
NTPDase1 Modulates Smooth Muscle Contraction in Mice Bladder by Regulating Nucleotide Receptor Activation Distinctly in Male and Female

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Enzyme Histochemistry
2.4. Immunohistochemistry
2.5. Nucleotidase Activity Measurement
2.6. Quantitative Real-Time PCR (RT-qPCR)
2.7. Measurement of Bladder Strips Contraction
2.8. Statistical Analysis
3. Results
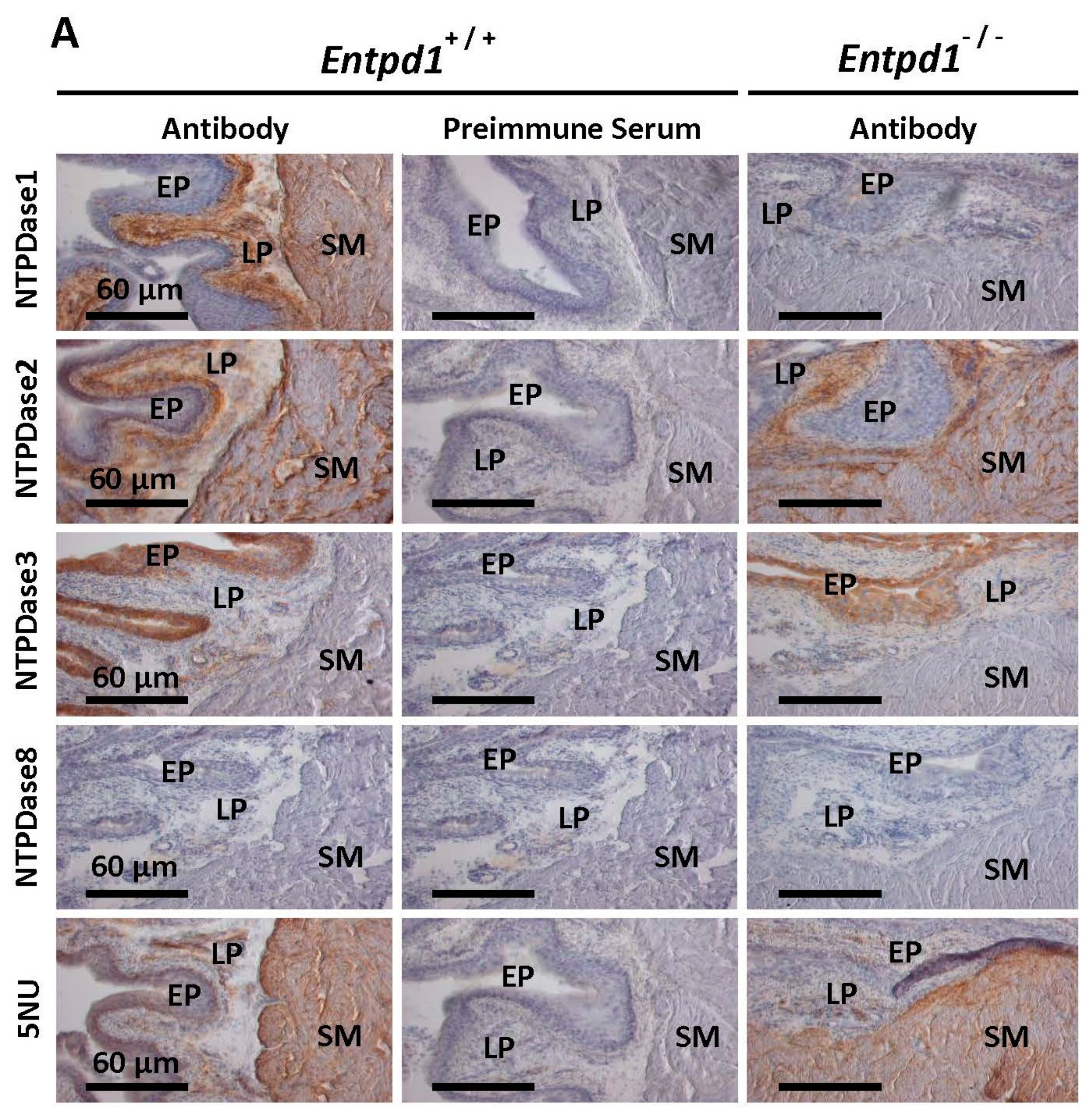
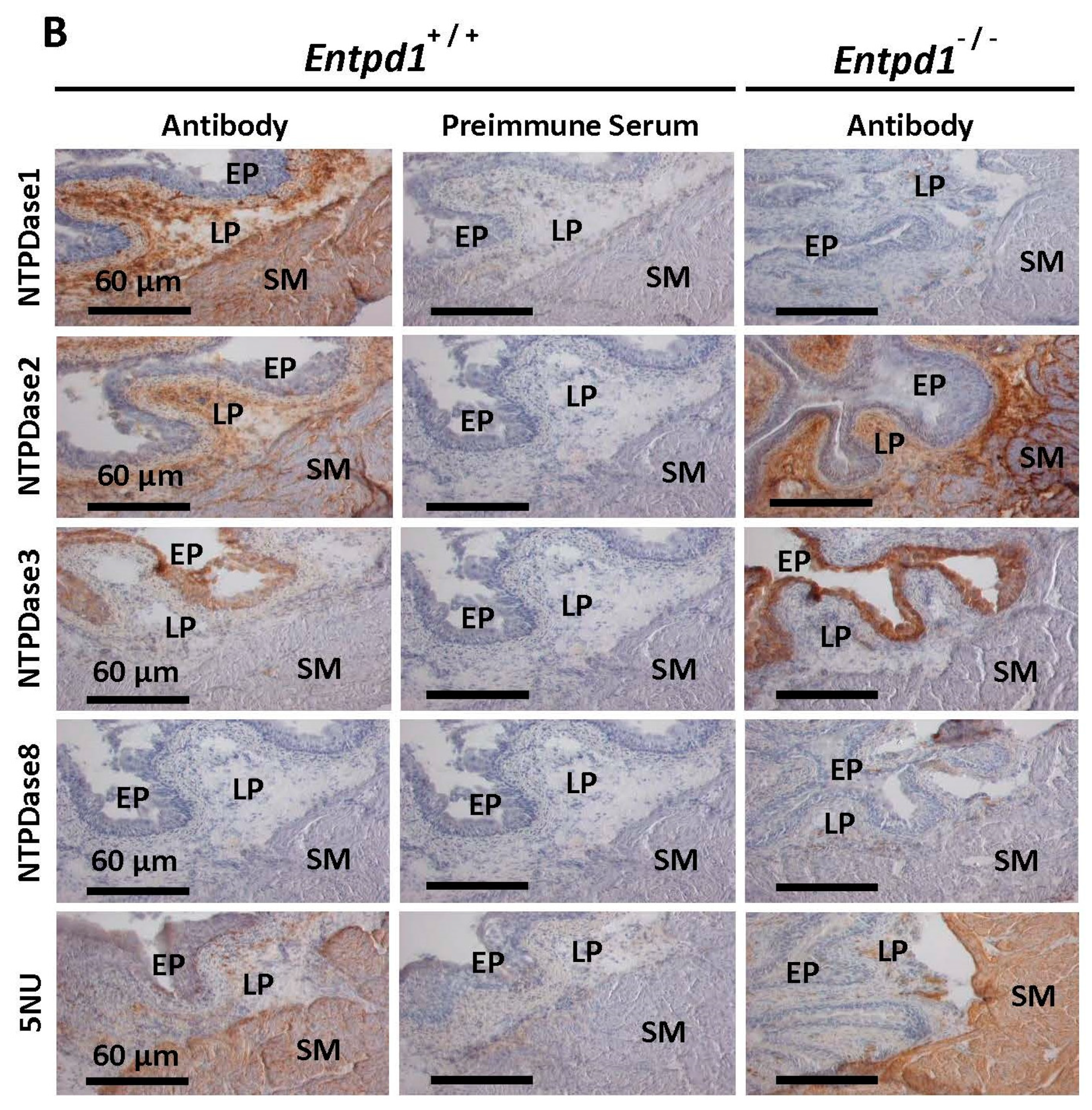
3.1. Immunolocalization and Activity of Ectonucleotidases in Bladder SMCs
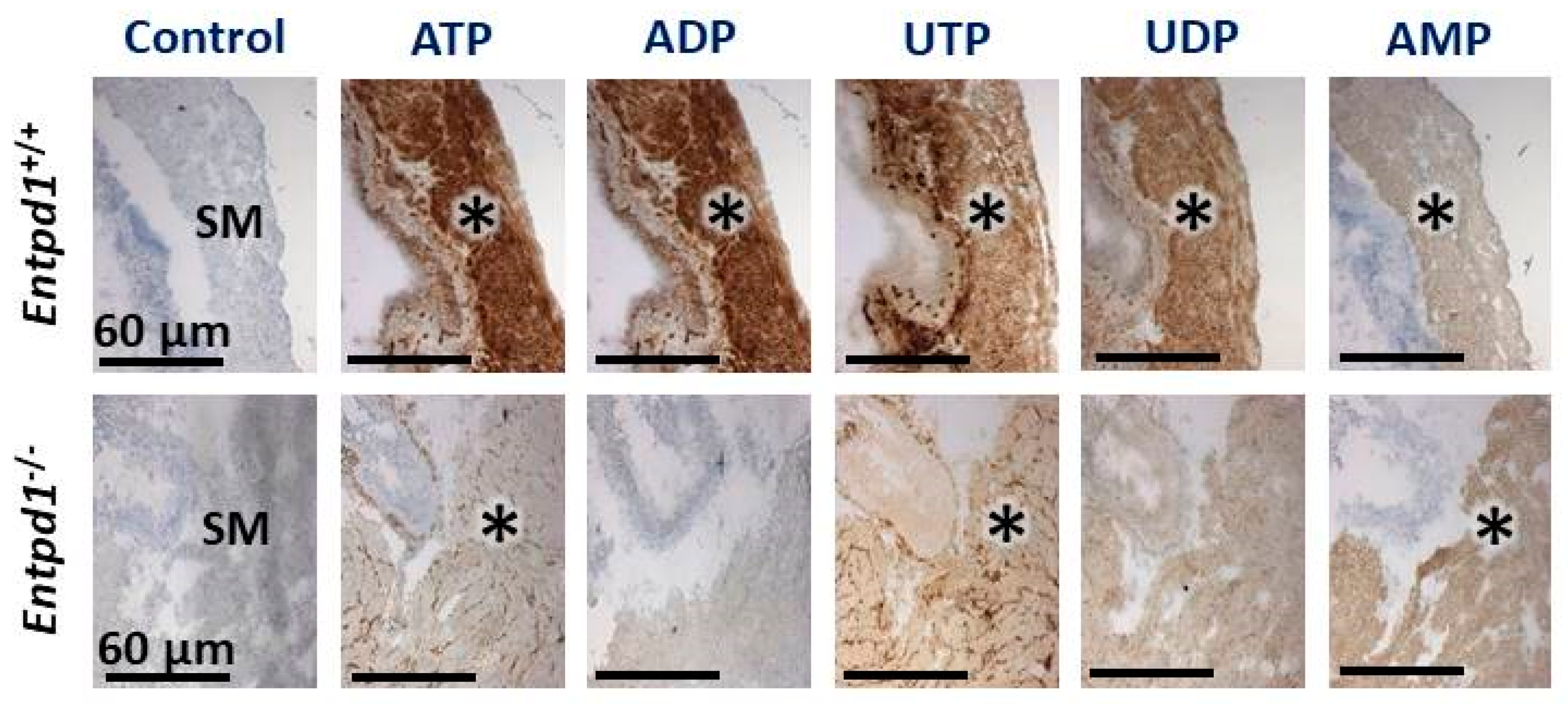
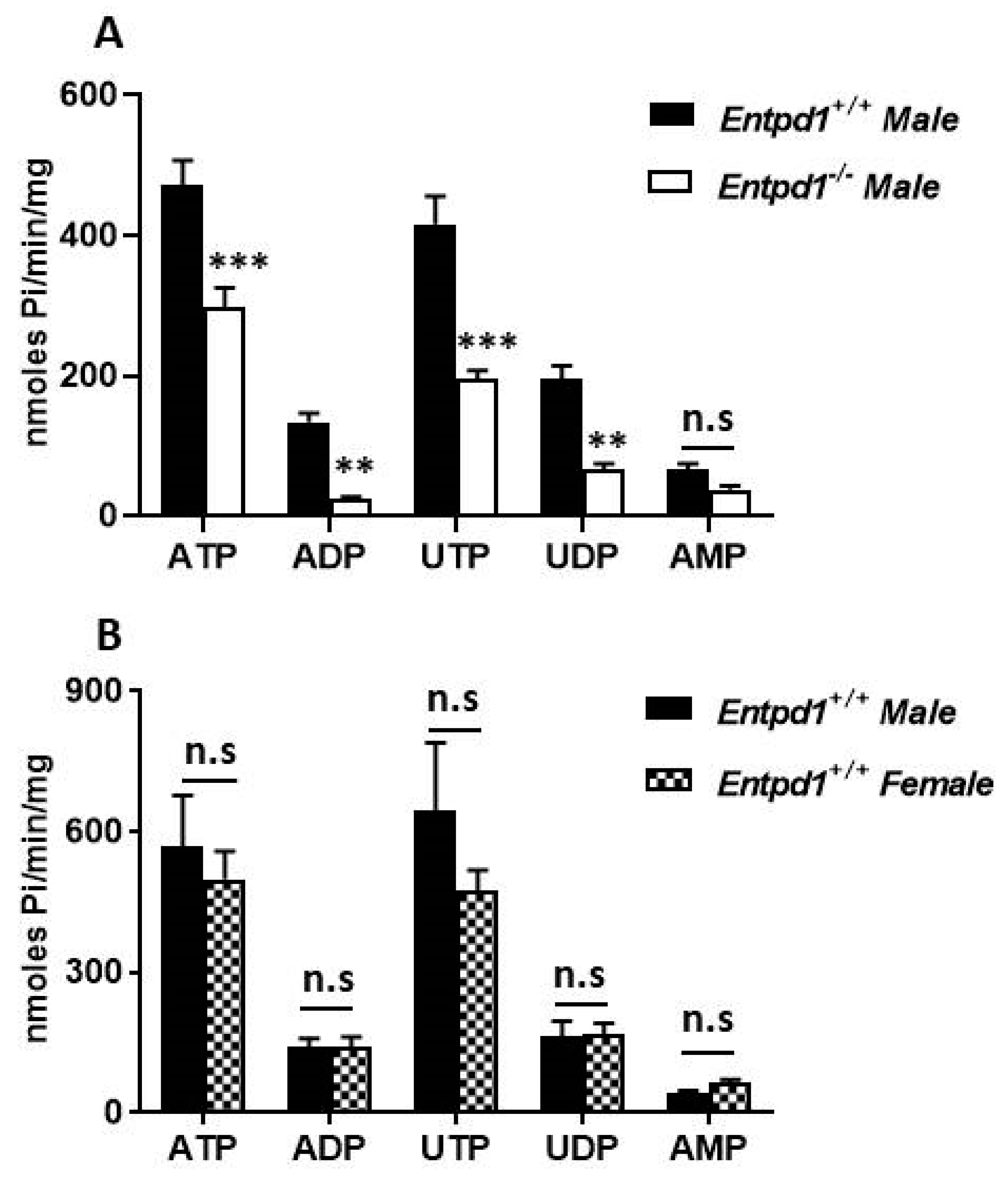
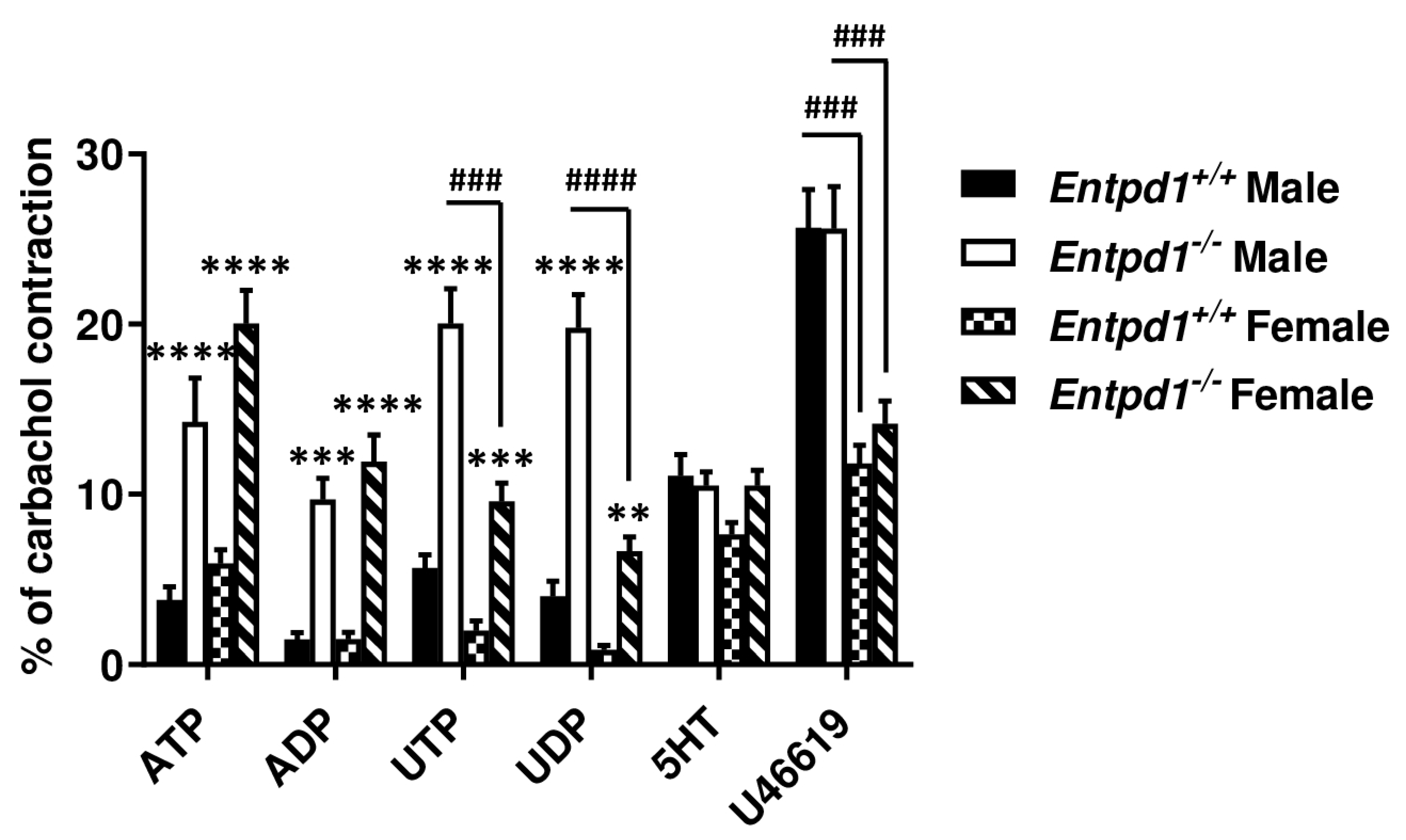
3.2. Contractile Effect of Nucleotides is Increased in Entpd1−/− Bladder Strips
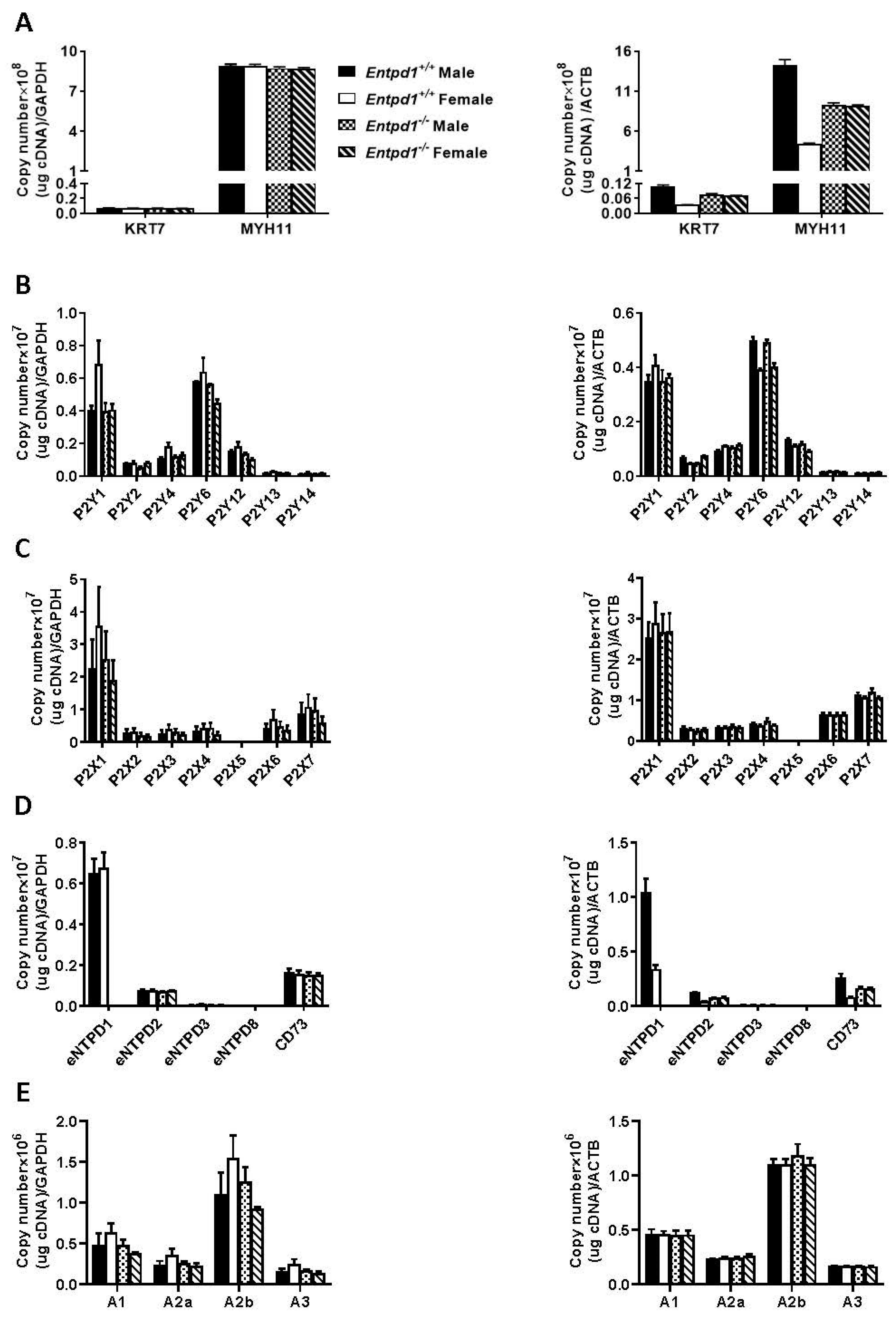
3.3. mRNA Expression of Ectonucleotidases and Nucleotide Receptors in Mouse Bladder Smooth Muscle Layer
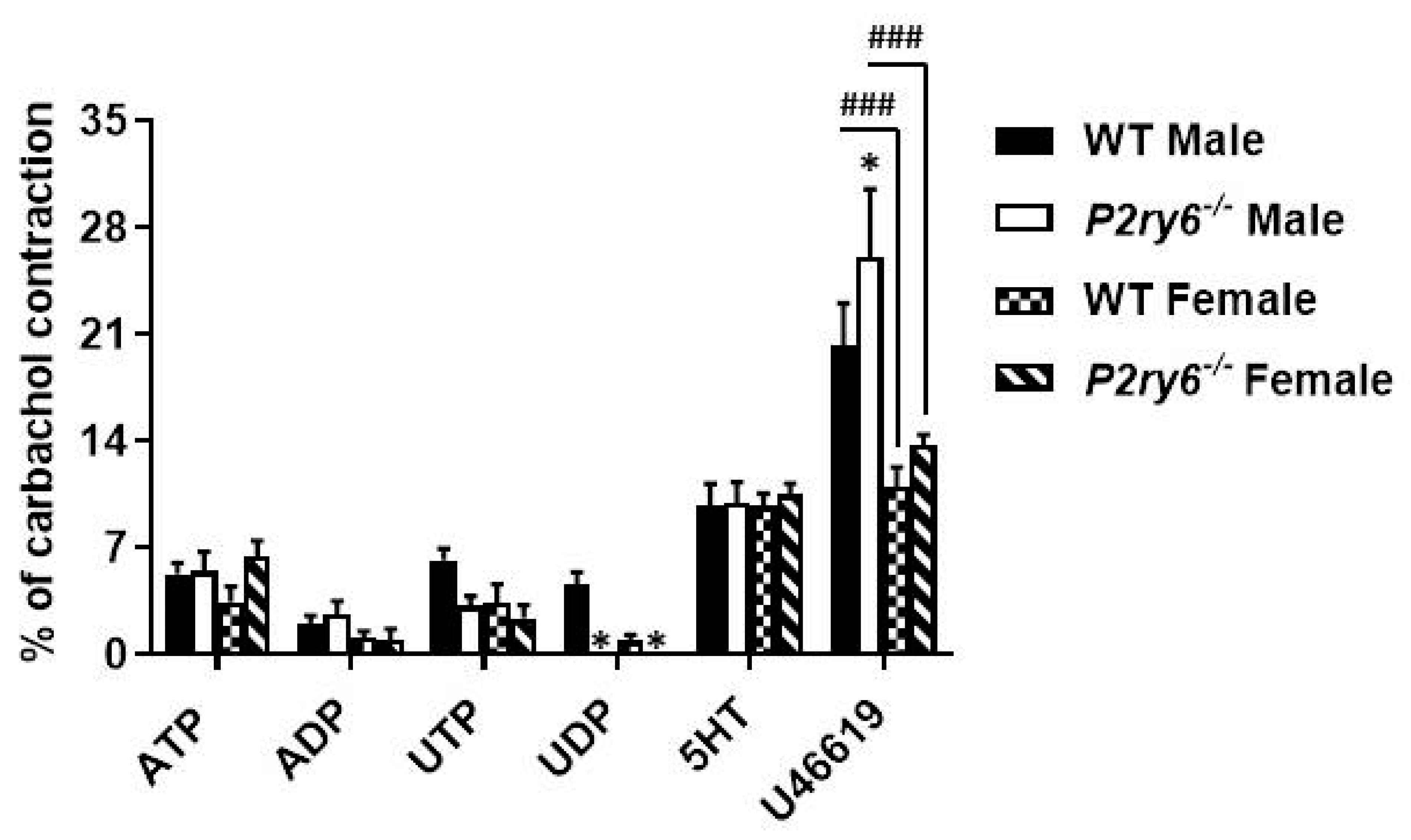
3.4. Implication of P2Y6 Receptor in the Contractile Response of Bladder SMCs
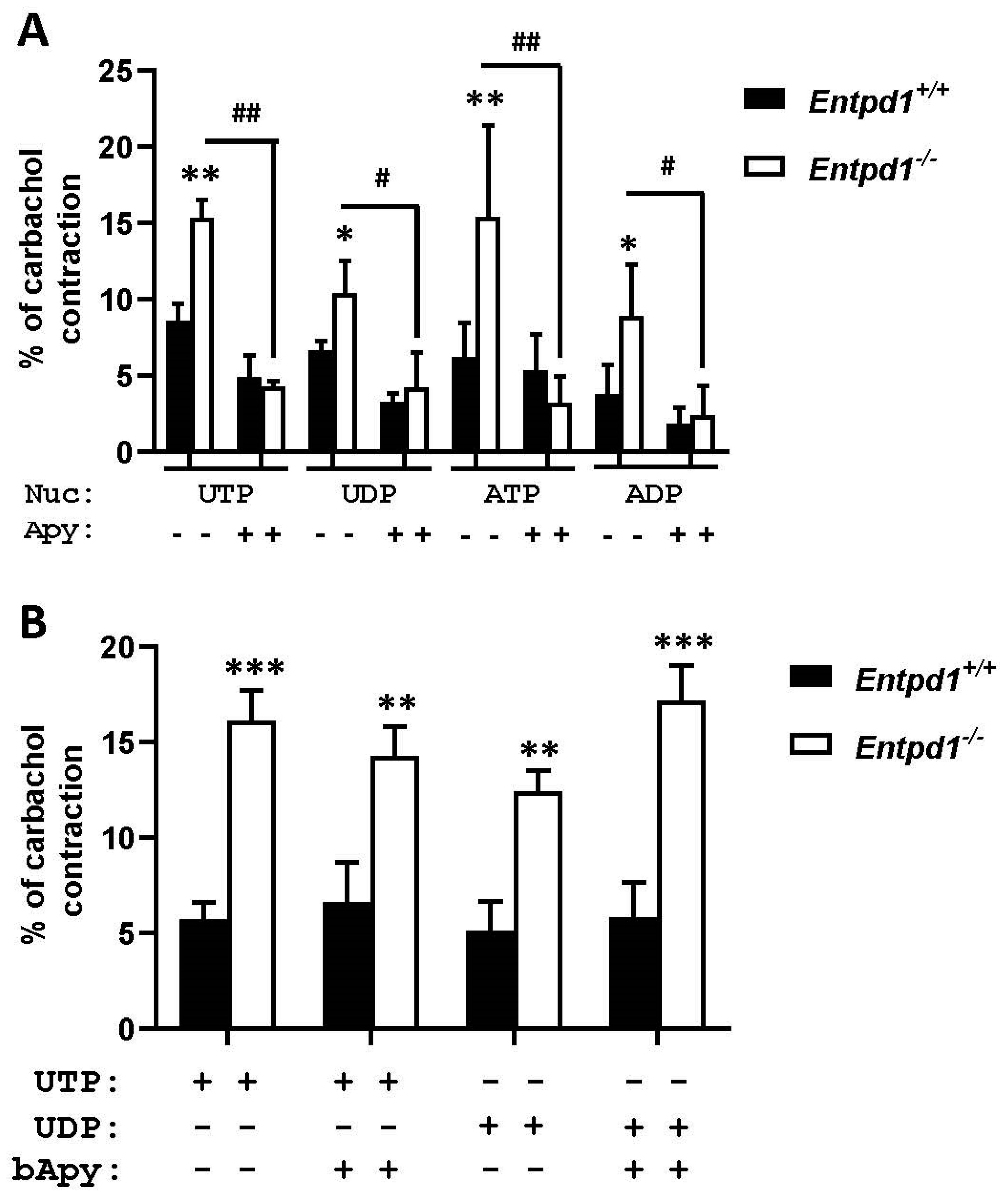
3.5. Apyrase Reversed the Increased Contraction in Entpd1−/− Bladders
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| SMC | Smooth muscle cell |
| ATP | Adenosine 5′-triphosphate |
| ADP | Adenosine 5′-diphosphate |
| UTP | Uridine 5′-triphosphate |
| UDP | Uridine 5′-diphosphate |
| AMP | Adenosine 5′-monophosphate |
| UMP | Uridine 5′-monophosphate |
| RT-qPCR | Quantitative Real time polymerase chain reaction |
| E-NTPDase | Ecto-nucleoside triphosphate diphosphohydrolase |
| OCT | Optimal cutting temperature |
| Pi | Inorganic phosphate |
| SEM | Standard error of mean |
| 5HT | 5-hydroxytryptamine |
| U46619 | Thromboxane A2 analogue |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| ACTB | Actin beta |
References
- Vassort, G. Adenosine 5′-triphosphate: A P2-purinergic agonist in the myocardium. Physiol. Rev. 2001, 81, 767–806. [Google Scholar] [CrossRef] [PubMed]
- Kukulski, F.; Levesque, S.A.; Sevigny, J. Impact of ectoenzymes on p2 and p1 receptor signaling. Adv. Pharmacol. 2011, 61, 263–299. [Google Scholar] [CrossRef] [PubMed]
- Yegutkin, G.G. Nucleotide- and nucleoside-converting ectoenzymes: Important modulators of purinergic signalling cascade. Biochim. Biophys. Acta 2008, 1783, 673–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbracchio, M.P.; Burnstock, G.; Boeynaems, J.M.; Barnard, E.A.; Boyer, J.L.; Kennedy, C.; Knight, G.E.; Fumagalli, M.; Gachet, C.; Jacobson, K.A.; et al. International Union of Pharmacology LVIII: Update on the P2Y G protein-coupled nucleotide receptors: From molecular mechanisms and pathophysiology to therapy. Pharmacol. Rev. 2006, 58, 281–341. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Corriden, R.; Inoue, Y.; Yip, L.; Hashiguchi, N.; Zinkernagel, A.; Nizet, V.; Insel, P.A.; Junger, W.G. ATP release guides neutrophil chemotaxis via P2Y2 and A3 receptors. Science 2006, 314, 1792–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazarowski, E.R.; Boucher, R.C.; Harden, T.K. Mechanisms of release of nucleotides and integration of their action as P2X- and P2Y-receptor activating molecules. Mol. Pharmacol. 2003, 64, 785–795. [Google Scholar] [CrossRef] [Green Version]
- Eltzschig, H.K.; Eckle, T.; Mager, A.; Kuper, N.; Karcher, C.; Weissmuller, T.; Boengler, K.; Schulz, R.; Robson, S.C.; Colgan, S.P. ATP release from activated neutrophils occurs via connexin 43 and modulates adenosine-dependent endothelial cell function. Circ. Res. 2006, 99, 1100–1108. [Google Scholar] [CrossRef] [Green Version]
- Woehrle, T.; Yip, L.; Elkhal, A.; Sumi, Y.; Chen, Y.; Yao, Y.; Insel, P.A.; Junger, W.G. Pannexin-1 hemichannel-mediated ATP release together with P2X1 and P2X4 receptors regulate T-cell activation at the immune synapse. Blood 2010, 116, 3475–3484. [Google Scholar] [CrossRef] [Green Version]
- Kukulski, F.; Bahrami, F.; Ben Yebdri, F.; Lecka, J.; Martín-Satué, M.; Lévesque, S.A.; Sévigny, J. NTPDase1 controls IL-8 production by buman neutrophils. J. Immunol. 2011, 187, 644–653. [Google Scholar] [CrossRef] [Green Version]
- Corriden, R.; Insel, P.A. Basal release of ATP: An autocrine-paracrine mechanism for cell regulation. Sci. Signal. 2010, 3, re1. [Google Scholar] [CrossRef] [Green Version]
- Katsuragi, T.; Tokunaga, T.; Ogawa, S.; Soejima, O.; Sato, C.; Furukawa, T. Existence of ATP-evoked ATP release system in smooth muscles. J. Pharmacol. Exp. Ther. 1991, 259, 513–518. [Google Scholar] [PubMed]
- Bergfeld, G.R.; Forrester, T. Release of ATP from human erythrocytes in response to a brief period of hypoxia and hypercapnia. Cardiovasc. Res. 1992, 26, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Bodin, P.; Burnstock, G. Synergistic effect of acute hypoxia on flow-induced release of ATP from cultured endothelial cells. Experientia 1995, 51, 256–259. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Purine and purinergic receptors. Brain Neurosci. Adv. 2018, 2. [Google Scholar] [CrossRef] [Green Version]
- Ciruela, F.; Jacobson, K.A. Editorial: Purinergic Pharmacology. Front. Pharmacol. 2019, 10, 21. [Google Scholar] [CrossRef]
- Von Kugelgen, I.; Wetter, A. Molecular pharmacology of P2Y-receptors. Naunyn Schmiedebergs Arch. Pharmacol. 2000, 362, 310–323. [Google Scholar] [CrossRef]
- Von Kugelgen, I.; Harden, T.K. Molecular pharmacology, physiology, and structure of the P2Y receptors. Adv. Pharmacol. 2011, 61, 373–415. [Google Scholar] [CrossRef]
- Jacobson, K.A.; Paoletta, S.; Katritch, V.; Wu, B.; Gao, Z.G.; Zhao, Q.; Stevens, R.C.; Kiselev, E. Nucleotides Acting at P2Y Receptors: Connecting Structure and Function. Mol. Pharmacol. 2015, 88, 220–230. [Google Scholar] [CrossRef] [Green Version]
- Kauffenstein, G.; Drouin, A.; Thorin-Trescases, N.; Bachelard, H.; Robaye, B.; D’Orleans-Juste, P.; Marceau, F.; Thorin, E.; Sevigny, J. NTPDase1 (CD39) controls nucleotide-dependent vasoconstriction in mouse. Cardiovasc. Res. 2010, 85, 204–213. [Google Scholar] [CrossRef] [Green Version]
- Bours, M.J.; Swennen, E.L.; Di Virgilio, F.; Cronstein, B.N.; Dagnelie, P.C. Adenosine 5′-triphosphate and adenosine as endogenous signaling molecules in immunity and inflammation. Pharmacol. Ther. 2006, 112, 358–404. [Google Scholar] [CrossRef]
- Ralevic, V.; Burnstock, G. Receptors for purines and pyrimidines. Pharmacol. Rev. 1998, 50, 413–492. [Google Scholar]
- Erlinge, D.; Burnstock, G. P2 receptors in cardiovascular regulation and disease. Purinergic. Signal. 2008, 4, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Gitterman, D.P.; Evans, R.J. Properties of P2X and P2Y receptors are dependent on artery diameter in the rat mesenteric bed. Br. J. Pharmacol. 2000, 131, 1561–1568. [Google Scholar] [CrossRef] [Green Version]
- Shryock, J.C.; Belardinelli, L. Adenosine and adenosine receptors in the cardiovascular system: Biochemistry, physiology, and pharmacology. Am. J. Cardiol. 1997, 79, 2–10. [Google Scholar] [CrossRef]
- Robson, S.C.; Sevigny, J.; Zimmermann, H. The E-NTPDase family of ectonucleotidases: Structure function relationships and pathophysiological significance. Purinergic Signal. 2006, 2, 409–430. [Google Scholar] [CrossRef] [Green Version]
- Kukulski, F.; Levesque, S.A.; Lavoie, E.G.; Lecka, J.; Bigonnesse, F.; Knowles, A.F.; Robson, S.C.; Kirley, T.L.; Sevigny, J. Comparative hydrolysis of P2 receptor agonists by NTPDases 1, 2, 3 and 8. Purinergic Signal. 2005, 1, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Colgan, S.P.; Eltzschig, H.K.; Eckle, T.; Thompson, L.F. Physiological roles for ecto-5′-nucleotidase (CD73). Purinergic Signal. 2006, 2, 351–360. [Google Scholar] [CrossRef] [Green Version]
- Vorhoff, T.; Zimmermann, H.; Pelletier, J.; Sevigny, J.; Braun, N. Cloning and characterization of the ecto-nucleotidase NTPDase3 from rat brain: Predicted secondary structure and relation to other members of the E-NTPDase family and actin. Purinergic Signal. 2005, 1, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Sevigny, J.; Sundberg, C.; Braun, N.; Guckelberger, O.; Csizmadia, E.; Qawi, I.; Imai, M.; Zimmermann, H.; Robson, S.C. Differential catalytic properties and vascular topography of murine nucleoside triphosphate diphosphohydrolase 1 (NTPDase1) and NTPDase2 have implications for thromboregulation. Blood 2002, 99, 2801–2809. [Google Scholar] [CrossRef] [Green Version]
- Enjyoji, K.; Sevigny, J.; Lin, Y.; Frenette, P.S.; Christie, P.D.; Esch, J.S., II; Imai, M.; Edelberg, J.M.; Rayburn, H.; Lech, M.; et al. Targeted disruption of cd39/ATP diphosphohydrolase results in disordered hemostasis and thromboregulation. Nat. Med. 1999, 5, 1010–1017. [Google Scholar] [CrossRef]
- Lavoie, E.G.; Gulbransen, B.D.; Martin-Satue, M.; Aliagas, E.; Sharkey, K.A.; Sevigny, J. Ectonucleotidases in the digestive system: Focus on NTPDase3 localization. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G608–G620. [Google Scholar] [CrossRef]
- Kauffenstein, G.; Pelletier, J.; Lavoie, E.G.; Kukulski, F.; Martin-Satue, M.; Dufresne, S.S.; Frenette, J.; Ribas Furstenau, C.; Sereda, M.J.; Toutain, B.; et al. Nucleoside triphosphate diphosphohydrolase-1 ectonucleotidase is required for normal vas deferens contraction and male fertility through maintaining P2X1 receptor function. J. Biol. Chem. 2014, 289, 28629–28639. [Google Scholar] [CrossRef] [Green Version]
- Martin-Satue, M.; Lavoie, E.G.; Pelletier, J.; Fausther, M.; Csizmadia, E.; Guckelberger, O.; Robson, S.C.; Sevigny, J. Localization of plasma membrane bound NTPDases in the murine reproductive tract. Histochem. Cell Biol. 2009, 131, 615–628. [Google Scholar] [CrossRef]
- Bartel, D.L.; Sullivan, S.L.; Lavoie, E.G.; Sevigny, J.; Finger, T.E. Nucleoside triphosphate diphosphohydrolase-2 is the ecto-ATPase of type I cells in taste buds. J. Comp. Neurol. 2006, 497, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Pelletier, J.; Salem, M.; Lecka, J.; Fausther, M.; Bigonnesse, F.; Sevigny, J. Generation and Characterization of Specific Antibodies to the Murine and Human Ectonucleotidase NTPDase8. Front. Pharmacol. 2017, 8, 115. [Google Scholar] [CrossRef] [Green Version]
- Fausther, M.; Lecka, J.; Soliman, E.; Kauffenstein, G.; Pelletier, J.; Sheung, N.; Dranoff, J.A.; Sevigny, J. Coexpression of ecto-5′-nucleotidase/CD73 with specific NTPDases differentially regulates adenosine formation in the rat liver. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G447–G459. [Google Scholar] [CrossRef]
- Bar, I.; Guns, P.J.; Metallo, J.; Cammarata, D.; Wilkin, F.; Boeynams, J.M.; Bult, H.; Robaye, B. Knockout mice reveal a role for P2Y6 receptor in macrophages, endothelial cells, and vascular smooth muscle cells. Mol. Pharmacol. 2008, 74, 777–784. [Google Scholar] [CrossRef] [Green Version]
- Braun, N.; Sevigny, J.; Mishra, S.K.; Robson, S.C.; Barth, S.W.; Gerstberger, R.; Hammer, K.; Zimmermann, H. Expression of the ecto-ATPase NTPDase2 in the germinal zones of the developing and adult rat brain. Eur. J. Neurosci. 2003, 17, 1355–1364. [Google Scholar] [CrossRef]
- Sevigny, J.; Levesque, F.P.; Grondin, G.; Beaudoin, A.R. Purification of the blood vessel ATP diphosphohydrolase, identification and localisation by immunological techniques. Biochim. Biophys. Acta 1997, 1334, 73–88. [Google Scholar] [CrossRef]
- Baykov, A.A.; Evtushenko, O.A.; Avaeva, S.M. A malachite green procedure for orthophosphate determination and its use in alkaline phosphatase-based enzyme immunoassay. Anal. Biochem. 1988, 171, 266–270. [Google Scholar] [CrossRef]
- Lu, M.; Zhu, K.; Schulam, P.G.; Chai, T.C. A non-enzymatic method for dissection of mouse bladder urothelial tissue. Nat. Protoc. 2019, 14, 1280–1292. [Google Scholar] [CrossRef]
- Heine, P.; Braun, N.; Heilbronn, A.; Zimmermann, H. Functional characterization of rat ecto-ATPase and ecto-ATP diphosphohydrolase after heterologous expression in CHO cells. Eur. J. Biochem. 1999, 262, 102–107. [Google Scholar] [CrossRef]
- Goepfert, C.; Sundberg, C.; Sevigny, J.; Enjyoji, K.; Hoshi, T.; Csizmadia, E.; Robson, S. Disordered cellular migration and angiogenesis in cd39-null mice. Circulation 2001, 104, 3109–3115. [Google Scholar] [CrossRef] [Green Version]
- Guckelberger, O.; Sun, X.F.; Sevigny, J.; Imai, M.; Kaczmarek, E.; Enjyoji, K.; Kruskal, J.B.; Robson, S.C. Beneficial effects of CD39/ecto-nucleoside triphosphate diphosphohydrolase-1 in murine intestinal ischemia-reperfusion injury. Thromb. Haemost. 2004, 91, 576–586. [Google Scholar] [CrossRef]
- Yu, W.; Robson, S.C.; Hill, W.G. Expression and distribution of ectonucleotidases in mouse urinary bladder. PLoS ONE 2011, 6, e18704. [Google Scholar] [CrossRef] [Green Version]
- Kauffenstein, G.; Tamareille, S.; Prunier, F.; Roy, C.; Ayer, A.; Toutain, B.; Billaud, M.; Isakson, B.E.; Grimaud, L.; Loufrani, L.; et al. Central Role of P2Y6 UDP Receptor in Arteriolar Myogenic Tone. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 1598–1606. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Sun, X.; Robson, S.C.; Hill, W.G. Extracellular UDP enhances P2X-mediated bladder smooth muscle contractility via P2Y(6) activation of the phospholipase C/inositol trisphosphate pathway. FASEB J. 2013, 27, 1895–1903. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, I.; Timoteo, M.A.; Silva, I.; Vieira, C.; Baldaia, C.; Ferreirinha, F.; Silva-Ramos, M.; Correia-de-Sa, P. Activation of P2Y6 receptors increases the voiding frequency in anaesthetized rats by releasing ATP from the bladder urothelium. Br. J. Pharmacol. 2014, 171, 3404–3419. [Google Scholar] [CrossRef] [Green Version]
- Kira, S.; Yoshiyama, M.; Tsuchiya, S.; Shigetomi, E.; Miyamoto, T.; Nakagomi, H.; Shibata, K.; Mochizuki, T.; Takeda, M.; Koizumi, S. P2Y6-deficiency increases micturition frequency and attenuates sustained contractility of the urinary bladder in mice. Sci. Rep. 2017, 7, 771. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Sun, X.; Robson, S.C.; Hill, W.G. ADP-induced bladder contractility is mediated by P2Y12 receptor and temporally regulated by ectonucleotidases and adenosine signaling. FASEB J. 2014, 28, 5288–5298. [Google Scholar] [CrossRef]
- Burnstock, G.; Dumsday, B.; Smythe, A. Atropine resistant excitation of the urinary bladder: The possibility of transmission via nerves releasing a purine nucleotide. Br. J. Pharmacol. 1972, 44, 451–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnstock, G.; Satchell, D.G.; Smythe, A. A comparison of the excitatory and inhibitory effects of non-adrenergic, non-cholinergic nerve stimulation and exogenously applied ATP on a variety of smooth muscle preparations from different vertebrate species. Br. J. Pharmacol. 1972, 46, 234–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vial, C.; Evans, R.J. P2X receptor expression in mouse urinary bladder and the requirement of P2X(1) receptors for functional P2X receptor responses in the mouse urinary bladder smooth muscle. Br. J. Pharmacol. 2000, 131, 1489–1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heppner, T.J.; Werner, M.E.; Nausch, B.; Vial, C.; Evans, R.J.; Nelson, M.T. Nerve-evoked purinergic signalling suppresses action potentials, Ca2+ flashes and contractility evoked by muscarinic receptor activation in mouse urinary bladder smooth muscle. J. Physiol. 2009, 587, 5275–5288. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, C.; Tasker, P.N.; Gallacher, G.; Westfall, T.D. Identification of atropine- and P2X1 receptor antagonist-resistant, neurogenic contractions of the urinary bladder. J. Neurosci. 2007, 27, 845–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, W.; Hill, W.G.; Robson, S.C.; Zeidel, M.L. Role of P2X4 Receptor in Mouse Voiding Function. Sci. Rep. 2018, 8, 1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaefer, U.; Machida, T.; Broekman, M.J.; Marcus, A.J.; Levi, R. Targeted deletion of ectonucleoside triphosphate diphosphohydrolase 1/CD39 leads to desensitization of pre- and postsynaptic purinergic P2 receptors. J. Pharmacol. Exp. Ther. 2007, 322, 1269–1277. [Google Scholar] [CrossRef]
- Mizumoto, N.; Kumamoto, T.; Robson, S.C.; Sevigny, J.; Matsue, H.; Enjyoji, K.; Takashima, A. CD39 is the dominant Langerhans cell-associated ecto-NTPDase: Modulatory roles in inflammation and immune responsiveness. Nat. Med. 2002, 8, 358–365. [Google Scholar] [CrossRef]
- Song, P.; Zhang, M.; Wang, S.; Xu, J.; Choi, H.C.; Zou, M.H. Thromboxane A2 receptor activates a Rho-associated kinase/LKB1/PTEN pathway to attenuate endothelium insulin signaling. J. Biol. Chem. 2009, 284, 17120–17128. [Google Scholar] [CrossRef] [Green Version]
- Goulopoulou, S.; Hannan, J.L.; Matsumoto, T.; Webb, R.C. Pregnancy reduces RhoA/Rho kinase and protein kinase C signaling pathways downstream of thromboxane receptor activation in the rat uterine artery. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H2477–H2488. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, T.; Kakami, M.; Kobayashi, T.; Kamata, K. Gender differences in vascular reactivity to endothelin-1 (1-31) in mesenteric arteries from diabetic mice. Peptides 2008, 29, 1338–1346. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.G.; Khalil, R.A. Gender-specific reduction in contractility and [Ca(2+)](i) in vascular smooth muscle cells of female rat. Am. J. Physiol. Cell Physiol. 2000, 278, C834–C844. [Google Scholar] [CrossRef] [PubMed]
- Barber, D.A.; Miller, V.M. Gender differences in endothelium-dependent relaxations do not involve NO in porcine coronary arteries. Am. J. Physiol. 1997, 273, H2325–H2332. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Qiao, X.; Falone, A.E.; Reslan, O.M.; Sheppard, S.J.; Khalil, R.A. Gender-specific reduction in contraction is associated with increased estrogen receptor expression in single vascular smooth muscle cells of female rat. Cell Physiol. Biochem. 2010, 26, 457–470. [Google Scholar] [CrossRef]
- Caiazzo, E.; Bilancia, R.; Rossi, A.; Ialenti, A.; Cicala, C. Ectonucleoside Triphosphate Diphosphohydrolase-1/CD39 Affects the Response to ADP of Female Rat Platelets. Front. Pharmacol. 2019, 10, 1689. [Google Scholar] [CrossRef] [Green Version]
- Koziak, K.; Bojakowska, M.; Robson, S.C.; Bojakowski, K.; Soin, J.; Csizmadia, E.; Religa, P.; Gaciong, Z.; Kaczmarek, E. Overexpression of CD39/nucleoside triphosphate diphosphohydrolase-1 decreases smooth muscle cell proliferation and prevents neointima formation after angioplasty. J. Thromb. Haemost. 2008, 6, 1191–1197. [Google Scholar] [CrossRef] [Green Version]
- Huttinger, Z.M.; Milks, M.W.; Nickoli, M.S.; Aurand, W.L.; Long, L.C.; Wheeler, D.G.; Dwyer, K.M.; d’Apice, A.J.; Robson, S.C.; Cowan, P.J.; et al. Ectonucleotide triphosphate diphosphohydrolase-1 (CD39) mediates resistance to occlusive arterial thrombus formation after vascular injury in mice. Am. J. Pathol. 2012, 181, 322–333. [Google Scholar] [CrossRef] [Green Version]
- Patra, P.B.; Patra, S. Sex differences in the physiology and pharmacology of the lower urinary tract. Curr. Urol. 2013, 6, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Kamei, J.; Ito, H.; Aizawa, N.; Hotta, H.; Kojima, T.; Fujita, Y.; Ito, M.; Homma, Y.; Igawa, Y. Age-related changes in function and gene expression of the male and female mouse bladder. Sci. Rep. 2018, 8, 2089. [Google Scholar] [CrossRef]
- Searl, T.J.; Dynda, D.I.; Alanee, S.R.; El-Zawahry, A.M.; McVary, K.T.; Silinsky, E.M. A1 Adenosine Receptor-Mediated Inhibition of Parasympathetic Neuromuscular Transmission in Human and Murine Urinary Bladder. J. Pharmacol. Exp. Ther. 2016, 356, 116–122. [Google Scholar] [CrossRef] [Green Version]
- Hao, Y.; Wang, L.; Chen, H.; Hill, W.G.; Robson, S.C.; Zeidel, M.L.; Yu, W. Targetable purinergic receptors P2Y12 and A2b antagonistically regulate bladder function. JCI Insight 2019, 4. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer | Amplicon (bp) |
|---|---|---|---|
| Krt7 | Qiagen | Qiagen | 134 |
| Myh11 | Qiagen | Qiagen | 112 |
| Gapdh | CCA TCA CCA TCT TCC AGG AG | GTG GTT CAC ACC CAT CAC AA | 194 |
| Actb | GGC TGT ATT CCC CTC CAT CG | CCA GTT GGT AAC AAT GCC ATG T | 154 |
| Entpdl | AGC TGC CCC TTA TGG AAG AT | TCA GTC CCA CAG CAA TCA AA | 123 |
| Entpd2 | TTC CTG GGA TGT CAG GTC TC | GTC TCT GGT GCT TGC CTT TC | 132 |
| Entpd3 | ACC TGT CCC GTG CTT AAA TG | AGA CAG AGT GAA GCC CCT GA | 183 |
| Entpd8 | Qiagen | Qiagen | 146 |
| Nt5e/CD73 | CAG GAA ATC CAC CTT CCA AA | AAC CTT CAG GTA GCC CAG GT | 128 |
| P2ry1 | TCG TGT CTC CAT TCT GCT TG | CGA CAG GGT TTA TGC CAC TT | 218 |
| P2ry2 | TGA CGA CTC AAG ACG GAC AG | GTC CCC TAC AGC TCC CCT AC | 108 |
| P2ry4 | AGA CGG GCC TGA TGT GTA TC | AGG TTC ACA TGC CCT GTA CC | 126 |
| P2ry6 | GGT AGC GCT GGA AGC TAA TG | TTT CAA GCG ACT GCT GCT AA | 308 |
| P2ry12 | GGC AGC CTT GAG TGT TCT TC | ATA ACG TGC TAC CCG ACC TG | 130 |
| P2ry13 | ATA GAG AAC CGG GAA CAG CA | CAA AAC AAA GCT GAT GCT CG | 115 |
| P2ry14 | TTT TGT CGT CTG CTT TGT GC | GCA GCC GAG AGT AGC AGA GT | 135 |
| P2rx1 | CAA CTG TGT GCC CTT CAA TG | GGT ACC ATT CAC CTC CTC CA | 114 |
| Pr2x2 | GCT GGG CTT CAT TGT AGA GC | CCT GTC CAT GCA CAA TAA CG | 281 |
| P2rx3 | ATT TCC TCA AAG GGG CTG AT | GTT CTG CAG CCC AAG GAT AA | 204 |
| P2rx4 | CAC AAC GTG TCT CCT GGC TA | GCC TTT CCA AAC ACG ATG AT | 125 |
| P2rx6 | TCA CCC GCT AAC CCT GTT AC | TAG TCC CGC TGA AGC TTT GT | 242 |
| P2rx7 | AAT CGG TGT GTT TCC TTT GG | CCG GGT GAC TTT GTT TGT CT | 165 |
| Adora1 | GTG ATT TGG GCT GTG AAG GT | AGT AGG TCT GTG GCC CAA TG | 142 |
| Adora2a | TCA ACA GCA ACC TGC AGA AC | GGC TGA AGA TGG AAC TCT GC | 186 |
| Adora2b | TCT GGC CTT TTG GAG AAG AA | TTT CCG GAA TCA ATT CAA GC | 246 |
| Adora3 | TGT GGA GGG AGT CTC GTC TT | TCC TTC TGT TCC CCA CAT TC | 101 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Babou Kammoe, R.B.; Kauffenstein, G.; Pelletier, J.; Robaye, B.; Sévigny, J. NTPDase1 Modulates Smooth Muscle Contraction in Mice Bladder by Regulating Nucleotide Receptor Activation Distinctly in Male and Female. Biomolecules 2021, 11, 147. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11020147
Babou Kammoe RB, Kauffenstein G, Pelletier J, Robaye B, Sévigny J. NTPDase1 Modulates Smooth Muscle Contraction in Mice Bladder by Regulating Nucleotide Receptor Activation Distinctly in Male and Female. Biomolecules. 2021; 11(2):147. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11020147
Chicago/Turabian StyleBabou Kammoe, Romuald Brice, Gilles Kauffenstein, Julie Pelletier, Bernard Robaye, and Jean Sévigny. 2021. "NTPDase1 Modulates Smooth Muscle Contraction in Mice Bladder by Regulating Nucleotide Receptor Activation Distinctly in Male and Female" Biomolecules 11, no. 2: 147. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11020147