
Mollusc N-glycosylation: Structures, Functions and Perspectives


Department of Chemistry, University of Natural Resources and Life Sciences, 1190 Vienna, Austria
Biomolecules 2021, 11(12), 1820; https://0-doi-org.brum.beds.ac.uk/10.3390/biom11121820
Submission received: 16 November 2021
/
Revised: 1 December 2021
/
Accepted: 1 December 2021
/
Published: 3 December 2021
(This article belongs to the Special Issue Glycosylation—The Most Diverse Post-Translational Modification)
Abstract
:Molluscs display a sophisticated N-glycan pattern on their proteins, which is, in terms of involved structural features, even more diverse than that of vertebrates. This review summarises the current knowledge of mollusc N-glycan structures, with a focus on the functional aspects of the corresponding glycoproteins. Furthermore, the potential of mollusc-derived biomolecules for medical applications is addressed, emphasising the importance of mollusc research.
1. Introduction
Mollusca is a large and evolutionary very successful phylum of animals comprising about 200,000 recognised species (alive or fossil) with very heterogenous morphology (gastropods, cephalopods, bivalvia). For more than 500 million years, since the Upper Cambrian, this phylum has populated freshwater, marine and terrestrial habitats worldwide. Some species are recognized as important members of several ecosystems in terms of waste disposal and cleaning, others are used due to their nutritional value and shells [1]. One class of molluscs, the cephalopoda (squid, octopus), is used in the medical context as a model for neuronal studies. However, other species, especially snails and slugs, are ill-reputed and known as pests in agriculture or hosts of parasite life cycles.
In general, their success in survival, their adaptability to changing environmental conditions and their immense potential in medical and pharmaceutical application (see below), make molluscs an interesting target for research. However, despite their significance, there is a general lack of knowledge regarding mollusc biochemistry or molecular biology. In addition, only few genomic resources are available, further hampering research [2]. However, the investigation of mollusc biosynthetic pathways, recognition processes and interactions in general can provide valuable knowledge.
Glycosylation plays an important role in altering the properties of proteins and lipids, modifying their interactions by changing the biophysical characteristics of a target molecule significantly. Attached glycans contribute not only to physical properties, such as conformational stability, protease resistance, charge, or hydrophilicity, but also modulate several types of recognition processes ranging from reproductive biology, self/non-self-recognition, cell-cell communication, cell trafficking, development, differentiation, host-pathogen or host-symbiont interaction, immune activation, cell death and even to evolution [3,4].
N-glycosylation, one of the most common types of glycosylation, is conserved between eukaryotes. The process starts in the endoplasmic reticulum (ER) by forming a lipid-linked precursor which is transferred en bloc onto the amide of an asparagine side chain located in the consensus sequence Asn-X-Ser/Thr, where X may be any amino acid except proline. Most commonly, except for unicellular species, the precursor is a Glc3Man9GlcNAc2 [5]. Subsequent modifications of N-glycans are then conducted in different areas of the Golgi apparatus. In a stepwise process, specific glycosidases and glycosyltransferases sequentially add and remove monosaccharides to form high mannosidic, paucimannosidic, hybrid or complex type N-glycans [6]. Further modifications, such as methyl groups, sulphate, phosphate, phosphorylcholine or phosphoethanolamine may also occur [7,8]. The final structure of the glycan depends on the organism, the tissue, the developmental and physiological stage of the cell and even the glycosylation site within the protein. Molluscs display a very broad spectrum of such N-glycans on their proteome. They combine structural features from mammals, plants, insects and nematodes, supplementing them with additional rare features.
This publication gives an overview on the still scattered knowledge of mollusc N-glycosylation. The data are arranged around probable functions of the corresponding glycoproteins. Within the chapters, the development of the methodical approach in glycan identification can also be observed. At the beginning of carbohydrate analysis, simple monosaccharide component analysis and selected lectin binding studies were state of the art, which was then followed by the determination of detailed structures by NMR. Recently, sensitivity of the methodical portfolio has increased dramatically with wide availability of different mass spectrometry and array techniques. However, these technically sophisticated methods must be supported by further tools, for example digestion by specific exoglycosidases or chemical treatment, to confirm linkages or to distinguish isoforms [9].
Besides the focus on N-glycosylation, this review also wants to emphasise the importance of further investigations on molluscs. Hence, it is supplemented by two further short chapters on lectins and cone glycopeptides, which do not deal directly with N-glycans, but aim to highlight the importance of gastropod research. The past months of the pandemic clearly showed the need to know much more about any biological interaction, even when a species is not so important at first glance. For molluscs, we still do not know what surprises, be they treasures or hazards, they contain.
2. Shell Matrix Protein Glycosylation
Mollusc shells are built of calcium carbonate and an elaborated organic matrix. Shell matrix proteins, components of this organic matrix, control the necessary biomineralisation process [10]. Posttranslational modifications of these proteins, including glycosylation, often contribute to this process by modulating the protein activity [11]. The significance of glycoproteins in the biomineralisation process could be confirmed clearly by deglycosylation experiments or the comparison of glycosylated with recombinant non-glycosylated versions [12,13,14]. Because most conducted studies on shell matrix protein glycosylation present only monosaccharide compositions, ratios of sugar to protein, or lectin binding experiments, structural details and linkage information are still rare. Only two molluscs have been studied in greater detail for their shell protein N-glycosylation: the tropical freshwater snail Biomphalaria glabrata and the marine bivalve Mytilus edulis.
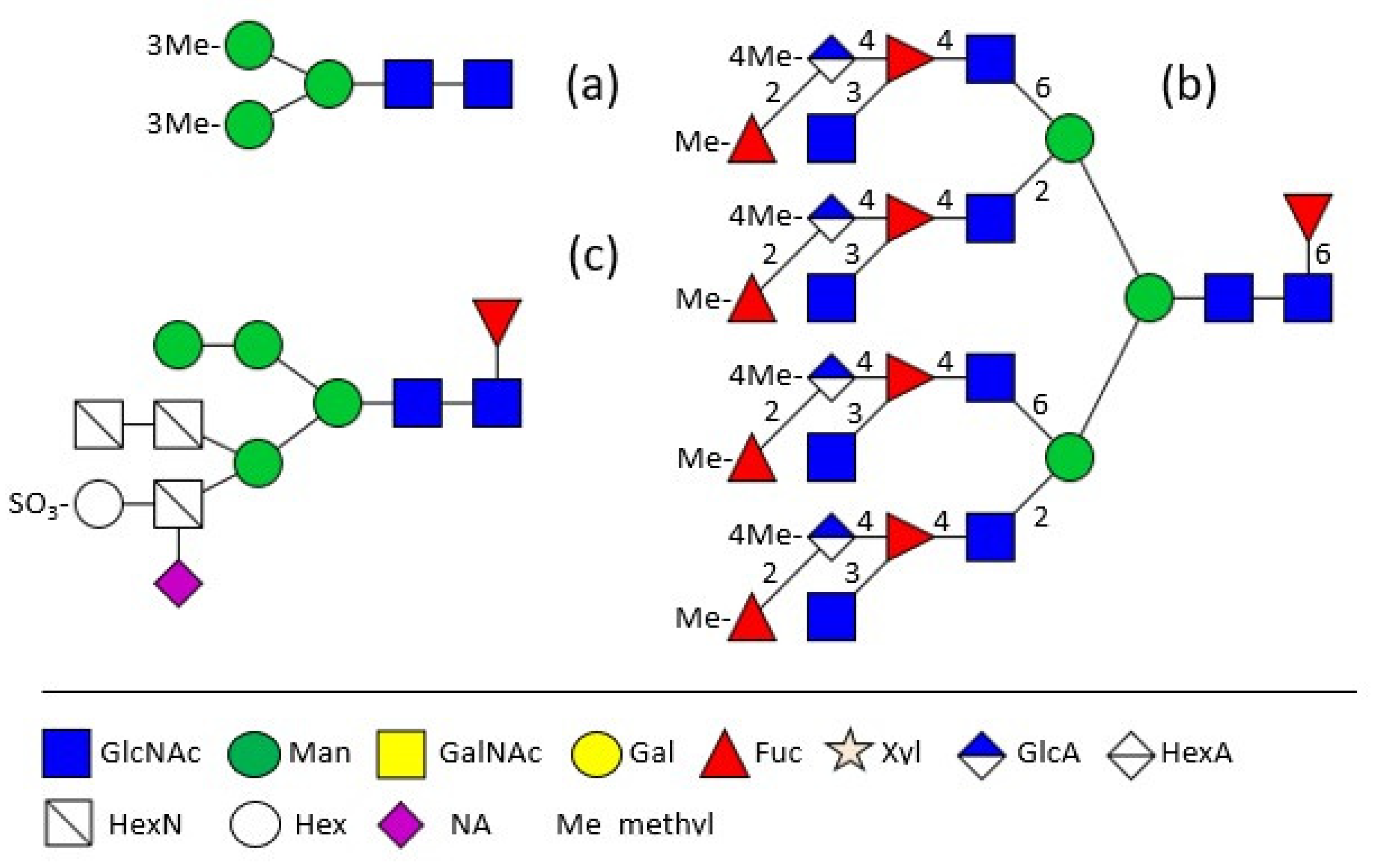
The major shell matrix protein of Biomphalaria glabrata, dermatopontin, is a 148 amino acid protein with one N-glycosylation site which is occupied by a 3-O-methylated pentasaccharide (Figure 1a) [15]. Completely different to this, the 28 kDa-sized major extrapallial protein of Mytilus edulis displays on its single N-glycosylation site a variation of large tetra- and penta-antennary 4-O-methylated structures with the rarely occurring structural feature of branched fucose residues (Figure 1b,c) [16]. In another marine bivalve, the Akoya pearl oyster Pincatada fucata, acidic compounds, namely sulphite linked to a terminal hexose as well as sialic acid, linked to a non-terminal hexosamine in the α1,3-antenna, have been identified [17].
3. Hemolymph Proteins
Hemocyanins are blue copper-containing respiratory proteins in the hemolymph of arthropods and molluscs. They are usually of high molecular weight and display a complex quaternary structure [18,19]. Due to their immunostimulating properties, which are at least partly caused by their glycans, hemocyanins are by far the best investigated proteins of snails.
Over the last decades, detailed investigation of these glycan structures has been made possible by the development and improvement of a number of analytical tools and instruments, especially mass spectrometry. Today, specific glycosylation sites within a polypeptide chain as well as minimal amounts of material can be analysed. By using these tools, mollusc glycans never cease to surprise researchers due to their various glycan structures and additional features. Here, a short overview is given, focusing in particular on new and uncommon structural features found for the first time in mollusc hemocyanins.
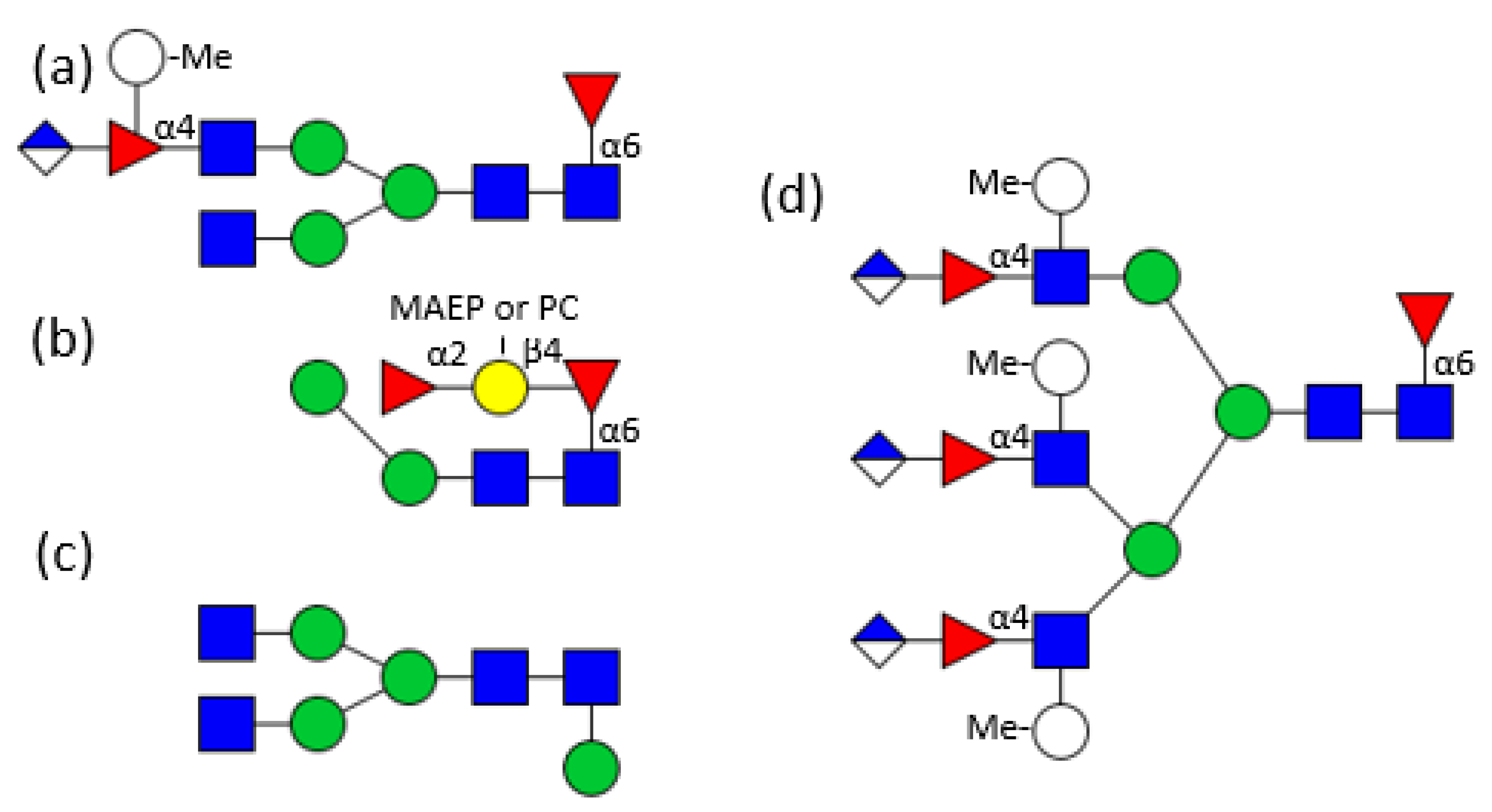
Detailed analysis started at the end of the seventies with the identification of 3-O-methylated mannose and 3-O-methylated galactose in the course of monosaccharide analyses of Helix pomatia and Lymnaea stagnalis glycans [20]. Then, in the mid-eighties, the first glycan structure was determined by 500 MHz 1H NMR spectroscopy. Xylose, known from cartilage and plants, was found in the Helix pomatia α-hemocyanin (Figure 2a) [21], followed by the identification and exact location of 3-O-methylated mannoses in the hemocyanin of the freshwater snail Lymnaea stagnalis (Figure 2b) [22]. 3-O-methylated galactose could be found in Lymnaea stagnalis as well (Figure 2c), but also in the marine snail Rapana thomasiana (today named Rapana venosa) and to an even higher extend in the land snail Helix pomatia [23,24,25]. In Helix pomatia 4-O-methylated galactose was determined, additionally [25]. Another feature, 6-O-methylated mannose, which is a constituent of the otherwise high mannosidic glycans of tridacnin, a lectin found in the hemolymph of the clam Hippopus hippopus, could be identified as well [26]. At first they were considered a novelty, but by now xylose and methylated hexoses have been found in many other molluscs and are recognised as a typical feature of mollusc glycans.
Even more unusual structures were found in the functional unit of RvH1-a of Rapana venosa hemocyanin. Besides a biantennary structure terminated by two methylated galactoses, a sulphated glycan with a methylated bisecting GlcNAc was identified (Figure 2d) [27].
Analysis of keyhole limpet (Megathura crenulata) hemocyanin revealed another two novel modifications. Firstly, a galactose residue directly β1,6 linked to mannose was identified on high mannosidic or hybrid N-glycans, some substituted by an additional core α-fucosylation (Figure 2e) [28] and, secondly, a chain of two β1,4 linked galactoses substituting the α1,6 linked core fucose (Galβ1,4Galβ1,4Fucα1,6-) of another group of structures, was found (Figure 2f) [29]. A single galactose linked to the inner fucose had already been seen before in another protein derived from mollusc origin, the octopus’s rhodopsin, but this structure with two galactoses had been completely unknown until then [30].
Soon afterwards, not only mono-substituted, but also branched fucoses were identified in the glycan antennae. RtH2-e, the functional unit of Rapana venosa hemocyanin, contains a hybrid glycan structure with a fucose located in the α1,3 antenna substituted by GlcNAc as well as by 3-O-methyl-galactose (Figure 3a) [31]. In RvH1 and RvH2, however, the fucose is substituted by an amino sugar and a hexuronic acid (Figure 3b) [32,33]. In contrast to that, HtH1, the hemocyanin functional unit of Haliotis tuberculata, displays the more common structure with unsubstituted fucoses linked to the antennal GlcNAc residues (Figure 3c) [34].
Biomphalaria glabrata hemolymph hybrid N-glycans mostly carry a xylose in the 2-position to the central mannose and an α1,6 fucose linked to the innermost GlcNAc of the core. Their α1,3 linked antenna is elongated by a GalNAc-GlcNAc moiety, with both amino sugars fucosylated (Figure 3d) [35], whereas Helix pomatia displays up to six methylated galactoses on the antennae (Figure 3e) [25,36]. Similar structures decorate the 13 potential glycosylation sites of the β HlH subunit of Helix lucorum hemocyanin. The inner core of the 32 different identified glycans is often α1,6 fucosylated and/or β1,2 xylosylated. A significant number of these glycans is of the hybrid type with only one mannose on the α1,6 antenna, but a GalNAc-GlcNAc elongation, terminated by several methylated hexoses, on the α1,3 antenna [37].
A recent review compares on a very high technical level N-glycan structures, glycan-protein binding sites and conformational aspects of hemocyanins found in the Black Sea snail Rapana venosa, the garden snail Helix lucorum and the abalone Haliotis tuberculata [38]. All investigated organisms show a high content of methylated hexoses (3-O-methyl mannose and 3-O-methyl galactose) and acidic structures. In fact, comparative analysis of amino acid sequences, close to the putative N-glycosylation sites, on the surface of the functional units show a high degree of homology, thus implying an important contribution of the glycans to the function [38].
Hemocyanins carrying typical mollusc structural features on their glycans, such as fucosylation, xylosylation or methylated hexose residues, display strong immune stimulatory properties in animal studies and therefore, are used to enhance the efficiency of anti-tumour vaccines [39]. The importance of glycans and their contribution to the structure of these large protein complexes became evident in deglycosylation experiments. Deglycosylation of hemocyanins from different mollusc origin (Rapana thomasiana, Concholepas concholepas, Fissurella latimarginata, Megathura crenulata) disrupts the quaternary structure of the protein complexes and reduces their immunogenic capacities. Especially methylated galactoses and xylose seem to be the key players of this effect [36,40,41]. In vitro studies on hemocyanins from the marine snail Rapana venosa and the garden snails Helix aspersa and Helix lucorum revealed antitumor effects on human colorectal adenocarcinoma cells [42]. A recent study shows that hemocyanins from Concholepas concholepas and Fissurella latimarginata are safe and useful carriers of carbohydrate mimotopes and therefore new suitable candidates for the enhancement of the immunogenicity of peptides in cancer vaccine research [43]. For the last several years, keyhole limpet hemocyanin from Megathura crenulata has been successfully utilised for treatment of various types of cancer. These treatments have led to enhanced recognition of the importance of carbohydrates [for a review see [44]. In the specific case of superficial bladder cancer, however, the involvement of glycans on the hemocyanins from Megathura crenulata, Concholepas concholepas and Fissurella latimarginata seem not be necessary to bias the immune response towards the cancer cells through natural, complement-activating antibodies [45].
Research on immuno-stimulative properties of mollusc glycoproteins is ongoing and expanded to additional applications. Besides cancer treatment, the immune stimulatory benefits of snail hemocyanins have been successfully evaluated for viral infections. Rapana venosa RvH2-e, for example, which contains glycans essential for its activity, exhibits an antiviral effect against herpes simplex virus type 1 (HSV-1) [33].
4. Glycosylation of Other Mollusc Proteins
Much less is known about N-glycosylation of other proteins. Multiple studies compared N-glycan patterns of complete adult tissue preparations (viscera and skin) of terrestrial slugs (Arion lusitanicus, Limax maximus), terrestrial snails with shells (Cepaea hortensis, Arianta arbustorum, Achatina fulica) and a freshwater snail (Planorbarius corneus) for their similarities and differences. All analysed snails contained mainly high mannosidic and paucimannosidic structures terminated by 3-O-methylated mannose residues. The inner core was decorated with a β1,2-linked xylose at the β-mannose and often fucosylated at the inner GlcNAc, mainly in α1,6 position. Only in Limax maximus and Achatina fulica some larger structures containing galactose and/or 3-O-methylated galactose were observed [46,47]. Neither in Arion lusitanicus eggs nor on the extracellular coat of the bivalve Unio elongatulus were methylated hexoses discovered, thus glycosylation patterns undoubtedly are developmentally regulated and vary between different stages [46,48]. This finding is supported by a study on Achatina fulica eggs, young snails and adults showing remarkable differences in their glycosylation patterns [49].
Rhodopsin, the visual pigment in photoreceptor cells of the cephalopod Todarodes pacificus, has been studied as well and is mainly glycosylated (85%) by a α1,3/α1,6 difucosylated pentasaccharide core with the α1,6 fucose elongated by a galactose residue in β1,4 position. No methylated sugars were detected here [50].
A whole glycome analysis was performed on the total tissue of the marine bivalve Volvarina rubella revealing a high complexity of about 100 different N-glycan structures. While the common mollusc structural characteristics, such as xylose or methylation of hexoses were either not present or in low abundance respectively, some new features were identified. Additional mannose residues were found linked to the innermost GlcNAc in α1,3 position and peripheral in an unusual α1,6 position. The α1,6 linked fucose of the inner core was not only substituted by a β1,4 linked galactose, but the chain was further substituted by an α1,2 linked fucose residue. Phosphorylcholine and up to four N-methyl-2-aminoethylphosphonate molecules on mannose and GlcNAc residues increased the heterogeneity. Additionally, sulphatation of mannose residues or trisaccharide units containing a branched fucose substituted by a methylated hexose and uronic acid were revealed to be responsible for the charge of anionic N-glycans (Figure 4a–d) [51].
So far, nothing is known about the function of these unusual decorations of N-glycans found in the different studied species. Presumably, similar to all other phyla, glycans in molluscs play an important role in many recognition processes within and among cells. It is clear, however, that at least neuronal processes are influenced by glycoproteins in snails. Helix pomatia nervous tissue contains an N-glycosylated glycoprotein (F3/contactin immunologically related protein, HCRP), which is involved in axonal growth control and neurotransmitter release and in Lymnaea stagnalis an increase of glycosylation during long–time-memory formation was detected. However, no detailed information is given regarding the glycan structures involved [52,53].
5. Molluscs as Intermediate Hosts for Parasites
Snails are the intermediate hosts for several parasites, mainly trematodes, which need the molluscs for some steps of their life cycle. In many of the diverse recognition events during this process, for example, recognition of a proper host, adhesion or invasion, protein linked carbohydrates are at least one of the binding partners.
Biomphalaria glabrata is the intermediate host of the trematode Schistosoma mansoni, the causative agent of human schistosomiasis, which affects hundreds of millions of people worldwide. Hence, it is the best investigated intermediate host/parasite system. The snails are essential for asexual reproduction in this parasite’s life cycle. [54,55,56]. Because of its human relevance, a Biomphalaria glabrata embryonic (Bge) cell line, the only established cell line from mollusc origin, is available for research [57].
Several galectins, lectins or lectin-like proteins have been found in snails which play a role in the recognition of parasite-specific glycan motifs and may also be correlated to susceptibility or resistance of snail strains towards infection by the parasite [58]. Several glycan structures have been identified which play major roles in the blood fluke—snail interaction [59,60,61,62]. For example, a tandem-repeat like galectin was identified in Bge cells, as well as in circulating hemocytes of adults of Biomphalaria glabrata, displaying Lac or Gal binding activity [63]. These N-glycans of hemolymph glycoproteins, which cross react with Schistosoma mansoni glycoconjugates, are paucimannosidic or hybrid structures containing often an α1,6 linked fucose and/or a β1,2 xylose linked to the core pentasaccharide. The α1,3 antenna of the glycan is elongated by a LacdiNAc unit (GalNAcβ1,4GlcNAcβ1-) containing up to two α1,3 linked fucose residues (Figure 3d) [35]. These structural features are shared with Schistosoma mansoni sporocytes and miracidia, two snail-pathogenic larval stages of the parasite [64]. Further studies demonstrated that especially the fucose residues are essential for the interaction between host and parasite [65]. Enhanced occurrence of glycans with two or three fucose residues on the hemolymph glycoproteins of the Puerto Rico Biomphalaria glabrata strain (BgPR) are correlated with the high susceptibility of this strain towards parasite infection [66].
The great pond snail Lymnaea stagnalis is the intermediate host for the larvae of a variety of bird schistosomes (genus Trichobilharzia), the causative agent of cercarial dermatitis, as well as for the liver fluke Fasciola hepatica, a common parasite in ruminants and also humans [67,68]. As a reaction to infection, Lymnaea stagnalis down-regulates cell signalling, which is at least partly caused by fucose and galactose [69]. Fasciola hepatica and its further host, Galba truncatula, display structural similarities of surface carbohydrate residues in the contact zone between both organisms. This suggests that carbohydrate mimicry is used by the parasite to evade host immune response [70].
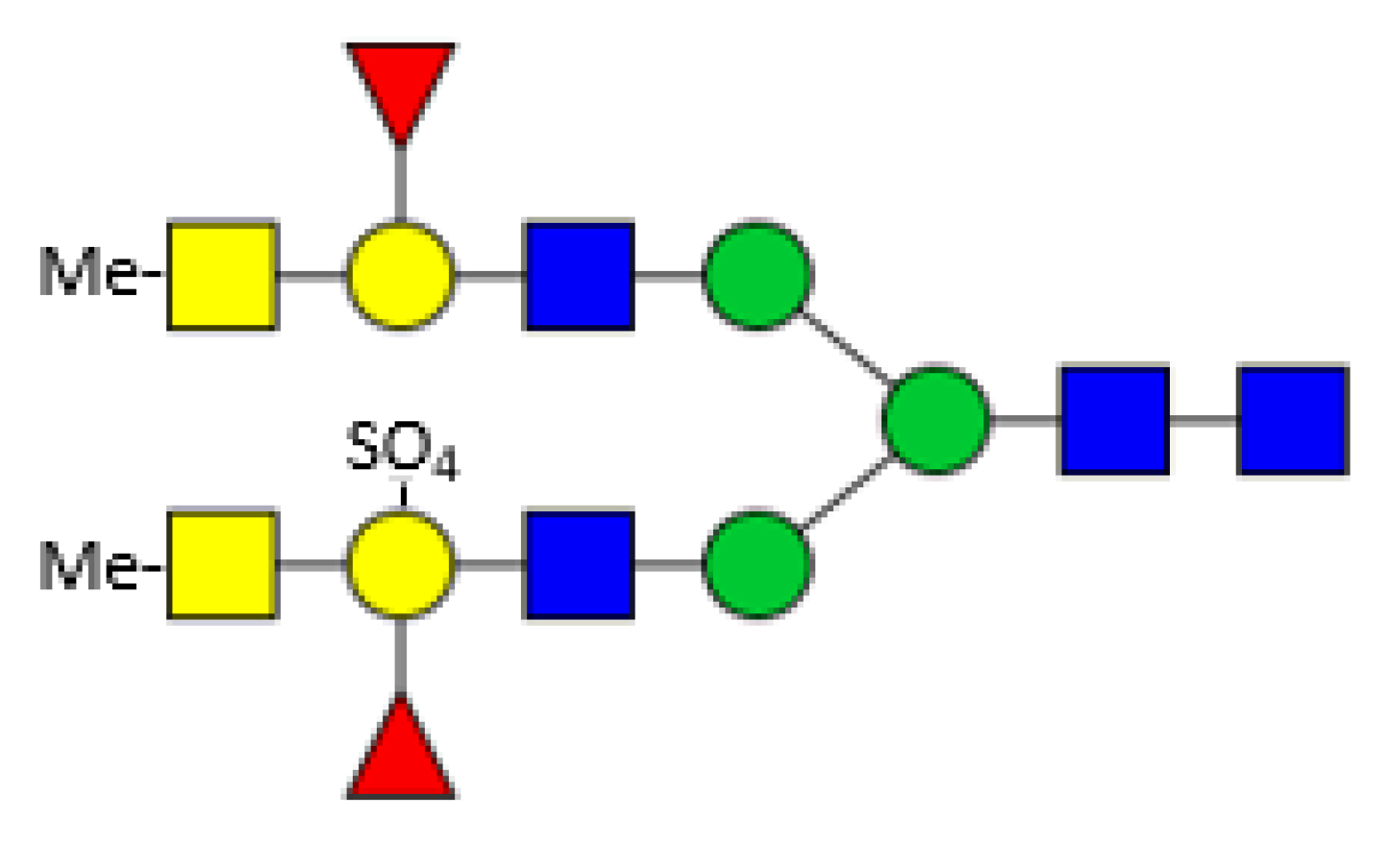
Eastern oysters (Crassostrea virginica) contain two galectins (CvGal1 and CvGal2) which are involved in the immunological response upon infection by the protozoan parasite Perkinsus marinus. CvGal1 binds in vitro strongly to type-2 blood group A oligosaccharides, but in the biological context binding to type-1 blood group A glycans may be preferred [71,72]. CvGal2 displays a broader specificity and can distinguish between different Perkinsus species [73]. The oyster plasma and hemocyte glycans comprise a broad spectrum of paucimannosicic, hybrid and complex structures containing core α1,6 fucosylation, core α1,3/α1,6 difucosylation and some structures with an α1,3 linked hexose at the innermost GlcNAc residue. The antennae are often terminated by methylated GalNAc residues. The acidic glycans are substituted by one or more sulphate groups linked to terminal or innermost galactose residues comprising modified forms of human histo-blood group A antigens (Figure 5) [74]. These glycans fit perfectly to the specificity of the previously mentioned Biomphalaria glabrata galectins. Surprisingly, no xylose could be detected here.
In addition, different nematodes use snails, for example Achatina fulica, as intermediate hosts in their life cycles, but no detailed information on glycan involvement is given [75].
Keyhole limpet (Megathura crenulata) is not susceptible to Schistosoma mansoni infection, but its hemocyanin glycans cross-react serologically with glycoconjugates of this parasite. The epitope responsible for this was identified as the Fucα1,3GalNAcβ1,4(Fucα1,3)GlcNAcβ1- motif. Using the cross-reaction behaviour, keyhole limpet is a valuable tool in diagnosis and may act as a model antigen for further studies on immunopathological mechanisms [76]. Our knowledge about intermediate host-pathogen interaction processes is still scattered. Efforts in this research area need to be increased in order to provide ways to interrupt parasite life cycles, to avoid tremendous numbers of infections worldwide.
6. Enzymes Involved in N-glycan Biosynthesis
By now, the decreasing costs of DNA sequencing has enabled full sequencing of various species. However, for molluscs there is still much less genetic information available, compared to other phyla. Fewer than 50 studies describe partial or complete genome mollusc assemblies, for example those of the Pacific oyster Crassostrea gigas (synonym Magallana gigas) or the freshwater snail Biomphalaria glabrata, the intermediate host of the human blood fluke Schistosoma mansoni [2,77,78]. From the latter source an immortalised cell line has been developed (Biomphalaria glabrata embryonic cell line—Bge), which thus is the only available lophotrochozoan in vitro cell model. However, it must be emphasised that the laboratory strain differs both in sequence and structure from the reference genome [79]. For many other mollusc species only limited data are available so far. The missing opportunity for homology search is one main reason why the knowledge of enzymes and their characteristics is so limited within this phylum.
Molluscs are important players as waste removers in the environment. To accomplish this, they contain a number of oligosaccharide degradation enzymes, such as mannanases, cellulases, chitinases or glucosidases. Several such exo- and endoglycosidases have been identified, characterised and partly recombinantly produced. As this review mainly focuses on N-glycosylation, here only enzymes taking part in the N-glycosylation process are described.
In respect of N-glycan biosynthetic pathway enzymes, only the freshwater snail Lymnaea stagnalis has been well investigated. Several enzymes responsible for the formation of typical complex mollusc glycans were identified in the connective tissue of this snail. In nearly all phyla, including molluscs, biosynthesis of complex N-glycans is initiated by the transfer of a GlcNAc residue to the α1,3 antenna of an N-glycan as prerequisite for further enzymes. GlcNAc-transferase I (β1,2-N-acetylglucosaminyltransferase I) action then allows subsequent modification by GlcNAc-transferase II (β1,2-N-acetylglucosaminyltransferase II) and β1,2-xylosyltransferase [80]. Terminal GlcNAc residues can be further elongated by β1,4-GalNAc-transferase (UDP-GalNAc: GlcNAc β-R β1,4-N-acetylgalactosaminyltransferase) forming a LacdiNAc-chain (GalNAcβ1,4GlcNAc), which occurs frequently in invertebrates [81,82]. Onward elongation is performed by β1,3-galactosyltransferase [83]. In addition, three types of fucosyltransferases have been described. Two of them act on terminal Gal or GlcNAc residues (GDP-Fuc:Galβ1,3GalNAc-R(Fuc to Gal) α1,2 fucosyltransferase and GDP-Fuc:Gal β1,4GlcNAc (Fuc to GlcNAc) α1,3 fucosyltransferase [84], while the third one (core α1,3 fucosyltransferase) adds a fucose onto the innermost GlcNAc residue forming an α1,3 core fucosylation [85]. Even though many glycans have been identified, which contain fucose α1,6 linked to the inner GlcNAc, the corresponding α1,6 fucosyltransferase in mollusc species is still unknown.
In contrast to vertebrates, where GlcNAc residues of the antennae are usually substituted by Gal residues, in molluscs a terminal GalNAc residue is frequently added. Interestingly, the responsible enzyme, the β1,4-GalNAc-transferase, shows high DNA homology and structural similarity to the mammalian β1,4-galactosyltransferase [81,82]. In addition, a β1,4-GlcNAc-transferase with clear homology to the mammalian β1,4-galactosyltransferase was identified, but this enzyme is more likely involved in O-glycan biosynthesis [86]. In the prostate gland of Lymnaea stagnalis a β1,4-glucosyltransferase was found, which can glucosylate terminal GlcNAc residues of O-glycans as well as of N-glycans. However, the products of this enzyme’s activity could not be determined in tissues so far [87].
An essential requirement requisite for the production of any glycan is the availability of the appropriate activated sugar nucleotides. One of these vital genes was identified in the genome of the Pacific oyster Magallana gigas (synonym: Crassostrea gigas). It was proven to encode UDP-Glucose-4-epimerase as the corresponding protein could catalyse the conversion of UDP-Glc into UDP-Gal [88]. In the same study, another gene, a β1,4-galactosyltransferase from oyster, was expressed and purified but not checked for activity on N-glycan substrates [88].
Overall, the biosynthesis of N-glycans in molluscs seems to obey in general what we know from other phyla regarding the first basic steps. When the N-glycosylation process of mammals was compared to that of Mytilus galloprovincialis digestive cells along the Golgi apparatus, only a handful of differences were found: a lack of sialic acid, a very prompt galactosylation and an increased fucosylation of the antennae [89]. However, even though the first steps are quite similar, further modifications of the glycans are far more variable in terms of compounds (sugars and other additions, such as methyl or sulphate groups) and possible linkages. The consequence is a much wider spectrum of possible glycan structures than identified in vertebrates or plants.
Currently the only application for mollusc enzymes discussed is a usage of the impressive transglycosylation ability of some glycosidases. This would enable an easy and environmentally friendly production of various disaccharides. For example, the β-D-glycosidase from the China white jade snail (a special breeding of Achatina fulica) can transfer fucose, glucose as well as galactose to different acceptor substrates [90].
Due to the broad spectrum of glycan structures in molluscs, it can be assumed that a number of glycosyltransferases with new specificities are present. Those enzymes, recombinantly expressed, would allow new advances in chemoenzymatic synthesis of glycans which may be useful for diagnosis and treatment of several diseases.
7. Current and Prospective Applications of Mollusc Lectins
Lectins are proteins that bind specifically to carbohydrate structures. They are part of numerous recognition and adhesion processes in all phyla. Their strict restriction to specific sugars in specific linkages allows a high selectivity and discrimination for interactions in the living organism, but also provide a powerful tool for basic research, diagnostic and therapeutic applications. Already for more than forty years, lectins from mollusc origin have been isolated and characterised. Presumably, they play a crucial role in innate immune protection. Due to their binding specificity, they are targeted to structural elements characteristic for bacteria. For this purpose, sialic acid or galactose-binding lectins are found in many mollusc species (Achatina fulica, Cepeaea hortensis, Limax flavus, Mytilus californianus) with a putative role in the defence against microorganisms [91,92,93,94,95]. An upregulation of a sialic acid-binding lectin of Crassostrea hongkongensis is visible upon bacterial infection [96]. In Achatina fulica a high molecular weight lectin specific for galactose does not inhibit bacterial growth, but rather induces agglutination of the microorganisms [97]. Furthermore, lectins also have other tasks in molluscs, for example the protection of eggs by an anti-nutritive defence function. A highly stable, non-digestible lectin was described and characterised revealing this function in Pomacea diffusa [98]. For a review on carbohydrate recognition in mollusc immune defence see [99].
Many of these lectins have the potential to be used as powerful tools in medicine. Malignant transformations of cells often correlate with an altered glycosylation pattern on the surface of those cells. Lectins, which specifically bind to these structures, meanwhile are used in diagnosis and treatment of various kinds of cancer. This includes several mollusc derived lectins which are under investigation or already in application as biomarkers. For example, a Helix pomatia lectin, which recognises O-GlcNAc residues in metastatic breast cancer and a Helix aspersa lectin, which recognises O-GalNAc on various cancer cells are already used for diagnosis [100,101,102,103,104].
A completely different potential application, but nonetheless important, is possible with the C-type hemolytic lectin from the freshwater clam Villorita cyprinoides. This lectin has a significant clot lysis activity and could be developed as a new antithrombotic drug molecule [105].
8. Glycopeptides from Conus Marine Molluscs
Predatory cone snails (genus: Conus) use complex venoms to paralyse and kill their prey. These venoms are mostly composed of multiple small peptides with a high content of disulphide bridges and posttranslational modifications. Due to the high diversity between Conus species, the total number of different existing peptides was estimated to be around 50,000 [106]. Many of the already identified peptides are mucin-type O-glycosylated, but no N-glycosylation has been described so far [107,108]. Some of the peptides interact with nicotinic acetylcholine receptors or are able to desensitise neurotensin receptors [109,110]. Peptides with low toxicity but a capacity to block these receptors irreversibly may be of pharmaceutical use in a broad field of applications, such as pain, epilepsy, and myocardial infarction. For a detailed review on cone toxins see [111].
9. Conclusions
Molluscs are able to produce all kinds of N-glycan structures ranging from those typical in mammals to structures similar to those found in plants, insects or nematodes. They also add further interesting features to the portfolio of glycosylation in living cells. This makes them a valuable model for research on N-glycosylation in general and may help to understand several interaction processes between cells or organisms, such as fertilisation, immunomodulatory activity or host-parasite recognition events.
Potential medical applications of mollusc components range, due to the high immunomodulatory capacity, from diagnostic and therapeutic tools in cancer to models for neuroscience research (cephalopoda) or vaccines against parasites. Furthermore, antimicrobial peptides derived from snail origin may contribute significantly to the development of novel antimicrobial therapies, especially in times of increasing numbers of antibiotic-resistant bacteria and antibiotic-allergic patients.
However, other fields also could benefit from mollusc research, for example material sciences. Some interesting inputs might be gained from shell formation for the development of new robust and stable biocompatible materials or from the mucus for the development of different kinds of surface lubrication.
Last, but not least, gastropods may play an increasing role in future protein supply solutions, perhaps a necessity for the nutrition of a growing world population.
Mollusc glycosylation is a treasure chest of different structural features and combinations thereof. We are far away from understanding their function, but some of these glycoproteins might be promising candidates for future applications in human health. The detailed knowledge of this very complex glycosylation system could become a valuable tool to understand the principal rules of glycosylation in all organisms. However, the challenging and time-consuming analysis methods, the limited genetic data and the unavailability of fitting bioinformatics tools, most based only on mammalian structural features, currently hinder a fast progress in mollusc research. Nevertheless, the evolutionary, medical and veterinary importance of this phylum is worth the investment of money and time to advance research.
This review gives an overview on the current knowledge on mollusc N-glycosylation, in terms of structure, corresponding enzymes and putative function. However, we are still far away from forming a complete picture of this immense glycosylation potential. Increasing scientific effort in this complex field will be a benefit for advancing glycoscience in general.
Funding
This research was funded by the Austrian Science Fund (FWF), grant number P33239-B.
Acknowledgments
Open Access Funding by the Austrian Science Fund (FWF).
Conflicts of Interest
The author declares no conflict of interest.
Abbreviations
Fuc, fucose; Gal, galactose; GalNAc, N-Acetylgalactosamine; Glc, glucose; GlcA, glucuronic acid; GlcNAc, N-Acetylglucosamine; Hex, hexose; HexA, hexuronic acid; HexN, Hexosamine, Man, mannose; Me, methyl; NA, neuraminc acid; Xyl, xylose.
References
- Wanninger, A.; Wollesen, T. The evolution of molluscs. Biol. Rev. Camb. Philos. Soc. 2018, 94, 102–115. [Google Scholar] [CrossRef]
- Gomes dos Santos, A.; Lopes-Lima, M.; Castro, L.F.C.; Froufe, E. Molluscan genomics: The road so far and the way forward. Hydrobiologia 2020, 847, 1705–1726. [Google Scholar] [CrossRef]
- Varki, A. Nothing in glycobiology makes sense, except in the light of evolution. Cell 2006, 126, 841–845. [Google Scholar] [CrossRef] [Green Version]
- Varki, A. Evolutionary forces shaping the Golgi glycosylation machinery: Why cell surface glycans are universal to living cells. Cold Spring Harb. Perspect. Biol. 2011, 3, a005462. [Google Scholar] [CrossRef] [Green Version]
- Aebi, M. N-linked protein glycoylation in the ER. Biochim. Biophys. Acta 2013, 1833, 2430–2437. [Google Scholar] [CrossRef] [Green Version]
- Stanley, P.; Taniguchi, N.; Aebi, M. N-glycans. Structure and Biosynthesis. In Essentials of Glycobiology, 3rd ed.; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G.W., Aebi, M., Darvill, A.G., Kinoshita, T., Packer, N.H., Prestegard, J.H., et al., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2015. [Google Scholar]
- Staudacher, E. Methylation—An uncommon modification of glycans. Biol. Chem. 2012, 393, 675–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paschinger, K.; Wilson, I.B.H. Anionic and zwitterionic moieties as widespread glycan modifications in non-vertebrates. Glycoconj. J. 2020, 37, 27–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hykollari, A.; Paschinger, K.; Wilson, I.B.H. Negative-mode mass spectrometry in the analysis of invertebrate, fungal, and protist N-glycans. Mass Spec. Rev. 2021, 1–19. [Google Scholar] [CrossRef]
- Addadi, L.; Joester, D.; Nudelman, F.; Weiner, S. Mollusk shell formation: A source of new concepts for understanding biomineralization processes. Chemistry 2006, 12, 980–987. [Google Scholar] [CrossRef]
- Rivera-Pérez, C.; Hernández-Saavedra, N.Y. Post-translational modifications of marine shell matrix proteins. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2021, 256, 110641. [Google Scholar] [CrossRef]
- Marie, B.; Zanella-Cléon, I.; Corneillat, M.; Becchi, M.; Alcaraz, G.; Plasseraud, L.; Luquet, G.; Marin, F. Nautilin-63, a novel acidic glycoprotein from the shell nacre of Nautilus macromphalus. FEBS J. 2011, 278, 2117–2130. [Google Scholar] [CrossRef] [PubMed]
- Chang, E.P.; Perovic, I.; Rao, A.; Cölfen, H.; Evans, J.S. Insect cell glycosylation and its impact on the functionality of a recombinant intracrystalline nacre protein, AP24. Biochemistry 2016, 55, 1024–1035. [Google Scholar] [CrossRef]
- Juan-Colas, J.; Jung, Y.S.; Johnson, S.; Evans, J.S. A complicated relationship: Glycosylation, Ca(II), and primary sequence affect the interactions and kinetics between two model mollusk shell intracrystalline nacre proteins. Biochemistry 2020, 59, 346–350. [Google Scholar] [CrossRef]
- Marxen, J.C.; Nimtz, M.; Becker, W.; Mann, K. The major soluble 19.6 kDa protein of the organic shell matrix of the freshwater snail Biomphalaria glabrata is an N-glycosylated dermatopontin. Biochim. Biophys. Acta. 2003, 1650, 92–98. [Google Scholar] [CrossRef]
- Zhou, H.; Hanneman, A.J.; Chasteen, N.D.; Reinhold, V.N. Anomalous N-glycan structures with an internal fucose branched to GlcA and GlcN residues isolated from a mollusk shell-forming fluid. J. Proteome. Res. 2013, 12, 4547–4555. [Google Scholar] [CrossRef] [Green Version]
- Takakura, D.; Norizuki, M.; Ishikawa, F.; Samata, T. Isolation and characterization of the N-linked oligosaccharides in nacrein from Pinctada fucata. Mar. Biotechnol. 2008, 10, 290–296. [Google Scholar] [CrossRef]
- Kato, S.; Matsui, T.; Gatsogiannis, C.; Tanaka, Y. Molluscan hemocyanin: Structure, evolution, and physiology. Biophys. Rev. 2018, 10, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Kato, S.; Matsui, T.; Tanaka, Y. Molluscan Hemocyanins. Subcell. Biochem. 2020, 94, 195–218. [Google Scholar] [CrossRef]
- Hall, R.L.; Wood, E.J.; Kamerling, J.P.; Gerwig, G.J.; Vliegenthart, J.F.G. 3-O-methyl sugars as constituents of glycoproteins. Identification of 3-O-methylgalactose and 3-O-methylmannose in pulmonate gastropod haemocyanins. Biochem. J. 1977, 165, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Van Kuik, J.A.; van Halbeek, H.; Kamerling, J.P.; Vliegenthart, J.F. Primary structure of the low-molecular-weight carbohydrate chains of Helix pomatia α-hemocyanin. Xylose as a constituent of N-linked oligosaccharides in an animal glycoprotein. J. Biol. Chem. 1985, 260, 13984–13988. [Google Scholar] [CrossRef]
- Van Kuik, J.A.; Sijbesma, R.P.; Kamerling, J.P.; Vliegenthart, J.F.; Wood, E.J. Primary structure of a low-molecular-mass N-linked oligosaccharide from hemocyanin of Lymnaea stagnalis. 3-O-methyl-D-mannose as a constituent of the xylose-containing core structure in an animal glycoprotein. Eur. J. Biochem. 1986, 160, 621–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Kuik, J.A.; Sijbesma, R.P.; Kamerling, J.P.; Vliegenthart, J.F.; Wood, E.J. Primary structure determination of seven novel N-linked carbohydrate chains derived from hemocyanin of Lymnaea stagnalis. 3-O-methyl-D-galactose and N-acetyl-D-galactosamine as constituents of xylose-containing N-linked oligosaccharides in an animal glycoprotein. Eur. J. Biochem. 1987, 169, 399–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoeva, S.; Rachev, R.; Severov, S.; Voelter, W.; Genov, N. Carbohydrate content and monosaccharide composition of Rapana thomasiana grosse (Gastropoda) hemocyanin and its structural subunits. Comparison with gastropodan hemocyanins. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1995, 110, 761–765. [Google Scholar] [CrossRef]
- Lommerse, J.P.; Thomas-Oates, J.E.; Gielens, C.; Préaux, G.; Kamerling, J.P.; Vliegenthart, J.F. Primary structure of 21 novel monoantennary and diantennary N-linked carbohydrate chains from alpha D-hemocyanin of Helix pomatia. Eur. J. Biochem. 1997, 249, 195–222. [Google Scholar] [CrossRef]
- Puanglarp, N.; Oxley, D.; Currie, G.J.; Bacic, A.; Craik, D.J.; Yellowlees, D. Structure of the N-linked oligosaccharides from tridacnin, a lectin found in the haemolymph of the giant clam Hippopus hippopus. Eur. J. Biochem. 1995, 232, 873–880. [Google Scholar] [CrossRef]
- Dolashka-Angelova, P.; Beck, A.; Dolashki, A.; Beltramini, M.; Stevanovic, S.; Salvato, B.; Voelter, W. Characterization of the carbohydrate moieties of the functional unit RvH1-a of Rapana venosa haemocyanin using HPLC/electrospray ionization MS and glycosidase digestion. Biochem. J. 2003, 374, 185–192. [Google Scholar] [CrossRef]
- Kurokawa, T.; Wuhrer, M.; Lochnit, G.; Geyer, H.; Markl, J.; Geyer, R. Hemocyanin from the keyhole limpet Megathura crenulata (KLH) carries a novel type of N-glycan with Gal(β1,6)Man-motifs. Eur. J. Biochem. 2002, 269, 5459–5473. [Google Scholar] [CrossRef]
- Wuhrer, M.; Robijn, M.L.; Koeleman, C.A.; Balog, C.I.; Geyer, R.; Deelder, A.M.; Hokke, C.H. A novel Gal(β1,4)Gal(β1,4)Fuc(α1,6)-core modification attached to the proximal N-acetylglucosamine of keyhole limpet haemocyanin (KLH) N-glycans. Biochem. J. 2004, 378, 625–632. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Iwasa, T.; Tsuda, M.; Kobata, A.; Takasaki, S. A novel monoantennary complex-type sugar chain found in octopus rhodopsin: Occurrence of the Gal beta 1-4Fuc group linked to the proximal N-acetylglucosamine residue of the trimannosyl core. Glycobiology 1997, 7, 1153–1158. [Google Scholar] [CrossRef] [Green Version]
- Gielens, C.; Idakieva, K.; Van den Bergh, V.; Siddiqui, N.I.; Parvanova, K.; Compernolle, F. Mass spectral evidence for N-glycans with branching on fucose in a molluscan hemocyanin. Biochem. Biophys. Res. Commun. 2005, 331, 562–570. [Google Scholar] [CrossRef]
- Sandra, K.; Dolashka-Angelova, P.; Devreese, B.; Van Beeumen, J. New insights in Rapana venosa hemocyanin N-glycosylation resulting from on-line mass spectrometric analyses. Glycobiology 2007, 17, 141–156. [Google Scholar] [CrossRef] [Green Version]
- Dolashka, P.; Velkova, L.; Shishkov, S.; Kostova, K.; Dolashki, A.; Dimitrov, I.; Atanasov, B.; Devreese, B.; Voelter, W.; Van Beeumen, J. Glycan structures and antiviral effect of the structural subunit RvH2 of Rapana hemocyanin. Carbohydr. Res. 2010, 345, 2361–2367. [Google Scholar] [CrossRef] [PubMed]
- Velkova, L.; Dolashka, P.; Lieb, B.; Dolashki, A.; Voelter, W.; Van Beeumen, J.; Devreese, B. Glycan structures of the structural subunit (HtH1) of Haliotis tuberculata hemocyanin. Glycoconj. J. 2011, 28, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Lehr, T.; Geyer, H.; Maass, K.; Doenhoff, M.J.; Geyer, R. Structural characterization of N-glycans from the freshwater snail Biomphalaria glabrata cross-reacting with Schistosoma mansoni glycoconjugates. Glycobiology 2007, 17, 82–103. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, N.I.; Yigzaw, Y.; Préaux, G.; Gielens, C. Involvement of glycans in the immunological cross-reaction between alpha-macroglobulin and hemocyanin of the gastropod Helix pomatia. Biochimie 2009, 91, 508–516. [Google Scholar] [CrossRef]
- Velkova, L.; Dolashka, P.; Van Beeumen, J.; Devreese, B. N-glycan structures of beta-HlH subunit of Helix lucorum hemocyanin. Carbohydr. Res. 2017, 449, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Dolashka, P.; Daskalova, A.; Dolashki, A.; Voelter, W. De Novo Structural Determination of the Oligosaccharide Structure of Hemocyanins from Molluscs. Biomolecules 2020, 10, 1470. [Google Scholar] [CrossRef]
- Dolashka, P.; Velkova, L.; Iliev, I.; Beck, A.; Dolashki, A.; Yossifova, L.; Toshkova, R.; Voelter, W.; Zacharieva, S. Antitumor activity of glycosylated molluscan hemocyanins via Guerin ascites tumor. Immunol. Investig. 2011, 40, 130–149. [Google Scholar] [CrossRef]
- Siddiqui, N.I.; Idakieva, K.; Demarsin, B.; Doumanova, L.; Compernolle, F.; Gielens, C. Involvement of glycan chains in the antigenicity of Rapana thomasiania hemocyanin. Biochem. Biophys. Res. Commun. 2007, 361, 705–711. [Google Scholar] [CrossRef]
- Salazar, M.L.; Jiménez, J.M.; Villar, J.; Rivera, M.; Báez, M.; Manubens, A.; Becker, M.I. N-Glycosylation of mollusk hemocyanins contributes to their structural stability and immunomodulatory properties in mammals. J. Biol. Chem. 2019, 294, 19546–19564. [Google Scholar] [CrossRef] [Green Version]
- Georgieva, A.; Todorova, K.; Iliev, I.; Dilcheva, V.; Vladov, I.; Petkova, S.; Toshkova, R.; Velkova, L.; Dolashki, A.; Dolashka, P. Hemocyanins from Helix and Rapana snails exhibit in vitro antitumor effects in human colorectal adenocarcinoma. Biomedicines 2020, 8, 194. [Google Scholar] [CrossRef] [PubMed]
- Palacios, M.; Tampe, R.; Del Campo, M.; Zhong, T.Y.; López, M.N.; Salazar-Onfray, F.; Becker, M.I. Antitumor activity and carrier properties of novel hemocyanins coupled to a mimotope of GD2 ganglioside. Eur. J. Med. Chem. 2018, 150, 74–86. [Google Scholar] [CrossRef]
- Harris, J.R.; Markl, J. Keyhole limpet hemocyanin (KLH): A biomedical review. Micron 1999, 30, 597–623. [Google Scholar] [CrossRef]
- Pizarro-Bauerle, J.; Maldonado, I.; Sosoniuk-Roche, E.; Vallejos, G.; López, M.N.; Salazar-Onfray, F.; Aguilar-Guzmán, L.; Valck, C.; Ferreira, A.; Becker, M.I. Molluskan hemocyanins activate the classical pathway of the human complement system through natural antibodies. Front. Immunol. 2017, 8, 188. [Google Scholar] [CrossRef]
- Gutternigg, M.; Ahrer, K.; Grabher-Meier, H.; Bürgmayr, S.; Staudacher, E. Neutral N-glycans of the gastropod Arion lusitanicus. Eur. J. Biochem. 2004, 271, 1348–1356. [Google Scholar] [CrossRef] [PubMed]
- Gutternigg, M.; Bürgmayr, S.; Pöltl, G.; Rudolf, J.; Staudacher, E. Neutral N-glycan patterns of the gastropods Limax maximus, Cepaea hortensis, Planorbarius corneus, Arianta arbustorum and Achatina fulica. Glycoconj. J. 2007, 24, 475–489. [Google Scholar] [CrossRef]
- Di Patrizi, L.; Capone, A.; Focarelli, R.; Rosati, F.; Gallego, R.G.; Gerwig, G.J.; Vliegenthart, J.F. Structural characterization of the N-glycans of gp273, the ligand for sperm-egg interaction in the mollusk bivalve Unio elongatulus. Glycoconj. J. 2001, 18, 511–518. [Google Scholar] [CrossRef]
- Park, Y.; Zhang, Z.; Laremore, T.N.; Li, B.; Sim, J.S.; Im, A.R.; Ahn, M.Y.; Kim, Y.S.; Linhardt, R.J. Variation of acharan sulfate and monosaccharide composition and analysis of neutral N-glycans in African giant snail (Achatina fulica). Glycoconj. J. 2008, 25, 863–877. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, N.; Masuda, K.; Hiraki, K.; Yoshihara, K.; Huang, H.-H.; Khoo, K.-H.; Kato, K. N-Glycan structures of rhodopsin. Existence of the α1,3 and α1,6 difucosylated innermost GlcNAc residue in a molluscan glycoprotein. Eur. J. Biochem. 2003, 270, 2617–2632. [Google Scholar] [CrossRef]
- Eckmair, B.; Jin, C.; Abed-Navandi, D.; Paschinger, K. Multistep fractionation and mass spectrometry reveal zwitterionic and anionic modifications of the N- and O-glycans of a marine snail. Mol. Cell Proteomics 2016, 15, 573–597. [Google Scholar] [CrossRef] [Green Version]
- Milanese, C.; Fiumara, F.; Bizzoca, A.; Giachello, C.; Leitinger, G.; Gennarini, G.; Montarolo, P.G.; Ghirardi, M. F3/contactin-related proteins in Helix pomatia nervous tissue (HCRPs): Distribution and function in neurite growth and neurotransmitter release. J. Neurosci. Res. 2008, 86, 821–831. [Google Scholar] [CrossRef] [Green Version]
- Silverman-Gavrila, L.B.; Senzel, A.G.; Charlton, M.P.; Feng, Z.P. Expression, phosphorylation, and glycosylation of CNS proteins in aversive operant conditioning associated memory in Lymnaea stagnalis. Neuroscience 2011, 186, 94–109. [Google Scholar] [CrossRef]
- Mitta, G.; Gourbal, B.; Grunau, C.; Knight, M.; Bridger, J.M.; Théron, A. The compatibility between Biomphalaria glabrata snails and Schistosoma mansoni: An increasingly complex puzzle. Adv. Parasitol. 2016, 97, 111–145. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.-J.; Dinguirard, N.; Sabat, G.; Lui, H.; Gonzalez, L.; Gehring, M.; Bickham-Wright, U.; Yoshino, T.P. Proteomic analysis of Biomphalaria glabrata plasma proteins with binding affinity to those expressed by early developing larval Schistosoma mansoni. PLOS Pathoges 2017, 13, e1006081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Famakinde, D.O. Molecular context of Schistosoma mansoni transmission in the molluscan environments: A mini-review. Acta Trop. 2017, 176, 98–104. [Google Scholar] [CrossRef]
- Odoemelan, E.; Raghavan, N.; Miller, A.; Bridger, J.M.; Knight, M. Revised karyotyping and gene mapping of the Biomphalaria glabrata embryonic (Bge) cell line. Int. J. Parasitol. 2009, 39, 675–681. [Google Scholar] [CrossRef] [Green Version]
- Martins -Souza, R.L.; Pereira, C.A.J.; Martins Filho, O.A.; Coelho, P.M.Z.; Corrêa Jr., A.; Negrão- Corrêa, D. Differential lectin labelling of circulating hemocytes from Biomphalaria glabrata and Biomphalaria tenagophila resistant or susceptible to Schistosoma mansoni infection. Mem. Inst. Oswaldeo Cruz 2006, 101, 185–192. [Google Scholar] [CrossRef]
- Yoshino, T.P.; Wu, X.J.; Liu, H.; Gonzalez, L.A.; Deelder, A.M.; Hokke, C.H. Glycotope sharing between snail hemolymph and larval schistosomes: Larval transformation products alter shared glycan patterns of plasma proteins. PLoS Negl. Trop. Dis. 2012, 6, e1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshino, T.P.; Wu, X.J.; Gonzalez, L.A.; Hokke, C.H. Circulating Biomphalaria glabrata hemocyte subpopulations possess shared schistosome glycans and receptors capable of binding larval glycoconjugates. Exp. Parasitol. 2013, 133, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Mansour, M.H. Evidence for a family of schistosome glycan-binding lectins in Biomphalaria alexandrina. Dev. Comp. Immunol. 1995, 19, 365–376. [Google Scholar] [CrossRef]
- Mansour, M.H.; Abdul-Salam, F. Characterization of fucose-binding lectins in rock- and mud-dwelling snails inhabiting Kuwait Bay. Immunobiology 2009, 214, 77–85. [Google Scholar] [CrossRef]
- Yoshino, T.P.; Dinguirard, N.; Kunert, J.; Hokke, C.H. Molecular and functional characterization of a tandem-repeat galectin from the freshwater snail Biomphalaria glabrata, intermediate host of the human blood fluke Schistosoma mansoni. Gene 2008, 411, 46–58. [Google Scholar] [CrossRef] [Green Version]
- Lehr, T.; Beuerlein, K.; Doenhoff, M.J.; Grevelding, C.G.; Geyer, R. Localization of carbohydrate determinants common to Biomphalaria glabrata as well as to sporocysts and miracidia of Schistosoma mansoni. Parasitology 2008, 135, 931–942. [Google Scholar] [CrossRef]
- Castillo, M.G.; Wu, X.J.; Dinguirard, N.; Nyame, A.K.; Cummings, R.D.; Yoshino, T.P. Surface membrane proteins of Biomphalaria glabrata embryonic cells bind fucosyl determinants on the tegumental surface of Schistosoma mansoni primary sporocysts. J. Parasitol. 2007, 93, 832–840. [Google Scholar] [CrossRef]
- Lehr, T.; Frank, S.; Natsuka, S.; Geyer, H.; Beuerlein, K.; Doenhoff, M.J.; Hase, S.; Geyer, R. N-Glycosylation patterns of hemolymph glycoproteins from Biomphalaria glabrata strains expressing different susceptibility to Schistosoma mansoni infection. Exp. Parasitol. 2010, 126, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Faltýnková, A.; Nasincová, V.; Kablásková, L. Larval trematodes (Digenea) of the great pond snail, Lymnaea stagnalis (L.), (Gastropoda, Pulmonata) in Central Europe: A survey of species and key to their identification. Parasite 2007, 14, 39–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davison, A.; Blaxter, M.L. An expressed sequence tag survey of gene expression in the pond snail Lymnaea stagnalis, an intermediate vector of Fasciola hepatica. Parasitology 2005, 130, 539–552. [Google Scholar] [CrossRef] [Green Version]
- Plows, L.D.; Cook, R.T.; Davies, A.J.; Walker, A.J. Carbohydrates that mimic schistosome surface coat components affect ERK and PKC signaling in Lymnaea stagnalis haemocytes. Int. J. Parasitol. 2005, 35, 293–302. [Google Scholar] [CrossRef]
- Georgieva, K.; Georgieva, L.; Mizinska-Boevska, Y.; Stoitsova, S.R. Study of surface carbohydrates in Galba truncatula tissues before and after infection with Fasciola hepatica. Memórias Inst. Oswaldo Cruz 2016, 111, 475–483. [Google Scholar] [CrossRef]
- Feng, C.; Ghosh, A.; Amin, M.N.; Giomarelli, B.; Shridhar, S.; Banerjee, A.; Fernández-Robledo, J.A.; Bianchet, M.A.; Wang, L.-X.; Wilson, I.B.H.; et al. The galectin CvGal1 from Eastern oyster (Crassostrea virginica) binds to blood group A oligosaccharides on the hemocyte surface. J. Biol. Chem. 2013, 288, 24394–24409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tasumi, S.; Vasta, G.R. A galectin of unique domain organization from hemocytes of the Eastern oyster (Crassostrea virginica) is a receptor for the protistan parasite Perkinsus marinus. J. Immunol. 2007, 179, 3086–3098. [Google Scholar] [CrossRef] [Green Version]
- Feng, C.; Ghosh, A.; Amin, M.N.; Bachvaroff, T.R.; Tasumi, S.; Pasek, M.; Banerjee, A.; Shridhar, S.; Wang, L.-X.; Bianchet, M.A.; et al. Galectin CvGal2 from the Eastern oyster (Crassostrea virginica) displays unique specificity for ABH blood group oligosaccharides and differentially recognizes sympatric Perkinsus species. Biochemistry 2015, 54, 4711–4730. [Google Scholar] [CrossRef] [Green Version]
- Kurz, S.; Jin, C.; Hykollari, A.; Gregorich, D.; Giomarelli, B.; Vasta, G.R.; Wilson, I.B.; Paschinger, K. Hemocytes and plasma of the eastern oyster (Crassostrea virginica) display a diverse repertoire of sulfated and blood group A-modified N-glycans. J. Biol. Chem. 2013, 288, 24410–24428. [Google Scholar] [CrossRef] [Green Version]
- Thiengo, S.C.; Fernandez, M.A.; Torres, E.J.L.; Coelho, P.M.; Lanfredi, R.M. First record of a nematode Metastrongyloidea (Aelurostrongylus abstrusus larvae) in Achatina (Lissachatina) fulica (Mollusca, Achatinidae) in Brazil. J. Invertebr. Pathol. 2008, 98, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Geyer, H.; Wuhrer, M.; Resemann, A.; Geyer, R. Identification and characterization of keyhole limpet hemocyanin N-glycans mediating cross-reactivity with Schistosoma mansoni. J. Biol. Chem. 2005, 280, 40731–40748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Fang, X.; Guo, X.; Li, L.; Luo, R.; Xu, F.; Yang, P.; Zhang, L.; Wang, X.; Qi, H.; et al. The oyster genome reveals stress adaptation and complexity of shell formation. Nature 2012, 490, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Adema, C.M.; Hillier, L.W.; Jones, C.S.; Loker, E.S.; Knight, M.; Minx, P.; Oliveira, G.; Raghavan, N.; Shedlock, A.; do Amaral, L.R.; et al. Whole genome analysis of a schistosomiasis-transmitting freshwater snail. Nat. Commun. 2017, 16, 15451. [Google Scholar] [CrossRef]
- Wheeler, N.J.; Dinguirard, N.; Marquez, J.; Gonzalez, A.; Zamanian, M.; Yoshino, T.P.; Castillo, M.G. Sequence and structural variation in the genome of the Biomphalaria glabrata embryonic (Bge) cell line. Parasites Vectors 2018, 11, 496. [Google Scholar] [CrossRef] [PubMed]
- Mulder, H.; Dideberg, F.; Schachter, H.; Spronk, B.A.; De Jong-Brink, M.; Kamerling, J.P.; Vliegenthart, J.F. In the biosynthesis of N-glycans in connective tissue of the snail Lymnaea stagnalis of incorporation GlcNAc by β2GlcNAc-transferase I is an essential prerequisite for the action of β2GlcNAc-transferase II and β2Xyl-transferase. Eur. J. Biochem. 1995, 232, 272–283. [Google Scholar] [CrossRef]
- Mulder, H.; Spronk, B.A.; Schachter, H.; Neeleman, A.P.; van den Eijnden, D.; De Jong-Brink, M.; Kamerling, J.P.; Vliegenthart, J.F.G. Identification of a novel UDP-GalNAc:GlcNAcβ-R β1,4 N-acetylgalactosaminyltransferase from the albumen gland and connective tissue of the snail Lymnaea stagnalis. Eur. J. Biochem. 1995, 227, 175–185. [Google Scholar] [CrossRef]
- Neeleman, A.P.; van den Eijnden, D.H. α-Lactalbumin affects the acceptor specificity of Lymnaea stagnalis albumen gland UDP-GalNAc:GlcNAcβ-R β1,4-N-acetylgalactosaminyltransferase: Synthesis of GalNAcβ1,4Glc. Proc. Natl. Acad. Sci. USA 1996, 93, 10111–10116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulder, H.; Schachter, H.; De Jong-Brink, M.; Van der Ven, J.G.; Kamerling, J.P.; Vliegenthart, J.F. Identification of a novel UDP-Gal:GalNAc β1,4GlcNAc-R β1,3-galactosyltransferase in the connective tissue of the snail Lymnaea stagnalis. Eur. J. Biochem. 1991, 201, 459–465. [Google Scholar] [CrossRef]
- Mulder, H.; Schachter, H.; Thomas, J.R.; Halkes, K.M.; Kamerling, J.P.; Vliegenthart, J.F.G. Identification of a GDP-Fuc:Galβ1,3GalNAc-R(Fuc to Gal) α1,2 fucosyltransferase and a GDP-Fuc:Galβ1,4GlcNAc (Fuc to GlcNAc) α1,3 fucosyltransferase in connective tissue of the snail Lymnaea stagnalis. Glycoconj. J. 1996, 13, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Van Tetering, A.; Schiphorst, W.; van den Eijnden, D.H.; van Die, I. Characterization of a core alpha1-->3-fucosyltransferase from the snail Lymnaea stagnalis that is involved in the synthesis of complex-type N-glycans. FEBS Lett. 1999, 461, 311–314. [Google Scholar] [CrossRef] [Green Version]
- Bakker, H.; Agterberg, M.; Van Tetering, A.; Koeleman, C.A.; Van den Eijnden, D.H.; Van Die, I. A Lymnaea stagnalis gene, with sequence similarity to that of mammalian beta 1-->4-galactosyltransferases, encodes a novel UDP-GlcNAc:GlcNAc beta-R beta 1-->4-N-acetylglucosaminyltransferase. J. Biol. Chem. 1994, 269, 30326–30333. [Google Scholar] [CrossRef]
- Van Die, I.; Cummings, R.D.; van Tetering, A.; Hokke, C.H.; Koeleman, C.A.; van den Eijnden, D.H. Identification of a novel UDP-Glc:GlcNAc beta1-->4-glucosyltransferase in Lymnaea stagnalis that may be involved in the synthesis of complex-type oligosaccharide chains. Glycobiology 2000, 10, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Song, H.-B.; He, M.; Cai, Z.-P.; Huang, K.; Flitsch, S.L.; Liu, L.; Voglmeir, J. UDP-Glucose 4-epimerase and β-1,4-galactosyltransferase from the oyster Magallana gigas as valuable biocatalysts for the production of galactosylated products. Int. J. Mol. Sci. 2018, 19, 1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robledo, Y.; Marigómez, I.; Angulo, E.; Cajaraville, M.P. Glycosylation and sorting pathways of lysosomal enzymes in mussel digestive cells. Cell Tissue Res. 2006, 324, 319–333. [Google Scholar] [CrossRef]
- Hu, Y.; Luan, H.; Liu, H.; Ge, G.; Zhou, K.; Liu, Y.; Yang, L. Acceptor specificity and transfer efficiency of a beta-glycosidase from the Chinese white jade snail. Biosci. Biotechnol. Biochem. 2009, 73, 671–676. [Google Scholar] [CrossRef] [Green Version]
- Biswas, C.; Sinha, D.; Mandal, C. Investigation on interaction of Achatinin, a 9-O-acetyl sialic acid-binding lectin, with lipopolysaccharide in the innate immunity of Achatina fulica snails. Mol. Immunol. 2000, 37, 745–754. [Google Scholar] [CrossRef]
- Brossmer, R.; Wagner, M.; Fischer, E. Specificity of the sialic acid-binding lectin from the snail Cepaea hortensis. J. Biol. Chem. 1992, 267, 8752–8756. [Google Scholar] [CrossRef]
- Gerlach, D.; Schlott, B.; Schmidt, K.H. Cloning and expression of a sialic acid-binding lectin from the snail Cepaea hortensis. FEMS Immunol. Med. Microbiol. 2004, 40, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Miller, R.L.; Collawn, J.F., Jr.; Fish, W.W. Purification and macromolecular properties of a sialic acid-specific lectin from the slug Limax flavus. J. Biol. Chem. 1982, 257, 7574–7580. [Google Scholar] [CrossRef]
- García-Maldonado, E.; Cano-Sánchez, P.; Hernández-Santoyo, A. Molecular and functional characterization of a glycosylated Galactose-Binding lectin from Mytilus californianus. Fish Shellfish Immunol. 2017, 66, 564–574. [Google Scholar] [CrossRef]
- He, X.; Zhang, Y.; Yu, F.; Yu, Z. A novel sialic acid binding lectin with anti-bacterial activity from the Hong Kong oyster (Crassostrea hongkongensis). Fish Shellfish Immunol. 2011, 31, 1247–1250. [Google Scholar] [CrossRef]
- Ito, S.; Shimizu, M.; Nagatsuka, M.; Kitajima, S.; Honda, M.; Tsuchiya, T.; Kanzawa, N. High molecular weight lectin isolated from the mucus of the Giant African Snail Achatina fulica. Biosci. Biotechnol. Biochem. 2011, 75, 20–25. [Google Scholar] [CrossRef] [Green Version]
- Brola, T.R.; Dreon, M.S.; Qiu, J.W.; Heras, H. A highly stable, non-digestible lectin from Pomacea diffusa unveils clade-related protection systems in apple snail eggs. J. Exp. Biol. 2020, 223, jeb231878. [Google Scholar] [CrossRef]
- Wang, W.; Song, X.; Wang, L.; Song, L. Pathogen-derived carbohydrate recognition in molluscs immune defense. Int. J. Mol. Sci. 2018, 19, 721. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, U.; Adam, E.; Brooks, S.A.; Leathem, A.J. Lectin-binding properties of human breast cancer cell lines and human milk with particular reference to Helix pomatia agglutinin. J. Histochem. Cytochem. 1995, 43, 275–281. [Google Scholar] [CrossRef] [Green Version]
- Rambaruth, N.D.S.; Greenwell, P.; Dwek, M.V. The lectin Helix pomatia agglutinin recognizes O-GlcNAc containing glycoproteins in human breast cancer. Glycobiology 2012, 22, 839–848. [Google Scholar] [CrossRef] [Green Version]
- Pietrzyk, A.J.; Bujacz, A.; Mak, P.; Potempa, B.; Niedziela, T. Structural studies of Helix aspersa agglutinin complexed with GalNAc: A lectin that serves as a diagnostic tool. Int. J. Biol. Macromol. 2015, 81, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.M.; Suzuki, H.; Brooks, M.T.; Tomana, M.; Moldoveanu, Z.; Mestecky, J.; Julian, B.A.; Novak, J.; Herr, A.B. Recognition of galactose-deficient O-glycans in the hinge region of IgA1 by N-acetylgalactosamine-specific snail lectins: A comparative binding study. Biochemistry 2010, 49, 5671–5682. [Google Scholar] [CrossRef] [Green Version]
- Markiv, A.; Peiris, D.; Curley, G.P.; Odell, M.; Dwek, M.V. Identification, cloning, and characterization of two N-acetylgalactosamine-binding lectins from the albumen gland of Helix pomatia. J. Biol. Chem. 2011, 286, 20260–20266. [Google Scholar] [CrossRef] [Green Version]
- Sudhakar, G.R.L.; Vincent, S.G.P. Purification and characterization of a novel C-type lectin for clot lysis from the fresh water clam Villorita cyprinoides: A possible natural thrombolytic agent against myocardial infarction. Fish Shellfish Immunol. 2015, 36, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.G.; Zafaralla, G.; Cruz, L.J.; Santos, A.D.; Hillyard, D.R.; Dykert, J.; Rivier, J.E.; Gray, W.R.; Imperial, J.; DelaCruz, R.G.; et al. An O-glycosylated neuroexcitatory conus peptide. Biochemistry 1998, 37, 16019–16025. [Google Scholar] [CrossRef]
- Hocking, H.G.; Gerwig, G.J.; Dutertre, S.; Violette, A.; Favreau, P.; Stöcklin, R.; Kamerling, J.P.; Boelens, R. Structure of the O-glycosylated conopeptide CcTx from Conus consors venom. Chemistry 2013, 19, 870–879. [Google Scholar] [CrossRef]
- Gerwig, G.J.; Hocking, H.G.; Stöcklin, R.; Kamerling, J.P.; Boelens, R. Glycosylation of conotoxins. Mar. Drugs 2013, 11, 623–642. [Google Scholar] [CrossRef] [PubMed]
- Tsetlin, V.I.; Hucho, F. Snake and snail toxins acting on nicotinic acetylcholine receptors: Fundamental aspects and medical applications. FEBS Lett. 2004, 557, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.K.; Zhang, L.; Smith, M.D.; Walewska, A.; Vellore, N.A.; Baron, R.; McIntosh, J.M.; White, H.S.; Olivera, B.M.; Bulaj, G. A marine analgesic peptide, Contulakin-G, and neurotensin are distinct agonists for neurotensin receptors: Uncovering structural determinants of desensitization properties. Front. Pharmacol. 2015, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, E.C. Post-translationally modified conopeptides: Biological activities and pharmacological applications. Peptides 2021, 139, 170525. [Google Scholar] [CrossRef]
Figure 1.
Shell matrix protein N-glycans of (a) Biomphalaria glabrata [15]; (b) Mytilus edulis [16]; (c) Pincatada fucata [17]. The structure plots were generated in the notation of the Consortium for Functional Glycomics (http://www.functionalglycomics.org, accessed on 16 November 2021). All figures were drawn especially for this review, based on previously published data as cited.
Figure 1.
Shell matrix protein N-glycans of (a) Biomphalaria glabrata [15]; (b) Mytilus edulis [16]; (c) Pincatada fucata [17]. The structure plots were generated in the notation of the Consortium for Functional Glycomics (http://www.functionalglycomics.org, accessed on 16 November 2021). All figures were drawn especially for this review, based on previously published data as cited.

Figure 2.
Examples for hemocyanin N-glycan structures from different molluscs. (a) Helix pomatia [21]; (b) Lymnaea stagnalis [22]; (c) Lymnaea stagnalis [23]; (d) Rapana venosa [27]; (e) Megathura crenulata [28]; (f) Megathura crenulata [29]. For the symbols key see Figure 1.

Figure 3.
Examples for hemocyanin N-glycan structures from different molluscs. (a) 7 RtH2-e of Rapana thomasiana [31]; (b) RvH1 and RvH2 of Rapana venosa [32,33], (c) HtH1 of Haliotis tuberculata [34], (d) Biomphalaria glabrata [35], (e) Helix pomatia [25,36]. For the symbols key see Figure 1.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Staudacher, E. Mollusc N-glycosylation: Structures, Functions and Perspectives. Biomolecules 2021, 11, 1820. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11121820
AMA Style
Staudacher E. Mollusc N-glycosylation: Structures, Functions and Perspectives. Biomolecules. 2021; 11(12):1820. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11121820
Chicago/Turabian StyleStaudacher, Erika. 2021. "Mollusc N-glycosylation: Structures, Functions and Perspectives" Biomolecules 11, no. 12: 1820. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11121820
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.