
Molecular Targets and Strategies for Inhibition of the Bacterial Type III Secretion System (T3SS); Inhibitors Directly Binding to T3SS Components



Abstract
:1. Introduction
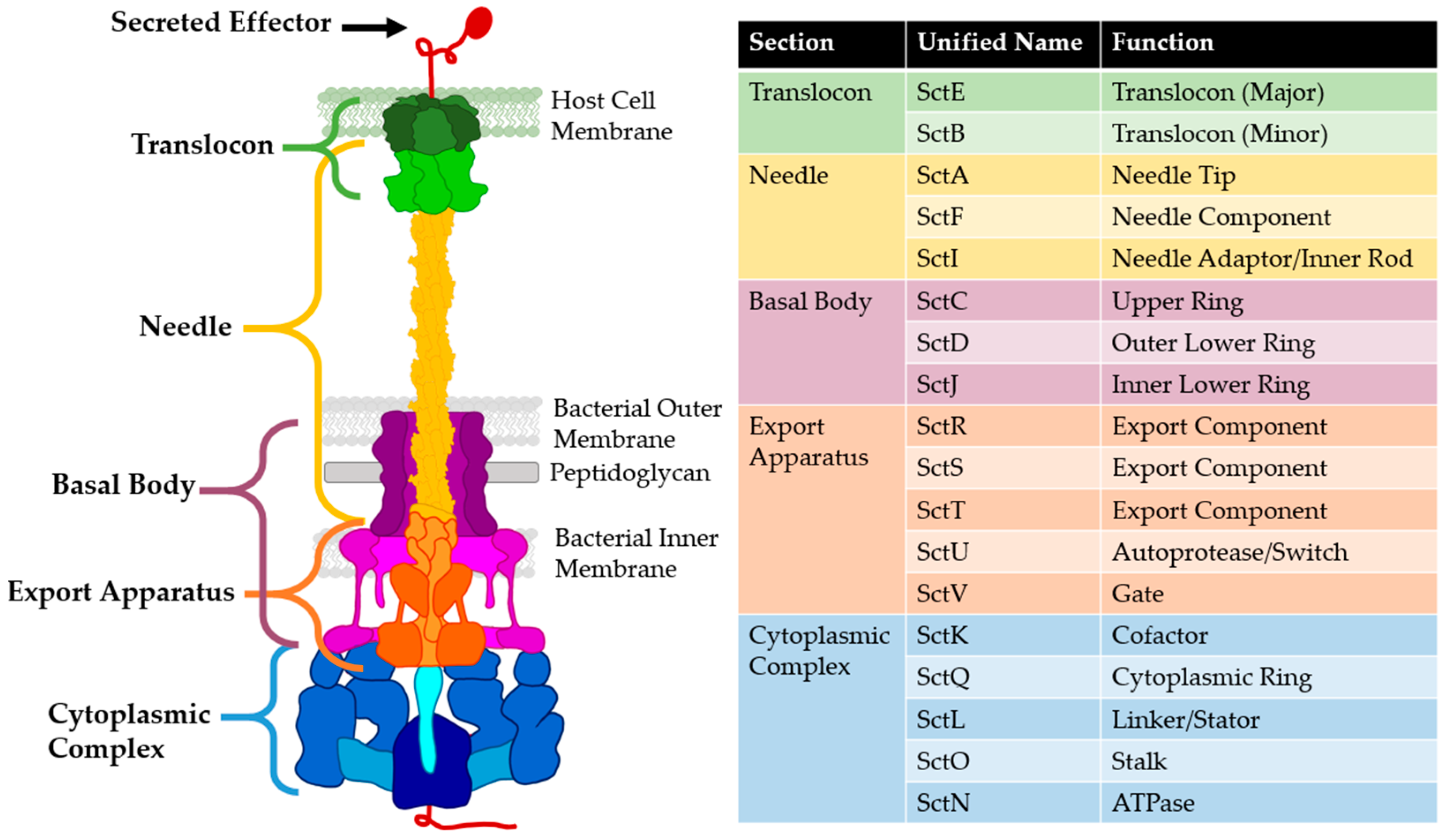
2. Needle and Translocon
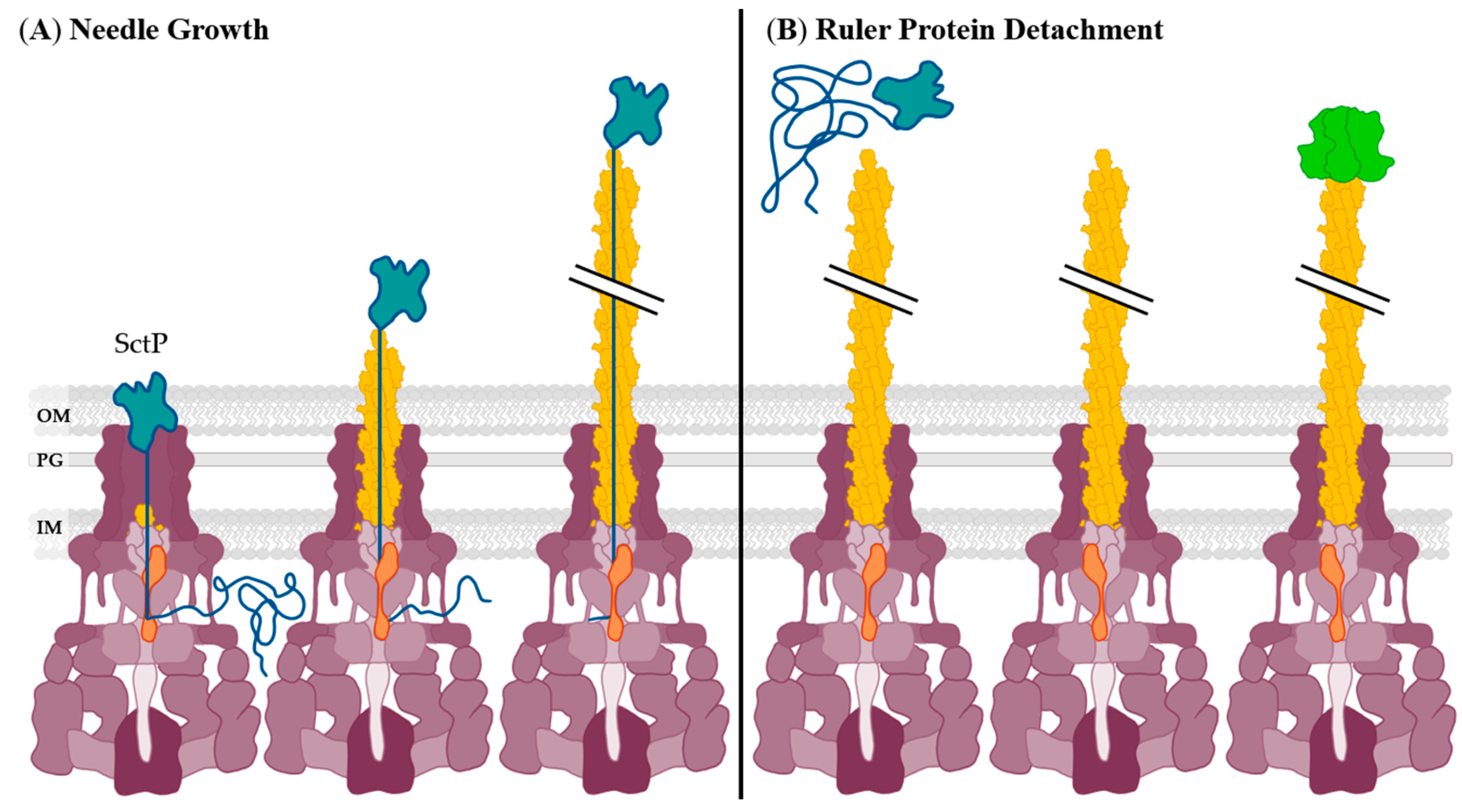
2.1. Needle Formation Inhibition
2.2. Needle Tip and Translocon Inhibition
2.3. Structural Chaperone Inhibition
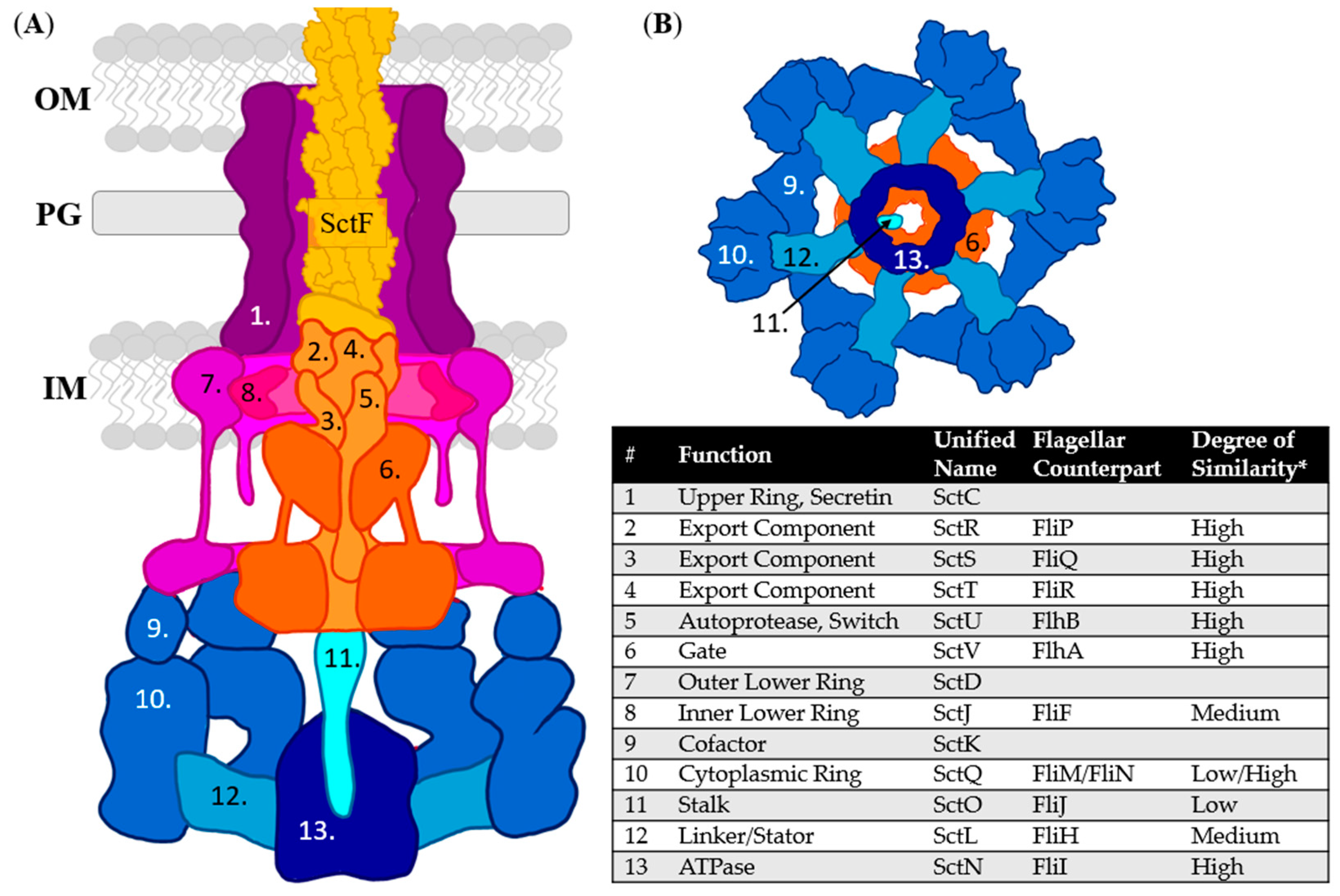
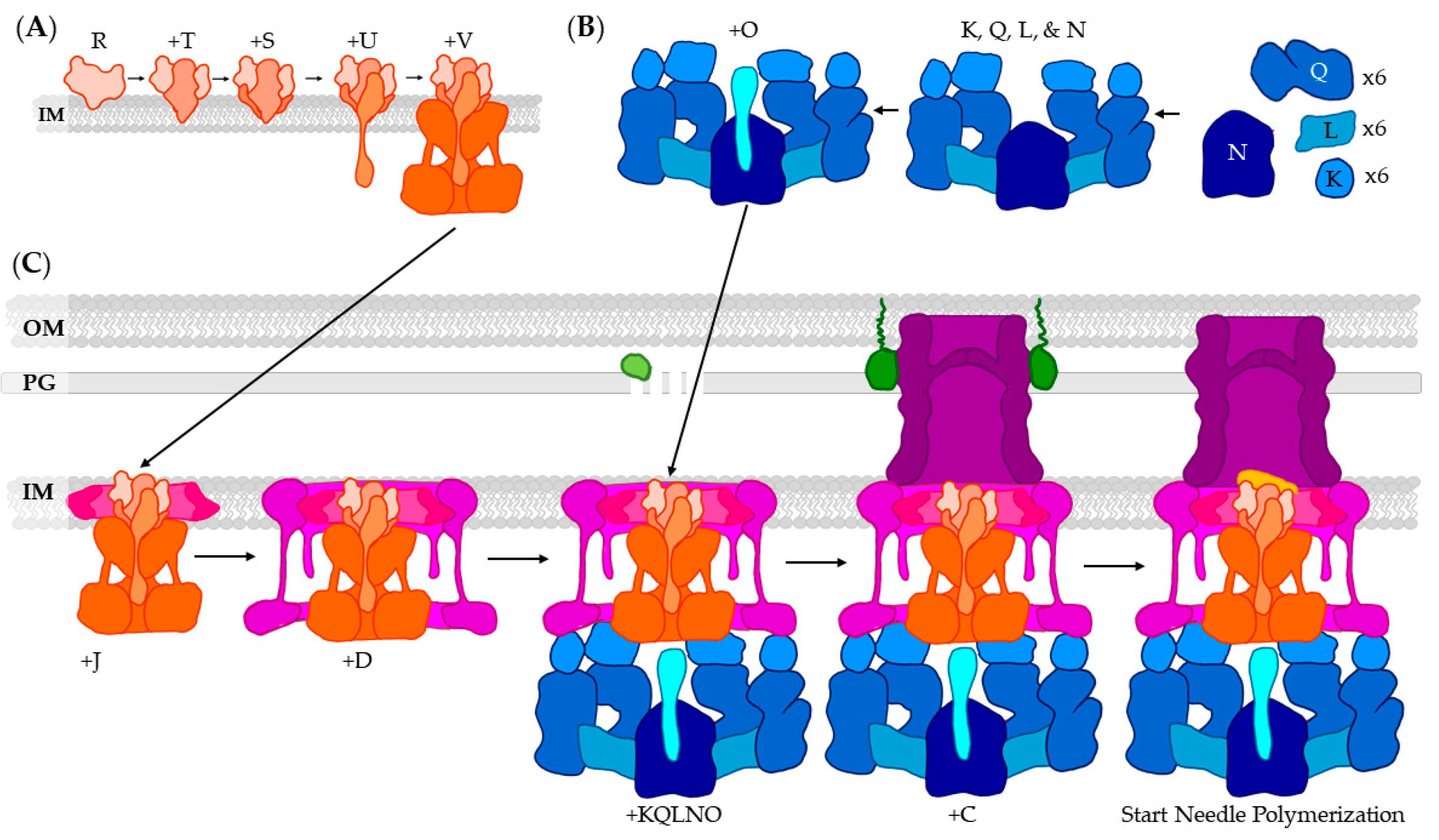
3. T3SS Base: Basal Body, Export Apparatus, and Cytoplasmic Complex
3.1. Formation Inhibition
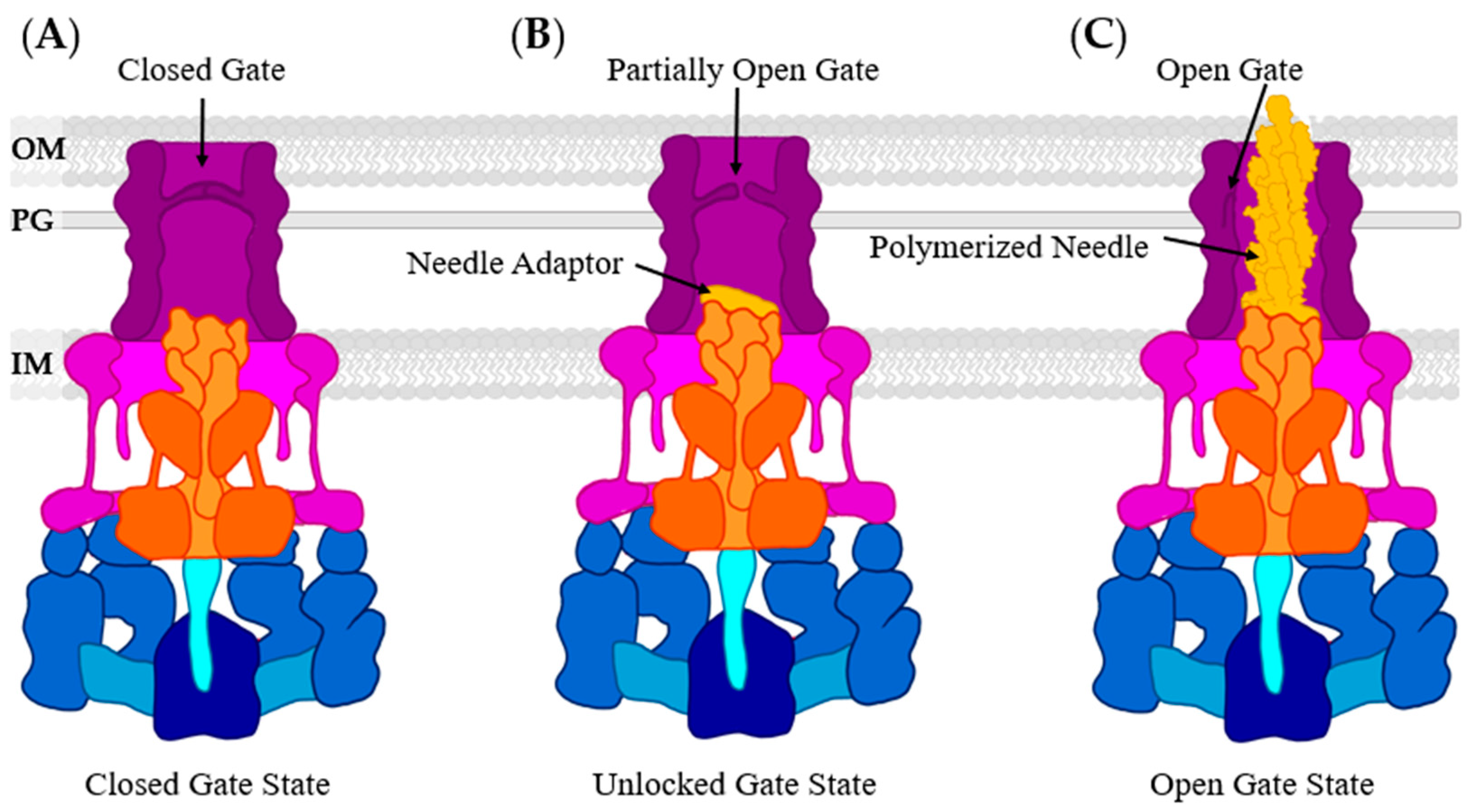
3.2. Basal Body: The Periplasmic Gate
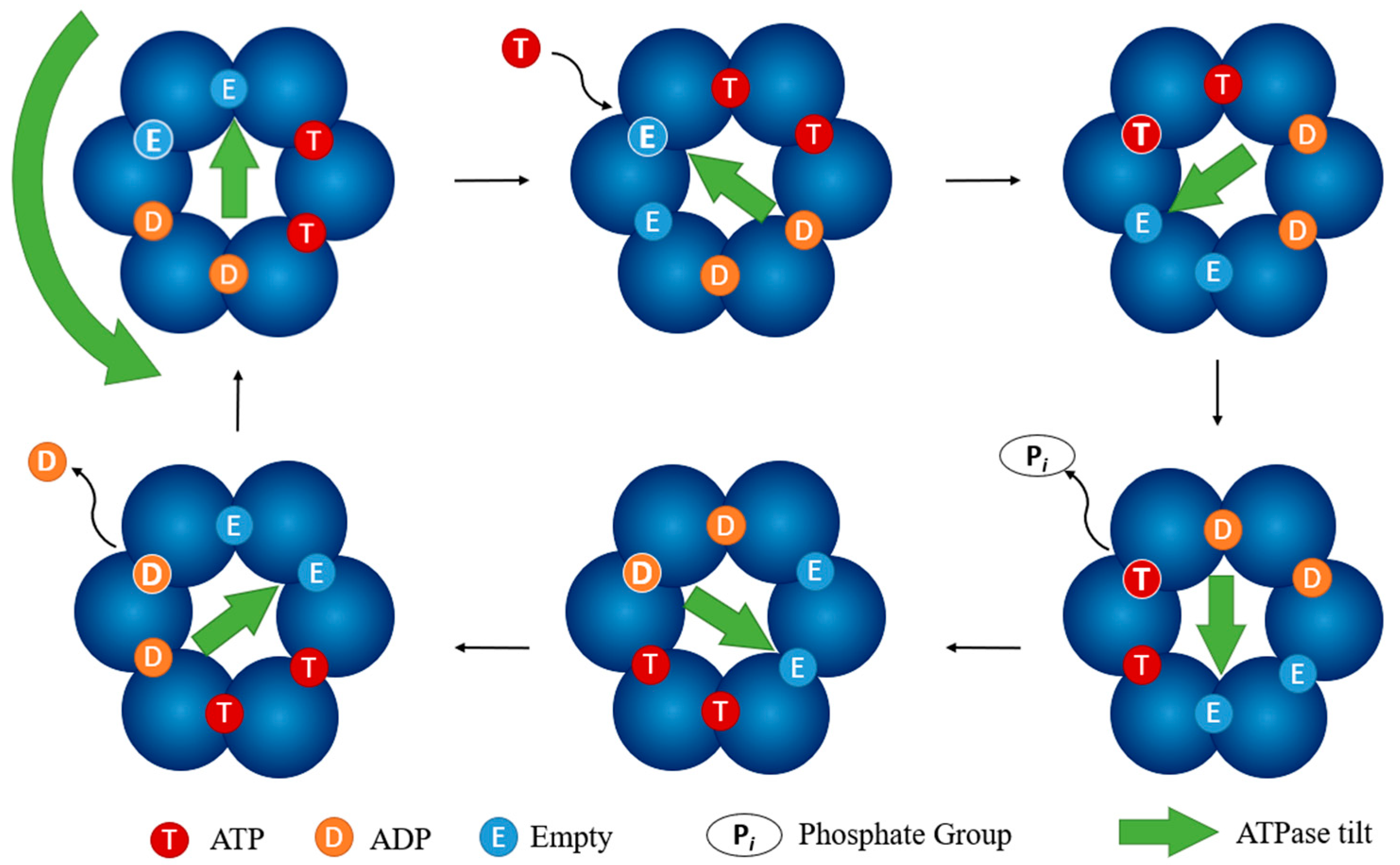
3.3. Cytoplasmic Complex: Prevention of ATPase Function
4. Inhibition of Effectors
4.1. Adhesion Inhibition
4.2. Internalization Inhibition
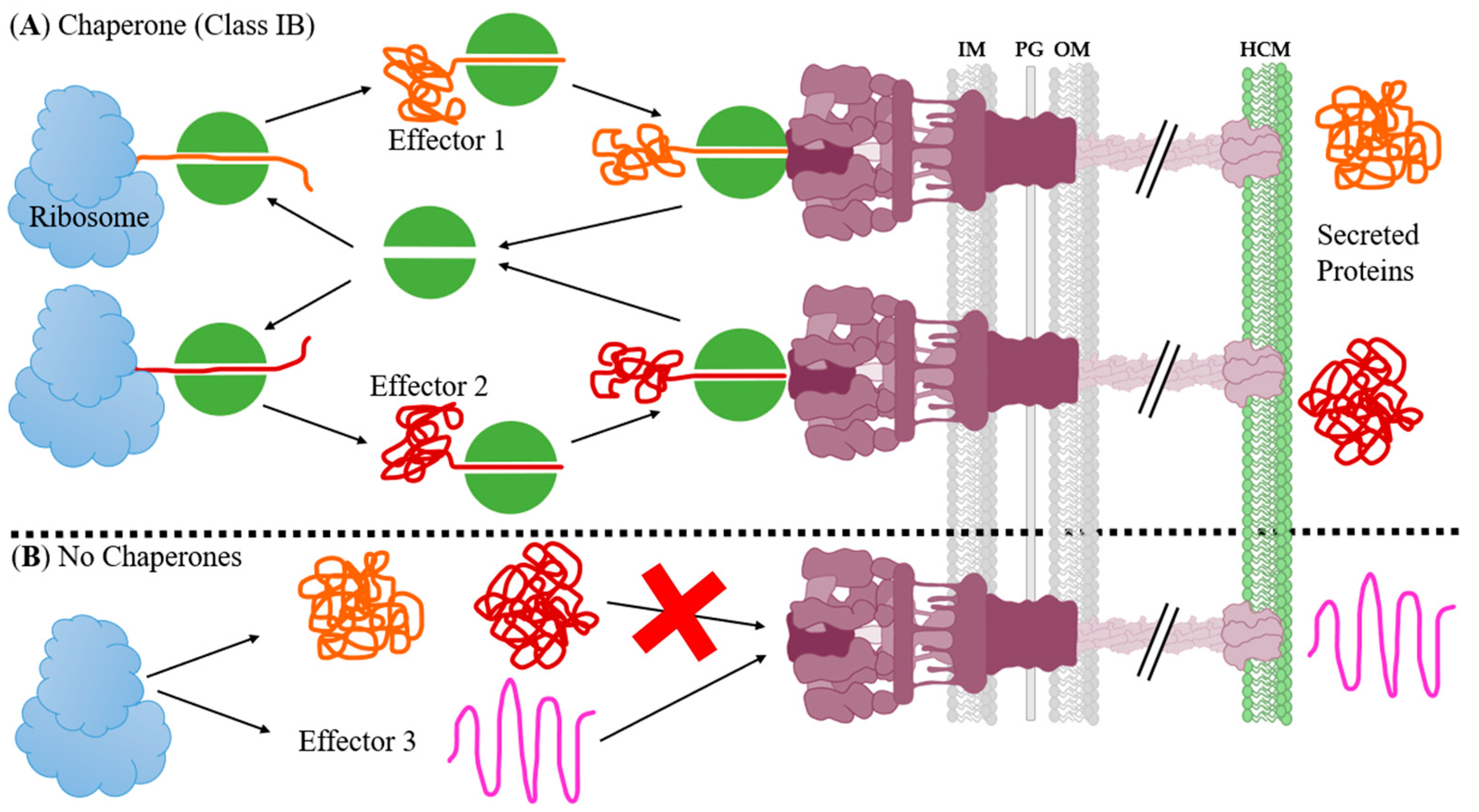
4.3. Effector Chaperone Inhibition
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- CDC National Centre for Health Statistics. Expectancy. Available online: https://www.cdc.gov/nchs/fastats/life-expectancy.htm (accessed on 12 October 2019).
- Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keyser, P.; Elofsson, M.; Rosell, S.; Wolf-Watz, H. Virulence blockers as alternatives to antibiotics: Type III secretion inhibitors against Gram-negative bacteria. J. Intern. Med. 2008, 264, 17–29. [Google Scholar] [CrossRef]
- Cornelis, G.R.; Van Gijsegem, F. Assembly and Function of Type III Secretory Systems. Annu. Rev. Microbiol. 2000, 54, 735–774. [Google Scholar] [CrossRef]
- Salmond, G.P.; Reeves, P.J. Membrance traffic wardens and protein secretion in Gram-negative bacteria. Trends Biochem. Sci. 1993, 18, 7–12. [Google Scholar] [CrossRef]
- Gaillard, M.E.; Bottero, D.; Castuma, C.E.; Basile, L.A.; Hozbor, D. Laboratory adaptation of Bordetella pertussis is associated with the loss of type three secretion system functionalityᰔ. Infect. Immun. 2011, 79, 3677–3682. [Google Scholar] [CrossRef] [Green Version]
- Stevens, M.P.; Wood, M.W.; Taylor, L.A.; Monaghan, P.; Hawes, P.; Jones, P.W.; Wallis, T.S.; Galyov, E.E. An Inv/Mxi-Spa-like type III protein secretion system in Burkholderia pseudomallei modulates intracellular behaviour of the pathogen. Mol. Microbiol. 2002, 46, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Li, Z. Conserved type III secretion system exerts important roles in Chlamydia trachomatis. Int. J. Clin. Exp. Pathol. 2014, 7, 5404–5414. [Google Scholar]
- Croxen, M.A.; Finlay, B.B. Molecular mechanisms of Escherichia coli pathogenicity. Nat. Rev. Genet. 2009, 8, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Franzin, F.M.; Sircili, M.P. Locus of enterocyte effacement: A pathogenicity island involved in the virulence of enteropathogenic and enterohemorrhagic Escherichia coli subjected to a complex network of gene regulation. Biomed Res. Int. 2015, 2015, 534738. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, K.G.; Giron, J.A.; Jerse, A.E.; McDaniel, T.K.; Donnenberg, M.S.; Kaper, J.B. Enteropathogenic Escherichia coli contains a putative type III secretion system necessary for the export of proteins involved in attaching and effacing lesion formation. Proc. Natl. Acad. Sci. USA 1995, 92, 7996–8000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontaine, A.; Arondel, J.; Sansonetti, P.J. Role of Shiga toxin in the pathogenesis of bacillary dysentery, studied by using a Tox- mutant of Shigella dysenteriae 1. Infect. Immun. 1988, 56, 3099–3109. [Google Scholar] [CrossRef] [Green Version]
- Alam, A.; Tam, V.; Hamilton, E.; Dziejman, M. VttR A and vttR B encode ToxR family proteins that mediate bile-induced expression of type three secretion system genes in a Non-O1/Non-O139 Vibrio cholerae strain. Infect. Immun. 2010, 78, 2554–2570. [Google Scholar] [CrossRef] [Green Version]
- Marketon, M.M.; DePaolo, R.W.; Debord, K.L.; Jabri, B.; Schneewind, O. Plague bacteria target immune cells during infection. Sci. 2005, 309, 1739–1741. [Google Scholar] [CrossRef] [Green Version]
- Bartra, S.S.; Lorica, C.; Qian, L.; Gong, X.; Bahnan, W.; Barreras, H.B., Jr.; Hernandez, R.; Li, Z.; Plano, G.V.; Schesser, K. Chromosomally-encoded Yersinia pestis type III secretion effector proteins promote infection in cells and in mice. Front. Cell. Infect. Microbiol. 2019, 9, 23. [Google Scholar] [CrossRef] [Green Version]
- Fast, M.D.; Tse, B.; Boyd, J.M.; Johnson, S.C. Mutations in the Aeromonas salmonicida subsp. salmonicida type III secretion system affect Atlantic salmon leucocyte activation and downstream immune responses. Fish Shellfish. Immunol. 2009, 27, 721–728. [Google Scholar] [CrossRef]
- Zheng, J.; Li, N.; Tan, Y.P.; Sivaraman, J.; Mok, Y.-K.; Mo, Z.L.; Leung, K.Y. EscC is a chaperone for the Edwardsiella tarda type III secretion system putative translocon components EseB and EseD. Microbiology 2007, 153, 1953–1962. [Google Scholar] [CrossRef] [Green Version]
- Liverman, A.D.B.; Cheng, H.C.; Trosky, J.E.; Leung, D.W.; Yarbrough, M.L.; Burdette, D.L.; Rosen, M.K.; Orth, K. Arp2/3-independent assembly of actin by Vibrio type III effector VopL. Proc. Natl. Acad. Sci. USA 2007, 104, 17117–17122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogdanove, A.J.; Wei, Z.M.; Zhao, L.; Beer, S.V. Erwinia amylovora secretes harpin via a type III pathway and contains a homolog of yopN of Yersinia spp. J. Bacteriol. 1996, 178, 1720–1730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hueck, C.J. Type III protein secretion systems in bacterial pathogens of animals and plants. Microbiol. Mol. Biol. Rev. 1998, 62, 379–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badel, J.L.; Nomura, K.; Bandyopadhyay, S.; Shimizu, R.; Collmer, A.; He, S.Y. Pseudomonas syringae pv. tomato DC3000 HopPtoM (CEL ORF3) is important for lesion formation but not growth in tomato and is secreted and translocated by the Hrp type III secretion system in a chaperone-dependent manner. Mol. Microbiol. 2003, 49, 1239–1251. [Google Scholar] [CrossRef]
- Rossier, O.; Wengelnik, K.; Hahn, K.; Bonas, U. The Xanthomonas Hrp type III system secretes proteins from plant and mammalian bacterial pathogens. Proc. Natl. Acad. Sci. USA 1999, 96, 9368–9373. [Google Scholar] [CrossRef] [Green Version]
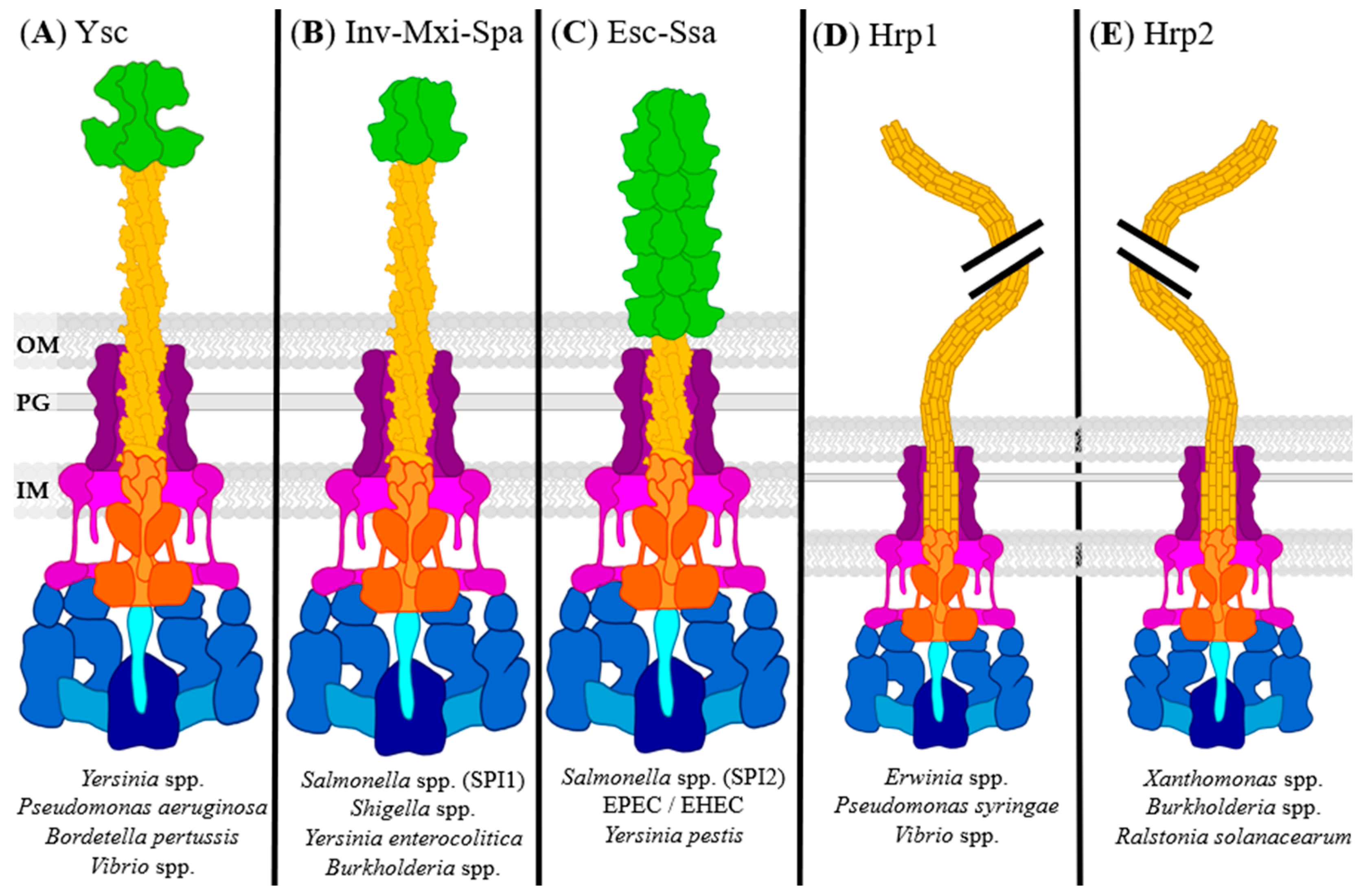
- Wagner, S.; Diepold, A. A unified nomenclature for injectisome-type type III secretion systems. Curr. Topics Microbiol. Immunol. 2020, 427, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jessen Condry, D.L.; Nilles, M.L. Identification of the targets of type III secretion system inhibitors. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2017; Volume 1531, pp. 203–211. [Google Scholar]
- Jessen, D.L.; Bradley, D.S.; Nilles, M.L. A type III secretion system inhibitor targets YopD while revealing differential regulation of secretion in calcium-blind mutants of Yersinia pestis. Antimicrob. Agents Chemother. 2014, 58, 839–850. [Google Scholar] [CrossRef] [Green Version]
- Dean, P. Functional domains and motifs of bacterial type III effector proteins and their roles in infection. FEMS Microbiol. Rev. 2011, 35, 1100–1125. [Google Scholar] [CrossRef] [Green Version]
- De Grado, M.; Abe, A.; Gauthier, A.; Steele-Mortimer, O.; DeVinney, R.; Finlay, B.B. Identification of the intimin-binding domain of Tir of enteropathogenic Escherichia coli. Cell. Microbiol. 1999, 1, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Pendergrass, H.A.; May, A.E. Natural product type III secretion system inhibitors. Antibiotics 2019, 8, 162. [Google Scholar] [CrossRef] [Green Version]
- Hume, P.J.; Singh, V.; Davidson, A.C.; Koronakis, V. Swiss army pathogen: The Salmonella entry toolkit. Front. Cell. Infect. Microbiol. 2017, 7, 348. [Google Scholar] [CrossRef] [PubMed]
- Mattock, E.; Blocker, A.J. How do the virulence factors of Shigella work together to cause disease? Front. Cell. Infect. Microbiol. 2017, 7, 64. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Mei, M.; Yu, C.; Shen, W.; Ma, L.; He, J.; Yi, L. The functions of effector proteins in Yersinia virulence. Pol. J. Microbiol. 2016, 65, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Nissim-Eliraz, E.; Nir, E.; Shoval, I.; Marsiano, N.; Nissan, I.; Shemesh, H.; Nagy, N.; Goldstein, A.M.; Gutnick, M.; Rosenshine, I.; et al. Microvascular thrombosis and ischemic enteritis in human gut xenografts infected with enteropathogenic E. Coli. Infect. Immun. 2017, 85, e00558-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browne, S.H.; Hasegawa, P.; Okamoto, S.; Fierer, J.; Guiney, D.G. Identification of Salmonella SPI-2 secretion system components required for SpvB-mediated cytotoxicity in macrophages and virulence in mice. FEMS 2008, 1, 194–201. [Google Scholar]
- Berube, B.J.; Murphy, K.R.; Torhan, M.C.; Bowlin, N.O.; Williams, J.D.; Bowlin, T.L.; Moir, D.T.; Hauser, A.R. Impact of type III secretion effectors and of phenoxyacetamide inhibitors of type III secretion on abscess formation in a mouse model of Pseudomonas aeruginosa infection. Antimicrob. Agents Chemother. 2017, 61, e01202-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marteyn, B.; West, N.P.; Browning, D.F.; Cole, J.A.; Shaw, J.G.; Palm, F.; Mounier, J.; Prévost, M.-C.; Sansonetti, P.; Tang, C.M. Modulation of Shigella virulence in response to available oxygen in vivo. Nat. Cell Biol. 2010, 465, 355–358. [Google Scholar] [CrossRef] [Green Version]
- Sampaio, S.C.F.; Moreira, F.C.; Liberatore, A.M.A.; Vieira, M.A.M.; Knobl, T.; Romão, F.T.; Hernandes, R.T.; Ferreira, C.S.A.; Ferreira, A.P.; Felipe-silva, A.; et al. Analysis of the virulence of an atypical enteropathogenic Escherichia coli strain in vitro and in vivo and the influence of type three secretion system. Biomed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [Green Version]
- Bergeron, J.R.C.; Worrall, L.J.; Sgourakis, N.G.; DiMaio, F.; Pfuetzner, R.A.; Felise, H.B.; Vuckovic, M.; Yu, A.C.; Miller, S.I.; Baker, D.; et al. A refined model of the prototypical Salmonella SPI-1 T3SS basal body reveals the molecular basis for its assembly. PLOS Pathog. 2013, 9, e1003307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernal, I.; Börnicke, J.; Heidemann, J.; Svergun, D.; Horstmann, J.A.; Erhardt, M.; Tuukkanen, A.; Uetrecht, C.; Kolbe, M. Molecular organization of soluble type III secretion system sorting platform complexes. J. Mol. Biol. 2019, 431, 3787–3803. [Google Scholar] [CrossRef] [PubMed]
- Stevens, T.C.; Ochoa, C.D.; Morrow, K.A.; Robson, M.J.; Prasain, N.; Zhou, C.; Alvarez, D.F.; Frank, D.W.; Balczon, R.; Stevens, T. The Pseudomonas aeruginosa exoenzyme Y impairs endothelial cell proliferation and vascular repair following lung injury. Am. J. Physiol. Cell. Mol. Physiol. 2014, 306, L915–L924. [Google Scholar] [CrossRef] [Green Version]
- Duncan, M.C.; Linington, R.G.; Auerbuch, V. Chemical inhibitors of the type three secretion system: Disarming bacterial pathogens. Antimicrob. Agents Chemother. 2012, 56, 5433–5441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minamino, T.; Kawamoto, A.; Kinoshita, M.; Namba, K. Molecular organization and assembly of the export apparatus off flagellar type III secretion systems. Curr. Topics Microbiol. Immunol. 2019, 427, 91–107. [Google Scholar] [CrossRef]
- Hauser, A.R. The type III secretion system of Pseudomonas aeruginosa: Infection by injection. Nat. Rev. Genet. 2009, 7, 654–665. [Google Scholar] [CrossRef] [Green Version]
- Galán, J.E. Common themes in the design and function of bacterial effectors. Cell Host Microbe 2009, 5, 571–579. [Google Scholar] [CrossRef] [Green Version]
- Golubeva, Y.A.; Sadik, A.Y.; Ellermeier, J.R.; Slauch, J.M. Integrating global regulatory input into the Salmonella Pathogenicity Island 1 type III secretion system. Genetics 2012, 190, 79–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, K.; Shinoda, N.; Hiramatsu, Y.; Ohnishi, S.; Kamitani, S.; Ogura, Y.; Hayashi, T.; Horiguchi, Y. BspR/BtrA, an anti-σ factor, regulates the ability of Bordetella bronchiseptica to cause cough in rats. mSphere 2019, 4, e00093-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, S.; Okada, R.; Tandhavanant, S.; Hiyoshi, H.; Gotoh, K.; Iida, T.; Kodama, T. Export of a Vibrio parahaemolyticus toxin by the Sec and type III secretion machineries in tandem. Nat. Microbiol. 2019, 4, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, H.; Sun, M.; Zhang, Q.; Guo, D. T3DB: An integrated database for bacterial type III secretion system. BMC Bioinform. 2012, 13, 66. [Google Scholar] [CrossRef] [Green Version]
- Barret, M.; Egan, F.; O’Gara, F. Distribution and diversity of bacterial secretion systems across metagenomic datasets. Environ. Microbiol. Rep. 2013, 5, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Aiello, D.; Williams, J.D.; Majgier-Baranowska, H.; Patel, I.; Peet, N.P.; Huang, J.; Lory, S.; Bowlin, T.L.; Moir, D.T. Discovery and characterization of inhibitors of Pseudomonas aeruginosa type III secretion. Antimicrob. Agents Chemother. 2010, 54, 1988–1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasciano, A.C.; Shaban, L.; Mecsas, J. Promises and challenges of the type three secretion system injectisome as an antivirulence target. Protein Secretion in Bacteria 2019, 8, 261–276. [Google Scholar] [CrossRef]
- Linington, R.G.; Robertson, M.; Gauthier, A.; Finlay, B.B.; van Soest, R.; Andersen, R.J. Caminoside A, an antimicrobial glycolipid isolated from the marine sponge Caminus sphaeroconia. Org. Lett. 2002, 4, 4089–4092. [Google Scholar] [CrossRef]
- May, A.E.; Khosla, C. Discovery and mechanism of type III secretion system inhibitors. Isr. J. Chem. 2013, 53, 577–587. [Google Scholar] [CrossRef]
- Kolář, M.; Urbánek, K.; Látal, T. Antibiotic selective pressure and development of bacterial resistance. Int. J. Antimicrob. Agents 2001, 17, 357–363. [Google Scholar] [CrossRef]
- Yuan, X.; Yu, M.; Yang, C.-H. Innovation and application of the type III secretion system inhibitors in plant pathogenic bacteria. Microorganisms 2020, 8, 1956. [Google Scholar] [CrossRef] [PubMed]
- Troisfontaines, P.; Cornelis, G.R. Type III secretion: More systems than you think. Physiology 2005, 20, 326–339. [Google Scholar] [CrossRef]
- Cornelis, G.R. The type III secretion injectisome. Nat. Rev. Genet. 2006, 4, 811–825. [Google Scholar] [CrossRef] [PubMed]
- Vishwakarma, V.; Pati, N.B.; Chandel, H.S.; Sahoo, S.S.; Saha, B.; Suar, M. Evaluation of Salmonella enterica serovar Typhimurium TTSS-2 deficient fur mutant as safe live-attenuated vaccine candidate for immunocompromised mice. PLoS ONE 2012, 7, e52043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodou, A.; Ankrah, D.O.; Stathopoulos, C. Toxins and secretion systems of Photorhabdus luminescens. Toxins 2010, 2, 1250–1264. [Google Scholar] [CrossRef]
- Cornelis, G.R. The Yersinia Ysc–Yop “type III” weaponry. Nat. Rev. Mol. Cell Biol. 2002, 3, 742–753. [Google Scholar] [CrossRef]
- Matsumoto, H.; Young, G.M. Translocated effectors of Yersinia. Curr. Opin. Microbiol. 2009, 12, 94–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dale, C. The insect endosymbiont Sodalis glossinidius utilizes a type III secretion system for cell invasion. Proc. Natl. Acad. Sci. USA 2001, 98, 1883–1888. [Google Scholar] [CrossRef]
- Winstanley, C.; Hales, B.A.; Hart, C.A. Evidence for the presence in Burkholderia pseudomallei of a type III secretion system-associated gene cluster. J. Med. Microbiol. 1999, 48, 649–656. [Google Scholar] [CrossRef] [Green Version]
- Miki, T.; Akiba, K.; Iguchi, M.; Danbara, H.; Okada, N. The Chromobacterium violaceum type III effector CopE, a guanine nucleotide exchange factor for Rac1 and Cdc42, is involved in bacterial invasion of epithelial cells and pathogenesis. Mol. Microbiol. 2011, 80, 1186–1203. [Google Scholar] [CrossRef]
- Daniell, S.J.; Kocsis, E.; Morris, E.; Knutton, S.; Booy, F.P.; Frankel, G. 3D structure of EspA filaments from enteropathogenic Escherichia coli. Mol. Microbiol. 2003, 49, 301–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McHugh, R.E.; O’Boyle, N.; Connolly, J.P.R.; Hoskisson, P.A.; Roe, A.J. Characterization of the mode of action of aurodox, a type III secretion system inhibitor from Streptomyces goldiniensis. Infect. Immun. 2018, 87, e00595-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallen, M.J.; Beatson, S.A.; Bailey, C.M. Bioinformatics, genomics and evolution of non-flagellar type-III secretion systems: A Darwinian perpective. FEMS Microbiol. Rev. 2005, 29, 201–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfano, J.R.; Collmer, A. Type III secretion system effector proteins: Double agents in bacterial disease and plant defense. Annu. Rev. Phytopathol. 2004, 42, 385–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfano, J.R.; Collmer, A. The type III (Hrp) secretion pathway of plant pathogenic bacteria: Trafficking harpins, Avr proteins, and death. J. Bacteriol. 1997, 179, 5655–5662. [Google Scholar] [CrossRef] [Green Version]
- Rainbow, L.; Hart, C.A.; Winstanley, C. Distribution of type III secretion gene clusters in Burkholderia pseudomallei, B. thailandensis and B. mallei. J. Med Microbiol. 2002, 51, 374–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makino, K.; Oshima, K.; Kurokawa, K.; Yokoyama, K.; Uda, T.; Tagomori, K.; Iijima, Y.; Najima, M.; Nakano, M.; Yamashita, A.; et al. Genome sequence of Vibrio parahaemolyticus: A pathogenic mechanism distinct from that of V. cholerae. Lancet 2003, 361, 743–749. [Google Scholar] [CrossRef]
- Feng, J.; Yuan, F.; Hsu, S.; Song, W. Transcriptomic analysis provides insights into the molecular mechanisms of epigallocatechin-3-gallate to attenuate schistosomiasis hepatic fibrosis in mice. Int. J. Agric. Biol. 2019, 21, 314–324. [Google Scholar]
- Marie, C.; Broughton, W.J.; Deakin, W.J. Rhizobium type III secretion systems: Legume charmers or alarmers? Curr. Opin. Plant Biol. 2001, 4, 336–342. [Google Scholar] [CrossRef]
- Moriya, N.; Minamino, T.; Hughes, K.T.; Macnab, R.M.; Namba, K. The type III flagellar export specificity switch is dependent on FliK ruler and a molecular clock. J. Mol. Biol. 2006, 359, 466–477. [Google Scholar] [CrossRef]
- Bergeron, J.R.C.; Fernández, L.; Wasney, G.A.; Vuckovic, M.; Reffuveille, F.; Hancock, R.E.W.; Strynadka, N.C.J. The structure of a type 3 secretion system (T3SS) ruler protein suggests a molecular mechanism for needle length sensing. J. Biol. Chem. 2016, 291, 1676–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sal-Man, N.; Deng, W.; Finlay, B.B. EscI: A crucial component of the type III secretion system forms the inner rod structure in enteropathogenic Escherichia coli. Biochem. J. 2012, 442, 119–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burghout, P.; Van Boxtel, R.; Van Gelder, P.; Ringler, P.; Mu, S.A.; Tommassen, J.; Koster, M. Structure and electrophysiological properties of the YscC secretin from the type III secretion system of Yersinia enterocolitica. J. Bacteriol. 2004, 186, 4645–4654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekiya, K.; Ohishi, M.; Ogino, T.; Tamano, K.; Sasakawa, C.; Abe, A. Supermolecular structure of the enteropathogenic Escherichia coli type III secretion system and its direct interaction with the EspA-sheath-like structure. Proc. Natl. Acad. Sci. USA 2001, 98, 11638–11643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, R.K.; Shaw, R.K.; Daniell, S.; Knutton, S.; Frankel, G. Role of EscF, a putative needle complex protein, in the type III protein translocation system of enteropathogenic Escherichia coli. Cell. Microbiol. 2001, 3, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Mueller, C.A.; Broz, P.; Cornelis, G.R. The type III secretion system tip complex and translocon. Mol. Microbiol. 2008, 68, 1085–1095. [Google Scholar] [CrossRef] [PubMed]
- Delahay, R.M.; Knutton, S.; Shaw, R.K.; Hartland, E.L.; Pallen, M.J.; Frankel, G. The coiled-coil domain of EspA is essential for the assembly of the type III secretion translocon on the surface of enteropathogenic Escherichia coli. J. Biol. Chem. 1999, 274, 35969–35974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knutton, S.; Rosenshine, M.; Pallen, M.J.; Nisan, I.; Neves, B.C.; Bain, C.; Wolff, C.; Dougan, G.; Frankel, G. A novel EspA-associated surface organelle of enteropathogenic Escherichia coli involved in protein translocation into epithelial cells. EMBO J. 1998, 17, 2166–2176. [Google Scholar] [CrossRef]
- Shaw, R.K.; Daniell, S.; Ebel, F.; Frankel, G.; Knutton, S. EspA filament-mediated protein translocation into red blood cells. Cell. Microbiol. 2001, 3, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Ide, T.; Laarmann, S.; Greune, L.; Schillers, H.; Oberleithner, H.; Schmidt, M.A. Characterization of translocation pores inserted into plasma membranes by type III-secreted Esp proteins of enteropathogenic Escherichia coli. Cell. Microbiol. 2001, 3, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Donnenberg, M.S. Interactions and predicted host membrane topology of the enteropathogenic Escherichia coli translocator protein EspB. J. Bacteriol. 2011, 193, 2972–2980. [Google Scholar] [CrossRef] [Green Version]
- Larzábal, M.; Baldoni, H.A.; Suvire, F.D.; Curto, L.M.; Gomez, G.E.; Da Silva, W.M.; Giudicessi, S.L.; Camperi, S.A.; Delfino, J.M.; Cataldi, A.A.; et al. An inhibitory mechanism of action of coiled-coil peptides against type three secretion system from enteropathogenic Escherichia coli. J. Pept. Sci. 2019, 25, e3149. [Google Scholar] [CrossRef] [PubMed]
- Kaspar, A.A.; Reichert, J.M. Future directions for peptide therapeutics development. Drug Discov. Today 2013, 18, 807–817. [Google Scholar] [CrossRef]
- Nordfelth, R.; Elofsson, M. Small-molecule inhibitors specically targeting type III secretion. Infect. Immun. 2005, 73, 3104–3114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muschiol, S.; Bailey, L.; Gylfe, Å.; Sundin, C.; Hultenby, K.; Bergström, S.; Elofsson, M.; Wolf-Watz, H.; Normark, S.; Henriques-Normark, B. A small-molecule inhibitor of type III secretion inhibits different stages of the infectious cycle of Chlamydia trachomatis. Proc. Natl. Acad. Sci. USA 2006, 103, 14566–14571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, L.; Gylfe, Å.; Sundin, C.; Muschiol, S.; Elofsson, M.; Nordström, P.; Henriques-Normark, B.; Lugert, R.; Waldenström, A.; Wolf-Watz, H.; et al. Small molecule inhibitors of type III secretion in Yersinia block the Chlamydia pneumoniae infection cycle. FEBS Lett. 2007, 581, 587–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, D.L.; Layton, A.N.; Field, T.R.; Bowen, A.J.; Wolf-Watz, H.; Elofsson, M.; Stevens, M.P.; Galyov, E.E. Inhibition of type III secretion in Salmonella enterica serovar Typhimurium by small-molecule inhibitors. Antimicrob. Agents Chemother. 2007, 51, 2631–2635. [Google Scholar] [CrossRef] [Green Version]
- Veenendaal, A.K.J.; Sundin, C.; Blocker, A.J. Small-molecule type III secretion system inhibitors block assembly of the Shigella type III secretion. J. Bacteriol. 2009, 191, 563–570. [Google Scholar] [CrossRef] [Green Version]
- Tree, J.J.; Wang, D.; McInally, C.; Mahajan, A.; Layton, A.; Houghton, I.; Elofsson, M.; Stevens, M.P.; Gally, D.L.; Roe, A.J. Characterization of the effects of salicylidene acylhydrazide compounds on type III secretion in Escherichia coli O157:H7. Infect. Immun. 2009, 77, 4209–4220. [Google Scholar] [CrossRef] [Green Version]
- Tao, H.; Fan, S.-S.; Jiang, S.; Xiang, X.; Yan, X.; Zhang, L.-H.; Cui, Z.-N. Small molecule inhibitors specifically targeting the type III secretion system of Xanthomonas oryzae on rice. Int. J. Mol. Sci. 2019, 20, 971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, S.; Tian, F.; Li, J.; Hutchins, W.; Chen, H.; Yang, F.; Yuan, X.; Cui, Z.; Yang, C.-H.; He, C. Identification of phenolic compounds that suppress the virulence of Xanthomonas oryzae on rice via the type III secretion system. Mol. Plant Pathol. 2016, 18, 555–568. [Google Scholar] [CrossRef]
- Yang, F.; Korban, S.S.; Pusey, P.L.; Elofsson, M.; Sundin, G.W.; Zhao, Y. Small-molecule inhibitors suppress the expression of both type III secretion and amylovoran biosynthesis genes in Erwinia amylovora. Mol. Plant Pathol. 2014, 15, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Bowlin, N.O.; Williams, J.D.; Knoten, C.A.; Torhan, M.C.; Tashjian, T.F.; Li, B.; Aiello, D.; Mecsas, J.; Hauser, A.R.; Peet, N.P.; et al. Mutations in the Pseudomonas aeruginosa needle protein gene pscF confer resistance to phenoxyacetamide inhibitors of the type III secretion system. Antimicrob. Agents Chemother. 2014, 58, 2211–2220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
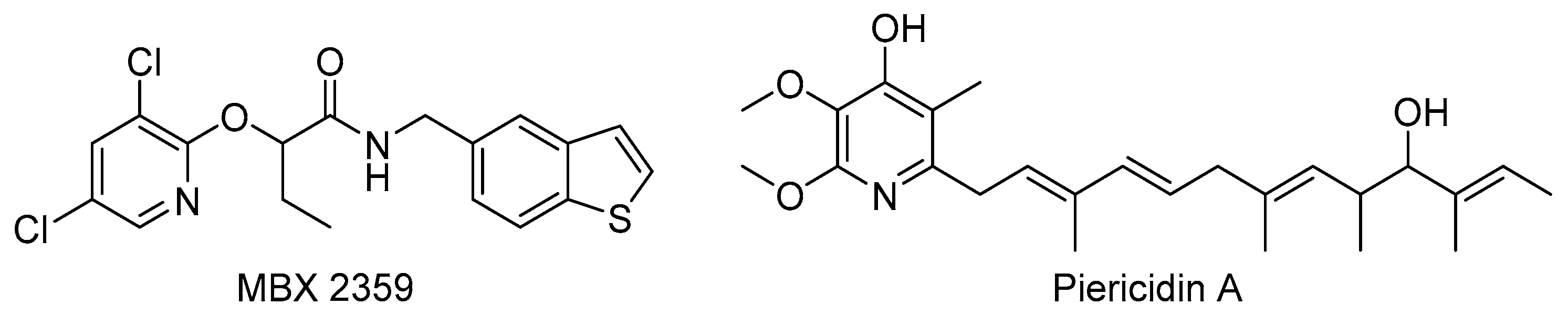
- Duncan, M.C.; Wong, W.R.; Dupzyk, A.J.; Bray, W.M.; Linington, R.G.; Auerbuch, V. An NF-κB-based high-throughput screen identifies piericidins as inhibitors of the Yersinia pseudotuberculosis type III secretion system. Antimicrob. Agents Chemother. 2014, 58, 1118–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, J.M.; Duncan, M.C.; Johnson, K.S.; Diepold, A.; Lam, H.; Dupzyk, A.J.; Martin, L.R.; Wong, W.R.; Armitage, J.P.; Linington, R.G.; et al. Piericidin A1 blocks Yersinia Ysc type III secretion system needle assembly. mSphere 2017, 2, e00030-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auerbuch, V.; Golenbock, D.T.; Isberg, R.R. Innate immune recognition of Yersinia pseudotuberculosis type III secretion. PLoS Pathog. 2009, 5, e1000686. [Google Scholar] [CrossRef] [Green Version]
- Sawa, T.; Kinoshita, M.; Inoue, K.; Ohara, J.; Moriyama, K. Immunoglobulin for treating bacterial infections: One more mechanism of action. Antibodies 2019, 8, 52. [Google Scholar] [CrossRef] [Green Version]
- Pendergrass, H.A.; May, A.E. Delivery of heterologous proteins, enzymes, and antigens via the bacterial type III secretion system. Microorganisms 2020, 8, 777. [Google Scholar] [CrossRef]
- Hotinger, J.A.; May, A.E. Antibodies inhibiting the type III secretion system of Gram-negative pathogenic bacteria. Antibodies 2020, 9, 35. [Google Scholar] [CrossRef]
- Huang, Y.-M.; Hu, W.; Rustandi, E.; Chang, K.; Yusuf-Makagiansar, H.; Ryll, T. Maximizing productivity of CHO cell-based fed-batch culture using chemically defined media conditions and typical manufacturing equipment. Biotechnol. Prog. 2010, 26, 1400–1410. [Google Scholar] [CrossRef]
- Sittner, A.; Mechaly, A.; Vitner, E.; Aftalion, M.; Levy, Y.; Levy, H.; Mamroud, E.; Fisher, M. Improved production of monoclonal antibodies against the LcrV antigen of Yersinia pestis using FACS-aided hybridoma selection. J. Biol. Methods 2018, 5, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Embry, A.; Meng, X.; Cantwell, A.; Dube, P.H.; Xiang, Y. Enhancement of immune response to an antigen delivered by vaccinia virus by displaying the antigen on the surface of intracellular mature virion. Vaccine 2011, 29, 5331–5339. [Google Scholar] [CrossRef] [Green Version]
- Cowan, C.; Philipovskiy, A.V.; Wulff-Strobel, C.R.; Ye, Z.; Straley, S.C. Anti-LcrV antibody inhibits delivery of Yops by Yersinia pestis KIM5 by directly promoting phagocytosis. Infect. Immun. 2005, 73, 6127–6137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, M.; Shimizu, M.; Akiyama, K.; Kato, H.; Moriyama, K.; Sawa, T. Epidemiological survey of serum titers from adults against various Gram-negative bacterial V-antigens. PLOS ONE 2020, 15, e0220924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abramov, V.M.; Kosarev, I.V.; Motin, V.L.; Khlebnikov, V.S.; Vasilenko, R.N.; Sakulin, V.K.; Machulin, A.V.; Uversky, V.N.; Karlyshev, A.V. Binding of LcrV protein from Yersinia pestis to human T-cells induces apoptosis, which is completely blocked by specific antibodies. Int. J. Biol. Macromol. 2019, 122, 1062–1070. [Google Scholar] [CrossRef] [Green Version]
- Sawa, T.; Ito, E.; Nguyen, V.H.; Haight, M. Anti-PcrV antibody strategies against virulent Pseudomonas aeruginosa. Hum. Vaccines Immunother. 2014, 10, 2843–2852. [Google Scholar] [CrossRef] [Green Version]
- DiGiandomenico, A.; Keller, A.E.; Gao, C.; Rainey, G.J.; Warrener, P.; Camara, M.M.; Bonnell, J.; Fleming, R.; Bezabeh, B.; DiMasi, N.; et al. A multifunctional bispecific antibody protects against Pseudomonas aeruginosa. Sci. Transl. Med. 2014, 6, 262ra155. [Google Scholar] [CrossRef]
- Tabor, D.E.; Oganesyan, V.; Keller, A.E.; Yu, L.; McLaughlin, R.E.; Song, E.; Warrener, P.; Rosenthal, K.; Esser, M.; Qi, Y.; et al. Pseudomonas aeruginosa PcrV and Psl, the molecular targets of bispecific antibody MEDI3902, are conserved among diverse global clinical isolates. J. Infect. Dis. 2018, 218, 1983–1994. [Google Scholar] [CrossRef]
- Le, H.N.; Quetz, J.S.; Tran, V.G.; Le, V.T.M.; Aguiar-Alves, F.; Pinheiro, M.G.; Cheng, L.; Yu, L.; Sellman, B.R.; Stover, C.K.; et al. MEDI3902 Correlates of protection against severe Pseudomonas aeruginosa pneumonia in a rabbit acute pneumonia model. Antimicrob. Agents Chemother. 2018, 62, e02565-17. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.O.; Yu, X.Q.; Robbie, G.J.; Wu, Y.; Shoemaker, K.; Yu, L.; DiGiandomenico, A.; Keller, A.E.; Anude, C.; Hernandez-Illas, M.; et al. Phase 1 study of MEDI3902, an investigational anti–Pseudomonas aeruginosa PcrV and Psl bispecific human monoclonal antibody, in healthy adults. Clin. Microbiol. Infect. 2019, 25, 629.e1–629.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chastre, J.; François, B.; Bourgeois, M.; Komnos, A.; Ferrer, R.; Rahav, G.; De Schryver, N.; Lepape, A.; Koksal, I.; Luyt, C.-E.; et al. 635. Efficacy, pharmacokinetics (PK), and safety profile of MEDI3902, an anti-Pseudomonas aeruginosa bispecific human monoclonal antibody in mechanically ventilated intensive care unit patients; Results of the phase 2 EVADE study conducted by the public-private COMBACTE-MAGNET consortium in the innovative medicines initiative (IMI) program. Open Forum Infect. Dis. 2020, 7, S377–S378. [Google Scholar] [CrossRef]
- Le, H.N.; Tran, V.G.; Vu, T.T.T.; Gras, E.; Le, V.T.M.; Pinheiro, M.G.; Aguiar-Alves, F.; Schneider-Smith, E.; Carter, H.C.; Sellman, B.R.; et al. Treatment efficacy of MEDI3902 in Pseudomonas aeruginosa bloodstream infection and acute pneumonia rabbit models. Antimicrob. Agents Chemother. 2019, 63, e00710-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
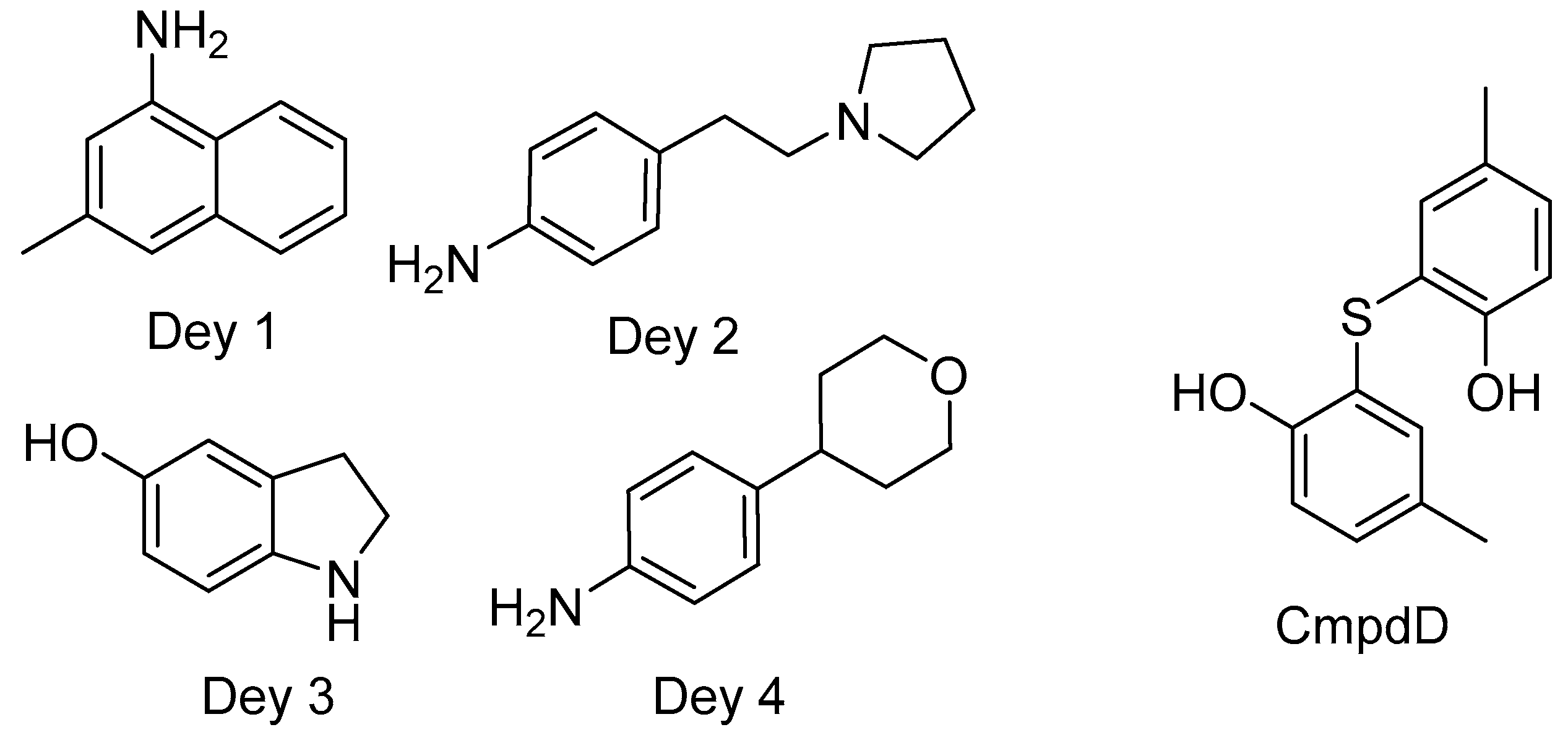
- Dey, S.; Anbanandam, A.; Mumford, B.E.; De Guzman, R.N. Characterization of small molecule scaffolds that bind to the Shigella type III secretion system protein IpaD. ChemMedChem 2017, 12, 1534–1541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barta, M.L.; Guragain, M.; Adam, P.; Dickenson, N.E.; Patil, M.; Geisbrecht, B.V.; Picking, W.L.; Picking, W.D. Identification of the bile salt binding site on ipad from Shigella flexneri and the influence of ligand binding on IpaD structure. Proteins Struct. Funct. Bioinform. 2011, 80, 935–945. [Google Scholar] [CrossRef] [Green Version]
- Dickenson, N.E.; Zhang, L.; Epler, C.R.; Adam, P.R.; Picking, W.L.; Picking, W.D. Conformational changes in IpaD from Shigella flexneri upon binding bile salts provide insight into the second step of type III secretion. Biochemistry 2011, 50, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Stensrud, K.F.; Adam, P.R.; La Mar, C.D.; Olive, A.J.; Lushington, G.H.; Sudharsan, R.; Shelton, N.L.; Givens, R.S.; Picking, W.L.; Picking, W.D. Deoxycholate interacts with IpaD of Shigella flexneri in inducing the recruitment of IpaB to the type III secretion apparatus needle tip. J. Biol. Chem. 2008, 283, 18646–18654. [Google Scholar] [CrossRef] [Green Version]
- Dickenson, N.E.; Arizmendi, O.; Patil, M.K.; Toth, R.T.; Middaugh, C.R.; Picking, W.D.; Picking, W.L. N-Terminus of IpaB provides a potential anchor to the Shigella type III secretion system tip complex protein IpaD. Biochemistry 2013, 52, 8790–8799. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Zetterström, C.E.; Gabrielsen, M.; Beckham, K.S.H.; Tree, J.J.; Macdonald, S.E.; Byron, O.; Mitchell, T.J.; Gally, D.L.; Herzyk, P.; et al. Identification of bacterial target proteins for the salicylidene acylhydrazide class of virulence-blocking compounds. J. Biol. Chem. 2011, 286, 29922–29931. [Google Scholar] [CrossRef] [Green Version]
- Zambelloni, R.; Connolly, J.P.R.; Huerta Uribe, A.; Burgess, K.; Marquez, R.; Roe, A.J. Novel compounds targeting the enterohemorrhagic Escherichia coli type three secretion system reveal insights into mechanisms of secretion inhibition. Mol. Microbiol. 2017, 105, 606–619. [Google Scholar] [CrossRef]
- Lloyd, S.A.; Sjöström, M.; Andersson, S.; Wolf-Watz, H. Molecular characterization of type III secretion signals via analysis of synthetic N-terminal amino acid sequences. Mol. Microbiol. 2002, 43, 51–59. [Google Scholar] [CrossRef]
- Stebbins, C.E.; Galán, J.E. Maintenance of an unfolded polypeptide by a cognate chaperone in bacterial type III secretion. Nat. Cell Biol. 2001, 414, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Wilharm, G.; Dittmann, S.; Schmid, A.; Heesemann, J. On the role of specific chaperones, the specific ATPase, and the proton motive force in type III secretion. Int. J. Med. Microbiol. 2007, 297, 27–36. [Google Scholar] [CrossRef]
- Woestyn, S.; Sory, M.-P.; Boland, A.; Lequenne, O.; Cornelis, G.R. The cytosolic SycE and SycH chaperones of Yersinia protect the region of YopE and YopH involved in translocation across eukaryotic cell membranes. Mol. Microbiol. 1996, 20, 1261–1271. [Google Scholar] [CrossRef] [PubMed]
- Thomas, N.A.; Deng, W.; Puente, J.L.; Frey, E.A.; Yip, C.K.; Strynadka, N.C.J.; Finlay, B.B. CesT is a multi-effector chaperone and recruitment factor required for the efficient type III secretion of both LEE- and non-LEE-encoded effectors of enteropathogenic Escherichia coli. Mol. Microbiol. 2005, 57, 1762–1779. [Google Scholar] [CrossRef] [PubMed]
- Burkinshaw, B.J.; Strynadka, N.C. Assembly and structure of the T3SS. Biochim. Biophys. Acta Bioenerg. 2014, 1843, 1649–1663. [Google Scholar] [CrossRef] [Green Version]
- Sal-Man, N.; Setiaputra, D.; Scholz, R.; Deng, W.; Yu, A.C.Y.; Strynadka, N.C.J.; Finlay, B.B. EscE and EscG are cochaperones for the type III needle protein EscF of enteropathogenic Escherichia coli. J. Bacteriol. 2013, 195, 2481–2489. [Google Scholar] [CrossRef] [Green Version]
- Yip, C.K.; Finlay, B.B.; Strynadka, N.C.J. Structural characterization of a type III secretion system filament protein in complex with its chaperone. Nat. Struct. Mol. Biol. 2005, 12, 75–81. [Google Scholar] [CrossRef]
- Creasey, E.A.; Friedberg, D.; Shaw, R.K.; Umanski, T.; Knutton, S.; Rosenshine, I.; Frankel, G. CesAB is an enteropathogenic Escherichia coli chaperone for the type-III translocator proteins EspA and EspB. Microbiology 2003, 149, 3639–3647. [Google Scholar] [CrossRef] [Green Version]
- Ku, C.P.; Lio, J.C.W.; Wang, S.H.; Lin, C.N.; Syu, W.J. Identification of a third EspA-binding protein that forms part of the type III secretion system of enterohemorrhagic Escherichia coli. J. Biol. Chem. 2009, 284, 1686–1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, S.; Roversi, P.; Espina, M.; Olive, A.; Deane, J.E.; Birket, S.; Field, T.; Picking, W.D.; Blocker, A.J.; Galyov, E.E.; et al. Self-chaperoning of the type III secretion system needle tip proteins IpaD and BipD. J. Biol. Chem. 2006, 282, 4035–4044. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, V.S.; Jobichen, C.; Tan, K.W.; Tan, Y.W.; Chan, S.L.; Ramesh, K.; Yuan, Y.; Hong, Y.; Seetharaman, J.; Leung, K.Y.; et al. Structure of AcrH-AopB chaperone-translocator complex reveals a role for membrane hairpins in type III secretion system translocon assembly. Structure 2015, 23, 2022–2031. [Google Scholar] [CrossRef]
- Chatterjee, C.; Kumar, S.; Chakraborty, S.; Tan, Y.W.; Leung, K.Y.; Sivaraman, J.; Mok, Y.-K. Crystal structure of the heteromolecular chaperone, AscE-AscG, from the type III secretion system in Aeromonas hydrophila. PLoS ONE 2011, 6, e19208. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, J.; Miletic, S.; Gaildry, T.; Chin-Fatt, A.; Menassa, R. Co-expression with the type 3 secretion chaperone CesT from enterohemorrhagic E. coli increases accumulation of recombinant Tir in plant chloroplasts. Front. Plant Sci. 2017, 8, 283. [Google Scholar] [CrossRef] [Green Version]
- Wainwright, L.A.; Kaper, J.B. EspB and EspD require a specific chaperone for proper secretion from enteropathogenic Escherichia coil. Mol. Microbiol. 1998, 27, 1247–1260. [Google Scholar] [CrossRef]
- Neves, B.C.; Mundy, R.; Petrovska, L.; Dougan, G.; Knutton, S.; Frankel, G. CesD2 of Enteropathogenic Escherichia coli is a second chaperone for the type III secretion translocator protein EspD. Infect. Immun. 2003, 71, 2130–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, M.S.-W.; Kao, H.-C.; Lin, C.-N.; Syu, W.-J. Gene l0017 encodes a second chaperone for EspA of enterohaemorrhagic Escherichia coli O157: H7. Microbiology 2008, 154, 1094–1103. [Google Scholar] [CrossRef] [Green Version]
- Schreiner, M.; Niemann, H.H. Crystal structure of the Yersinia enterocolitica type III secretion chaperone SycD in complex with a peptide of the minor translocator YopD. BMC Struct. Biol. 2012, 12, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanao, M.; Ricard-Blum, S.; Di Guilmi, A.M.; Lemaire, D.; Lascoux, D.; Chabert, J.; Attree, I.; Dessen, A. Type III secretion proteins PcrV and PcrG from Pseudomonas aeruginosa form a 1:1 complex through high affinity interactions. BMC Microbiol. 2003, 3, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plé, S.; Job, V.; Dessen, A.; Attree, I. Cochaperone interactions in export of the type III needle component PscF of Pseudomonas aeruginosa. J. Bacteriol. 2010, 192, 3801–3808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darwin, K.H.; Miller, V.L. The putative invasion protein chaperone SicA acts together with InvF to activate the expression of Salmonella typhimurium virulence genes. Mol. Microbiol. 2000, 35, 949–960. [Google Scholar] [CrossRef]
- Kaur, K.; Chatterjee, S.; De Guzman, R.N. Characterization of the Shigella and Salmonella type III secretion system tip-translocon protein-protein interaction by paramagnetic relaxation enhancement. ChemBioChem 2016, 17, 745–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, Y.; Miki, T.; Ono, S.; Haneda, T.; Ito, M.; Okada, N. Functional characterization of the type III secretion ATPase SsaN encoded by Salmonella Pathogenicity Island 2. PLoS ONE 2014, 9, e94347. [Google Scholar] [CrossRef]
- Zurawski, D.V.; Stein, M.A. SseA acts as the chaperone for the SseB component of the Salmonella Pathogenicity Island 2 translocon. Mol. Microbiol. 2003, 47, 1341–1351. [Google Scholar] [CrossRef]
- Takaya, A.; Takeda, H.; Tashiro, S.; Kawashima, H.; Yamamoto, T. Chaperone-mediated secretion switching from early to middle substrates in the type III secretion system encoded by Salmonella pathogenicity island 2. J. Biol. Chem. 2019, 294, 3783–3793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Becerra, F.J.; Chen, X.; Dickenson, N.E.; Choudhari, S.P.; Harrison, K.; Clements, J.D.; Picking, W.D.; Van De Verg, L.L.; Walker, R.I.; Picking, W.L. Characterization of a novel fusion protein from IpaB and IpaD of Shigella spp. and its potential as a pan-Shigella vaccine. Infect. Immun. 2013, 81, 4470–4477. [Google Scholar] [CrossRef] [Green Version]
- Cherradi, Y.; Hachani, A.; Allaoui, A. Spa13 of Shigella flexneri has a dual role: Chaperone escort and export gate-activator switch of the type III secretion system. Microbiology 2014, 160, 130–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukerjea, R.; Ghosh, P. Functionally essential interaction between Yersinia YscO and the T3S4 domain of YscP. J. Bacteriol. 2013, 195, 4631–4638. [Google Scholar] [CrossRef] [Green Version]
- Chaudhury, S.; Souza, C.A.; Plano, G.V.; De Guzman, R.N. The LcrG tip chaperone protein of the Yersinia pestis type III secretion system is partially folded. J. Mol. Biol. 2015, 427, 3096–3109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, C.D.A.; Richards, K.L.; Park, Y.; Schwartz, M.; Garcia, J.T.; Bartra, S.S.; Plano, G.V. The YscE/YscG chaperone and YscF N-terminal sequences target YscF to the Yersinia pestis type III secretion apparatus. Microbiology 2018, 164, 338–348. [Google Scholar] [CrossRef]
- Francis, M.S.; Lloyd, S.A.; Wolf-Watz, H. The type III secretion chaperone LcrH co-operates with YopD to establish a negative, regulatory loop for control of Yop synthesis in Yersinia pseudotuberculosis. Mol. Microbiol. 2001, 42, 1075–1093. [Google Scholar] [CrossRef] [PubMed]
- Ngo, T.D.; Plé, S.; Thomas, A.; Barette, C.; Fortuné, A.; Bouzidi, Y.; Fauvarque, M.-O.; de Freitas, R.P.; Hilário, F.F.; Attrée, I.; et al. Chimeric protein–protein interface inhibitors allow efficient inhibition of type III secretion machinery and Pseudomonas aeruginosa virulence. ACS Infect. Dis. 2019, 5, 1843–1854. [Google Scholar] [CrossRef] [PubMed]
- Diepold, A.; Armitage, J.P. Type III secretion systems: The bacterial flagellum and the injectisome. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20150020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abby, S.S.; Rocha, E.P.C. The non-flagellar type III secretion system evolved from the bacterial flagellum and diversified into host-cell adapted systems. PLoS Genet. 2012, 8, e1002983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, W.R. An introduction to sequence similarity (“homology”) searching. Curr. Protoc. Bioinform. 2013, 42, 3.1.1–3.1.8. [Google Scholar] [CrossRef]
- Korotkov, K.V.; Gonen, T.; Hol, W.G.J. Secretins: Dynamic channels for protein transport across membranes. Trends Biochem. Sci. 2011, 36, 433–443. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, A.; Puente, J.L.; Finlay, B.B. Secretin of the enteropathogenic Escherichia coli type III secretion system requires components of the type III apparatus for assembly and localization. Infect. Immun. 2003, 71, 3310–3319. [Google Scholar] [CrossRef] [Green Version]
- Kauppi, A.M.; Nordfelth, R.; Uvell, H.; Wolf-Watz, H.; Elofsson, M. Targeting bacterial virulence: Inhibitors of type III secretion in Yersinia. Chem. Biol. 2003, 10, 241–249. [Google Scholar] [CrossRef] [Green Version]
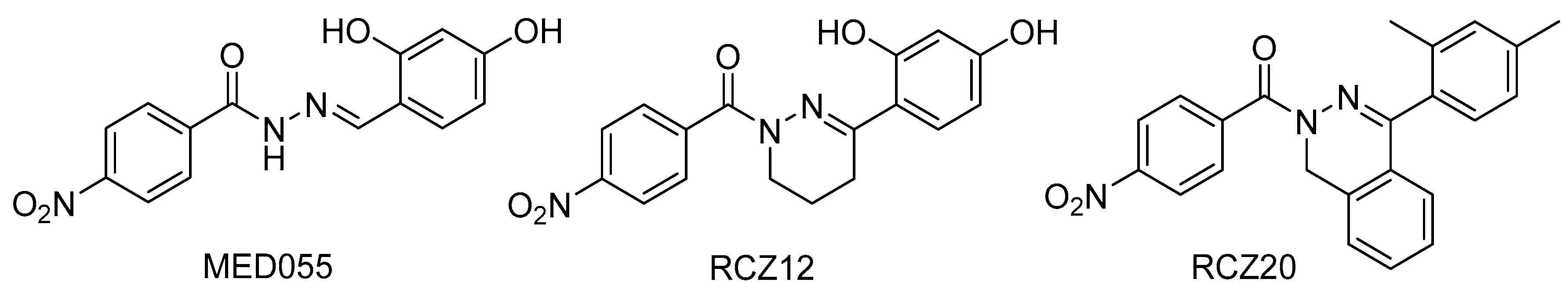
- Negrea, A.; Bjur, E.; Ygberg, S.E.; Elofsson, M.; Wolf-Watz, H.; Rhen, M. Salicylidene acylhydrazides that affect type III protein secretion in Salmonella enterica serovar Typhimurium. Antimicrob. Agents Chemother. 2007, 51, 2867–2876. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Argudo, I.; Veenendaal, A.K.J.; Liu, X.; Roehrich, A.D.; Ronessen, M.C.; Franzoni, G.; van Rietschoten, K.N.; Morimoto, Y.V.; Saijo-Hamano, Y.; Avison, M.B.; et al. Isolation of Salmonella mutants resistant to the inhibitory effect of salicylidene acylhydrazides on flagella-mediated motility. PLoS ONE 2013, 8, e52179. [Google Scholar]
- Wagner, S.; Grin, I.; Malmsheimer, S.; Singh, N.; Torres-Vargas, C.E.; Westerhausen, S. Bacterial type III secretion systems: A complex device for the delivery of bacterial effector proteins into eukaryotic host cells. FEMS Microbiol. Lett. 2018, 365, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseytin, I.; Madar, A.; Mitrovic, B.; Deng, W.; Finlay, B.B.; Sal-Man, N. The third transmembrane domain of EscR is critical for function of the enteropathogenic Escherichia coli type III secretion system. mSphere 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Elliott, S.J.; Wainwright, L.A.; McDaniel, T.K.; Jarvis, K.G.; Deng, Y.K.; Lai, L.C.; McNamara, B.P.; Donnenberg, M.S.; Kaper, J.B. The complete sequence of the locus of enterocyte effacement (LEE) from enteropathogenic Escherichia coli E2348/69. Mol. Microbiol. 1998, 28, 1–4. [Google Scholar] [CrossRef]
- Tseytin, I.; Mitrovic, B.; David, N.; Langenfeld, K.; Zarivach, R.; Diepold, A.; Sal-Man, N. The role of the small export apparatus protein, SctS, in the activity of the type III secretion system. Front. Microbiol. 2019, 10, 2551. [Google Scholar] [CrossRef] [PubMed]
- Thomassin, J.-L.; He, X.; Thomas, N.A. Role of EscU auto-cleavage in promoting type III effector translocation into host cells by enteropathogenic Escherichia coli. BMC Microbiol. 2011, 11, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minamino, T.; Morimoto, Y.V.; Hara, N.; Aldridge, P.D.; Namba, K. The bacterial flagellar type III export gate complex is a dual fuel engine that can use both H+ and Na+ for flagellar protein export. PLoS Pathog. 2016, 12, e1005495. [Google Scholar] [CrossRef]
- Majewski, D.D.; Worrall, L.J.; Hong, C.; Atkinson, C.E.; Vuckovic, M.; Watanabe, N.; Yu, Z.; Strynadka, N.C.J. Cryo-EM structure of the homohexameric T3SS ATPase-central stalk complex reveals rotary ATPase-like asymmetry. Nat. Commun. 2019, 10, 626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biemans-Oldehinkel, E.; Sal-Man, N.; Deng, W.; Foster, L.J.; Finlay, B.B. Quantitative proteomic analysis reveals formation of an EscL-EscQ-EscN type III complex in enteropathogenic Escherichia coli. J. Bacteriol. 2011, 193, 5514–5519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto, E.; Espinosa, N.; Díaz-Guerrero, M.; Gaytán, M.O.; Puente, J.L.; González-Pedrajo, B. Functional Characterization of EscK (Orf4), a sorting platform component of the enteropathogenic Escherichia coli injectisome. J. Bacteriol. 2016, 61, e00538-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadouloglou, V.E.; Tampakaki, A.P.; Glykos, N.M.; Bastaki, M.N.; Hadden, J.M.; Phillips, S.E.; Panopoulos, N.J.; Kokkinidis, M. Structure of HrcQB-C, a conserved component of the bacterial type III secretion systems. Proc. Natl. Acad. Sci. USA 2003, 101, 70–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseytin, I.; Dagan, A.; Oren, S.; Sal-Man, N. The role of EscD in supporting EscC polymerization in the type III secretion system of enteropathogenic Escherichia coli. Biochim. Biophys. Acta Biomembr. 2018, 1860, 384–395. [Google Scholar] [CrossRef]
- García-Gómez, E.; Espinosa, N.; De La Mora, J.; Dreyfus, G.; Gónzalez-Pedrajo, B. The muramidase EtgA from enteropathogenic Escherichia coli is required for efficient type III secretion. Microbiology 2011, 157, 1145–1160. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Wagner, S. Investigating the assembly of the bacterial type III secretion system injectisome by in vivo photocrosslinking. Int. J. Med. Microbiol. 2019, 309, 151331. [Google Scholar] [CrossRef] [PubMed]
- Diepold, A.; Wagner, S. Assembly of the bacterial type III secretion machinery. FEMS Microbiol. Rev. 2014, 38, 802–822. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Worrall, L.J.; Vuckovic, M.; Hong, C.; Deng, W.; Atkinson, C.E.; Brett Finlay, B.; Yu, Z.; Strynadka, N.C.J. T3S injectisome needle complex structures in four distinct states reveal the basis of membrane coupling and assembly. Nat. Microbiol. 2019, 4, 2010–2019. [Google Scholar] [CrossRef]
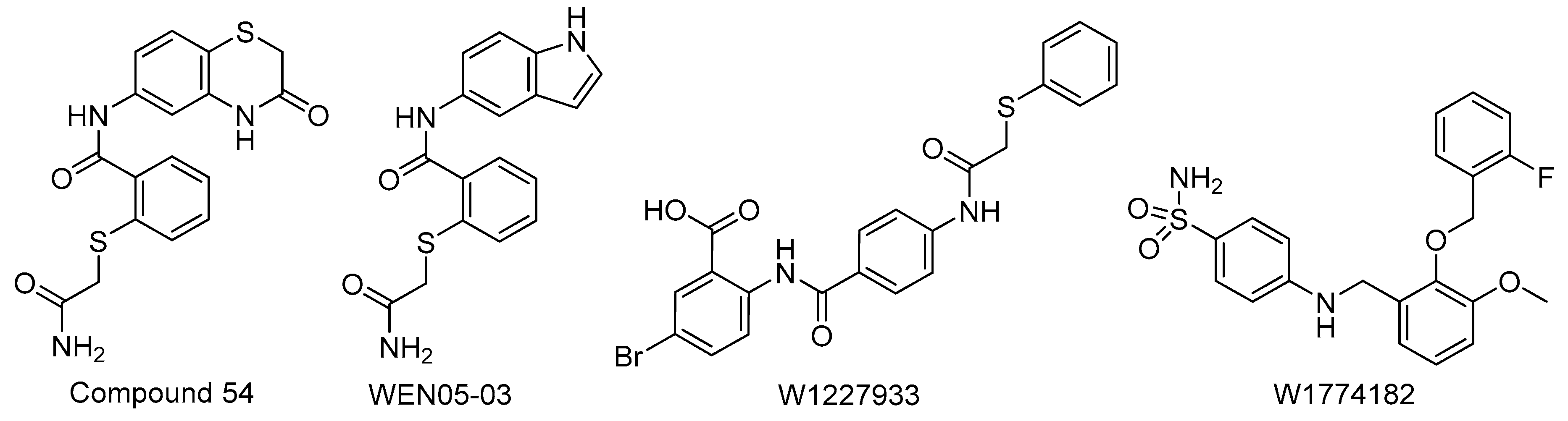
- Felise, H.B.; Nguyen, H.V.; Pfuetzner, R.A.; Barry, K.C.; Jackson, S.R.; Blanc, M.-P.; Bronstein, P.A.; Kline, T.; Miller, S.I. An inhibitor of Gram-negative bacterial virulence protein secretion. Cell Host Microbe 2008, 4, 325–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kline, T.; Barry, K.C.; Jackson, S.R.; Felise, H.B.; Nguyen, H.V.; Miller, S.I. Tethered thiazolidinone dimers as inhibitors of the bacterial type III secretion system. Bioorganic Med. Chem. Lett. 2009, 19, 1340–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Case, H.B.; Dickenson, N.E. Kinetic characterization of the Shigella type three secretion system ATPase Spa47 using α-32P ATP. Bio-Protocol 2018, 8, e3074. [Google Scholar] [CrossRef] [PubMed]
- Akeda, Y.; Galán, J.E. Chaperone release and unfolding of substrates in type III secretion. Nat. Cell Biol. 2005, 437, 911–915. [Google Scholar] [CrossRef]
- Paul, K.; Erhardt, M.; Hirano, T.; Blair, D.F.; Hughes, K.T. Energy source of flagellar type III secretion. Nat. Cell Biol. 2008, 451, 489–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, J.M.; Waldor, M.K. The locus of enterocyte effacement-encoded effector proteins all promote enterohemorrhagic Escherichia coli pathogenicity in infant rabbits. Infect. Immun. 2005, 73, 1466–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swietnicki, W.; Carmany, D.; Retford, M.; Guelta, M.; Dorsey, R.; Bozue, J.; Lee, M.S.; Olson, M.A. Identification of small-molecule inhibitors of Yersinia pestis type III secretion system YscN ATPase. PLoS ONE 2011, 6, e19716. [Google Scholar] [CrossRef]
- Bozue, J.; Cote, C.K.; Webster, W.; Bassett, A.; Tobery, S.; Little, S.; Swietnicki, W. A Yersinia pestis YscN ATPase mutant functions as a live attenuated vaccine against bubonic plague in mice. FEMS Microbiol. Lett. 2012, 332, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Quinaud, M.; Plé, S.; Job, V.; Contreras-Martel, C.; Simorre, J.-P.; Attree, I.; Dessen, A. Structure of the heterotrimeric complex that regulates type III secretion needle formation. Proc. Natl. Acad. Sci. USA 2007, 104, 7803–7808. [Google Scholar] [CrossRef] [Green Version]
- Gong, L.; Lai, S.-C.; Treerat, P.; Prescott, M.; Adler, B.; Boyce, J.D.; Devenish, R.J. Burkholderia pseudomallei type III secretion system cluster 3 ATPase BsaS, a chemotherapeutic target for small-molecule ATPase inhibitors. Infect. Immun. 2015, 83, 1276–1285. [Google Scholar] [CrossRef] [Green Version]
- Grishin, A.V.; Luyksaar, S.I.; Kapotina, L.N.; Kirsanov, D.D.; Zayakin, E.S.; Karyagina, A.S.; Zigangirova, N.A. Identification of chlamydial T3SS inhibitors through virtual screening against T3SS ATPase. Chem. Biol. Drug Des. 2018, 91, 717–727. [Google Scholar] [CrossRef]
- Stone, C.B.; Bulir, D.C.; Emdin, C.A.; Pirie, R.M.; Porfilio, E.A.; Slootstra, J.W.; Mahony, J.B. Chlamydia pneumoniae CdsL regulates CdsN ATPase activity, and disruption with a peptide mimetic prevents bacterial invasion. Front. Microbiol. 2011, 2, 21. [Google Scholar] [CrossRef] [Green Version]
- Bzdzion, L.; Krezel, H.; Wrzeszcz, K.; Grzegorek, I.; Nowinska, K.; Chodaczek, G.; Swietnicki, W. Design of small molecule inhibitors of type III secretion system ATPase EscN from enteropathogenic Escherichia coli. Acta Biochim. Pol. 2017, 64, 49–63. [Google Scholar] [CrossRef]
- Anantharajah, A.; Buyck, J.M.; Sundin, C.; Tulkens, P.M.; Mingeot-Leclercq, M.-P.; Van Bambeke, F. Salicylidene acylhydrazides and hydroxyquinolines act as inhibitors of type three secretion systems in Pseudomonas aeruginosa by distinct mechanisms. Antimicrob. Agents Chemother. 2017, 61, e02566-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerboneschi, M.; Decorosi, F.; Biancalani, C.; Ortenzi, M.V.; Macconi, S.; Giovannetti, L.; Viti, C.; Campanella, B.; Onor, M.; Bramanti, E.; et al. Indole-3-acetic acid in plant–pathogen interactions: A key molecule for in planta bacterial virulence and fitness. Res. Microbiol. 2016, 167, 774–787. [Google Scholar] [CrossRef] [Green Version]
- Enquist, P.A.; Gylfe, Å.; Hägglund, U.; Lindström, P.; Norberg-Scherman, H.; Sundin, C.; Elofsson, M. Derivatives of 8-hydroxyquinoline - Antibacterial agents that target intra- and extracellular Gram-negative pathogens. Bioorganic Med. Chem. Lett. 2012, 22, 3550–3553. [Google Scholar] [CrossRef]
- Case, H.B.; Mattock, D.S.; Dickenson, N.E. Shutting down Shigella secretion: Characterizing small molecule type three secretion system ATPase inhibitors. Biochemistry 2018, 57, 6906–6916. [Google Scholar] [CrossRef]
- Tachiyama, S.; Chang, Y.; Muthuramalingam, M.; Hu, B.; Barta, M.L.; Picking, W.L.; Liu, J.; Picking, W.D. The cytoplasmic domain of MxiG interacts with MxiK and directs assembly of the sorting platform in the Shigella type III secretion system. J. Biol. Chem. 2019, 294, 19184–19196. [Google Scholar] [CrossRef]
- Swietnicki, W.; Czarny, A.; Antkowiak, L.; Zaczynska, E.; Kolodziejczak, M.; Sycz, J.; Stachowicz, L.; Alicka, M.; Marycz, K. Identification of a potent inhibitor of type II secretion system from Pseudomonas aeruginosa. Biochem. Biophys. Res. Commun. 2019, 513, 688–693. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Engel, J.; Balachandran, P. Role of Pseudomonas aeruginosa type III effectors in disease. Curr. Opin. Microbiol. 2009, 12, 61–66. [Google Scholar] [CrossRef]
- Shaver, C.M.; Hauser, A.R. Relative contributions of Pseudomonas aeruginosa ExoU, ExoS, and ExoT. Society 2004, 72, 6969–6977. [Google Scholar]
- Foulkes, D.M.; McLean, K.; Haneef, A.S.; Fernig, D.G.; Winstanley, C.; Berry, N.; Kaye, S.B. Pseudomonas aeruginosa toxin ExoU as a therapeutic target in the treatment of bacterial infections. Microorganisms 2019, 7, 707. [Google Scholar] [CrossRef] [Green Version]
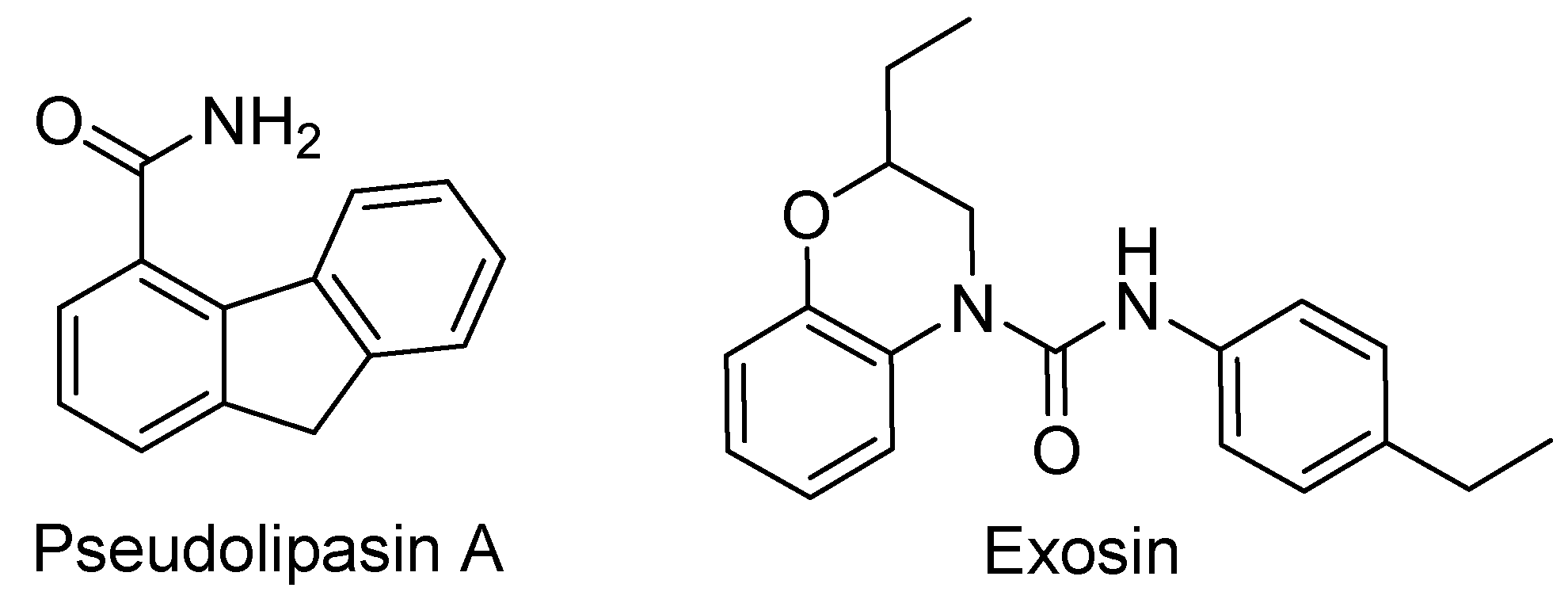
- Lee, V.T.; Pukatzki, S.; Sato, H.; Kikawada, E.; Kazimirova, A.A.; Huang, J.; Li, X.; Arm, J.P.; Frank, D.W.; Lory, S. Pseudolipasin A is a specific inhibitor for phospholipase A2 activity of Pseudomonas aeruginosa cytotoxin ExoU. Infect. Immun. 2007, 75, 1089–1098. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Baek, J.; Song, J.; Byeon, H.; Min, H.; Min, K.H. Identification of arylsulfonamides as ExoU inhibitors. Bioorganic Med. Chem. Lett. 2014, 24, 3823–3825. [Google Scholar] [CrossRef]
- Vareechon, C.; Zmina, S.E.; Karmakar, M.; Pearlman, E.; Rietsch, A. Pseudomonas aeruginosa effector ExoS inhibits ROS production in human neutrophils. Cell Host Microbe 2017, 21, 611.e5–618.e5. [Google Scholar] [CrossRef] [Green Version]
- Arnoldo, A.; Curak, J.; Kittanakom, S.; Chevelev, I.; Lee, V.T.; Sahebol-Amri, M.; Koscik, B.; Ljuma, L.; Roy, P.J.; Bedalov, A.; et al. Identification of small molecule inhibitors of Pseudomonas aeruginosa exoenzyme S using a yeast phenotypic screen. PLoS Genet. 2008, 4, e1000005. [Google Scholar] [CrossRef]
- Khanifar, J.; Hosseini, R.H.; Kazemi, R.; Ramandi, M.F.; Amani, J.; Salmanian, A.H. Prevention of EHEC infection by chitosan nano-structure coupled with synthetic recombinant antigen. J. Microbiol. Methods 2019, 157, 100–107. [Google Scholar] [CrossRef]
- Si, X.; Quan, X.; Wu, Y. A small-molecule norspermidine and norspermidine-hosting polyelectrolyte coatings inhibit biofilm formation by multi-species wastewater culture. Appl. Microbiol. Biotechnol. 2015, 99, 10861–10870. [Google Scholar] [CrossRef]
- Alnaqdy, A.; Al-Jabri, A.; Al Mahrooqi, Z.; Nzeako, B.; Nsanze, H. Inhibition effect of honey on the adherence of Salmonella to intestinal epithelial cells in vitro. Int. J. Food Microbiol. 2005, 103, 347–351. [Google Scholar] [CrossRef]
- Ur-Rehman, T.; Slepenkin, A.; Chu, H.; Blomgren, A.; Dahlgren, M.K.; Zetterström, C.E.; Peterson, E.M.; Elofsson, M.; Gylfe, Å. Pre-clinical pharmacokinetics and anti-chlamydial activity of salicylidene acylhydrazide inhibitors of bacterial type III secretion. J. Antibiot. 2012, 65, 397–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahlgren, M.K.; Kauppi, A.M.; Olsson, I.-M.; Linusson, A.; Elofsson, M. Design, synthesis, and multivariate quantitative structure−activity relationship of salicylanilides potent inhibitors of type III secretion in Yersinia. J. Med. Chem. 2007, 50, 6177–6188. [Google Scholar] [CrossRef] [PubMed]
- Sugita-Konishi, Y.; Sakanaka, S.; Sasaki, K.; Juneja, L.R.; Noda, T.; Amano, F. Inhibition of bacterial adhesion and Salmonella infection in BALB/c mice by sialyloligosaccharides and their derivatives from chicken egg yolk. J. Agric. Food Chem. 2002, 50, 3607–3613. [Google Scholar] [CrossRef]
- Weichert, S.; Jennewein, S.; Hüfner, E.; Weiss, C.; Borkowski, J.; Putze, J.; Schroten, H. Bioengineered 2′-fucosyllactose and 3-fucosyllactose inhibit the adhesion of Pseudomonas aeruginosa and enteric pathogens to human intestinal and respiratory cell lines. Nutr. Res. 2013, 33, 831–838. [Google Scholar] [CrossRef]
- Deng, W.; Li, Y.; Vallance, B.A.; Finlay, B.B. Locus of enterocyte effacement from Citrobacter rodentium: Sequence analysis and evidence for horizontal transfer among attaching and effacing pathogens. Infect. Immun. 2001, 69, 6323–6335. [Google Scholar] [CrossRef] [Green Version]
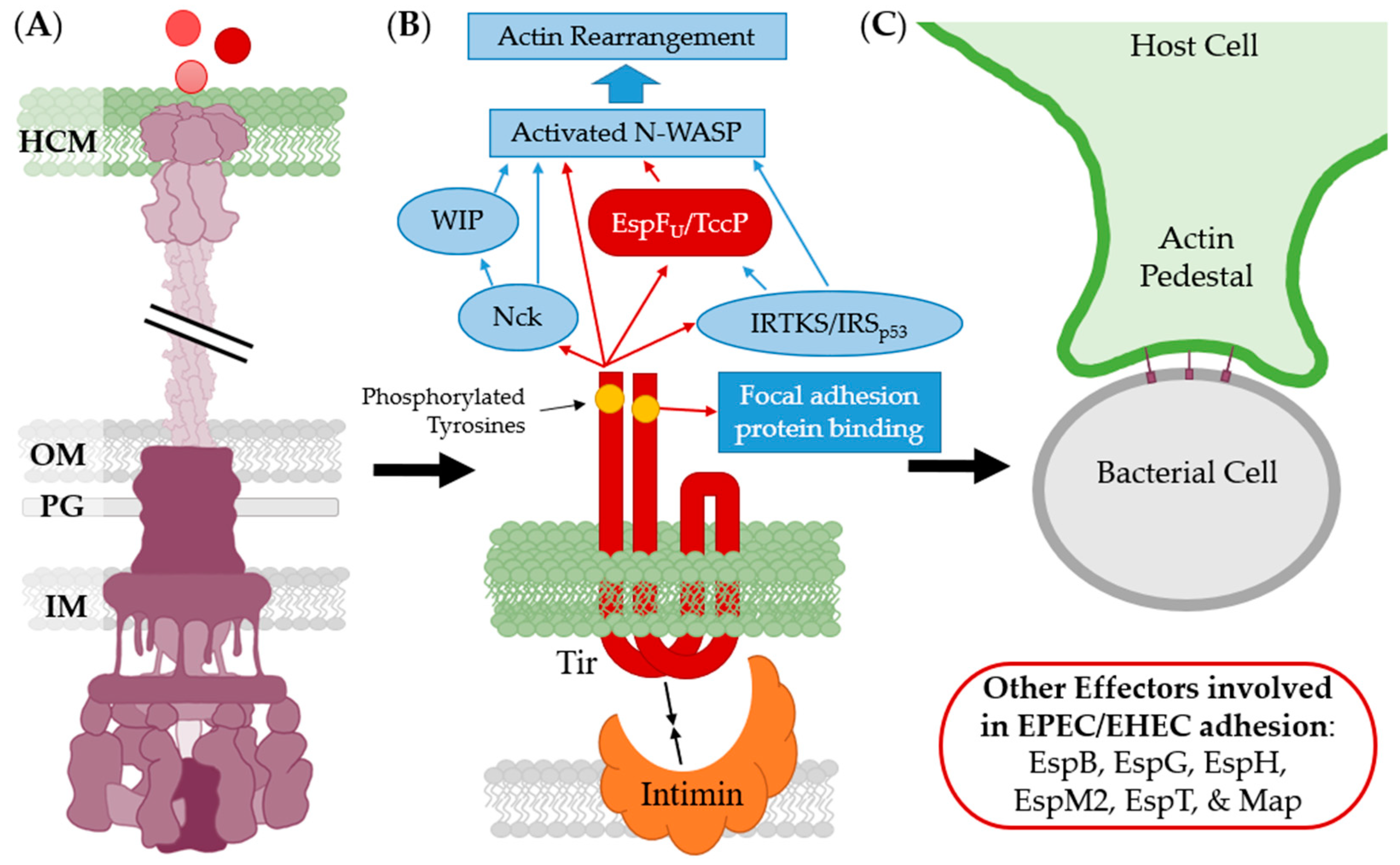
- Lai, Y.; Rosenshine, I.; Leong, J.M.; Frankel, G. Intimate host attachment: Enteropathogenic and enterohaemorrhagic Escherichia coli. Cell. Microbiol. 2013, 15, 1796–1808. [Google Scholar] [CrossRef] [Green Version]
- DeVinney, R.; Stein, M.; Reinscheid, D.; Abe, A.; Ruschkowski, S.; Finlay, B.B. Enterohemorrhagic Escherichia coli O157:H7 Produces Tir, which is translocated to the host cell membrane but is not tyrosine phosphorylated. Infect. Immun. 1999, 67, 2389–2398. [Google Scholar] [CrossRef] [Green Version]
- Kenny, B. Phosphorylation of tyrosine 474 of the enteropathogenic Escherichia coli (EPEC) Tir receptor molecule is essential for actin nucleating activity and is preceded by additional host modifications. Mol. Microbiol. 1999, 31, 1229–1241. [Google Scholar] [CrossRef]
- Donnenberg, M.S.; Tzipori, S.; Mckee, M.L.; O ’brien, A.D.; Alroy, J.; Kapert, J.B. The role of the eae gene of enterohemorrhagic Escherichia coli in intimate attachment in vitro and in a porcine model. Clin. Investig. 1993, 92, 1418–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goosney, D.L.; DeVinney, R.; Pfuetzner, R.A.; Frey, E.A.; Strynadka, N.C.; Finlay, B. Enteropathogenic E. coli translocated intimin receptor, Tir, interacts directly with α-actinin. Curr. Biol. 2000, 10, 735–738. [Google Scholar] [CrossRef] [Green Version]
- Freeman, N.L.; Zurawski, D.V.; Chowrashi, P.; Ayoob, J.C.; Huang, L.; Mittal, B.; Sanger, J.M.; Sanger, J.W. Interaction of the enteropathogenic Escherichia coli protein, translocated intimin receptor (Tir), with focal adhesion proteins. Cell Motil. Cytoskelet. 2000, 47, 307–318. [Google Scholar] [CrossRef]
- Garmendia, J.; Phillips, A.D.; Carlier, M.-F.; Chong, Y.; Schuller, S.; Marches, O.; Dahan, S.; Oswald, E.; Shaw, R.K.; Knutton, S.; et al. TccP is an enterohaemorrhagic Escherichia coli O157:H7 type III effector protein that couples Tir to the actin-cytoskeleton. Cell. Microbiol. 2004, 6, 1167–1183. [Google Scholar] [CrossRef] [PubMed]
- Gruenheid, S.; DeVinney, R.; Bladt, F.; Goosney, D.L.; Gelkop, S.; Gish, G.D.; Pawson, T.; Finlay, B.B. Enteropathogenic E. coli Tir binds Nck to initiate actin pedestal formation in host cells. Nat. Cell Biol. 2001, 3, 856–859. [Google Scholar] [CrossRef]
- Batchelor, M.; Guignot, J.; Patel, A.; Cummings, N.; Cleary, J.; Knutton, S.; Holden, D.W.; Connerton, I.; Frankel, G. Involvement of the intermediate filament protein cytokeratin-18 in actin pedestal formation during EPEC infection. EMBO Rep. 2004, 5, 104–110. [Google Scholar] [CrossRef] [Green Version]
- Goodin, J.L.; Raab, R.W.; McKown, R.L.; Coffman, G.L.; Powell, B.S.; Enama, J.T.; Ligon, J.A.; Andrews, G.P. Yersinia pestis outer membrane type III secretion protein YscC: Expression, purification, characterization, and induction of specific antiserum. Protein Expr. Purif. 2005, 40, 152–163. [Google Scholar] [CrossRef]
- Lynch, S.V.; Flanagan, J.L.; Sawa, T.; Fang, A.; Baek, M.S.; Rubio-Mills, A.; Ajayi, T.; Yanagihara, K.; Hirakata, Y.; Kohno, S.; et al. Polymorphisms in the Pseudomonas aeruginosa type III secretion protein, PcrV – Implications for anti-PcrV immunotherapy. Microb. Pathog. 2010, 48, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Hilf, M.; Yu, V.L.; Sharp, J.; Zuravleff, J.J.; Korvick, J.A.; Muder, R.R. Antibiotic therapy for Pseudomonas aeruginosa bacteremia: Outcome correlations in a prospective study of 200 patients. Am. J. Med. 1989, 87, 540–546. [Google Scholar] [CrossRef]
- Saberianfar, R.; Chin-Fatt, A.; Scott, A.; Henry, K.A.; Topp, E.; Menassa, R. Plant-produced chimeric VHH-sIgA against enterohemorrhagic E. coli intimin shows cross-serotype inhibition of bacterial adhesion to epithelial cells. Front. Plant Sci. 2019, 10, 270. [Google Scholar] [CrossRef] [Green Version]
- La Ragione, R.M.; Patel, S.; Maddison, B.; Woodward, M.J.; Best, A.; Whitelam, G.C.; Gough, K.C. Recombinant anti-EspA antibodies block Escherichia coli O157:H7-induced attaching and effacing lesions in vitro. Microbes Infect. 2006, 8, 426–433. [Google Scholar] [CrossRef]
- Selma, M.V.; Larrosa, M.; Beltrán, D.; Lucas, R.; Morales, J.C.; Tomás-Barberán, F.; Espín, J.C. Resveratrol and some glucosyl, glucosylacyl, and glucuronide derivatives reduce Escherichia coli O157:H7, Salmonella typhimurium, and Listeria monocytogenes Scott A adhesion to colonic epithelial cell lines. J. Agric. Food Chem. 2012, 60, 7367–7374. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Chen, Z.; Cheng, H.; Zheng, J.-X.; Pan, W.-G.; Yang, W.-Z.; Yu, Z.-J.; Deng, Q.-W. Inhibition of adhesion of enteropathogenic Escherichia coli to HEp-2 cells by binding of a novel peptide to EspB protein. Curr. Microbiol. 2016, 73, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Wang, H.; Guo, Y.-L.; Zen, K. Heparin blocks the adhesion of E. coli O157:H7 to human colonic epithelial cells. Biochem. Biophys. Res. Commun. 2008, 369, 1061–1064. [Google Scholar] [CrossRef]
- Wang, S.; Wang, J.; Mou, H.; Luo, B.; Jiang, X. Inhibition of adhesion of intestinal pathogens (Escherichia coli, Vibrio cholerae, Campylobacter jejuni, and Salmonella Typhimurium) by common oligosaccharides. Foodborne Pathog. Dis. 2015, 12, 360–365. [Google Scholar] [CrossRef]
- Ruano-Gallego, D.; Yara, D.A.; Di Ianni, L.; Frankel, G.; Schüller, S.; Fernández, L.Á. A nanobody targeting the translocated intimin receptor inhibits the attachment of enterohemorrhagic E. coli to human colonic mucosa. PLoS Pathog. 2019, 15, e1003081. [Google Scholar] [CrossRef]
- Xue, Y.; Du, M.; Zhu, M.J. Quercetin prevents Escherichia coli O157:H7 adhesion to epithelial cells via suppressing focal adhesions. Front. Microbiol. 2019, 10, 3278. [Google Scholar] [CrossRef]
- Lin, R.; Piao, M.; Song, Y. Dietary quercetin increases colonic microbial diversity and attenuates colitis severity in Citrobacter rodentium-infected mice. Front. Microbiol. 2019, 10, 1092. [Google Scholar] [CrossRef]
- Jones, G.W.; Richardson, L.A. The attachment to, and invasion of HeLa cells by Salmonella Typhimurium: The contribution of mannose-sensitive and mannose-resistant haemagglutinating activities. J. Gen. Microbiol. 1981, 127, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golberg, D.; Kroupitski, Y.; Belausov, E.; Pinto, R.; Sela, S. Salmonella Typhimurium internalization is variable in leafy vegetables and fresh herbs. Int. J. Food Microbiol. 2011, 145, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Yang, D.; Yang, J.; Zhang, X.; Zhang, J.; Fu, J.; Zhou, G.; Liu, H.; Lian, Z.; Han, H. Overexpression of Toll-like receptor 4 enhances LPS-induced inflammatory response and inhibits Salmonella Typhimurium growth in ovine macrophages. Eur. J. Cell Biol. 2019, 98, 36–50. [Google Scholar] [CrossRef]
- Zhang, K.; Riba, A.; Nietschke, M.; Torow, N.; Repnik, U.; Pütz, A.; Fulde, M.; Dupont, A.; Hensel, M.; Hornef, M. Minimal SPI1-T3SS effector requirement for Salmonella enterocyte invasion and intracellular proliferation in vivo. PLOS Pathog. 2018, 14, e1006925. [Google Scholar] [CrossRef] [Green Version]
- Ly, K.T.; Casanova, J.E. Mechanisms of Salmonella entry into host cells. Cell. Microbiol. 2007, 9, 2103–2111. [Google Scholar] [CrossRef]
- Schlumberger, M.C.; Müller, A.J.; Ehrbar, K.; Winnen, B.; Duss, I.; Stecher, B.; Hardt, W.D. Real-time imaging of type III secretion: Salmonella SipA injection into host cells. Proc. Natl. Acad. Sci. USA 2005, 102, 12548–12553. [Google Scholar] [CrossRef] [Green Version]
- Raffatellu, M.; Wilson, R.P.; Chessa, D.; Andrews-polymenis, H.; Tran, Q.T.; Lawhon, S.; Khare, S.; Adams, L.G.; Ba, A.J. SipA, SopA, SopB, SopD, and SopE2 contribute to Salmonella enterica serotype Typhimurium invasion of epithelial cells. Infect. Immun. 2005, 73, 146–154. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.; Mooseker, M.S.; Galán, J.E. Role of the S. Typhimurium actin-binding protein SipA in bacterial internalization. Science 1999, 283, 2092–2095. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, A.; Meikle, L.M.; Ormsby, M.J.; McCormick, B.A.; Christie, J.M.; Brewer, J.M.; Roberts, M.; Wall, D.M. SipA Activation of caspase-3 is a decisive mediator of host cell survival at early stages of Salmonella enterica serovar Typhimurium infection. Infect. Immun. 2017, 85, e00393-17. [Google Scholar] [CrossRef] [Green Version]
- Osiecki, J.C.; Barker, J.; Picking, W.L.; Serfis, A.B.; Berring, E.; Shah, S.; Harrington, A.; Picking, W.D. IpaC from Shigella and SipC from Salmonella possess similar biochemical properties but are functionally distinct. Mol. Microbiol. 2001, 42, 469–481. [Google Scholar] [CrossRef] [PubMed]
- Karavolos, M.H.; Wilson, M.; Henderson, J.; Lee, J.J.; Khan, C.M.A. Type III secretion of the Salmonella effector protein SopE is mediated via an N-terminal amino acid signal and not an mRNA sequence. J. Bacteriol. 2005, 187, 1559–1567. [Google Scholar] [CrossRef] [Green Version]
- Liebl, D.; Qi, X.; Zhe, Y.; Barnett, T.C.; Teasdale, R.D. SopB-mediated recruitment of SNX18 facilitates Salmonella Typhimurium internalization by the host cell. Front. Cell. Infect. Microbiol. 2017, 7, 257. [Google Scholar] [CrossRef] [Green Version]
- Giacomodonato, M.N.; Sarnacki, H.; Llana, N.; Cerquetti, C. SopB effector protein of Salmonella Typhimurium is translocated in mesenteric lymph nodes during murine salmonellosis. FEMS Microbiol. Lett. 2011, 317, 100–106. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, F.; Kawashima, Y.; Takeuchi, S.; Okimori, K.; Inobe, E.; Oosawa, K. SptP106–136plays a role in the complex formation with SptP-specific chaperone SicP. Biosci. Biotechnol. Biochem. 2014, 78, 1560–1563. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R.; Byrne, A.; Berger, C.N.; Klemm, E.; Crepin, V.F.; Dougan, G.; Frankel, G. The type III secretion system effector SptP of Salmonella enterica serovar Typhi. J. Bacteriol. 2017, 199, e00647-16. [Google Scholar] [CrossRef] [Green Version]
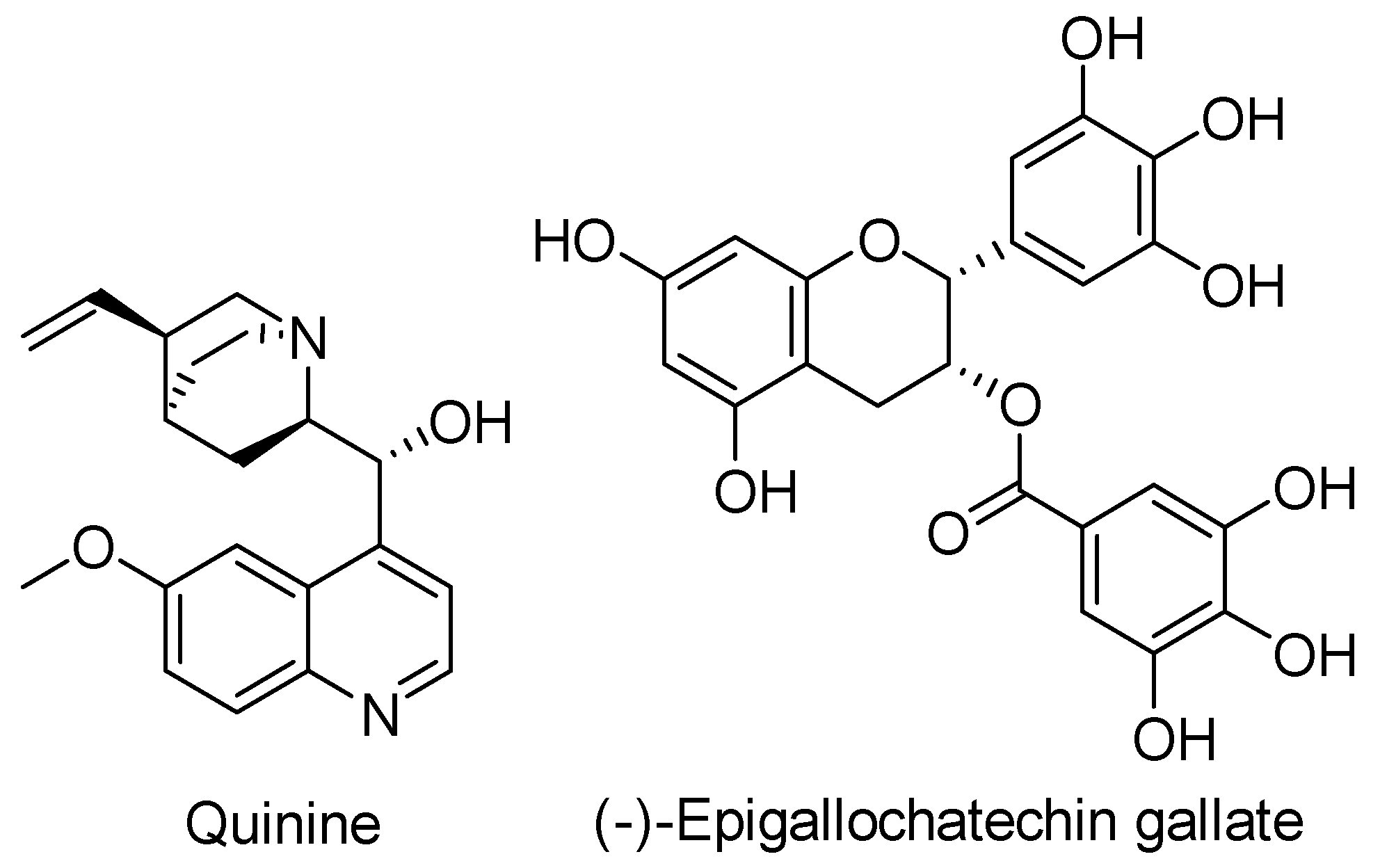
- Wolf, R.; Grimaldi, E.; Donnarumma, G.; Greco, R.; Auricchio, L.; De Filippis, A.; Tufano, M.A. Quinine sulfate inhibits invasion of Salmonella typhimurium and Shigella flexneri: A preliminary study. J. Travel Med. 2006, 12, 343–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kharal, S.A.; Hussain, Q.; Ali, S. Fakhuruddin Quinine is bactericidal. J. Pak. Med. Assoc. 2009, 59, 208–211. [Google Scholar] [PubMed]
- Nakasone, N.; Higa, N.; Toma, C.; Ogura, Y.; Suzuki, T.; Yamashiro, T. Epigallocatechin gallate inhibits the type III secretion system of Gram-negative enteropathogenic bacteria under model conditions. FEMS Microbiol. Lett. 2017, 364, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Tsou, L.K.; Yount, J.S.; Hang, H.C. Epigallocatechin-3-gallate inhibits bacterial virulence and invasion of host cells. Bioorganic Med. Chem. 2017, 25, 2883–2887. [Google Scholar] [CrossRef] [PubMed]
- Runte, C.S.; Jain, U.; Getz, L.J.; Secord, S.; Kuwae, A.; Abe, A.; LeBlanc, J.J.; Stadnyk, A.W.; Kaper, J.B.; Hansen, A.M.; et al. Tandem tyrosine phosphosites in the enteropathogenic Escherichia coli chaperone CesT are required for differential type III effector translocation and virulence. Mol. Microbiol. 2018, 108, 536–550. [Google Scholar] [CrossRef] [Green Version]
- Castiblanco, L.F.; Triplett, L.R.; Sundin, G.W. Regulation of effector delivery by type III secretion chaperone proteins in Erwinia amylovora. Front. Microbiol. 2018, 9, 146. [Google Scholar] [CrossRef]
- Little, D.J.; Coombes, B.K. Molecular basis for CesT recognition of type III secretion effectors in enteropathogenic Escherichia coli. PLOS Pathog. 2018, 14, e1007224. [Google Scholar] [CrossRef] [PubMed]
- Elbaz, N.; Socol, Y.; Katsowich, N.; Rosenshine, I. Control of type III secretion system effector/chaperone ratio fosters pathogen adaptation to host-adherent lifestyle. mBio 2019, 10, e02074-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramu, T.; Prasad, M.E.; Connors, E.; Mishra, A.; Thomassin, J.L.; Leblanc, J.; Rainey, J.K.; Thomas, N.A. A novel C-terminal region within the multicargo type III secretion chaperone CesT contributes to effector secretion. J. Bacteriol. 1999, 195, 740–756. [Google Scholar] [CrossRef] [Green Version]
- Elliott, S.J.; Hutcheson, S.W.; Dubois, M.S.; Mellies, J.L.; Wainwright, L.A.; Batchelor, M.; Frankel, G.; Knutton, S.; Kaper, J.B. Identification of CesT, a chaperone for the type III secretion of Tir in enteropathogenic Escherichia coli. Mol. Microbiol. 1999, 33, 1176–1189. [Google Scholar] [CrossRef]
- Creasey, E.A.; Delahay, R.M.; Bishop, A.A.; Shaw, R.K.; Kenny, B.; Knutton, S.; Frankel, G. CesT is a bivalent enteropathogenic Escherichia coli chaperone required for translocation of both Tir and Map. Mol. Microbiol. 2003, 47, 209–221. [Google Scholar] [CrossRef]
- Gauthier, A.; Finlay, B.B. Translocated intimin receptor and its chaperone interact with ATPase of the type III secretion apparatus of enteropathogenic Escherichia coli. J. Bacteriol. 2003, 185, 6747–6755. [Google Scholar] [CrossRef] [Green Version]
- Trülzsch, K.; Roggenkamp, A.; Aepfelbacher, M.; Wilharm, G.; Ruckdeschel, K.; Heesemann, J. Analysis of chaperone-dependent Yop secretion/translocation and effector function using a mini-virulence plasmid of Yersinia enterocolitica. Int. J. Med. Microbiol. 2003, 293, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Kerschen, E.J.; Cohen, D.A.; Kaplan, A.M.; Straley, S.C. The plague virulence protein YopM targets the innate immune response by causing a global depletion of NK cells. Infect. Immun. 2004, 72, 4589–4602. [Google Scholar] [CrossRef] [Green Version]
- Evdokimov, A.G.; Anderson, D.E.; Routzahn, K.M.; Waugh, D.S. Unusual molecular architecture of the Yersinia pestis cytotoxin YopM: A leucine-rich repeat protein with the shortest repeating unit. J. Mol. Biol. 2001, 312, 807–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
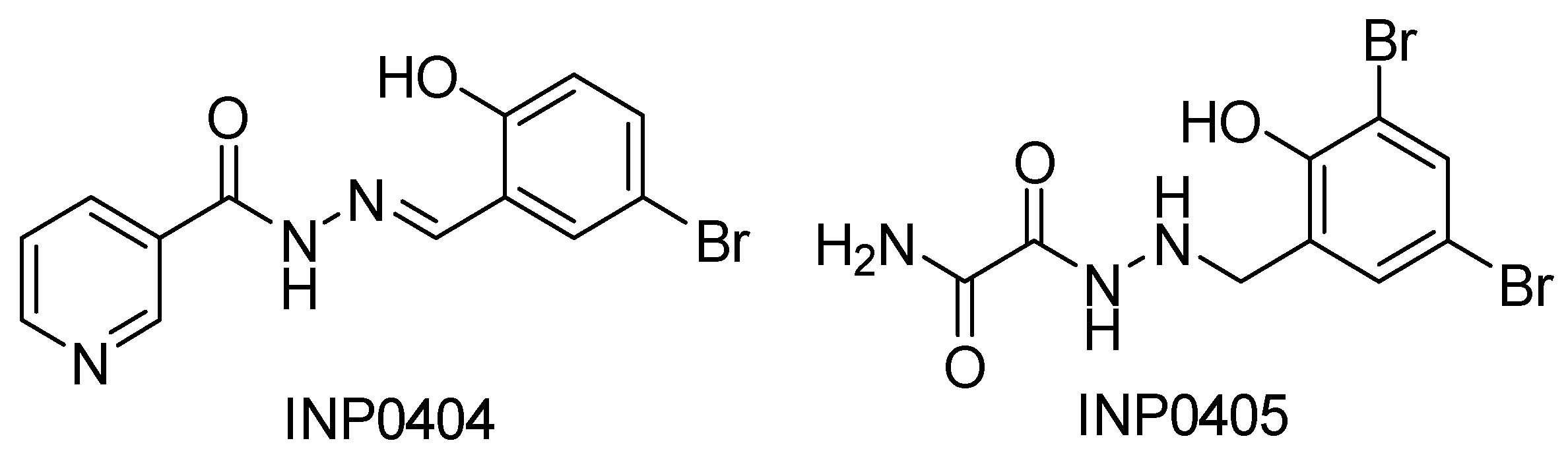
- Sharma, P.; Elofsson, M.; Roy, S. Attenuation of Pseudomonas aeruginosa infection by INP0341, a salicylidene acylhydrazide, in a murine model of keratitis. Virulence 2020, 11, 795–804. [Google Scholar] [CrossRef] [PubMed]























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence |
|---|---|
| EspA | KANNLTTTVNNSQLEIQQMSNTLNLLTSARSDMQSLQYRTISGI |
| Coil A | LTTTVNNSQLEIQQM |
| Coil B | MSNTLNLLTSARSDM |
| Coil AB1 | KMNNLTTKVNNLQLELQEMRNTLKRLKSAMRRMQ |
| Coil AB2 | KINNLTTKVNNLQLELQEMRNTLKRLKSAMRRMQ |
| EscF | LSDSVPELLNSTDLVNDPEKMLELQFAVQ |
| Coil C | LSDSVPELLNSTDLV |
| Coil D | VNDPEKMLELQFAVQ |
| CesA | IVSQTRNKELLDKKIRSEIEAIKKIIAEFDVVKESVNELSEKAK |
| Ces A1 | IVSQTRNKELLDKKIRSEIEA |
| Ces A2 | IKKIIAEFDVVKESVNELSEK |
| Species (T3SS) | Chaperone | Binding Partner | Class | PDB/REF |
|---|---|---|---|---|
| Aeromonas spp. | AcrH | AopB & AopD | II | 3WXX [134] |
| AscE/AscG | AscF | III | 3PH0 [135] | |
| EPEC/EHEC | CesD | EspB & EspD | II | [136,137] |
| CesD2 | EspD | II | [138] | |
| EscE/EscG | EscF | III | [129] | |
| CesA (CesAB) | EspA | III | 1XOU (2M1N) [130,131] | |
| EscG/CesA2 | EscF & EspA | III | [129,130,139] | |
| Pseudomonas spp. | PcrH | PopB & PopD | II | 2XCC, 2XCB [140] |
| PcrG | PcrV | III | [141] | |
| PscE/PscG | PscF | III | 2UWJ [142] | |
| Salmonella spp. (SPI1) | SicA | SipB & SipC | II | [143] |
| SipD | PrgI | III | 3ZQB [144] | |
| Salmonella spp. (SPI2) | SseA | SseC & SseD | II | [145] |
| SseA | SseB | III | [146] | |
| SsaH/SsaE | SsaG | III | [147] | |
| Shigella spp. | IpgC | IpaB & IpaC | II | 3GYZ [148] |
| Spa13 | MxiH | III | [149] | |
| Yersinia spp. | SycD | YopD & YscO | II | 4AM9 [150] |
| LcrG | LcrV | III | [151] | |
| YscE/YscG | YscF | III | 2P58 [152] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hotinger, J.A.; Pendergrass, H.A.; May, A.E. Molecular Targets and Strategies for Inhibition of the Bacterial Type III Secretion System (T3SS); Inhibitors Directly Binding to T3SS Components. Biomolecules 2021, 11, 316. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11020316
Hotinger JA, Pendergrass HA, May AE. Molecular Targets and Strategies for Inhibition of the Bacterial Type III Secretion System (T3SS); Inhibitors Directly Binding to T3SS Components. Biomolecules. 2021; 11(2):316. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11020316
Chicago/Turabian StyleHotinger, Julia A., Heather A. Pendergrass, and Aaron E. May. 2021. "Molecular Targets and Strategies for Inhibition of the Bacterial Type III Secretion System (T3SS); Inhibitors Directly Binding to T3SS Components" Biomolecules 11, no. 2: 316. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11020316