
Effect of Postnatal Nutritional Environment Due to Maternal Diabetes on Beta Cell Mass Programming and Glucose Intolerance Risk in Male and Female Offspring


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Embryo Harvesting and Transfer Procedures
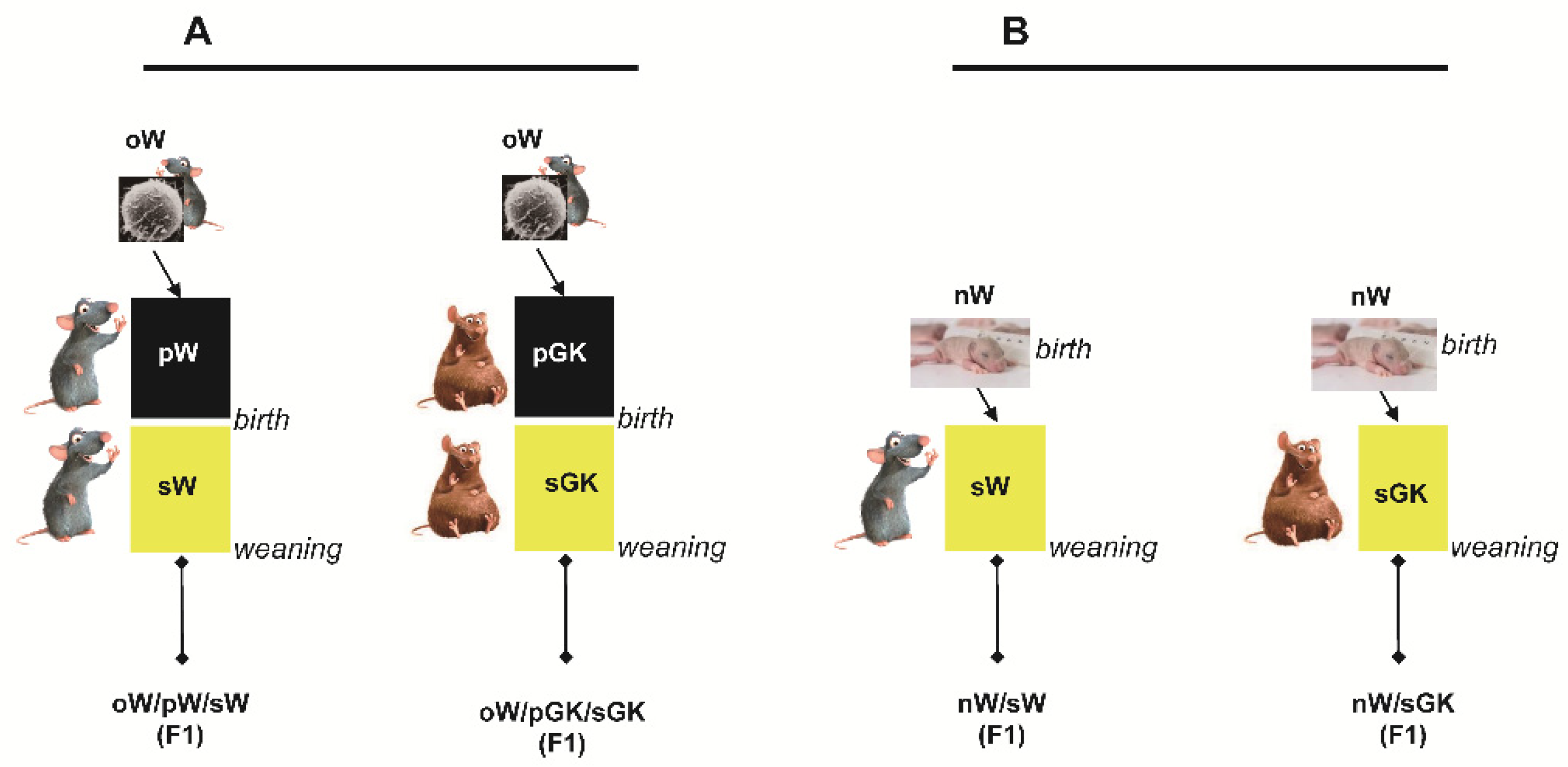
2.3. First Set of Experiments: Embryo-Transferred and Cross-Fostered Groups
2.4. Second Set of Experiments: Cross-Fostered Groups
2.5. Glucose Tolerance and Insulin Secretion Tests
2.6. Immunohistochemical Analysis
2.7. Morphometric Image Analysis
2.8. Calculations
3. Results
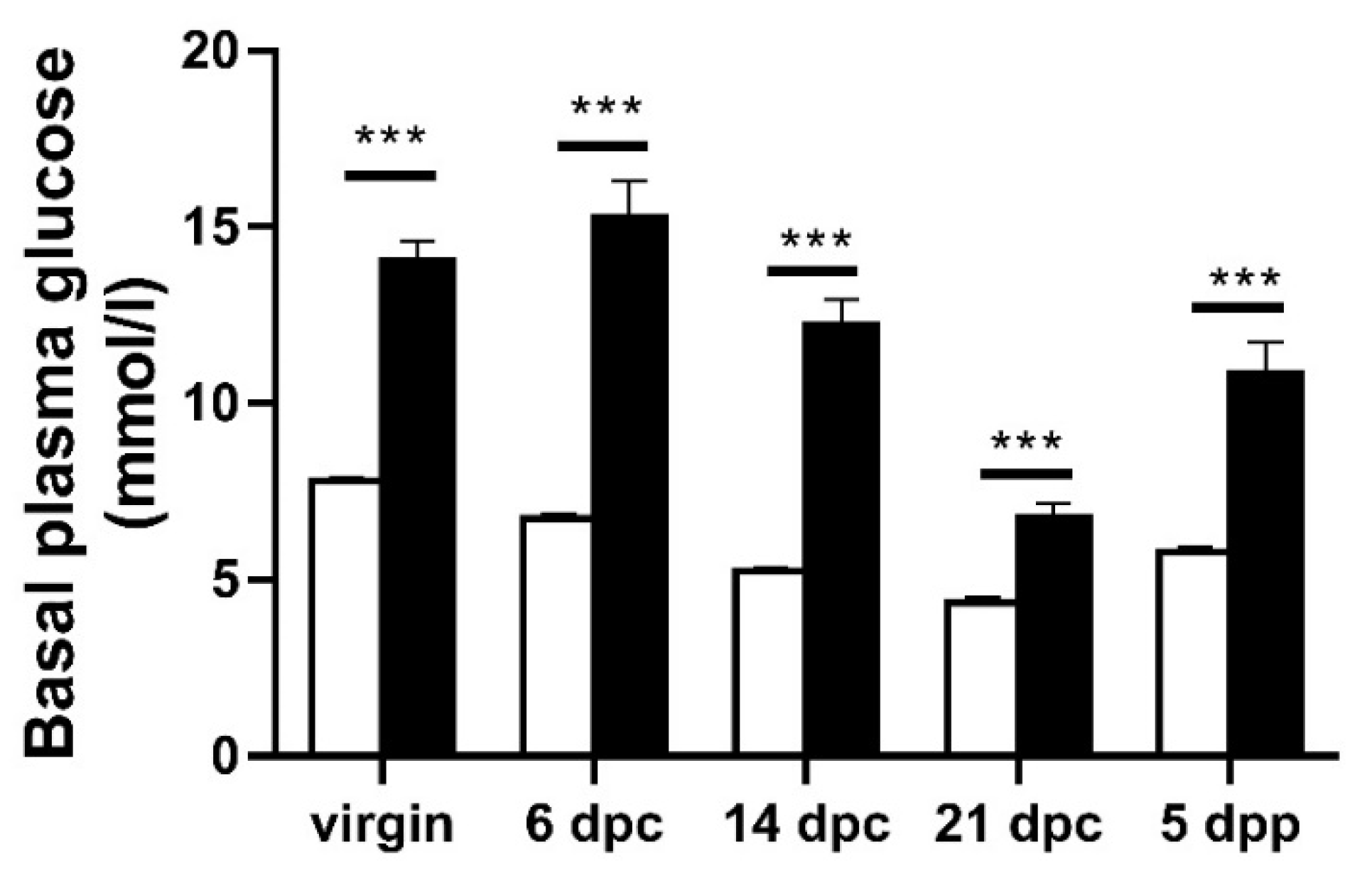
3.1. Diabetes Phenotype in Pregnant and Lactating GK Females
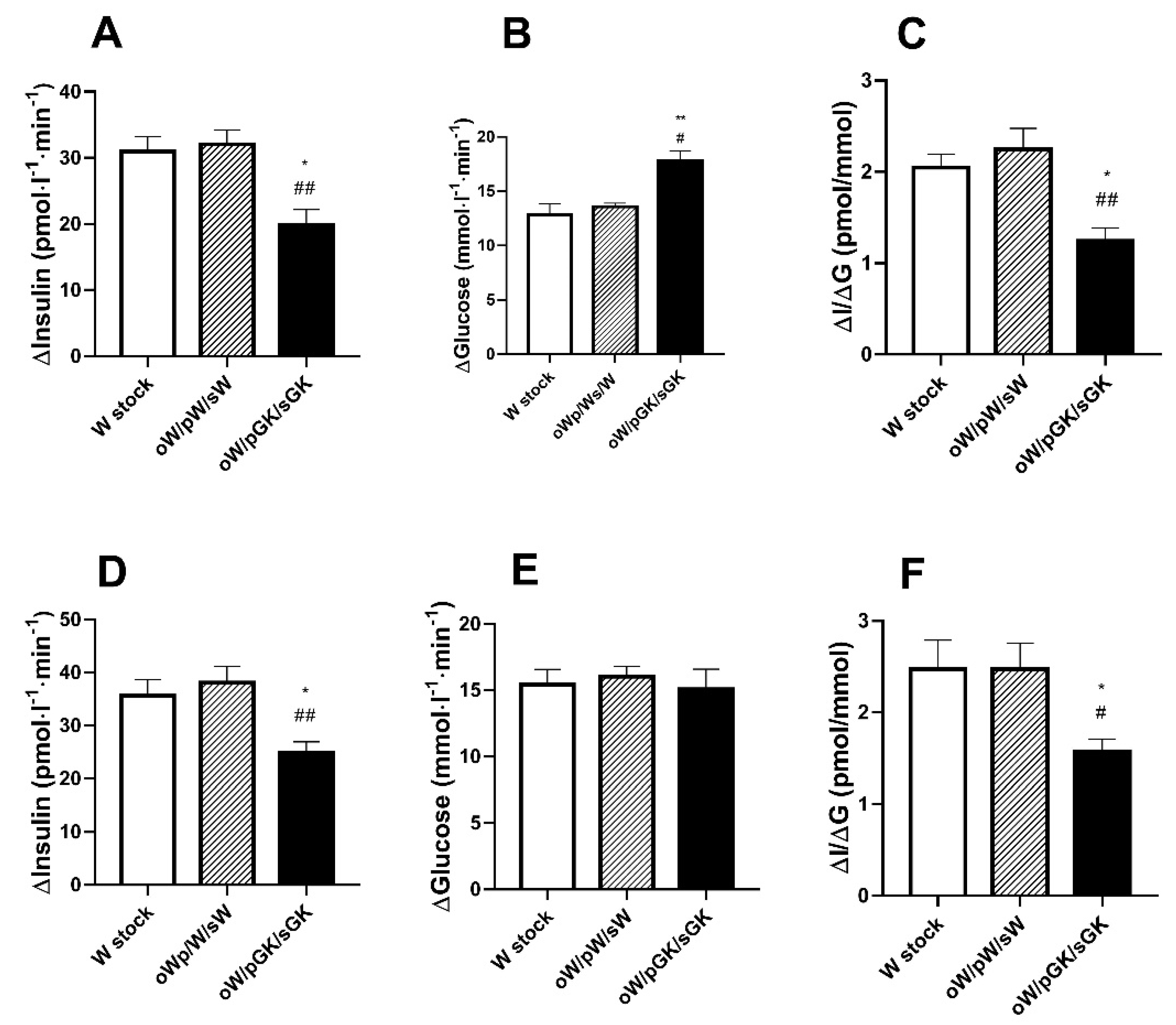
3.2. Adult W/pGK/sGK Offspring Have Decreased Beta Cell Mass and Low Glucose-Stimulated Insulin Secretion and They Are Glucose Intolerant
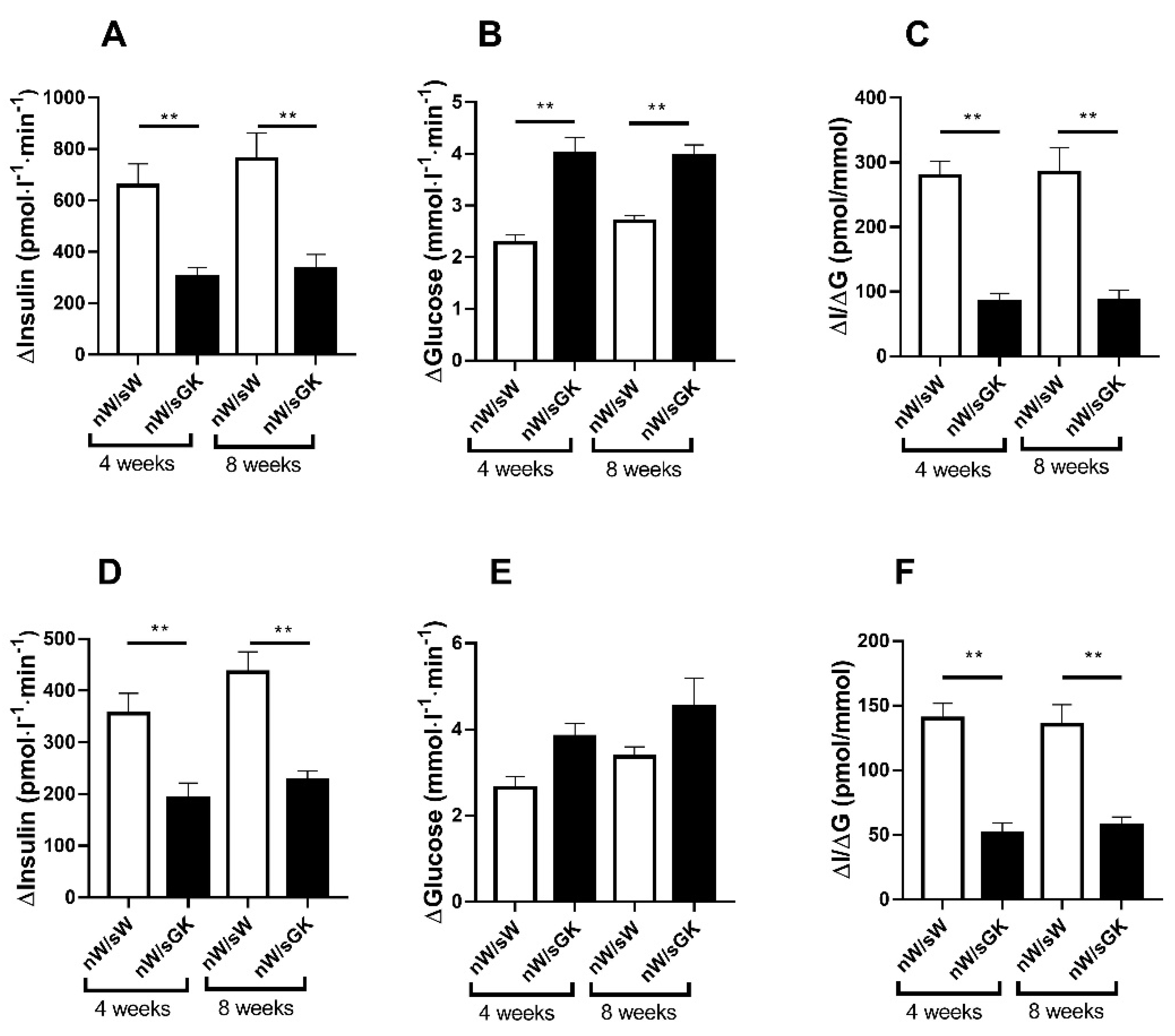
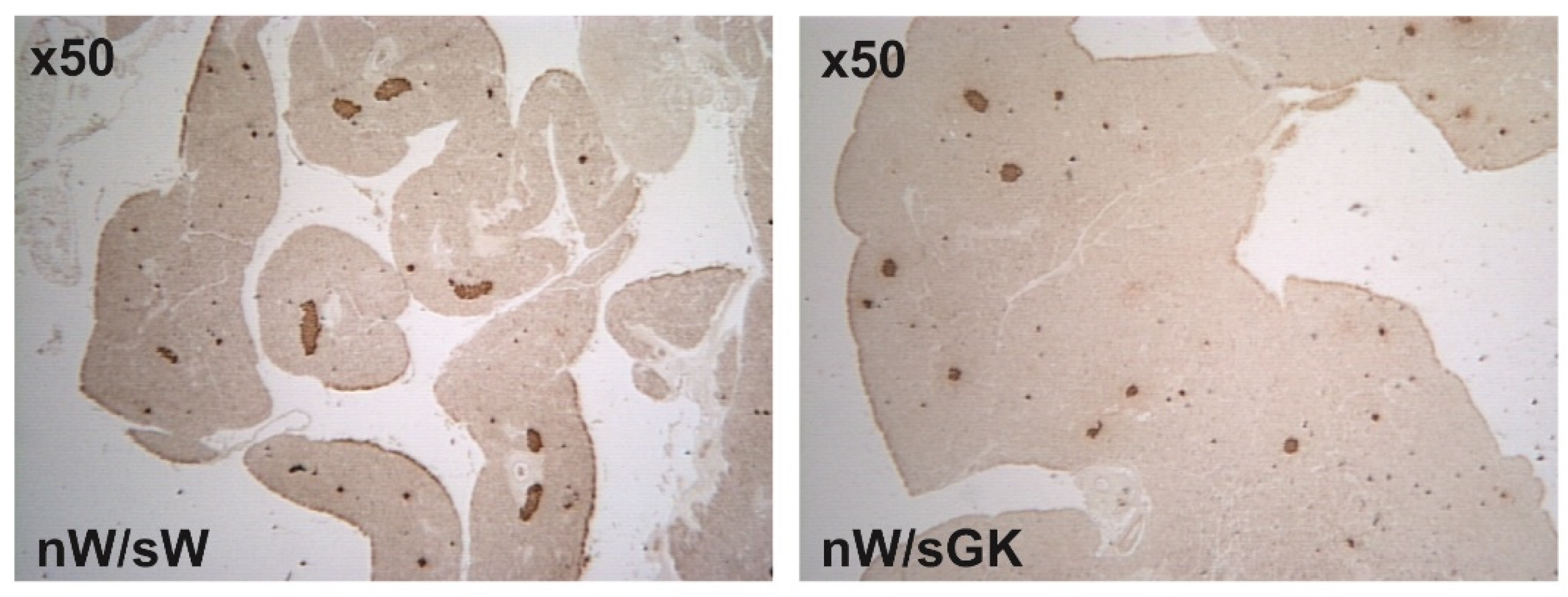
3.3. Adult nW/sGK Offspring Have Decreased Beta Cell Mass and Low Glucose-Stimulated Insulin Secretion and They Are Glucose Tolerant
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harris, M.I.; Klein, R.; Welborn, T.A.; Knuiman, M.W. Onset of NIDDM occurs at least 4–7 yr before clinical diagnosis. Diabetes Care 1992, 15, 815–819. [Google Scholar] [CrossRef] [PubMed]
- Defronzo, R.A. Banting Lecture. From the triumvirate to the ominous octet: A new paradigm for the treatment of type 2 diabetes mellitus. Diabetes 2009, 58, 773–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voight, B.F.; Scott, L.J.; Steinthorsdottir, V.; Morris, A.P.; Dina, C.; Welch, R.P.; Zeggini, E.; Huth, C.; Aulchenko, Y.S.; Thorleifsson, G.; et al. Twelve type 2 diabetes susceptibility loci identified through large-scale association analysis. Nat. Genet. 2010, 42, 579–589. [Google Scholar] [CrossRef]
- Ling, C.; Groop, L. Epigenetics: A molecular link between environmental factors and type 2 diabetes. Diabetes 2009, 58, 2718–2725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groop, L.; Pociot, F. Genetics of diabetes—Are we missing the genes or the disease? Mol. Cell. Endocrinol. 2014, 382, 726–739. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Twinn, D.S.; Constância, M.; Ozanne, S.E. Intergenerational epigenetic inheritance in models of developmental programming of adult disease. Semin. Cell Dev. Biol. 2015, 43, 85–95. [Google Scholar] [CrossRef]
- Rando, O.J.; Simmons, R.A. I’m eating for two: Parental dietary effects on offspring metabolism. Cell 2015, 161, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Chavey, A.; Ah Kioon, M.-D.; Bailbé, D.; Movassat, J.; Portha, B. Maternal diabetes, programming of beta-cell disorders and intergenerational risk of type 2 diabetes. Diabetes Metab. 2014, 40, 323–330. [Google Scholar] [CrossRef]
- Portha, B.; Grandjean, V.; Movassat, J. Mother or Father: Who Is in the Front Line? Mechanisms Underlying the Non-Genomic Transmission of Obesity/Diabetes via the Maternal or the Paternal Line. Nutrients 2019, 11, 233. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, M.; Patel, M.S. Metabolic programming in the immediate postnatal period. Trends Endocrinol. Metab. 2008, 19, 146–152. [Google Scholar] [CrossRef]
- Chavey, A.; Bailbé, D.; Maulny, L.; Renard, J.P.; Movassat, J.; Portha, B. A euglycaemic/non-diabetic perinatal environment does not alleviate early beta cell maldevelopment and type 2 diabetes risk in the GK/Par rat model. Diabetologia 2013, 56, 194–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portha, B.; Lacraz, G.; Chavey, A.; Figeac, F.; Fradet, M.; Tourrel-Cuzin, C.; Homo-Delarche, F.; Giroix, M.-H.; Bailbé, D.; Gangnerau, M.-N.; et al. Islet structure and function in the GK rat. Adv. Exp. Med. Biol. 2010, 654, 479–500. [Google Scholar] [CrossRef] [PubMed]
- Cozzi, J.; Wang, E.; Jacquet, C.; Fraichard, A.; Cherifi, Y.; Zhou, Q. Procedures for somatic cell nuclear transfer in the rat. Methods Mol. Biol. 2010, 597, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Movassat, J.; Saulnier, C.; Serradas, P.; Portha, B. Impaired development of pancreatic beta-cell mass is a primary event during the progression to diabetes in the GK rat. Diabetologia 1997, 40, 916–925. [Google Scholar] [CrossRef] [Green Version]
- Figeac, F.; Uzan, B.; Faro, M.; Chelali, N.; Portha, B.; Movassat, J. Neonatal growth and regeneration of beta-cells are regulated by the Wnt/beta-catenin signaling in normal and diabetic rats. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E245–E256. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.S.; Srinivasan, M. Metabolic programming: Causes and consequences. J. Biol. Chem. 2002, 277, 1629–1632. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, M.; Laychock, S.G.; Hill, D.J.; Patel, M.S. Neonatal nutrition: Metabolic programming of pancreatic islets and obesity. Exp. Biol. Med. 2003, 228, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Knittle, J.L.; Hirsch, J. Effect of early nutrition on the development of rat epididymal fat pads: Cellularity and metabolism. J. Clin. Investig. 1968, 47, 2091–2098. [Google Scholar] [CrossRef] [Green Version]
- Lemonnier, D.; Suquet, J.P.; Aubert, R.; Rosselin, G. Long term effect of mouse neonate food intake on adult body composition, insulin and glucose serum levels. Horm. Metab. Res. 1973, 5, 223–224. [Google Scholar] [CrossRef]
- Siebel, A.L.; Mibus, A.; De Blasio, M.J.; Westcott, K.T.; Morris, M.J.; Prior, L.; Owens, J.A.; Wlodek, M.E. Improved lactational nutrition and postnatal growth ameliorates impairment of glucose tolerance by uteroplacental insufficiency in male rat offspring. Endocrinology 2008, 149, 3067–3076. [Google Scholar] [CrossRef] [Green Version]
- Lau, C.; Sullivan, M.K.; Hazelwood, R.L. Effects of diabetes mellitus on lactation in the rat. Proc. Soc. Exp. Biol. Med. 1993, 204, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Oak, S.A.; Tran, C.; Pan, G.; Thamotharan, M.; Devaskar, S.U. Perturbed skeletal muscle insulin signaling in the adult female intrauterine growth-restricted rat. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E1321–E1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, D.I.; Barker, D.J.; Hales, C.N.; Hirst, S.; Osmond, C. Thinness at birth and insulin resistance in adult life. Diabetologia 1994, 37, 150–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thamotharan, M.; Garg, M.; Oak, S.; Rogers, L.M.; Pan, G.; Sangiorgi, F.; Lee, P.W.N.; Devaskar, S.U. Transgenerational inheritance of the insulin-resistant phenotype in embryo-transferred intrauterine growth-restricted adult female rat offspring. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E1270–E1279. [Google Scholar] [CrossRef] [Green Version]
- Garg, M.; Thamotharan, M.; Rogers, L.; Bassilian, S.; Lee, W.N.P.; Devaskar, S.U. Glucose metabolic adaptations in the intrauterine growth-restricted adult female rat offspring. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E1218–E1226. [Google Scholar] [CrossRef] [Green Version]
- Matveyenko, A.V.; Singh, I.; Shin, B.-C.; Georgia, S.; Devaskar, S.U. Differential effects of prenatal and postnatal nutritional environment on ß-cell mass development and turnover in male and female rats. Endocrinology 2010, 151, 5647–5656. [Google Scholar] [CrossRef]
- Darnaudery, M.; (Université Bordeaux, Bordeaux, France); Movassat, J.; (Université de Paris, Paris, France). Personal communication, 2019.
- Grasemann, C.; Devlin, M.J.; Rzeczkowska, P.A.; Herrmann, R.; Horsthemke, B.; Hauffa, B.P.; Grynpas, M.; Alm, C.; Bouxsein, M.L.; Palmert, M.R. Parental diabetes: The Akita mouse as a model of the effects of maternal and paternal hyperglycemia in wildtype offspring. PLoS ONE 2012, 7, e50210. [Google Scholar] [CrossRef] [Green Version]
- Yokomizo, H.; Inoguchi, T.; Sonoda, N.; Sakaki, Y.; Maeda, Y.; Inoue, T.; Hirata, E.; Takei, R.; Ikeda, N.; Fujii, M.; et al. Maternal high-fat diet induces insulin resistance and deterioration of pancreatic β-cell function in adult offspring with sex differences in mice. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E1163–E1175. [Google Scholar] [CrossRef] [Green Version]
- Louet, J.-F.; LeMay, C.; Mauvais-Jarvis, F. Antidiabetic actions of estrogen: Insight from human and genetic mouse models. Curr. Atheroscler. Rep. 2004, 6, 180–185. [Google Scholar] [CrossRef]
- Garofano, A.; Czernichow, P.; Bréant, B. Beta-cell mass and proliferation following late fetal and early postnatal malnutrition in the rat. Diabetologia 1998, 41, 1114–1120. [Google Scholar] [CrossRef] [Green Version]
- Fowden, A.L.; Forhead, A.J. Endocrine mechanisms of intrauterine programming. Reprod. Camb. Engl. 2004, 127, 515–526. [Google Scholar] [CrossRef] [PubMed]
- McMillen, I.C.; Robinson, J.S. Developmental origins of the metabolic syndrome: Prediction, plasticity, and programming. Physiol. Rev. 2005, 85, 571–633. [Google Scholar] [CrossRef] [PubMed]
- Bansal, A.; Simmons, R.A. Epigenetics and developmental origins of diabetes: Correlation or causation? Am. J. Physiol. Endocrinol. Metab. 2018, 315, E15–E28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Millán, E.; Gangnerau, M.N.; De Miguel-Santos, L.; Calderari, S.; Serradas, P.; Escrivá, F.; Portha, B.; Alvarez, C. Undernutrition of the GK rat during gestation improves pancreatic IGF-2 and beta-cell mass in the fetuses. Growth Factors Chur Switz. 2009, 27, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Stoffers, D.A.; Desai, B.M.; DeLeon, D.D.; Simmons, R.A. Neonatal exendin-4 prevents the development of diabetes in the intrauterine growth retarded rat. Diabetes 2003, 52, 734–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tourrel, C.; Bailbe, D.; Lacorne, M.; Meile, M.-J.; Kergoat, M.; Portha, B. Persistent improvement of type 2 diabetes in the Goto-Kakizaki rat model by expansion of the beta-cell mass during the prediabetic period with glucagon-like peptide-1 or exendin-4. Diabetes 2002, 51, 1443–1452. [Google Scholar] [CrossRef] [Green Version]
- Movassat, J.; Calderari, S.; Fernández, E.; Martín, M.A.; Escrivá, F.; Plachot, C.; Gangnerau, M.N.; Serradas, P.; Alvarez, C.; Portha, B. Type 2 diabetes—A matter of failing beta-cell neogenesis? Clues from the GK rat model. Diabetes Obes. Metab. 2007, 9, 187–195. [Google Scholar] [CrossRef]
- Meier, J.J. Linking the genetics of type 2 diabetes with low birth weight: A role for prenatal islet maldevelopment? Diabetes 2009, 58, 1255–1256. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rats | Sex | Body Weight (g) | Basal Plasma Glucose (mmol/L) | Pancreatic Insulin Stores (pmol/mg pancreas) | Pancreatic Beta Cell (%) | Beta Cell Mass (μg/mg pancreas) |
|---|---|---|---|---|---|---|
| oW/pW/sW | Male | 362 ± 18 (10) | 6.6 ± 0.2 (10) | 23.8 ± 1.0 (10) | 0.51 ± 0.03 (6) | 5.10 ± 0.30 (6) |
| Female | 246 ± 5 (11) | 6.4 ± 0.1 (11) | 25.3 ± 0.5 (11) | ND | ND | |
| oW/pGK/sGK | Male | 338 ± 10 (11) | 6.0 ± 0.1 (11) | 16.4 ± 0.8 ** (14) | 0.40 ± 0.03 * (6) | 3.98 ± 0.25 * (6) |
| Female | 225 ± 5 (19) | 5.9 ± 0.1 (19) | 16.3 ± 0.5 *** (11) | ND | ND |
| Age | Rats | Body Weight (g) | Basal Plasma Glucose (mmol/L) | Pancreatic Insulin Stores (pmol/mg pancreas) | Pancreatic Beta Cell (%) | Beta Cell Mass (μg/mg pancreas) |
|---|---|---|---|---|---|---|
| 4 wks | nW/sW | 85 ± 1 (10) | 6.7 ± 0.3 (10) | ND | ND | ND |
| 4 wks | nW/sGK | 65 ± 3 ** (10) | 7.2 ± 0.3 (16) | ND | ND | ND |
| 8 wks | nW/sW | 221 ± 10 (9) | 7.1 ± 0.2 (9) | 28.0 ± 1.0 (6) | 0.56 ± 0.03 (6) | 5.6 ± 0.3 (6) |
| 8 wks | nW/sGK | 212 ± 9 (9) | 7.4 ± 0.2 (9) | 19.5 ± 1.0 ** (6) | 0.43 ± 0.02 ** (6) | 4.3 ± 0.2 ** (6) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bailbe, D.; Liu, J.; Gong, P.; Portha, B. Effect of Postnatal Nutritional Environment Due to Maternal Diabetes on Beta Cell Mass Programming and Glucose Intolerance Risk in Male and Female Offspring. Biomolecules 2021, 11, 179. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11020179
Bailbe D, Liu J, Gong P, Portha B. Effect of Postnatal Nutritional Environment Due to Maternal Diabetes on Beta Cell Mass Programming and Glucose Intolerance Risk in Male and Female Offspring. Biomolecules. 2021; 11(2):179. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11020179
Chicago/Turabian StyleBailbe, Danièle, Junjun Liu, Pengfei Gong, and Bernard Portha. 2021. "Effect of Postnatal Nutritional Environment Due to Maternal Diabetes on Beta Cell Mass Programming and Glucose Intolerance Risk in Male and Female Offspring" Biomolecules 11, no. 2: 179. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11020179