastric In Vitro Digestion Model
2.3.2. Sample Preparation
2.4. Characterisation of the Grape Seed Oils
2.4.1. GC/FID Analysis of the Fatty Acid Composition
2.4.2. Peroxide Value
2.4.3. HS-GC/MS Quantitation of Hexanal
2.4.4. Extraction and High-Resolution LC-MS Analysis of Polyphenols
2.4.5. HPLC/UV Analysis of Tocopherols
2.5. Cell Culture
2.5.1. Cell Viability
2.5.2. Incubation of HGT-1 Cells and Cellular Extraction of Phospholipids
2.6. LC-MS and LC-MS/MS Analysis
2.6.1. Targeted Analysis by LC-MS/MS
2.6.2. High-Resolution LC-MS and LC-MS/MS Analysis
2.6.3. Comparative Analysis of Digested and Undigested Samples
2.7. Statistical Analysis
3. Results and Discussion
3.1. Characterisation of Cold-Pressed Grape Seed Oil
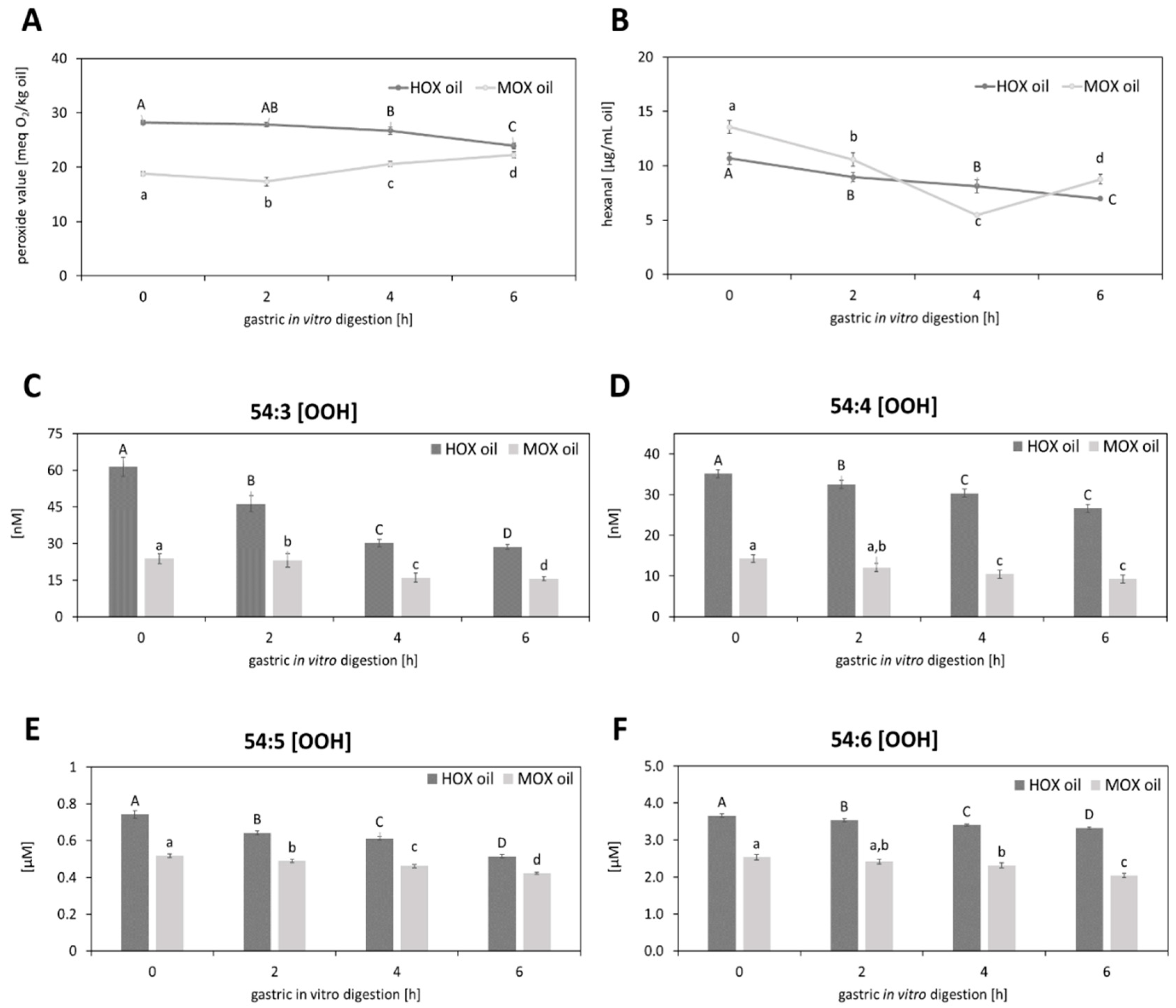
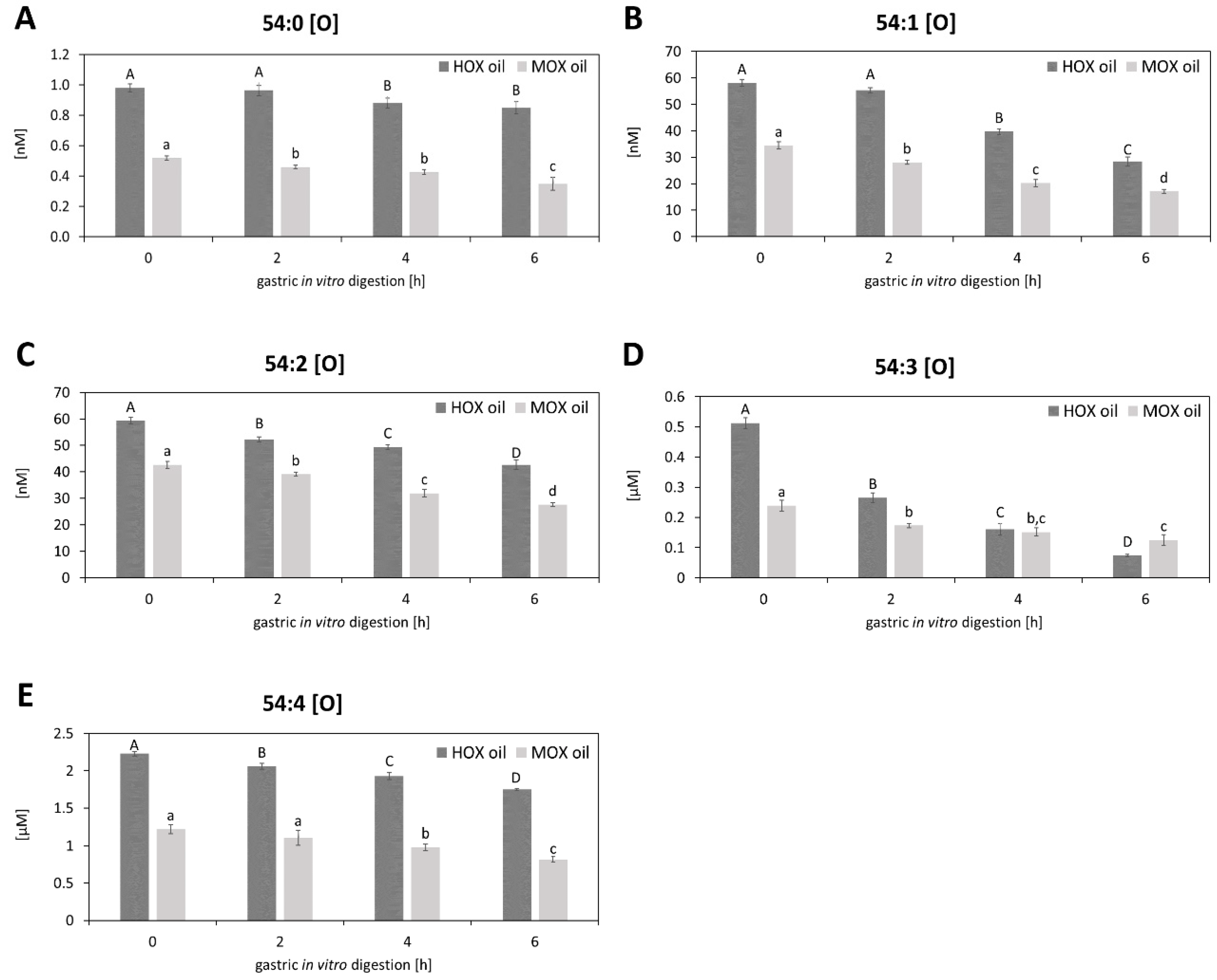
3.2. Effects of Gastric In Vitro Digestion on Cold-Pressed Grape Seed Oil
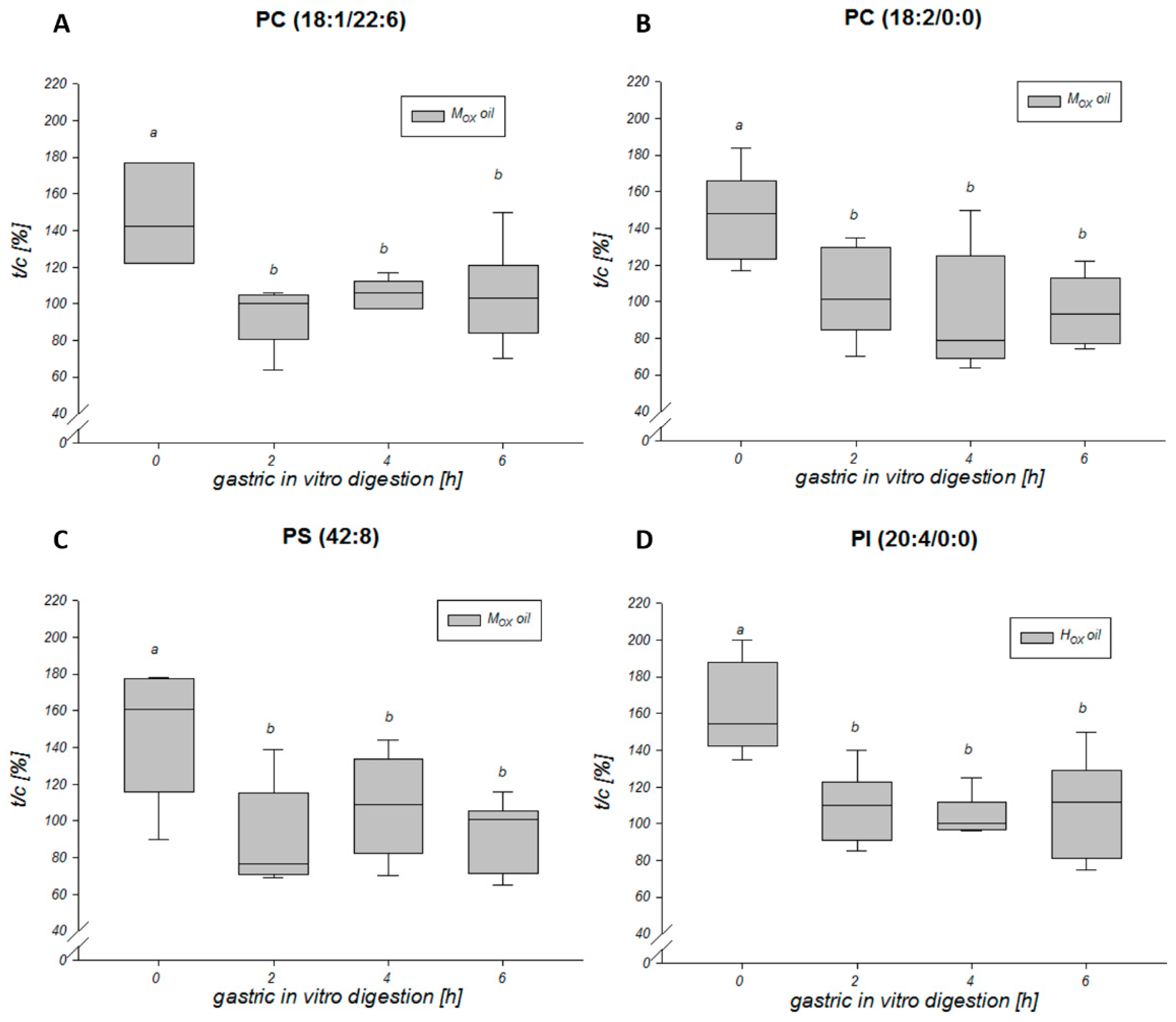
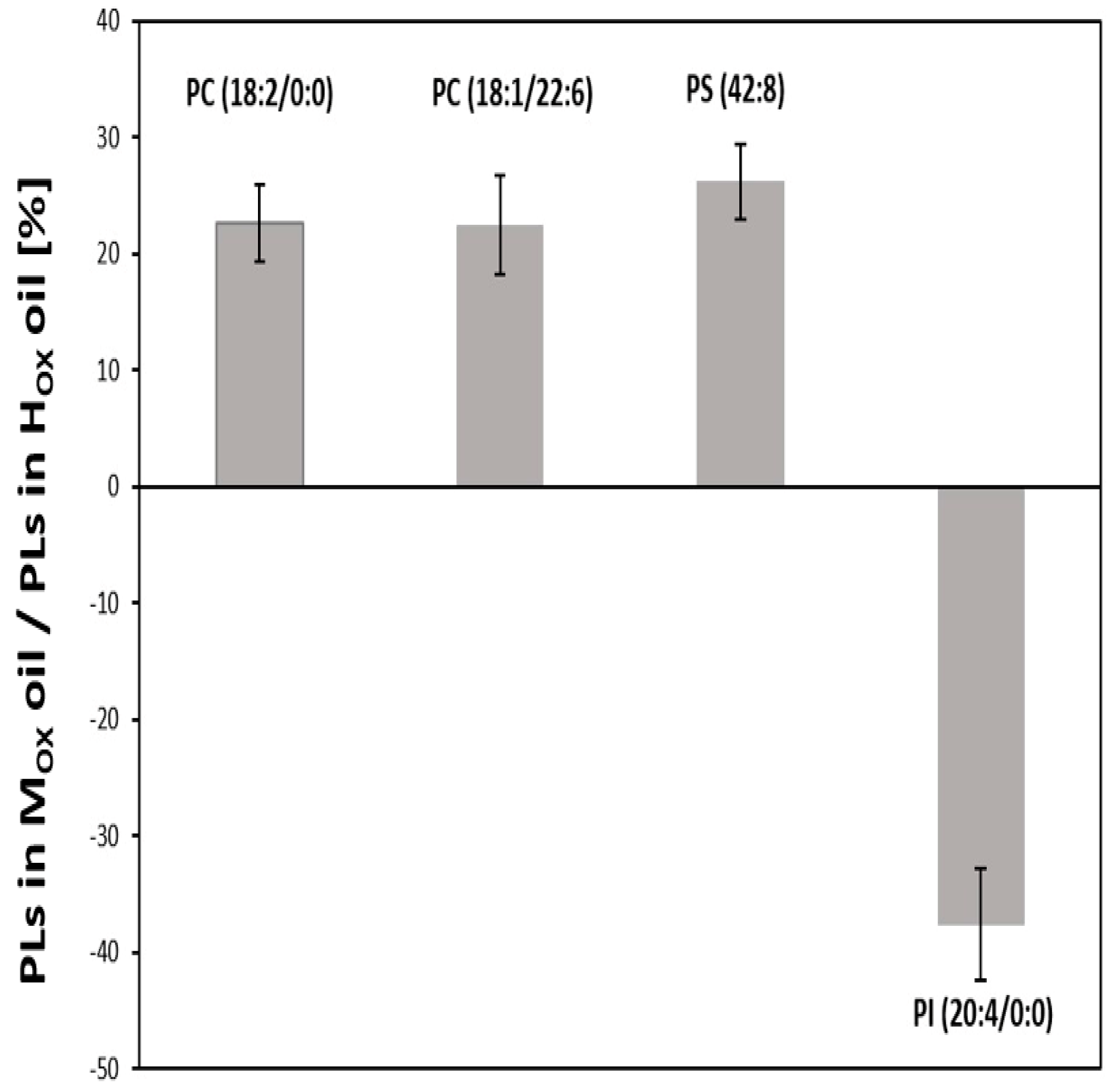
3.3. Effects of Oxidized Lipids on Phospholipid Composition in HGT-1 Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shahidi, F.; Zhong, Y. Lipid oxidation and improving the oxidative stability. Chem. Soc. Rev. 2010, 39, 4067–4079. [Google Scholar] [CrossRef] [PubMed]
- Abbott, S.K.; Else, P.L.; Hulbert, A.J. Membrane fatty acid composition of rat skeletal muscle is most responsive to the balance of dietary n-3 and n-6 PUFA. Br. J. Nutr. 2010, 103, 522–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutterodt, H.; Slavin, M.; Whent, M.; Turner, E.; Yu, L. Fatty acid composition, oxidative stability, antioxidant and antiproliferative properties of selected cold-pressed grape seed oils and flours. Food Chem. 2011, 128, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Choe, E.; Min, D.B. Mechanisms and Factors for Edible Oil Oxidation. Compr. Rev. Food Sci. Food Saf. 2006, 5, 169–186. [Google Scholar] [CrossRef]
- Kanner, J. Dietary advanced lipid oxidation endproducts are risk factors to human health. Mol. Nutr. Food Res. 2007, 51, 1094–1101. [Google Scholar] [CrossRef]
- Frey, L.; Hiller, S.; Riek, R.; Bibow, S. Lipid- and Cholesterol-Mediated Time-Scale-Specific Modulation of the Outer Membrane Protein X Dynamics in Lipid Bilayers. J. Am. Chem. Soc. 2018, 140, 15402–15411. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Rong, X.; Duerr, M.A.; Hermanson, D.J.; Hedde, P.N.; Wong, J.S.; Vallim, T.Q.; Cravatt, B.F.; Gratton, E.; Ford, D.A.; et al. Intestinal Phospholipid Remodeling Is Required for Dietary-Lipid Uptake and Survival on a High-Fat Diet. Cell Metab. 2016, 23, 492–504. [Google Scholar] [CrossRef] [Green Version]
- Zaunschirm, M.; Pignitter, M.; Kopic, A.; Keßler, C.; Hochkogler, C.; Kretschy, N.; Somoza, M.M.; Somoza, V. Exposure of Human Gastric Cells to Oxidized Lipids Stimulates Pathways of Amino Acid Biosynthesis on a Genomic and Metabolomic Level. Molecules 2019, 24. [Google Scholar] [CrossRef] [Green Version]
- Koziolek, M.; Carriere, F.; Porter, C.J.H. Lipids in the Stomach—Implications for the Evaluation of Food Effects on Oral Drug Absorption. Pharm. Res. 2018, 35, 55. [Google Scholar] [CrossRef]
- Kanner, J.; Lapidot, T. The stomache as a bioreactor: Dietary lipid peroxidation in the gastric fluid and the effects of plant-derived antioxidants. Free Radical. Bio. Med. 2001, 31, 1388–1395. [Google Scholar] [CrossRef]
- Ohnishi, M.; Hirose, S.; Kawaguchi, M.; Ito, S.; Fujino, Y. Chemical Composition of Lipids, Especially Triacylglycerol, in Grape Seeds. Agric. Biol. Chem. 2014, 54, 1035–1042. [Google Scholar] [CrossRef] [Green Version]
- Demirtas, I.; Pelvan, E.; Özdemir, İ.S.; Alasalvar, C.; Ertas, E. Lipid characteristics and phenolics of native grape seed oils grown in Turkey. Eur. J. Lipid. Sci. Tech. 2013, 115, 641–647. [Google Scholar] [CrossRef]
- Pignitter, M.; Dumhart, B.; Gartner, S.; Jirsa, F.; Steiger, G.; Kraemer, K.; Somoza, V. Vitamin A is rapidly degraded in retinyl palmitate-fortified soybean oil stored under household conditions. J. Agric. Food. Chem. 2014, 62, 7559–7566. [Google Scholar] [CrossRef] [PubMed]
- Grüneis, V.; Fruehwirth, S.; Zehl, M.; Ortner, J.; Schamann, A.; König, J.; Pignitter, M. Simultaneous Analysis of Epoxidized and Hydroperoxidized Triacylglycerols in Canola Oil and Margarine by LC-MS. J. Agric. Food Chem. 2019, 67, 10174–10184. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, D.H. Peroxide formation as a measure of autoxidative deterioriation. Oil Soap 1932, 9, 89–97. [Google Scholar] [CrossRef]
- Versantvoort, C.H.; Oomen, A.G.; Van de Kamp, E.; Rompelberg, C.J.; Sips, A.J. Applicability of an in vitro digestion model in assessing the bioaccessibility of mycotoxins from food. Food Chem. Toxicol. 2005, 43, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Nieva-Echevarría, B.; Goicoechea, E.; Manzanos, M.J.; Guillen, M.D. A study by (1)H NMR on the influence of some factors affecting lipid in vitro digestion. Food Chem. 2016, 211, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Cortot, A.; Phillips, S.F.; Malagelada, J.-R. Gastric emptying of lipids after ingestion of a solid-liquid meal in humans. Gastroenterology 1981, 80, 922–927. [Google Scholar] [CrossRef]
- Márquez-Ruiz, G.; Jorge, N.; Martín-Polvillo, M.; Dobarganes, M.C. Rapid, quantitative determination of polar compounds in fats and oils by solid-phase extraction and size-exclusion chromatography using monostearin as internal standard. J. Chromatogr. A 1996, 749, 55–60. [Google Scholar] [CrossRef]
- ICH. International Conference on Harmonization of Technical Requirements for the Registration of Pharmaceuticals for Human Use, Validation of Analytical Procedures: Text and Methodology Q2(R1); ICH-Q2B; ICH: Geneva, Switzerland, 1996. [Google Scholar]
- Shrivastava, A.; Gupta, V.B. Methods for the determination of limit of detection and limit of quantitation of the analytical methods. Chron. Young Sci. 2011, 2. [Google Scholar] [CrossRef]
- Giuffrida, F.; Golay, P.-A.; Destaillats, F.; Hug, B.; Dionisi, F. Accurate determination of hexanal in beef bouillons by headspace solid-phase microextraction gas-chromatography mass-spectrometry. Eur. J. Lipid. Sci. Tech. 2005, 107, 792–798. [Google Scholar] [CrossRef]
- Pignitter, M.; Stolze, K.; Gartner, S.; Dumhart, B.; Stoll, C.; Steiger, G.; Kraemer, K.; Somoza, V. Cold fluorescent light as major inducer of lipid oxidation in soybean oil stored at household conditions for eight weeks. J. Agric. Food Chem. 2014, 62, 2297–2305. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic and reagents. Am. J. Enol. Viticult. 1965, 16, 144–158. [Google Scholar]
- Liszt, K.I.; Ley, J.P.; Lieder, B.; Behrens, M.; Stoger, V.; Reiner, A.; Hochkogler, C.M.; Kock, E.; Marchiori, A.; Hans, J.; et al. Caffeine induces gastric acid secretion via bitter taste signaling in gastric parietal cells. Proc. Natl. Acad. Sci. USA 2017, 114, E6260–E6269. [Google Scholar] [CrossRef] [Green Version]
- Stoeger, V.; Liszt, K.I.; Lieder, B.; Wendelin, M.; Zopun, M.; Hans, J.; Ley, J.P.; Krammer, G.E.; Somoza, V. Identification of Bitter-Taste Intensity and Molecular Weight as Amino Acid Determinants for the Stimulating Mechanisms of Gastric Acid Secretion in Human Parietal Cells in Culture. J. Agric. Food Chem. 2018, 66, 6762–6771. [Google Scholar] [CrossRef]
- Liszt, K.I.; Eder, R.; Wendelin, S.; Somoza, V. Identification of Catechin, Syringic Acid, and Procyanidin B2 in Wine as Stimulants of Gastric Acid Secretion. J. Agric. Food Chem. 2015, 63, 7775–7783. [Google Scholar] [CrossRef]
- Hoi, J.K.; Lieder, B.; Pignitter, M.; Hans, J.; Ley, J.P.; Lietard, J.; Hoelz, K.; Somoza, M.; Somoza, V. Identification of Cinnamaldehyde as Most Effective Fatty Acid Uptake Reducing Cinnamon-Derived Compound in Differentiated Caco-2 Cells Compared to Its Structural Analogues Cinnamyl Alcohol, Cinnamic Acid, and Cinnamyl Isobutyrate. J. Agric. Food Chem. 2019, 67, 11638–11649. [Google Scholar] [CrossRef]
- Zhang, H.; Gao, Y.; Sun, J.; Fan, S.; Yao, X.; Ran, X.; Zheng, C.; Huang, M.; Bi, H. Optimization of lipid extraction and analytical protocols for UHPLC-ESI-HRMS-based lipidomic analysis of adherent mammalian cancer cells. Anal. Bioanal. Chem. 2017, 409, 5349–5358. [Google Scholar] [CrossRef]
- Pignitter, M.; Zaunschirm, M.; Lach, J.; Unterberger, L.; Kopic, A.; Kessler, C.; Kienesberger, J.; Pischetsrieder, M.; Eggersdorfer, M.; Riegger, C.; et al. Regioisomeric distribution of 9- and 13-hydroperoxy linoleic acid in vegetable oils during storage and heating. J. Sci. Food Agric. 2018, 98, 1240–1247. [Google Scholar] [CrossRef] [Green Version]
- Ovcharova, T.; Zlatanov, M.; Ivanov, A. Changes in grape seed oil during fermentation. Eur. Int. J. Sci. Technol. 2014, 3, 178–187. [Google Scholar]
- Tautenhahn, R.; Patti, G.J.; Rinehart, D.; Siuzdak, G. XCMS Online: A Web-Based Platform to Process Untargeted Metabolomic Data. Anal. Chem. 2012, 84, 5035–5039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, M.C.M.B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; Hoff, K.; et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef] [PubMed]
- Prince, J.T.; Marcotte, E.M. Chromatographic Alignment of ESI-LC-MS Proteomics Data Sets by Ordered Bijective Interpolated Warping. Anal. Chem. 2006, 78, 6140–6152. [Google Scholar] [CrossRef] [PubMed]
- Tautenhahn, R.; Bottcher, C.; Neumann, S. Highly sensitive feature detection for high resolution LC/MS. BMC Bioinform. 2008, 9, 504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beveridge, T.H.J.; Girard, B.; Kopp, T.; Drover, J.C.G. Yield and composition of grape seed oils extracted by supercritical carbon dioxide and petroleum ether: Varietal effects. J. Agric. Food Chem. 2005, 53, 1799–1804. [Google Scholar] [CrossRef] [PubMed]
- Sabir, A.; Unver, A.; Kara, Z. The fatty acid and tocopherol constituents of the seed oil extracted from 21 grape varieties (Vitis spp.). J. Agric. Food Chem. 2012, 92, 1982–1987. [Google Scholar] [CrossRef]
- Maszewska, M.; Florowska, A.; Dluzewska, E.; Wroniak, M.; Marciniak-Lukasiak, K.; Zbikowska, A. Oxidative Stability of Selected Edible Oils. Molecules 2018, 23. [Google Scholar] [CrossRef] [Green Version]
- The-Codex-Alimentarius. Joint FAO/WHO Food Standards Programme Codex Alimentarius Commission, 2nd ed.; FAO: Codex Alimentarius Commission: World Health Organization, Food and Agriculture Organization of the United Nations: Rome, Italy, 2001; Volume 8. [Google Scholar]
- Shahidi, F.; Pegg, R.B. Hexanal as an Indicator of the Flavor Deterioration of Meat and Meat Products. Lipids in Food Flavors; American Chemical Society: Washington, DC, USA, 1994. [Google Scholar]
- Fernandes, L.; Casal, S.; Cruz, R.; Pereira, J.A.; Ramalhosa, E. Seed oils of ten traditional Portuguese grape varieties with interesting chemical and antioxidant properties. Food Res. Int. 2013, 50, 161–166. [Google Scholar] [CrossRef]
- Assumpção, C.F.; Nunes, I.L.; Mendonça, T.A.; Bortolin, R.C.; Jablonski, A.; Flôres, S.H.; de Oliveira Rios, A. Bioactive Compounds and Stability of Organic and Conventional Vitis labrusca Grape Seed Oils. J. Am. Oil Chem. Soc. 2015, 93, 115–124. [Google Scholar] [CrossRef]
- Vieira, S.A.; Zhang, G.; Decker, E.A. Biological Implications of Lipid Oxidation Products. J. Am. Chem. Soc. 2017, 94, 339–351. [Google Scholar] [CrossRef]
- Kenmogne-Domguia, H.B.; Moisan, S.; Viau, M.; Genot, C.; Meynier, A. The initial characteristics of marine oil emulsions and the composition of the media inflect lipid oxidation during in vitro gastrointestinal digestion. Food Chem. 2014, 152, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Chew, S.-C.; Tan, C.-P.; Long, K.; Nyam, K.-L. In-vitro evaluation of kenaf seed oil in chitosan coated-high methoxyl pectin-alginate microcapsules. Ind. Crops Prod. 2015, 76, 230–236. [Google Scholar] [CrossRef]
- Cheong, A.M.; Tan, C.P.; Nyam, K.L. In-vitro gastrointestinal digestion of kenaf seed oil-in-water nanoemulsions. Ind. Crops Prod. 2016, 87, 1–8. [Google Scholar] [CrossRef]
- Garavaglia, J.; Markoski, M.M.; Oliveira, A.; Marcadenti, A. Grape Seed Oil Compounds: Biological and Chemical Actions for Health. Nutr. Metab. Insight 2016, 9, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjelica, M.; Vujasinović, V.; Rabrenović, B.; Dimić, S. Some Chemical Characteristics and Oxidative Stability of Cold Pressed Grape Seed Oils Obtained from Different Winery Waste. Eur. J. Lipid. Sci. Tech. 2019, 121. [Google Scholar] [CrossRef]
- Choe, E.; Min, D.B. Mechanisms of Antioxidants in the Oxidation of Foods. Compr. Rev. Food Sci. Food Saf. 2009, 8, 345–358. [Google Scholar] [CrossRef]
- Dai, F.; Chen, W.F.; Zhou, B. Antioxidant synergism of green tea polyphenols with alpha-tocopherol and L-ascorbic acid in SDS micelles. Biochimie 2008, 90, 1499–1505. [Google Scholar] [CrossRef]
- Réblová, Z.; Okrouhlá, P. Ability of Phenolic Acids to Protect α-Tocopherol. Czech J. Food Sci. 2010, 28, 290–297. [Google Scholar]
- Pignitter, M.; Somoza, V. Critical evaluation of methods for the measurement of oxidative rancidity in vegetable oils. J. Food Drug Anal. 2012, 20, 772–777. [Google Scholar] [CrossRef]
- Barriuso, B.; Astiasarán, I.; Ansorena, D. A review of analytical methods measuring lipid oxidation status in foods: A challenging task. Eur. Food Res. Technol. 2012, 236, 1–15. [Google Scholar] [CrossRef]
- N’Goma, J.-C.B.; Amara, S.; Dridi, K.; Jannin, V.; Carrière, F. Understanding the lipid-digestion processes in the GI tract before designing lipid-based drug-delivery systems. Ther. Deliv. 2012, 3, 105–124. [Google Scholar] [CrossRef] [PubMed]
- Armand, M. Lipases and lipolysis in the human digestive tract: Where do we stand? Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Elisia, I.; Kitts, D.D. Quantification of hexanal as an index of lipid oxidation in human milk and association with antioxidant components. J. Clin. Biochem. Nutr. 2011, 49, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Bernhard, W.; Postle, A.D.; Linck, M.; Sewing, K.-F. Composition of phospholipid classes and phosphatidylcholine molecular species of gastric mucosa and mucus. Biochim. Biophys. Acta 1994, 1255, 99–104. [Google Scholar] [CrossRef]
- Nardone, G.; Laccetti, P.; Civiletti, C.; Budillon, G. Phospholipid composition of human gastric mucosa: A study of endoscopic biopsy specimens. Gut 1993, 34, 456–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids | MOX (g/100 g oil) | HOX (g/100 g oil) |
|---|---|---|
| C 16:0 | 6.70 ± 0.33 a | 7.39 ± 0.19 b |
| C 18:0 | 3.28 ± 0.16 a | 3.39 ± 0.09 a |
| C 18:1 | 11.1 ± 0.54 a | 14.1 ± 0.42 b |
| C 18:2 | 78.1 ± 3.86 a | 74.0 ± 1.92 b |
| C 18:3 | 0.37 ± 0.02 a | 0.90 ± 0.02 b |
| SFA2 | 9.98 ± 0.49 a | 10.8 ± 0.28 b |
| MUFA3 | 11.1 ± 0.54 a | 14.1 ± 0.42 b |
| PUFA | 78.4 ± 3.88 a | 74.9 ± 1.94 b |
| total | 99.6 ± 4.91 a | 99.8 ± 2.63 a |
| peroxide value | MOX (meq O2/kg oil) | HOX (meq O2/kg oil) |
| 18.8 ± 0.39 a | 28.2 ± 0.39 b | |
| hexanal | MOX (µg/mL oil) | HOX (µg/mL oil) |
| 13.6 ± 0.61 a | 10.7 ± 0.54 b | |
| tocopherols | MOX (mg/kg oil) | HOX (mg/kg oil) |
| α-tocopherol | 325 ± 43.2 a | 329 ± 28.5 a |
| γ-tocopherol | 52.2 ± 3.46 a | 80.4 ± 2.12 b |
| δ-tocopherol | <LOQ | <LOQ |
| total | 410 ± 52.7 a | 454 ± 18.8 a |
| MOX-Polyphenols | Catechin (AUC × 104) | Change (%) | Ferulic Acid (AUC × 105) | Change (%) | p-Coumaric Acid (AUC × 106) | Change (%) |
|---|---|---|---|---|---|---|
| (M - H)- | 289.0712 | 193.0501 | 163.0395 | |||
| 0 h | 18.8 ± 2.23 a | 28.3 ± 3.63 a | 16.2 ± 2.25 a | |||
| 2 h | 4.28 ± 0.57 b | −77.3 | 8.41 ± 1.10 b | −70.3 | 4.41 ± 0.86 b | −72.8 |
| 4 h | 4.69 ± 0.63 b | −75.1 | 9.11 ± 0.19 b | −67.8 | 4.90 ± 0.54 b | −69.7 |
| 6 h | 4.48 ± 0.03 b | −76.2 | 9.71 ± 0.47 b | −65.7 | 4.76 ± 0.31 b | −70.6 |
| HOX-polyphenols | Catechin (AUC × 104) | Change (%) | Ferulic acid (AUC × 105) | Change (%) | p-Coumaric acid (AUC × 106) | Change (%) |
| (M - H)- | 289.0712 | 193.0501 | 163.0395 | |||
| 0 h | 21.8 ± 2.56 a | 45.15 ± 4.66 a | 25.0 ± 1.91 a | |||
| 2 h | 3.25 ± 0.41 b | −85.1 | 15.3 ± 0.76 b | −66.0 | 6.91 ± 0.47 b | −72.3 |
| 4 h | 3.97 ± 0.47 b | −81.8 | 17.0 ± 2.55 b | −62.4 | 7.57 ± 0.90 b | −69.7 |
| 6 h | 3.45 ± 0.27 b | −84.2 | 15.6 ± 1.23 b | −65.4 | 7.34 ± 0.66 b | −70.6 |
| MOX-Fatty Acids | 0 h (g/100 g oil) | 6 h (g/100 g oil) | Change (%) | |
|---|---|---|---|---|
| C 16:0 | 6.70 ± 0.33 a | 6.18 ± 0.16 a | −7.76 | |
| C 18:0 | 3.29 ± 0.16 a | 3.21 ± 0.06 a | −2.43 | |
| C 18:1 | 11.1 ± 0.54 a | 10.7 ± 0.14 a | −4.22 | |
| C 18:2 | 78.1 ± 3.86 a | 72.9 ± 0.80 b | −6.66 | |
| C 18:3 | 0.36 ± 0.02 a | 0.37 ± 0.01 a | +2.77 | |
| SFA | 9.98 ± 0.49 a | 10.0 ± 0.14 a | +0.30 | |
| MUFA | 11.1 ± 0.54 a | 10.7 ± 0.14 a | −4.22 | |
| PUFA | 78.4 ± 3.88 a | 73.2 ± 0.80 b | −6.63 | |
| total | 99.6 ± 4.91 a | 93.9 ± 1.07 b | −5.66 | |
| MOX-Tocopherols | 0 h (mg/kg oil) | 2 h (mg/kg oil) | 4 h (mg/kg oil) | 6 h (mg/kg oil) |
| α-tocopherol | 325 ± 43.2 a | 314.3 ± 6.88 a | 321 ± 7.74 a | 318 ± 7.74 a |
| γ-tocopherol | 52.2 ± 3.46 a | 51.8 ± 3.77 a | 53.6 ± 2.44 a | 52.4 ± 1.39 a |
| δ-tocopherol | <LOQ | <LOQ | <LOQ | <LOQ |
| total | 410 ± 52.7 a | 416 ± 16.3 a | 417 ± 7.75 a | 400 ± 17.7 a |
| HOX-Fatty Acids | 0 h (g/100 g oil) | 6 h (g/100 g oil) | Change (%) | |
| C 16:0 | 7.39 ± 0.19 a | 7.42 ± 0.07 a | +0.41 | |
| C 18:0 | 3.39 ± 0.09 a | 3.27 ± 0.03 a | −3.54 | |
| C 18:1 | 14.1 ± 0.42 a | 13.3 ± 0.11 b | −6.02 | |
| C 18:2 | 74.0 ± 1.92 a | 68.6 ± 0.56 b | −7.32 | |
| C 18:3 | 0.90 ± 0.02 a | 0.78 ± 0.01 b | −13.3 | |
| SFA | 10.8 ± 0.28 a | 10.7 ± 0.10 a | −0.93 | |
| MUFA | 14.1 ± 0.42 a | 13.3 ± 0.11 b | −5.67 | |
| PUFA | 74.9 ± 1.94 a | 69.4 ± 0.56 b | −7.34 | |
| total | 99.8 ± 2.63 a | 93.3 ± 0.75 b | −6.49 | |
| HOX-Tocopherols | 0 h (mg/kg oil) | 2 h (mg/kg oil) | 4 h (mg/kg oil) | 6 h (mg/kg oil) |
| α-tocopherol | 329 ± 28.5 a | 330 ± 9.27 a | 330 ± 3.64 a | 338 ± 8.25 a |
| γ-tocopherol | 80.4 ± 2.12 a | 86.3 ± 3.92 a | 84.1 ± 5.82 a | 87.3 ± 7.43 a |
| δ-tocopherol | < LOQ | < LOQ | 93.6 ± 9.18 a | 88.6 ± 12.9 a |
| total | 454 ± 18.8 a | 471 ± 15.3 a | 510 ± 12.2 b | 514 ± 10.9 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fruehwirth, S.; Zehentner, S.; Salim, M.; Sterneder, S.; Tiroch, J.; Lieder, B.; Zehl, M.; Somoza, V.; Pignitter, M. In Vitro Digestion of Grape Seed Oil Inhibits Phospholipid-Regulating Effects of Oxidized Lipids. Biomolecules 2020, 10, 708. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10050708
Fruehwirth S, Zehentner S, Salim M, Sterneder S, Tiroch J, Lieder B, Zehl M, Somoza V, Pignitter M. In Vitro Digestion of Grape Seed Oil Inhibits Phospholipid-Regulating Effects of Oxidized Lipids. Biomolecules. 2020; 10(5):708. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10050708
Chicago/Turabian StyleFruehwirth, Sarah, Sofie Zehentner, Mohammed Salim, Sonja Sterneder, Johanna Tiroch, Barbara Lieder, Martin Zehl, Veronika Somoza, and Marc Pignitter. 2020. "In Vitro Digestion of Grape Seed Oil Inhibits Phospholipid-Regulating Effects of Oxidized Lipids" Biomolecules 10, no. 5: 708. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10050708