Molecular Basis of NDT-Mediated Activation of Nucleoside-Based Prodrugs and Application in Suicide Gene Therapy

 ,
, 
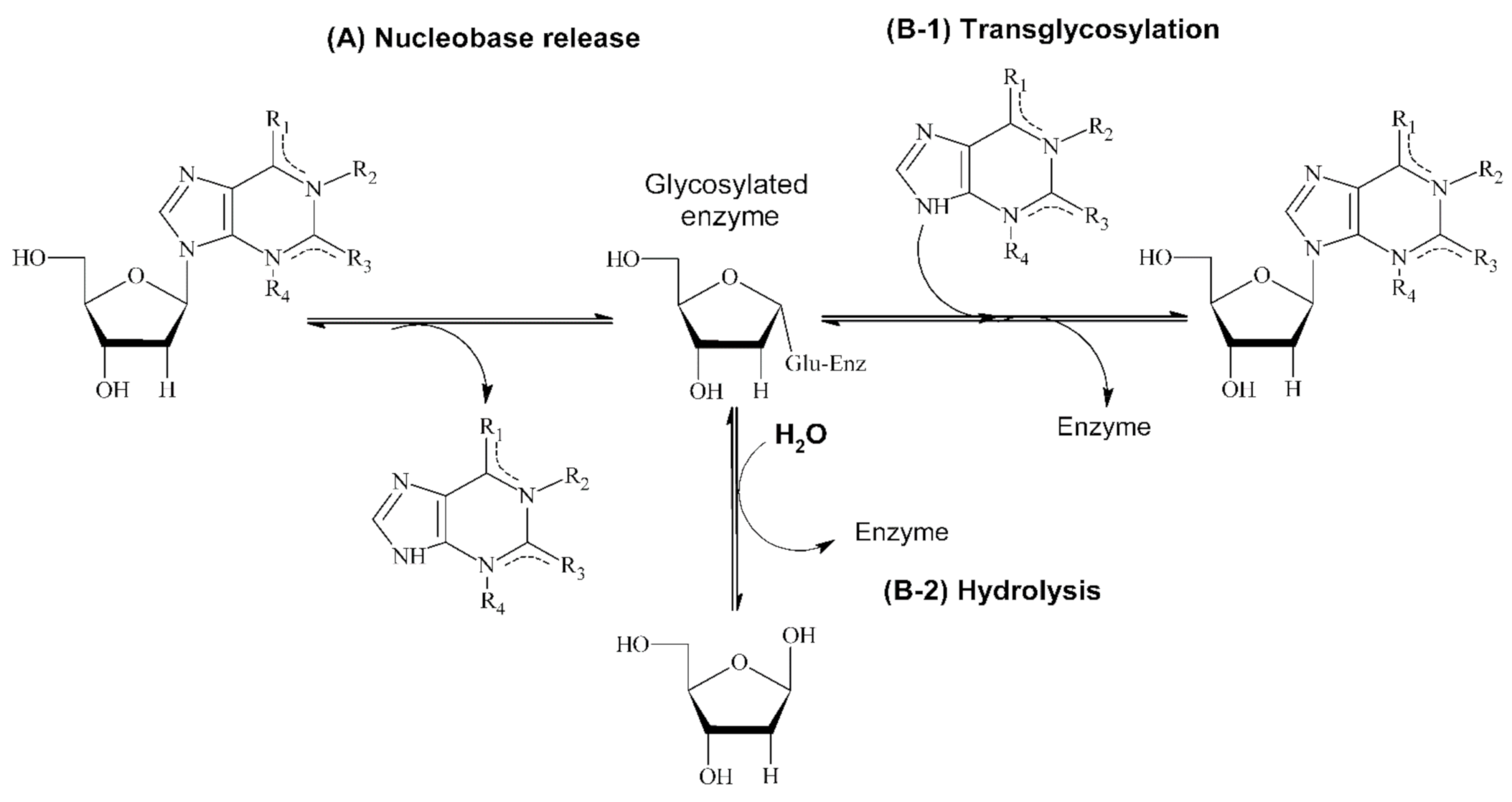{kind=link}
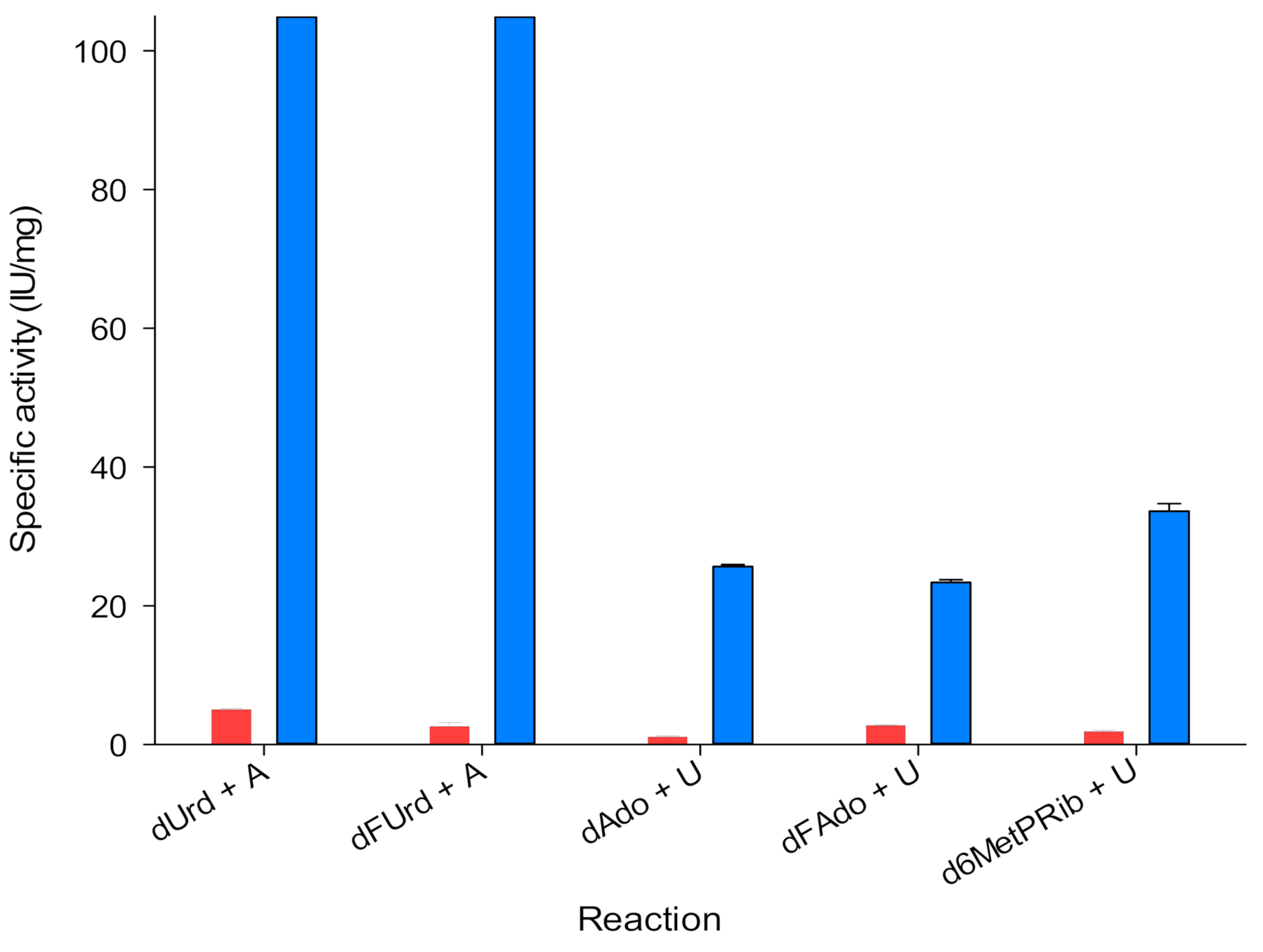{kind=link}
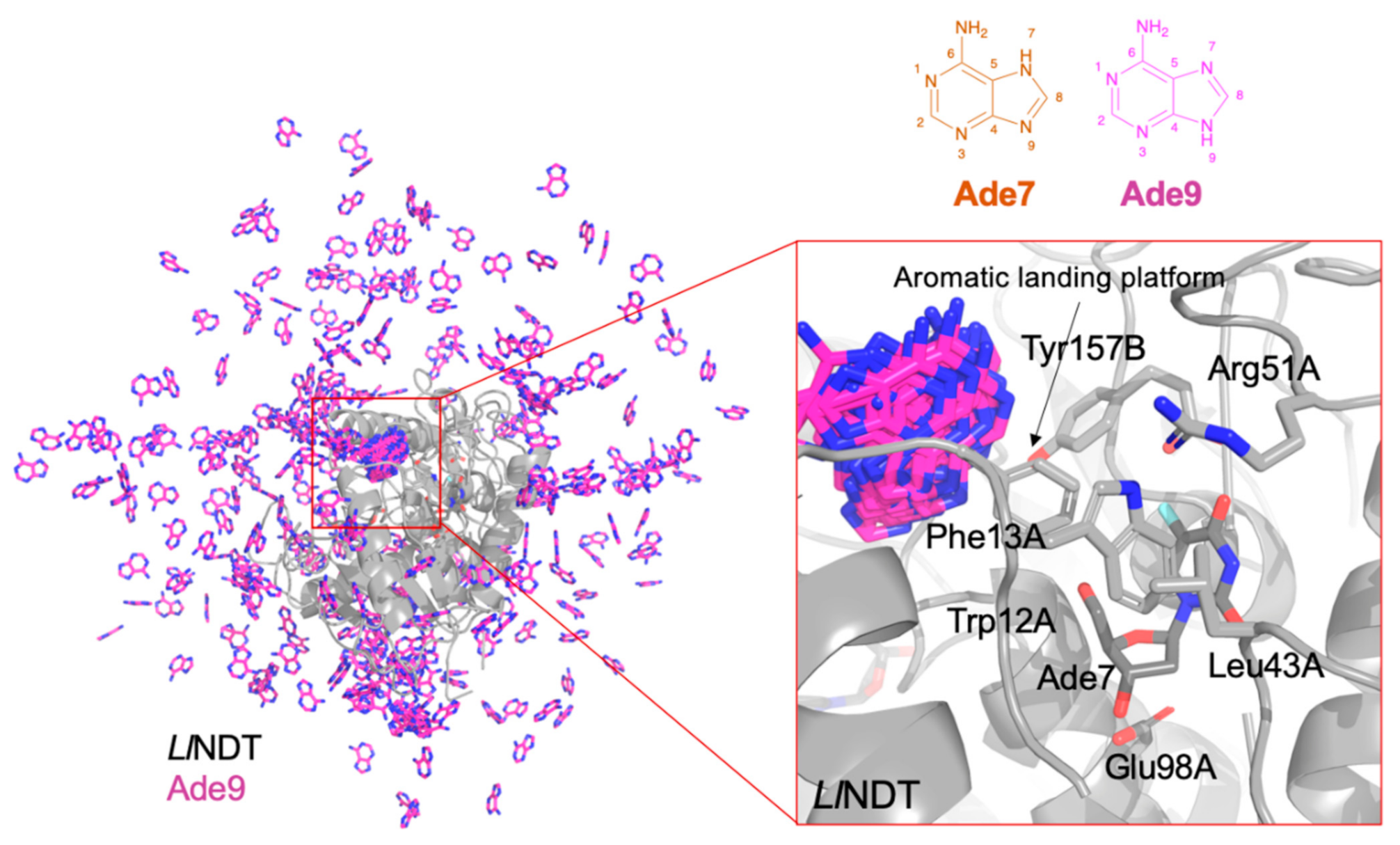{kind=link}
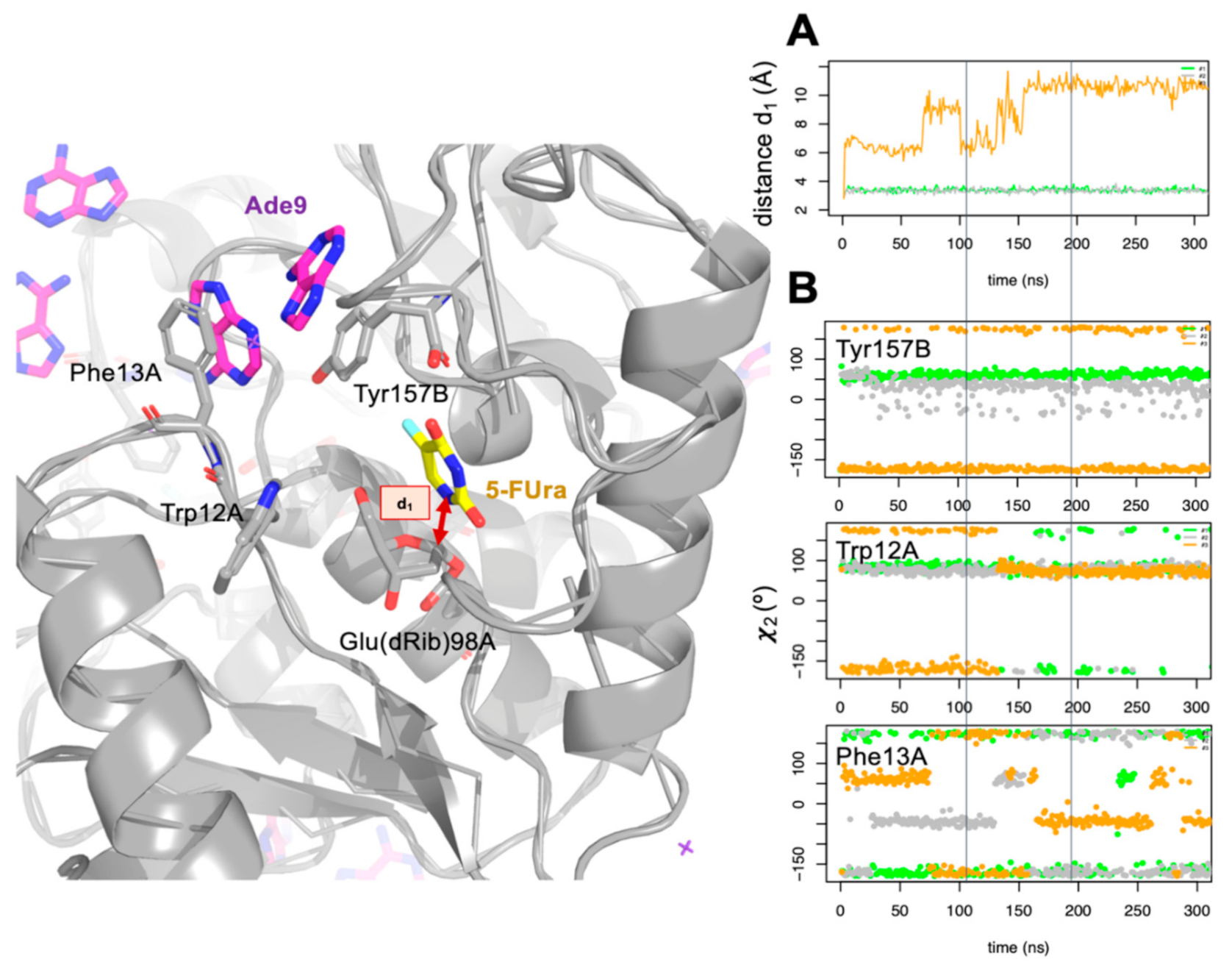{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Enzyme Production and Purification
2.3. Substrate Specificity Studies
2.4. Computational Methods
2.5. Cell Lines and Cell Culture
2.6. Chemical Transfections
2.7. Analytical Methods
3. Results and Discussion
3.1. Characterization of Glycosidase Activity of LdNDT
3.2. Molecular Dynamics: Understanding Nucleobase Release
3.3. Application of NDT/Prodrug Systems in Suicide Gene Therapy
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- WHO. Who Report on Cancer: Setting Priorities, Investing Wisely and Providing Care for All. Available online: https://www.who.int/publications/i/item/who-report-on-cancer-setting-priorities-investing-wisely-and-providing-care-for-all (accessed on 14 January 2021).
- Jordheim, L.P.; Durantel, D.; Zoulim, F.; Dumontet, C. Advances in the Development of Nucleoside and Nucleotide Analogues for Cancer and Viral Diseases. Nat. Rev. Drug Discov. 2013, 12, 447–464. [Google Scholar] [CrossRef] [PubMed]
- Galmarini, C.M.; Mackey, J.R.; Dumontet, C. Nucleoside Analogues and Nucleobases in Cancer Treatment. Lancet Oncol. 2002, 3, 415–424. [Google Scholar] [CrossRef]
- Parker, W.B. Enzymology of Purine and Pyrimidine Antimetabolites Used in the Treatment of Cancer. Chem. Rev. 2009, 109, 2880–2893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Kale, V.; Chen, M. Gene-Directed Enzyme Prodrug Therapy. AAPS J. 2015, 17, 102–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, W.B.; Sorscher, E.J. Use of E. Coli Purine Nucleoside Phosphorylase in the Treatment of Solid Tumors. Curr. Pharm. Des. 2018, 23, 7003–7024. [Google Scholar] [CrossRef]
- Sharma, S.K.; Bagshawe, K.D. Antibody Directed Enzyme Prodrug Therapy (ADEPT): Trials and Tribulations. Adv. Drug Deliv. Rev. 2017, 118, 2–7. [Google Scholar] [CrossRef]
- Nemani, K.V.; Ennis, R.C.; Griswold, K.E.; Gimi, B. Magnetic Nanoparticle Hyperthermia Induced Cytosine Deaminase Expression in Microencapsulated, E. Coli for Enzyme-Prodrug Therapy. J. Biotechnol. 2015, 203, 32–40. [Google Scholar] [CrossRef] [Green Version]
- Ardiani, A.J.; Johnson, A.; Ruan, H.; Sanchez-Bonilla, M.; Serve, K.E.; Black, M. Enzymes to Die For: Exploiting Nucleotide Metabolizing Enzymes for Cancer Gene Therapy. Curr. Gene Ther. 2012, 12, 77–91. [Google Scholar] [CrossRef]
- López-Estévez, S.; Ferrer, G.; Torres-Torronteras, J.; Mansilla, M.J.; Casacuberta-Serra, S.; Martorell, L.; Hirano, M.; Martí, R.; Barquinero, J. Thymidine Phosphorylase Is Both a Therapeutic and a Suicide Gene in a Murine Model of Mitochondrial Neurogastrointestinal Encephalomyopathy. Gene Ther. 2014, 21, 673–681. [Google Scholar] [CrossRef]
- Fresco-Taboada, A.; De La Mata, I.; Arroyo, M.; Fernández-Lucas, J. New Insights on Nucleoside 2′-Deoxyribosyltransferases: A Versatile Biocatalyst for One-Pot One-Step Synthesis of Nucleoside Analogs. Appl. Microbiol. Biotechnol. 2013, 97, 3773–3785. [Google Scholar] [CrossRef]
- Crespo, N.; Sánchez-Murcia, P.A.; Gago, F.; Cejudo-Sanches, J.; Galmes, M.A.; Fernández-Lucas, J.; Mancheño, J.M. 2′-Deoxyribosyltransferase from Leishmania Mexicana, an Efficient Biocatalyst for One-Pot, One-Step Synthesis of Nucleosides from Poorly Soluble Purine Bases. Appl. Microbiol. Biotechnol. 2017, 101, 7187–7200. [Google Scholar] [CrossRef] [Green Version]
- Pérez, E.; Sánchez-Murcia, P.A.; Jordaan, J.; Blanco, M.D.; Mancheño, J.M.; Gago, F.; Fernández-Lucas, J. Enzymatic Synthesis of Therapeutic Nucleosides Using a Highly Versatile Purine Nucleoside 2′-Deoxyribosyl Transferase from Trypanosoma Brucei. ChemCatChem 2018, 10, 4406–4416. [Google Scholar] [CrossRef]
- Del Arco, J.; Perona, A.; González, L.; Fernández-Lucas, J.; Gago, F.; Sánchez-Murcia, P.A. Reaction Mechanism of Nucleoside 2′-Deoxyribosyltransferases: Free-Energy Landscape Supports an Oxocarbenium Ion as the Reaction Intermediate. Org. Biomol. Chem. 2019, 17, 7891–7899. [Google Scholar] [CrossRef]
- Smar, M.; Short, S.A.; Wolfenden, R. Lyase Activity of Nucleoside 2-Deoxyribosyltransferase: Transient Generation of Ribal and Its Use in the Synthesis of 2′-Deoxynucleosides. Biochemistry 1991, 30, 7908–7912. [Google Scholar] [CrossRef]
- Del Arco, J.; Mills, A.; Gago, F.; Fernández-Lucas, J. Structure-Guided Tuning of a Selectivity Switch towards Ribonucleosides in Trypanosoma Brucei Purine Nucleoside 2′-Deoxyribosyltransferase. ChemBioChem 2019, 20, 2996–3000. [Google Scholar] [CrossRef]
- Kaminski, P.A.; Dacher, P.; Dugué, L.; Pochet, S. In Vivo Reshaping the Catalytic Site of Nucleoside 2′-Deoxyribosyltransferase for Dideoxy- and Didehydronucleosides via a Single Amino Acid Substitution. J. Biol. Chem. 2008, 283, 20053–20059. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Lucas, J.; Acebal, C.; Sinisterra, J.V.; Arroyo, M.; De La Mata, I. Lactobacillus Reuteri 2′-Deoxyribosyltransferase, a Novel Biocatalyst for Tailoring of Nucleosides. Appl. Environ. Microbiol. 2010, 76, 1462–1470. [Google Scholar] [CrossRef] [Green Version]
- Fresco-Taboada, A.; Fernández-Lucas, J.; Acebal, C.; Arroyo, M.; Ramón, F.; De La Mata, I.; Mancheño, J.M. 2′-Deoxyribosyltransferase from Bacillus Psychrosaccharolyticus: A Mesophilic-like Biocatalyst for the Synthesis of Modified Nucleosides from a Psychrotolerant Bacterium. Catalysts 2018, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Vichier-Guerre, S.; Dugué, L.; Bonhomme, F.; Pochet, S. Expedient and Generic Synthesis of Imidazole Nucleosides by Enzymatic Transglycosylation. Org. Biomol. Chem. 2016, 14, 3638–3653. [Google Scholar] [CrossRef]
- Lapponi, M.J.; Rivero, C.W.; Zinni, M.A.; Britos, C.N.; Trelles, J.A. New Developments in Nucleoside Analogues Biosynthesis: A Review. J. Mol. Catal B Enzym. 2016, 133, 218–233. [Google Scholar] [CrossRef]
- Acosta, J.; Del Arco, J.; Martinez-Pascual, S.; Clemente-Suárez, V.J.; Fernández-Lucas, J. One-Pot Multi-Enzymatic Production of Purine Derivatives with Application in Pharmaceutical and Food Industry. Catalysts 2018, 8, 9. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, K.A.; Jewett, M.W.; Rosa, P.A.; Gherardini, F.C. Borrelia Burgdorferi Bb0426 Encodes a 2′-Deoxyribosyltransferase That Plays a Central Role in Purine Salvage. Mol. Microbiol. 2009, 72, 1517–1529. [Google Scholar] [CrossRef] [Green Version]
- Bosch, J.; Robien, M.A.; Mehlin, C.; Boni, E.; Riechers, A.; Buckner, F.S.; Van Voorhis, W.C.; Myler, P.J.; Worthey, E.A.; DeTitta, G.; et al. Using Fragment Cocktail Crystallography to Assist Inhibitor Design of Trypanosoma Brucei Nucleoside 2-Deoxyribosyltransferase. J. Med. Chem. 2006, 49, 5939–5946. [Google Scholar] [CrossRef]
- Armstrong, S.R.; Cook, W.J.; Short, S.A.; Ealick, S.E. Crystal Structures of Nucleoside 2-Deoxyribosyltransferase in Native and Ligand-Bound Forms Reveal Architecture of the Active Site. Structure 1996, 4, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Anandakrishnan, R.; Aguilar, B.; Onufriev, A.V. H++ 3.0: Automating PK Prediction and the Preparation of Biomolecular Structures for Atomistic Molecular Modeling and Simulations. Nucleic Acids Res. 2012, 40, w537–w541. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of Simple Potential Functions for Simulating Liquid Water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Postma, J.P.M.; Van Gunsteren, W.F.; Dinola, A.; Haak, J.R. Molecular Dynamics with Coupling to an External Bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef] [Green Version]
- Darden, T.; York, D.; Pedersen, L. Particle Mesh Ewald: An N·log(N) Method for Ewald Sums in Large Systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef] [Green Version]
- Ausubel, F.M.; Brent, R.; Kingston, R.E.; Moore, D.D.; Seidman, J.G.; Smith, J.A.; Struhl, K. Current Protocols in Molecular Biology; John Wiley: New York, NY, USA, 1988; pp. 431–433. [Google Scholar] [CrossRef]
- Graham, F.L.; Van der Eb, A.J. A New Technique for the Assay of Infectivity of Human Adenovirus 5 DNA. Virology 1973, 52, 456–469. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Bennett, E.M.; Anand, R.; Allan, P.W.; Hassan, A.E.A.; Hong, J.S.; Levasseur, D.N.; McPherson, D.T.; Parker, W.B.; Secrist III, J.A.; Sorscher, E.J.; et al. Designer Gene Therapy Using an Escherichia Coli Purine Nucleoside Phosphorylase/Prodrug System. Chem. Biol. 2003, 10, 1173–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, W.J.; Short, S.A.; Ealick, S.E. Crystallization and Preliminary X-Ray Investigations of Recombinant Lactobacillus Leichmannii Nucleoside Deoxyribosyltransferase. J. Biol. Chem. 1990, 265, 2682–2683. [Google Scholar] [CrossRef]
- Short, S.A.; Armstrong, S.R.; Ealick, S.E.; Porter, D.J.T. Active Site Amino Acids That Participate in the Catalytic Mechanism of Nucleoside 2′-Deoxyribosyltransferase. J. Biol. Chem. 1996, 271, 4978–4987. [Google Scholar] [CrossRef] [Green Version]
- Parker, W.B.; Allan, P.W.; Hassan, A.E.A.; Secrist, J.A.; Sorscher, E.J.; Waud, W.R. Antitumor Activity of 2-Fluoro-2′-Deoxyadenosine against Tumors That Express Escherichia Coli Purine Nucleoside Phosphorylase. Cancer Gene Ther. 2003, 10, 23–29. [Google Scholar] [CrossRef]
- Silamkoti, A.V.; Allan, P.W.; Hassan, A.E.A.; Fowler, A.T.; Sorscher, E.J.; Parker, W.B.; Secrist, J.A. Synthesis and Biological Activity of 2-Fluoro Adenine and 6-Methyl Purine Nucleoside Analogs as Prodrugs for Suicide Gene Therapy of Cancer. Nucleosides Nucleotides Nucleic Acids. 2005, 24, 881–885. [Google Scholar] [CrossRef]
- Behbahani, T.E.; Rosenthal, E.L.; Parker, W.B.; Sorscher, E.J. Intratumoral Generation of 2-Fluoroadenine to Treat Solid Malignancies of the Head and Neck. Head Neck 2019, 41, 1979–1983. [Google Scholar] [CrossRef]
- Rosenthal, E.L.; Chung, T.K.; Parker, W.B.; Allan, P.W.; Clemons, L.; Lowman, D.; Hong, J.; Hunt, F.R.; Richman, J.; Conry, R.M.; et al. Phase I Dose-Escalating Trial of Escherichia Coli Purine Nucleoside Phosphorylase and Fludarabine Gene Therapy for Advanced Solid Tumors. Ann. Oncol. 2015, 26, 1481–1487. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acosta, J.; Pérez, E.; Sánchez-Murcia, P.A.; Fillat, C.; Fernández-Lucas, J. Molecular Basis of NDT-Mediated Activation of Nucleoside-Based Prodrugs and Application in Suicide Gene Therapy. Biomolecules 2021, 11, 120. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11010120
Acosta J, Pérez E, Sánchez-Murcia PA, Fillat C, Fernández-Lucas J. Molecular Basis of NDT-Mediated Activation of Nucleoside-Based Prodrugs and Application in Suicide Gene Therapy. Biomolecules. 2021; 11(1):120. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11010120
Chicago/Turabian StyleAcosta, Javier, Elena Pérez, Pedro A. Sánchez-Murcia, Cristina Fillat, and Jesús Fernández-Lucas. 2021. "Molecular Basis of NDT-Mediated Activation of Nucleoside-Based Prodrugs and Application in Suicide Gene Therapy" Biomolecules 11, no. 1: 120. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11010120