3.1.1. ‘Full’ Guest Encapsulation: TMB·HFA, TMB.PCA, and TMA·PCA
The term ‘full encapsulation’ is used here to signify that in these complexes, the respective guest molecules generally maintain planar conformations and span the entire length of the CD cavity, possibly with only slight protrusion of either or both of their terminal groups (phenolic/carboxylic acid) from the secondary or primary rims of the CD. Results for TMB·HFA and TMB.PCA, which happen to display some degree of isostructurality, are presented first. Initial examination of single crystals of TMB·HFA using HSM (
Figure 1) showed the development of fine parallel cracks and the release of small bubbles, indicative of dehydration. Melting commenced at 140 °C.
Figure 2 shows the TGA and DSC curves for TMB·HFA and the derived data are listed in
Table 2. The small water loss (1.4 ± 0.2%) over the 25–100 °C range in the TGA curve corresponded to 1.3 ± 0.2 H
2O molecules per 1:1 host-guest complex unit. Water loss is evident in the DSC curve as a broad endotherm in the same temperature range. The sharp endotherm peaking at 136 °C corresponded to the melting observed by HSM (
Figure 1) and was followed by a broad endotherm at ~230 °C, which signals a one-step mass loss in the TGA curve (T
on ~175 °C), representing loss of the guest hydroferulic acid. The measured mass loss of 12.0 ± 0.2% equates to a host-guest (H:G) stoichiometry of 1:(1.0 ± 0.1).
The complex TMB.PCA has a similar thermal profile to that of TMB·HFA except that, with a higher complement of water in the crystal (
Table 2), the dehydration process involves two overlapping endotherms followed by a small exotherm (
Figure S2, Supplementary Materials), attributed to a phase change of the anhydrous complex. This is in accord with the HSM observation (
Figure S3, Supplementary Materials), which indicates crystal fracture and the onset of opacity. This is followed by melting of the new phase at 179 °C and subsequent one-step loss of the guest
p-coumaric acid. As indicated in
Table 2, the complex also has a 1:1 H:G stoichiometry.
Single crystal X-ray diffraction (SCXRD) indicated that the complexes TMB·HFA and TMB·PCA crystallized in the orthorhombic space group
P2
12
12
1 with similar unit cell dimensions, indicating possible isostructurality of their host frameworks (a common occurrence for CD complexes [
14]). With
Z = 4, their modeled asymmetric units (ASUs) had the respective formulae TMB·HFA(1.3 H
2O) and TMB·PCA(7.0 H
2O). Crystal data and details of data-collection and refinement for these complexes are listed in
Table S1 (Supplementary Materials).
Figure 3 shows the structures, conformations, and atomic numbering for the complexed guest molecules. Non-planar conformations for these molecules are evident, the plane of the carboxylic acid group deviating somewhat from that of the phenyl group, as indicated by the respective torsion angles C7-C8-C9-O4 of −23(1)° for HFA and C3-C4-C7-C8 of 15(2)° for PCA.
In these 1:1 host-guest complexes, the common feature of particular interest is the mode of guest inclusion (
Figure 4), namely effectively complete encapsulation of both the HFA and PCA molecules within their respective host cavities, with their carboxylic acid group located near the narrow primary rim of the host (top) and the phenolic groups near the wider secondary rim (bottom).
Geometrical parameters defining the detailed conformations of cyclodextrins are well established [
15] and are based on the deviations from regularity of the polygon comprising the glycosidic oxygen atoms (O4) as well as the variations in the angles of tilt of glucose residues relative to the mean plane of the O4-polygon. For the TMB molecule, containing seven glucose rings, the O4-heptagon generally deviates significantly from planarity due to the steric clashes of methyl groups on the secondary rim, which also cause the individual glucose rings to adopt a wide range of tilt angles. For the TMB molecules shown in
Figure 4, the definitions of the geometrical parameters and their observed values are listed in
Tables S2 and S3 (Supplementary Materials). The view directions in
Figure 4 are approximately normal to the respective guest phenyl rings, located in the center of the CD molecule cavities. This common molecular orientation results in an elliptical distortion of each host molecule in the horizontal direction, as a result of the mutual induced fit of host and guest accompanying the inclusion process [
16]. The ellipticity of each TMB host molecule is evident from the fact that the values of the parameter
r (the distance of each O4 atom to the centroid of the O4-heptagon) are not equal, but span relatively wide ranges, namely 4.32–5.50 Å for TMB·HFA and 4.41–5.51 Å for TMB·PCA. The non-planarity of the O4-heptagon is evident from the ranges of values of
α (the positive or negative distance of each O4 atom from the mean (least-squares) plane through the seven O4 atoms), namely −0.529(3)–0.624(3) Å and −0.668(2)–0.517(3) Å for TMB·HFA and TMB·PCA, respectively. This puckering of the O4-heptagon is accompanied by wide ranges of the tilt angles (
τ2) for the seven permethylated glucose rings,
viz. 2.5(1)–39.0(1)° (TMB·HFA) and 8.3(1)–36.9(1)° (TMB·PCA).
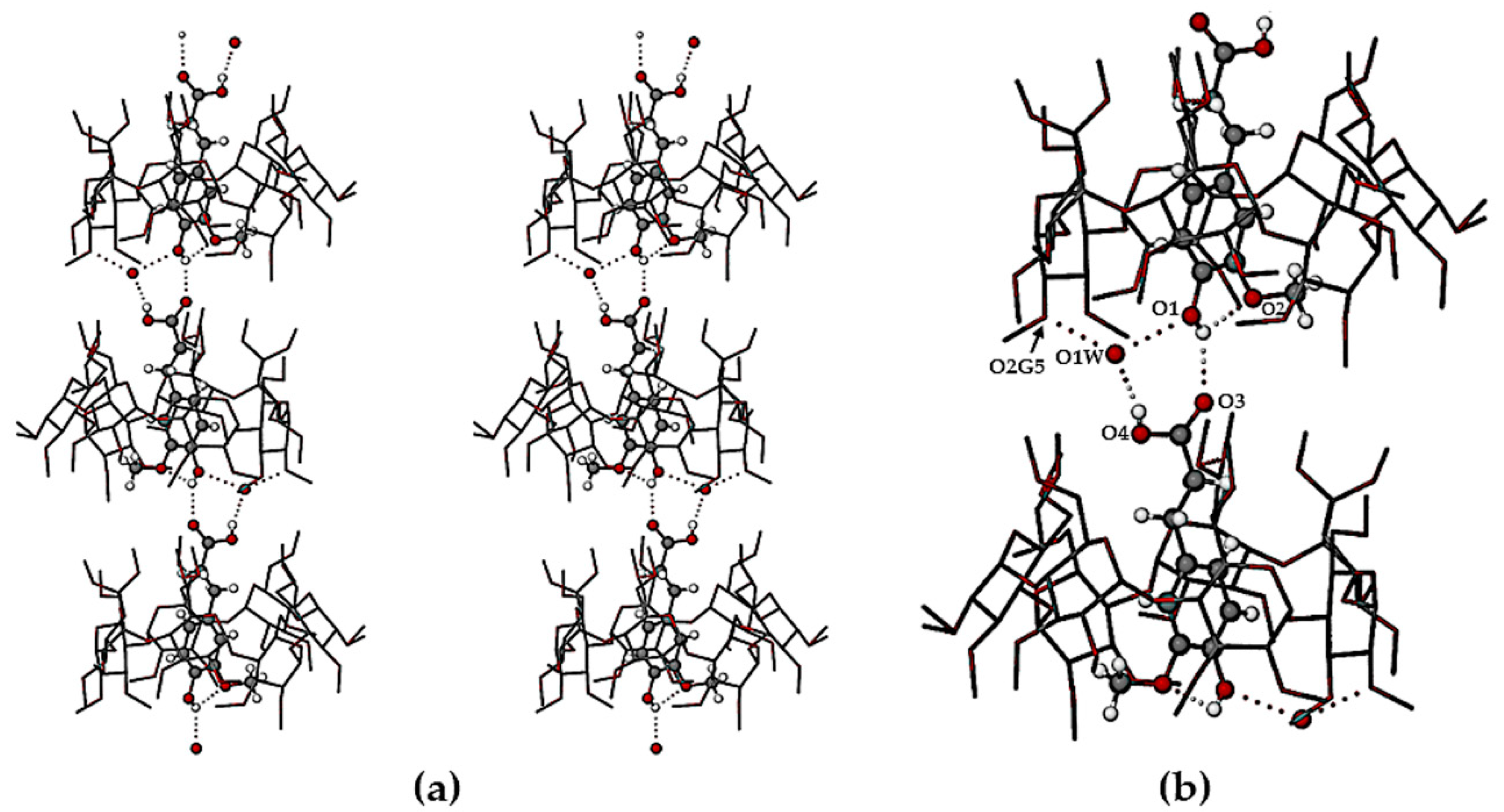
The crystal structures of TMB·HFA and TMB·PCA are formed by the close packing of infinite columns of complex units. A representative portion of the column in the TMB·HFA crystal is shown in
Figure 5 and an analogous figure for the TMB·PCA crystal is shown in the
Supplementary Materials (Figure S4). In these crystals, the complex units comprising the column are related by the twofold screw axis parallel to the crystal
b-axis. The resulting head-to-tail complex stacking arrangement (
Figure 5a) is stabilized by a series of hydrogen bonds, details of which appear in the magnified image (
Figure 5b). The principal intermolecular H-bonds in TMB·HFA include a direct guest–guest phenolic(O1-H)···O3(carbonyl) H-bond with O···O 2.753(6) Å and three additional hydrogen bonds mediated by the disordered water molecule O1W, namely carboxylic acid(O4-H)···O1W with O···O 2.692(7) Å, and two inferred H-bonds with the water molecule acting as donor to (a) atom O1 of the phenolic group (O···O 2.696(7) Å) and to (b) a secondary hydroxyl group O atom of the host molecule, O2G5 (O···O 2.793(6) Å). Evident also in
Figure 5b is the intramolecular H-bond in the HFA molecule O1-H···O2(methoxyl) with O···O 2.657(9) Å. Analogous head-to-tail guest inclusion of PCA molecules occurred in the crystal of TMB·PCA (
Figure S4, Supplementary Materials), but there was no direct H-bonding between successive guest molecules. Instead, the –COOH group of one PCA molecule was linked to the phenolic group of the neighboring PCA molecule via a bridging water molecule. The conformations of the TMB molecules in both TMB·HFA and TMB·PCA were also influenced by numerous intramolecular C–H···O hydrogen bonds. Intermolecular host-host and host-water C–H···O hydrogen bonds in both complexes contribute to their crystal stability. The higher water content in the TMB·PCA complex (7H
2O per complex unit vs. 1.3H
2O in the crystal of TMB·HFA) led to a more extensive network of H-bonds.
PXRD patterns for TMB·HFA and TMB·PCA are shown in
Figure S5 (Supplementary Material). For each complex, there was good agreement between the experimental pattern and the simulated pattern based on the single crystal X-ray data. In addition, the patterns of the two complexes showed similarities in a few major peak positions due to some degree of isostructurality referred to above.
The main features of the thermal analysis results for the crystals of TMA·PCA (the third complex displaying full guest inclusion) were similar to those reported for the previous complexes, namely initial dehydration, melting of the anhydrous phase, and loss of the guest. Salient quantitative thermal data for this complex phase appear in
Table 2. The TGA and DSC traces as well as HSM micrographs are shown in
Figure S6 (Supplementary Materials) and the crystallographic data for the 1:1 TMA·PCA complex are listed in
Table S4 (Supplementary Materials). In this complex, the guest molecule is disordered over two positions (
Figure S7, Supplementary Materials), the respective phenyl rings intersecting at an angle of 34.3(5)° and the major component having a site-occupancy factor (s.o.f.) of 0.59(1). The respective disordered –COOH group components were located near the host primary rim, while only the phenolic groups protruded from the secondary side, this inclusion mode (
Figure 6) being analogous to those described above for the TMB·HFA and TMB·PCA structures. For clarity, only the major component of the disordered guest is included in
Figure 6.
The ASU of TMA·PCA contains one complex unit plus 4.5 water molecules, the latter being disordered over eight sites forming a hydrogen bonded network (
Figure 7a) that includes methoxyl oxygen atom acceptors (generic labels OmGn) of neighboring host molecules. The network is concentrated in the region above the exposed guest –COOH group, to which it is linked via the H-bond –COOH···O4W. Detailed geometrical parameter data for the host conformation (
Table S5, Supplementary Materials) indicate some distortion of the O4-hexagon (parameter
r range 4.06–4.39 Å, maximum deviation
α of 0.192(3) Å from the mean O4-plane,
τ2 range 5.7(1)–14.4(1)°). In contrast to the previous complexes that associate in infinite columns, the complex units of TMA·PCA form layers parallel to the
ac-plane with sequence ABAB…, representative complex molecules in three layers are shown in
Figure 7b. The representative complex unit in layer B
1 is sandwiched between three complex units in layer A
1 and three identical complex units in layer A
2. This packing mode is similar to that occurring in α-CD cage-type structures [
17]. Water molecules are located at the A
1/B
1 junction, while the complex units in layers B
1 and A
2 are in somewhat closer contact with one another.
The experimental and computed PXRD traces were in good agreement (
Figure S8, Supplementary Materials), small differences being due to the different temperatures of intensity data capture for SCXRD and PXRD methods.
3.1.2. Partial Guest Inclusion: DMB·HFA, DMB.PCA, and TMA·FA
In this series of complexes, each guest molecule had one significant residue located within the CD cavity and a significant residue protruding from either the primary or secondary rim of the cavity into the interstitial space. The two DMB complexes showed very similar host packing arrangements and their thermal behaviors are described first.
The successive events on heating a crystal of DMB·HFA immersed in silicone oil are shown in
Figure 8. Bubbles initially appear due to dehydration and the crystal becomes opaque on further heating to 130 °C, evidently signifying a phase change (see DSC and TGA behavior in
Figure 9). The opacity was reduced at ~190 °C, at which temperature guest loss from the complex crystal commences. A color change observed at ~290 °C indicates decomposition of the host compound.
The assigned dehydration step is seen as a small mass loss in the range 25–100 °C in the TGA curve and as a broad endotherm in DSC (
Figure 9). The second endotherm peaking at 134 °C corresponded to the phase change evident in HSM, while the third endotherm, commencing at ~180 °C coincided with the first step of guest release; this was followed by a broad endotherm over the range 225 °C to 325 °C that accompanied the second release.
Figures showing the HSM, TGA, and DSC results for the structurally related complex DMB·PCA appear in the
Supplementary Materials (Figure S9). The thermal profiles displayed analogous events to those for DMB·HFA except that the enthalpy values for the corresponding endotherms were significantly smaller.
Table 2 lists the quantitative data for DMB·HFA and DMB.PCA derived from their thermal analyses. Both complexes have 1:1 CD:guest stoichiometry and neither displayed melting on HSM.
For the complex TMA·FA (
Figure S10, Supplementary Materials;
Table 2,) dehydration occurred between 30 and 150 °C. A small endotherm indicates a phase change at 160 °C, which was followed by a sharp endotherm at 185 °C corresponding to fusion of the complex, the latter also being evident in the HSM record. The TGA curve showed complete guest loss in the range 175–300 °C, corresponding to a 1:1 host-guest stoichiometry.
The DMB·HFA and DMB·PCA complexes crystallized in the orthorhombic space group
P2
12
12
1 with similar
a and
b unit cell dimensions, but with their
c-axes differing by ~1.6 Å, suggesting some level of host isostructurality. Their structures were solved by direct methods as attempts to use host atomic co-ordinates from known DMB complexes as trial structures for solution via isomorphous replacement failed. Crystallographic details are listed in
Table S6 in the Supplementary Materials.
The structures of the HFA and PCA molecules in their complexed state with the host DMB are shown in
Figure 10.
When compared with their counterparts in the complexed state with the TMB molecule (
Figure 4 above), it is evident that the HFA molecule adopted a significantly different conformation when it is included in the host DMB (
Figure 10a), while the PCA molecule adopted similar conformations when included in both DMB and TMB. For HFA, the torsion angle C3-C4-C7-C8 was −102.5(5)° upon inclusion in DMB and 179.8(8)° when included in TMB. The respective torsion angles in the case of the guest PCA had values of −14.7(1)° and 15(2)°). As shown in
Figure 11, both guest molecules were partially included within their respective DMB cavities, but with the larger volume of the HFA molecule protruding from the primary side of the host molecule, rotation of the plane of the carboxylic acid group relative to the phenyl ring was facilitated, enabling the –COOH group to achieve optimum interactions with molecules in the extra-cavity environment. These features are also evident from
Figure S11 in the Supplementary Materials, both hosts and guests being drawn in space-filling mode.
Generally, significant differences in the modes of inclusion of a given guest in DMB and TMB may arise from the radically different cavity topologies of these host molecules, the DMB macrocycle typically maintaining a round or slightly elliptical shape due to the belt of seven O2n···H-O3(n-1) hydrogen bonds on its secondary (wider) rim and relatively uniform inclinations of the seven glucose rings (these key features being evident in
Figure 11 above), while they are lacking in the TMB molecule (
Figure 4). However,
Figure 11 above, showing that both the bulk of the HFA molecule and a significant residue of the PCA molecule protrude from their respective host cavities, are deceptive, as the major factor determining their locations within the cavity is instead self-inclusion of the DMB molecules. This is shown in
Figure 12, where, for each complex, a portion of one DMB molecule (red) is inserted laterally into the secondary side of the DMB molecule above it (green), thereby limiting the cavity occupation by the guest molecules (blue). Quantitative measures of the extents of self-inclusion are discussed at the end of
Section 3.1.3.
The exposed –COOH group and the phenolic group of the HFA guest molecule are hydrogen bonded to distinct interstitial water clusters (
Figure S12, left, Supplementary Materials) which are enclosed by neighboring DMB molecules (the latter not shown for clarity). Instead, while the –COOH group of the PCA molecule is likewise hydrogen bonded to an interstitial water cluster, its phenolic group is a donor of a H-bond to oxygen atom O6G2 of the DMB molecule, which partly inserts into the parent host molecule (
Figure S12, right, Supplementary Materials). The geometrical parameters for the DMB host molecules in each of these complexes (
Tables S7 and S8 in Supplementary Materials) show typical variations indicating their slight ellipticity, the extents of which are guest-dependent. The tilt angle (
τ2) ranges of the glucose residues are somewhat wider for the DMB complex containing the sterically more bulky HFA molecule. Experimental and calculated PXRD traces for the DMB·HFA and DMB·PCA complexes are shown in
Figure S13 (Supplementary Materials).
X-ray analysis of the complex TMA.FA, crystallizing in the monoclinic space group
P2
1 with Z = 2 (
Table S9, Supplementary Materials), likewise revealed partial guest inclusion. In this case, the FA molecule adopts a planar conformation and is partially included in the host cavity from the secondary side (
Figure 13), while the host methoxyl groups on the primary side effectively close the cavity.
The guest molecule is tethered to the host via a hydrogen bond O–H···O (
Figure 13) between the included phenolic group and the host glycosidic oxygen atom O4G4 (O····O = 3.031(3) Å) while the carboxylic acid group protrudes from the host secondary rim. This mode of inclusion of FA in TMA contrasts strongly with that of the planar guest molecule PCA in the complex TMA·PCA (
Figure 6), which features ‘full’ guest encapsulation. The differences in the conformations of the host TMA molecules in these two complexes, evident from a comparison of the data in
Tables S5 (TMA·PCA) and S10 (TMA·FA) (Supplementary Materials) are very significant as a result of both the different modes of guest inclusion as well as a slightly greater steric bulk of the FA molecule, reflecting the flexibility of the common TMA molecule in guest accommodation. In particular, one notes that full encapsulation of PCA is associated with a narrow range of the TMA tilt angles
τ2, namely 3.5–14.4°, while for partial inclusion of FA in TMA, the
τ2 range is 7.7–41.4°.
While partial guest inclusion in the complexes DMB·HFA and DMB·PCA was attributed above to a significant level of host molecule interpenetration (‘self-inclusion’), the latter feature does not apply to the TMA·FA complex. Here, the closure of the primary side of the host TMA molecule by the methoxyl groups and the location of the bulk of the guest molecule within the host cavity leads to stacking of the discrete TMA·FA complex units into infinite columns, a representative portion of a column being shown in the stereodiagram (
Figure 14a). The crystal structure is based on anti-parallel columns of complex units that are bound together via the supramolecular heterosynthon shown in
Figure 14b. This heterosynthon, with graph-set notation
[
18], comprises two hydrogen bonds that link the carboxylic acid group of a guest FA molecule (at
x, y, z) of one complex column and a O–C–C–H unit of a methylglucose residue (G5) of a TMA molecule (at the symmetry-related position (a) = –x, –1/2 + y, –z) in the neighboring complex column. Details of these interactions are as follows: (1) O4-H···O3G5
a with O···O 2.748(3) Å, H···O 1.92 Å, bond angle 170°, and (2) C4G5
a -H···O3 with C···O 3.300(4) Å, H···O 2.40 Å, bond angle 149°. These strong intercolumnar links stabilize the complex crystal structure. Additional intercolumnar stabilization is provided by a water molecule (H-O1W-H) that links TMA molecules of neighboring columns via two O–H···O hydrogen bonds. Experimental and calculated PXRD traces for the complex were in good agreement (
Figure S14, Supplementary Materials).
3.1.3. ‘Non-Inclusion’ Complexes: DMB·FA and DMB·CAF
HSM micrographs for the representative complex DMB·FA are shown in
Figure 15. Upon heating the initially clear crystal, bubbles accumulate, indicating continued mass loss and the crystal eventually becomes opaque. Discoloration occurs at 280 °C and subsequent vigorous bubble formation indicates final decomposition of the residual material. For the second complex in this category, DMB·CAF, heating results in crystal fracture and at 160 °C, the crystal becomes opaque (
Figure S15b, Supplementary Materials). Discoloration from 220 °C indicates decomposition. Again, limited information was available from HSM, but these data complemented the quantitative results from TGA and DSC.
Figure 16 shows the combined TGA and DSC traces for the complex DMB·FA while those for DMB·CAF are shown in
Figure S15a (Supplementary Materials). Data derived from these traces are listed in
Table 2. As with all the previously described thermal profiles, the major events are initial dehydration and guest loss at a significantly higher temperature. For DMB·FA, the initial dehydration mass loss was associated with a broad endotherm spanning the range ~25–120 °C in DSC. A low enthalpy, broad endotherm with onset at 160 °C signals commencement of the guest loss, which is followed by decomposition of the host from ~360 °C. For DMB·CAF (
Figure S15b, Supplementary Materials), a small exotherm at 160 °C indicates a possible phase transition of the anhydrous complex, the latter undergoing guest loss from ~210 °C. For both DMB·FA and DMB·CAF, the guest mass losses corresponded to a DMB·guest stoichiometric ratio of 2:1, in contrast to the 1:1 host-guest stoichiometry determined for the previous six complexes. While the thermal analysis results for all eight complexes tended to follow a similar overall profile, with water and guest loss as the primary features, it turned out that the crystal structures of DMB·FA and DMB·CAF differed very significantly from those previously described in this report: as revealed in the next section, they exemplify the extreme situation of ‘non-inclusion’ of the guest molecules in the respective host cavities, a rare feature in the solid-state chemistry of CD complexes.
Single crystal X-ray diffraction indicated very similar unit cell dimensions and a common space group (
P2
1) for DMB·FA and DMB·CAF (
Table S11, Supplementary Materials), which strongly suggest isostructurality of the host arrangements [
14]. Following structural solution of DMB·CAF using phasing from program SHELXD [
12], the atomic co-ordinates of the two crystallographically independent host molecules were accordingly used as the trial model for successful structural solution of DMB·FA via isomorphous replacement, the guest molecule and water oxygen atoms being subsequently located in difference Fourier maps.
In the crystal of DMB·CAF, the guest molecule is disordered over two positions, the major component having a site-occupancy factor (s.o.f.) of 0.55(1). Minor disorder of the host molecules in these complexes was also modeled appropriately and oxygen atoms of the water molecules were included in the structural models. Each of the asymmetric units (ASUs) in the two complexes (
Figure 17) comprised two DMB molecules, one guest molecule and several water molecules (modeled as 11.2 water oxygen atoms occupying 14 sites in DMB·FA, and 11.9 water O atoms occupying 16 sites in DMB·CAF). The predicted isostructural arrangement of the host molecules in the two ASUs was quite evident and some level of isostructurality was also seen to extend to the respective guests and water molecules. As explained in the detailed discussion below, the proper classification of DMB·FA and DMB·CAF as ‘non-inclusion complexes’ emerged as a result of two important structural features evident in
Figure 17, namely the significant level of ‘self-inclusion’ displayed by the two DMB molecules, and the location of each guest molecule at the periphery of one of the DMB molecules in the ASU.
Application of the space group symmetry (2
1 parallel to
b) to each ASU and translation results in infinite spiral columns of self-included DMB molecules parallel to the
b-axis that preclude guest molecule entry, resulting in guest and water molecules being arranged in a spiral formation at the peripheries of the columns, as shown for the DMB·FA complex as representative (
Figure 18).
Complementing the binding of an FA molecule to the surface of the DMB column via its carboxylic acid group, the phenolic group and the methoxyl oxygen atom at the other terminus of the FA molecule engage in H-bonding to chains of water molecules located within the same interstitial channel (
Figure S16, Supplementary Materials). These water molecules link the FA molecules to neighboring columns of DMB molecules. Specifically, the phenolic group acts as both H-bond donor to a water oxygen atom [O1-H···O9W, O···O = 2.940(9) Å] and as an H-bond acceptor from a water molecule [O10W-H···O1, O···O = 2.68(1) Å], while the methoxyl atom (O2) is an acceptor of a H-bond from a water molecule [O9W-H···O2, O···O = 2.98(1) Å]. Thus, the FA molecule, being precluded from entry into the host molecule due to the latter’s self-inclusion, is wholly and strongly incorporated within the primarily hydrophilic environment of the interstitial channel, in contrast to the behaviors of the complexes described previously. Analogous observations and remarks apply to the crystal packing displayed by the isostructural complex DMB·CAF.
In view of the rarity of the type of self-inclusion described for these complexes and its possible significance in the context of the well-known phenomenon of CD aggregation, which occurs with increasingly higher CD concentrations in complex solutions [
19], further discussion of the geometrical features of this CD association is warranted. As shown in
Figure S17 (Supplementary Materials), the two DMB molecules (A and B) in the ASU of the 2:1 DMB·FA complex displayed the phenomenon of ‘self-inclusion’, two methoxyl groups of molecule B entering the cavity of molecule A via the secondary side, with their glucose residues abutting the inner surface of the DMB molecule A. The DMB molecules maintain their usual ‘round’ shape due to the presence of the O2n···H-O3(n-1) hydrogen bonds on their secondary sides [
20]. In addition, there is a single hydrogen bond that links the two cyclodextrin molecules directly, namely O3A4-H···O6B6 with H···A 1.85 Å, O···O 2.677(5) Å and bond angle 169°. We considered the following quantitative parameters as being suitable for defining the self-inclusion geometry: (a) the angle between the mean planes of the seven glycosidic (O4) atoms in each of molecules A and B (34.6° in DMB·FA), and (b) the distance between the centroids (Cgs) of the O4 atoms of molecules A and B (two independent distances in DMB·FA, namely CgA··CgB and CgB··CgA′ where A is located at
x, y, z and A′ at
a, 1 + y, z, namely 7.86 and 7.82 Å). The corresponding parameters for the closely isostructural complex DMB·CAF are 34.4°, 7.84, and 7.85 Å.
Interesting comparisons can be drawn between the isostructural non-inclusion complexes DMB·FA and DMB·CAF and other ‘irregular’ DMB complexes in the literature. An earlier example of a ‘non-inclusion’ complex is that of the hydrated α-CD complex with 2,5-dihydroxybenzoic acid (DHB), crystallizing in the space group
P2
12
12 [
21], originally described as a 1:1 α-CD-DHB noncovalent adduct (CSD refcode WIZQEB). In this crystal, the α-CD molecules retain the native CD channel-type packing features that they display in the crystal of the pure host, being aligned in head-to-tail mode, with intermolecular O–H···O hydrogen bonds maintaining the columns (
Figure S18, Supplementary Materials). The disordered DHB molecules, however, are located outside the host cavity region and are aligned in hydrogen bonded chains running parallel to the α-CD channel axis, but within the interstitial space created by four neighboring columns of α-CD molecules. Water molecules are also located in the interstitial space and engage in hydrogen bonded networks. This type of ‘non-inclusion’ complex differs very significantly from the type exemplified by the DMB complexes described in the present study, in particular due to the lack of the host ‘self-inclusion’ element.
The prolific X-ray structural studies of CDs and their complexes published by Harata [
22] include those of the isostructural complexes DMB·iodophenol and DMB·
p-nitrophenol [
23], with CSD refcodes DEZMOK10 and DEZMIE10, respectively. Our analysis of the published data show that in these structures, the DMB molecules also associate into columnar assemblies via self-inclusion, but to an extent that still permits simultaneous entry of the guest molecules into successive DMB molecules. This is illustrated in
Figure 19 for the iodophenol complex, as representative.
Comparing
Figure 18 and
Figure 19, the important difference is the absence of guest molecules within the host cavities in the former (showing the DMB·FA complex) due to the ‘tight’ self-inclusion of the DMB molecules that precludes guest entry, while in the latter (showing the DMB·iodophenol complex), the extent of guest inclusion is substantial. Specifically, a significant portion of the iodophenol molecule enters the extended cavity of the DMB molecule, the phenolic group engaging in hydrogen bonding with an intracavity water molecule, while the isostructural DMB·
p-nitrophenol complex displays analogous features. Such partial guest entry into the DMB cavity reduces the extent of self-inclusion of the DMB molecules, which is reflected in the following quantitative parameters for the DMB·iodophenol structure: (a) the angle between the mean planes of the seven glycosidic (O4) atoms of successive DMB molecules in the column was 48.7° (compared with only 34.6° in DMB·FA), and (b) the distance between the centroids (Cgs) of the O4 atoms of successive DMB molecules in the column was 8.69 Å (compared with only ~7.84 Å in the DMB·FA complex). The above self-inclusion parameters for the DMB·iodophenol crystal were very similar to those calculated for the isostructural DMB·
p-nitrophenol complex, namely 49.4° and 8.69 Å. Here, we conclude that, despite the common feature of host self-inclusion shown in
Figure 18 and
Figure 19, the new, isostructural complexes DMB·FA and DMB·CAF reported here are ‘non-inclusion’ complexes while DMB·iodophenol and DMB·
p-nitrophenol are ‘inclusion’ complexes. The latter complexes in fact resemble the DMB·HFA and DMB·PCA complexes that demonstrate partial guest inclusion (
Section 3.1.2) and for which the self-inclusion parameters are respectively 55.9°/8.66 Å, and 58.5°/8.96 Å, the somewhat larger angles that describe the self-inclusion being consistent with deeper guest inclusion within the host cavity.
A variation on the theme is a case of simultaneous inclusion and non-inclusion in a CD-containing crystal, reported by Harata [
24], and exemplified by the complex α-CD·(
m-nitrophenol)
2 (CSD refcode ACDMNP), crystallizing in the space group
P2
12
12. In this crystal, one of the
m-nitrophenol molecules is included within the host cavity while the other is located at the periphery of the host molecule (
Figure S19, Supplementary Materials), occupying a site within an interstitial channel generated by columns of complex units. This complex therefore represents the intermediate case between authentic inclusion complexes (such as DMB·iodophenol, DMB·
p-nitrophenol) and the authentic non-inclusion complexes DMB·FA and DMB·CAF.






















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}