
Rapamycin Improves Recognition Memory and Normalizes Amino-Acids and Amines Levels in the Hippocampal Dentate Gyrus in Adult Rats Exposed to Ethanol during the Neonatal Period

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
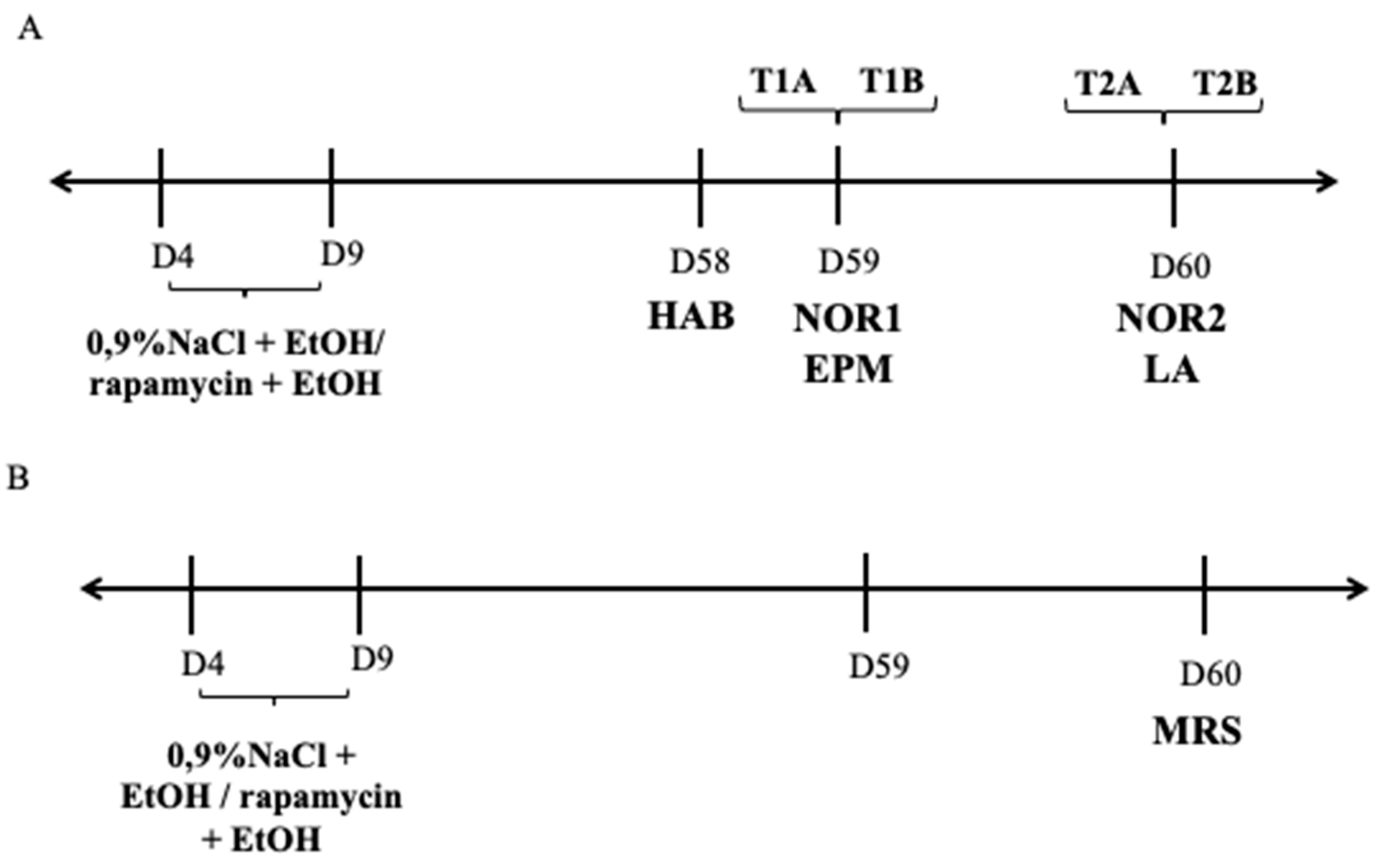
2.2. Drugs and Neonatal Treatment
2.3. Procedures
2.3.1. Novel Object Recognition Test
2.3.2. Elevated Plus Maze (EPM)
2.3.3. Locomotor Activity
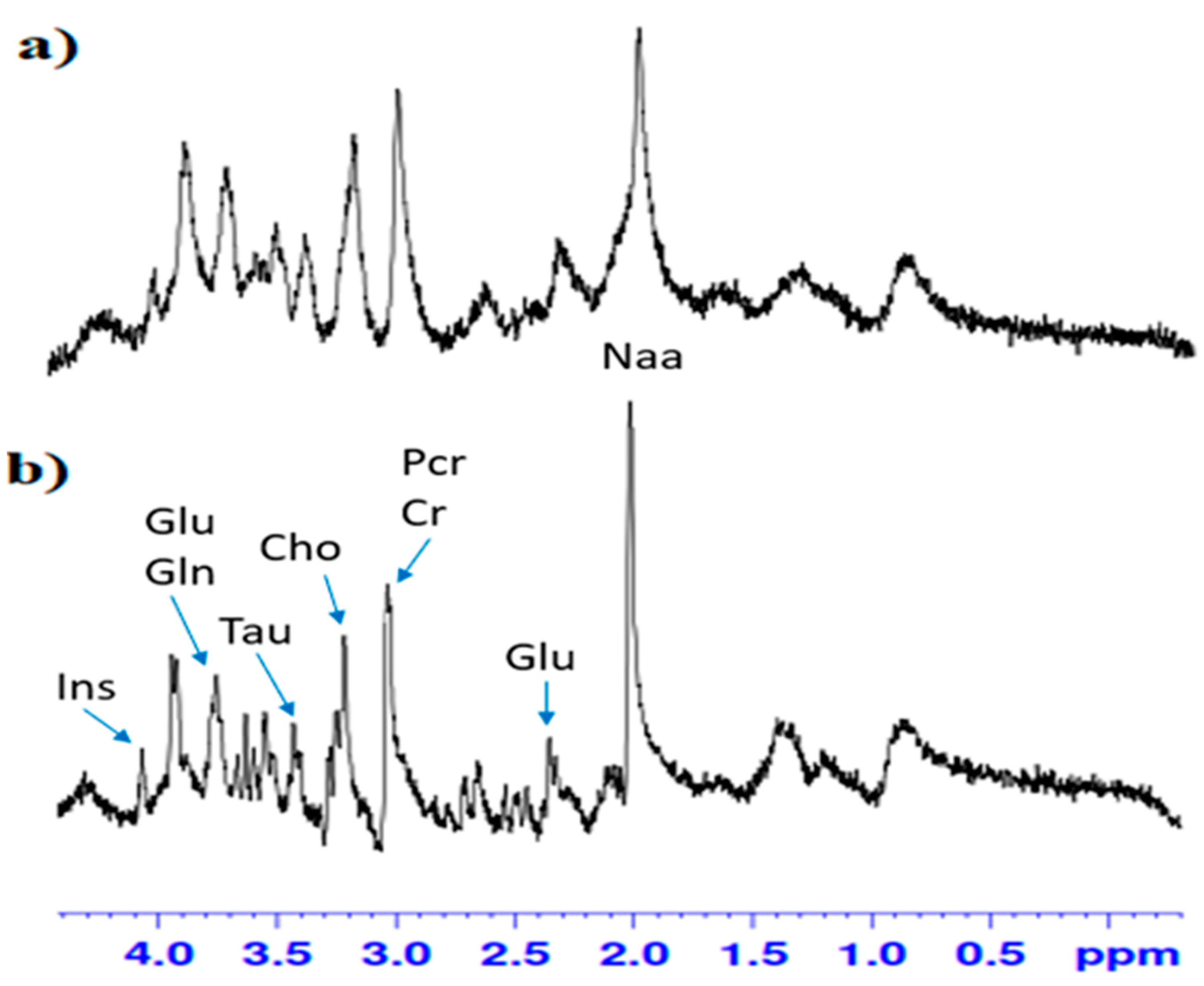
2.4. Spectral Analysis and Quantification of Neurochemicals in the Dentate Gyrus of the Hippocampus In Vivo
2.5. Statistical Analyses
3. Results
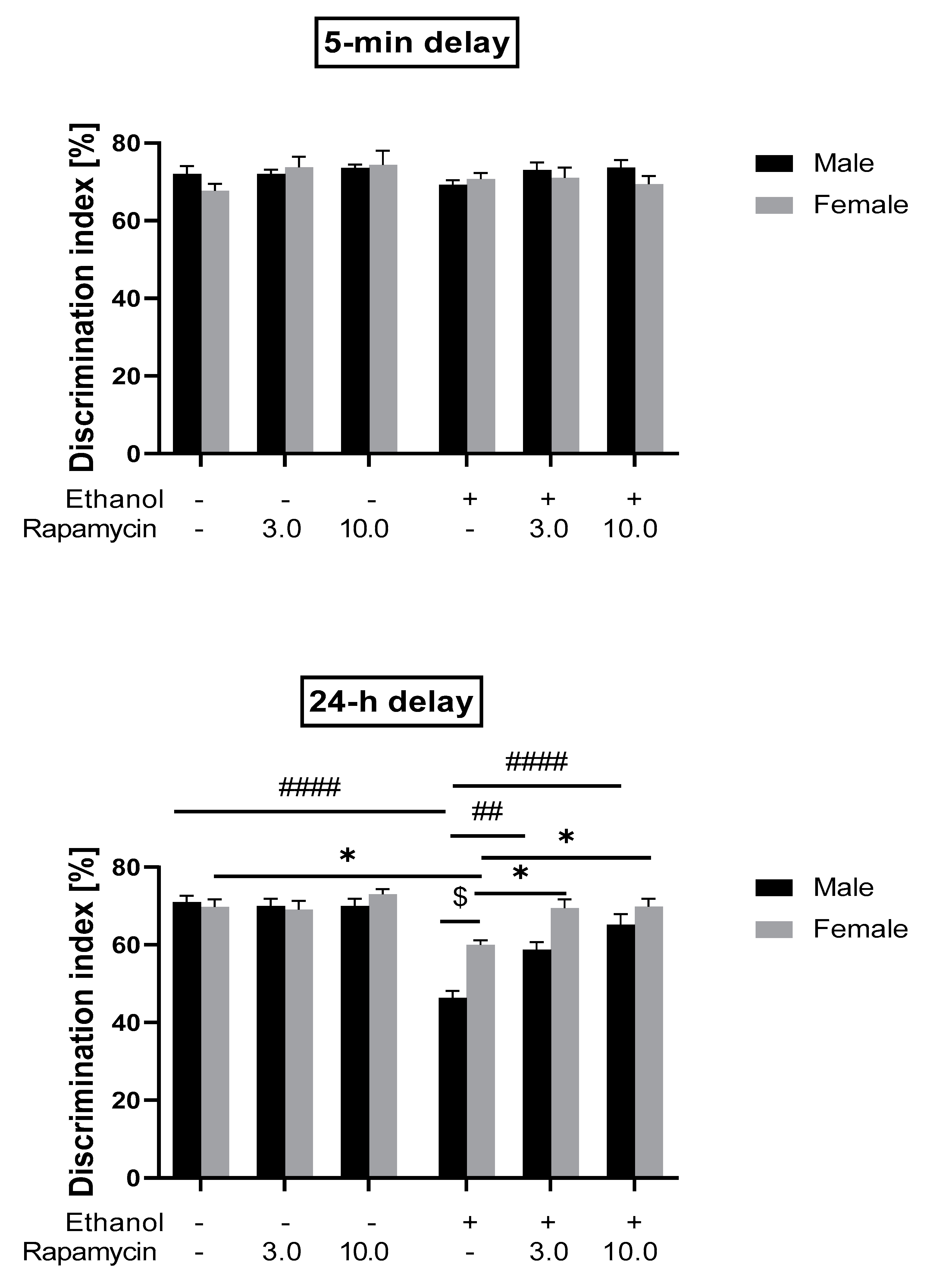
3.1. The Influence of Rapamycin Pre-Treatment before Every Ethanol Administration during PND 4-9 on the Short-and Long-Term Memory in the NOR Test in Adult (PND 59/60) Male and Female Rats
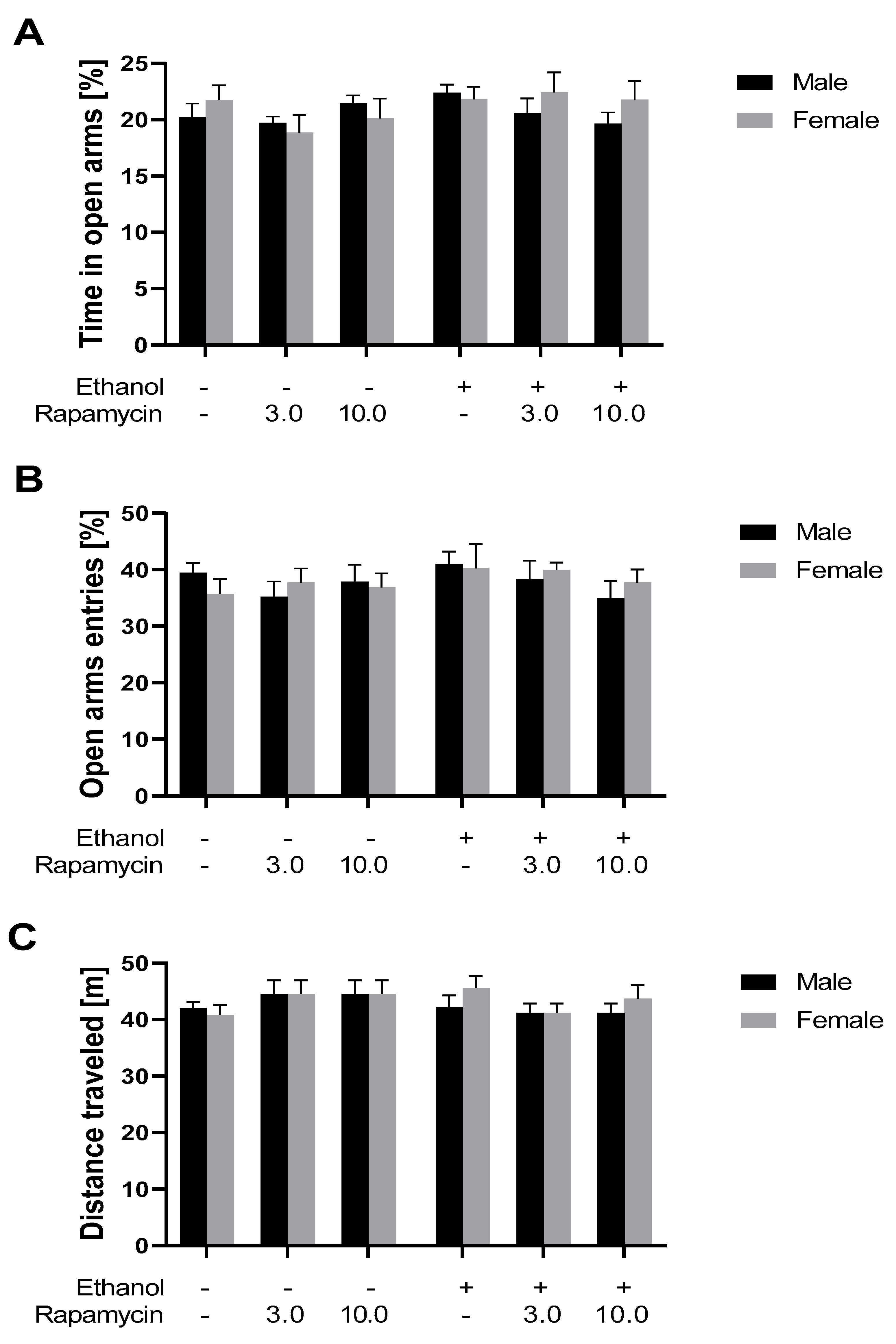
3.2. The Influence of Rapamycin Pre-Treatment before Every Ethanol Administration during PND4-9 on Anxiety-Like Behavior in Adult Male and Female (PND 59) Rats Using the EPM Test
3.3. The Influence of Rapamycin Pre-Treatment before Every Ethanol Administration during PND 4-9 on Locomotor Activity in adult (PND 60) Male and Female Rats
3.4. The Influence of Rapamycin Pre-Treatment before Every Ethanol Administration during PND 4-9 on Changes in Glutamate, Glutamine Concentration and Glutamate/Glutamine Ratio in the Hippocampal DG Measured by MRS Experiment in Adult (PND 60) Male Rats
3.4.1. Glutamate
3.4.2. Glutamine
3.5. The Influence of Rapamycin Pre-Treatment before Every Ethanol Administration during PND 4-9 on Gamma-Aminobutyric Acid, Taurine and Total Choline Concentration in the Hippocampal DG Measured by MRS Experiment In Adult (PND 60) Male Rats
3.5.1. Taurine
3.5.2. Total Choline
3.6. The Influence of Rapamycin Pre-Treatment before Every Ethanol Administration during PND 4-9 on N-acetylaspartate, Inosytol and Creatine Concentration in the Hippocampal DG Measured by MRS Experiment in Adult (PND 60), Male Rats
3.6.1. N-Acetylaspartate
3.6.2. Inosytol
3.6.3. Total Creatine
4. Discussion
4.1. Rapamycin Prevents Ethanol-Induced Recognition Memory Impairment
4.2. Rapamycin Prevents Neonatal Ethanol Induced Developmental Alteration in the DG. MRS Experiments
4.3. Potential Mechanism of Rapamycin
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hayes, D.M.; Deeny, M.A.; Shaner, C.A.; Nixon, K. Determining the threshold for alcohol-induced brain damage: New evidence with gliosis markers. Alcohol. Clin. Exp. Res. 2013, 37, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Riley, E.P.; Infante, M.A.; Warren, K.R. Fetal alcohol spectrum disorders: An overview. Neuropsychol. Rev. 2011, 21, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Mattson, S.N.; Crocker, N.; Nguyen, T.T. Fetal alcohol spectrum disorders: Neuropsychological and behavioral features. Neuropsychol. Rev. 2011, 21, 81–101. [Google Scholar] [CrossRef] [Green Version]
- Riley, E.P.; McGee, C.L. Fetal alcohol spectrum disorders: An overview with emphasis on changes in brain and behavior. Exp. Biol. Med. 2005, 230, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Norman, A.L.; Crocker, N.; Mattson, S.N.; Riley, E.P. Neuroimaging and fetal alcohol spectrum disorders. Dev. Disabil. Res. Rev. 2009, 15, 209–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil-Mohapel, J.; Boehme, F.; Kainer, L.; Christie, B.R. Hippocampal cell loss and neurogenesis after fetal alcohol exposure: Insights from different rodent models. Brain Res. Rev. 2010, 64, 283–303. [Google Scholar] [CrossRef] [PubMed]
- Wilhoit, L.F.; Scott, D.A.; Simecka, B.A. Fetal Alcohol Spectrum Disorders: Characteristics, Complications, and Treatment. Community Ment. Health J. 2017, 53, 711–718. [Google Scholar] [CrossRef]
- Maier, S.E.; West, J.R. Drinking patterns and alcohol-related birth defects. Alcohol. Res. Health 2001, 25, 168–174. [Google Scholar] [PubMed]
- Brown, L.M.; Leslie, S.; Gonzales, R.A. The effects of chronic ethanol exposure on N-methyl-D-aspartate-stimulated overflow of [3H] catecholamines from rat brain. Brain Res. 1991, 547, 289–294. [Google Scholar] [CrossRef]
- Korkman, M.; Kettunen, S.; Autti-Rämö, I. Neurocognitive impairment in early adolescence following prenatal alcohol exposure of varying duration. Child Neuropsychol. 2003, 9, 117–128. [Google Scholar] [CrossRef]
- Kodituwakku, P.W.; Handmaker, N.S.; Cutler, S.K.; Weathersby, E.K.; Handmaker, S.D. Specific impairments in self-regulation in children exposed to alcohol prenatally. Alcohol. Clin. Exp. Res. 1995, 19, 1558–1564. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Imaki, T.; Vale, W.; Rivier, C. Effect of prenatal exposure to ethanol on the activity of the hypothalamic-pituitary-adrenal axis of the offspring: Importance of the time of exposure to ethanol and possible modulating mechanisms. Mol. Cell. Neurosci. 1990, 1, 168–177. [Google Scholar] [CrossRef]
- Witter, M.P.; Van Hoesen, G.W.; Amaral, D.G. Topographical organization of the entorhinal projection to the dentate gyrus of the monkey. J. Neurosci. 1989, 9, 216–228. [Google Scholar] [CrossRef] [Green Version]
- Witter, M.P.; Amaral, D.G. Entorhinal cortex of the monkey: V. Projections to the dentate gyrus, hippocampus, and subicular complex. J. Comp. Neurol. 1991, 307, 437–459. [Google Scholar] [CrossRef]
- Jessberger, S.; Clark, R.E.; Broadbent, N.J.; Clemenson, G.D., Jr.; Consiglio, A.; Lie, D.C.; Squire, L.R.; Gage, F.H. Dentate gyrus-specific knockdown of adult neurogenesis impairs spatial and object recognition memory in adult rats. Learn Mem. 2009, 16, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, H.L.; Lu, Y.L.; Botterill, J.J.; Scharfman, H.E. Novelty and novel objects increase c-Fos immunoreactivity in mossy cells in the mouse dentate gyrus. Neural Plast. 2019, 27, 1815371. [Google Scholar] [CrossRef]
- Dillon, S.E.; Tsivos, D.; Knight, M.; McCann, B.; Pennington, C.; Shiel, A.I.; Conway, M.E.; Newson, M.A.; Kauppinen, R.A.; Coulthard, E.J. The impact of ageing reveals distinct roles for human dentate gyrus and CA3 in pattern separation and object recognition memory. Sci. Rep. 2017, 7, 14069. [Google Scholar] [CrossRef] [Green Version]
- Marr, D. Simple memory: A theory for archicortex. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1971, 262, 23–81. [Google Scholar] [CrossRef] [PubMed]
- Ennaceur, A.; Delacour, J. A new one-trial test for neurobiological studies of memory in rats. 1: Behavioral data. Behav. Brain Res. 1988, 31, 47–59. [Google Scholar] [CrossRef]
- Stough, S.; Shobe, J.L.; Carew, T.J. Intermediate-term processes in memory formation. Curr. Opin. Neurobiol. 2006, 16, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Taglialatela, G.; Hogan, D.; Zhang, W.R.; Dineley, K.T. Intermediate- and long-term recognition memory deficits in Tg2576 mice are reversed with acute calcineurin inhibition. Behav. Brain. Res. 2009, 200, 95–99. [Google Scholar] [CrossRef] [Green Version]
- Alberini, C.M.; Kandel, E.R. The regulation of transcription in memory consolidation. Cold Spring Harb. Perspect. Biol. 2015, 7, a021741. [Google Scholar] [CrossRef] [Green Version]
- Amtul, Z.; Rahman, A.U. Neural Plasticity and Memory: Is Memory Encoded in Hydrogen Bonding Patterns? Neuroscientist. 2016, 22, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Kapahi, P.; Zid, B. TOR pathway: Linking nutrient sensing to life span. Sci. Aging Knowl. Environ. 2004, 2004, 34. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.E.; Hall, M.N. The expanding TOR signaling network. Curr. Opin. Cell Biol. 2005, 17, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.F.; Hunter, R.W.; Hers, I. mTORC2 protein complex-mediated Akt (Protein Kinase B) Serine 473 Phosphorylation is not required for Akt1 activity in human platelets. J. Biol. Chem. 2011, 286, 24553–24560. [Google Scholar] [CrossRef] [Green Version]
- Jaworski, J.; Sheng, M. The growing role of mTOR in neuronal development and plasticity. Mol. Neurobiol. 2006, 34, 205–219. [Google Scholar] [CrossRef]
- Costa-Mattioli, M.; Sossin, W.S.; Klann, E.; Sonenberg, N. Translational control of long-lasting synaptic plasticity and memory. Neuron 2009, 61, 10–26. [Google Scholar] [CrossRef] [Green Version]
- Mameli, M.; Balland, B.; Luján, R.; Lüscher, C. Rapid synthesis and synaptic insertion of GluR2 for mGluR-LTD in the ventral tegmental area. Science 2007, 317, 530–533. [Google Scholar] [CrossRef] [Green Version]
- Blundell, J.; Kouser, M.; Powell, C.M. Systemic inhibition of mammalian target of rapamycin inhibits fear memory reconsolidation. Neurobiol. Learn Mem. 2008, 90, 28–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puighermanal, E.; Marsicano, G.; Busquets-Garcia, A.; Lutz, B.; Maldonado, R.; Ozaita, A. Cannabinoid modulation of hippocampal long-term memory is mediated by mTOR signaling. Nat. Neurosci. 2009, 12, 1152–1158. [Google Scholar] [CrossRef]
- Tang, S.J.; Reis, G.; Kang, H.; Gingras, A.C.; Sonenberg, N.; Schuman, E.M. A rapamycin-sensitive signaling pathway contributes to long-term synaptic plasticity in the hippocampus. Proc. Natl. Acad. Sci. USA 2002, 99, 467–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoeffer, C.A.; Klann, E. mTOR signaling: At the crossroads of plasticity, memory and disease. Trends Neurosci. 2010, 33, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Neasta, J.; Ben Hamida, S.; Yowell, Q.; Carnicella, S.; Ron, D. Role for mammalian target of rapamycin complex 1 signaling in neuroadaptations underlying alcohol-related disorders. Proc. Natl. Acad. Sci. USA 2010, 107, 20093–20098. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Lunde-Young, E.R.; Naik, V.; Ramirez, J.; Orzabal, M.; Ramadoss, J. Chronic Binge Alcohol Exposure During Pregnancy Alters mTOR System in Rat Fetal Hippocampus. Alcohol. Clin. Exp. Res. 2020, 44, 1329–1336. [Google Scholar] [CrossRef]
- Davis-Anderson, K.L.; Wesseling, H.; Siebert, L.M.; Lunde-Young, E.R.; Naik, V.D.; Steen, H.; Ramadoss, J. Fetal regional brain protein signature in FASD rat model. Reprod. Toxicol. 2018, 76, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Semple, B.D.; Blomgren, K.; Gimlin, K.; Ferriero, D.M.; Noble-Haeusslein, L.J. Brain development in rodents and humans: Identifying benchmarks of maturation and vulnerability to injury across species. Prog. Neurobiol. 2013, 106, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Chabenne, A.; Moon, C.; Ojo, C.; Khogali, A.; Nepal, B.; Sharma, S. Biomarkers in fetal alcohol syndrome. Biomark. Genom. Med. 2014, 6, 12–22. [Google Scholar] [CrossRef] [Green Version]
- Goodfellow, M.J.; Abdulla, K.A.; Lindquist, D.H. Neonatal Ethanol Exposure Impairs Trace Fear Conditioning and Alters NMDA Receptor Subunit Expression in Adult Male and Female Rats. Alcohol. Clin. Exp. Res. 2016, 40, 309–318. [Google Scholar] [CrossRef]
- MacIlvane, N.M.; Pochiro, J.M.; Hurwitz, N.R.; Goodfellow, M.J.; Lindquist, D.H. Recognition memory is selectively impaired in adult rats exposed to binge-like doses of ethanol during early postnatal life. Alcohol 2016, 57, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Dursun, I.; Jakubowska-Doğru, E.; Elibol-Can, B.; van der List, D.; Chapman, B.; Qi, L.; Berman, R.F. Effects of early postnatal alcohol exposure on the developing retinogeniculate projections in C57BL/6 mice. Alcohol 2013, 47, 173–179. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Liu, L.; Wen, Q.; Zheng, C.; Gao, Y.; Peng, S.; Tan, Y.; Li, Y. Rapamycin prevents drug seeking via disrupting reconsolidation of reward memory in rats. Int. J. Neuropsychopharmacol. 2014, 17, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Marszalek-Grabska, M.; Gibula-Bruzda, E.; Bodzon-Kulakowska, A.; Suder, P.; Gawel, K.; Filarowska, J.; Listos, J.; Danysz, W.; Kotlinska, J.H. Effects of the Positive Allosteric Modulator of Metabotropic Glutamate Receptor 5, VU-29, on Impairment of Novel Object Recognition Induced by Acute Ethanol and Ethanol Withdrawal in Rats. Neurotox. Res. 2018, 33, 607–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marszalek-Grabska, M.; Gibula-Bruzda, E.; Jenda, M.; Gawel, K.; Kotlinska, J.H. Memantine improves memory impairment and depressive-like behavior induced by amphetamine withdrawal in rats. Brain Res. 2016, 1642, 389–396. [Google Scholar] [CrossRef]
- Lueptow, L.M. Novel Object Recognition Test for the Investigation of Learning and Memory in Mice. J. Vis. Exp. 2017, 126, 55718. [Google Scholar] [CrossRef] [PubMed]
- Hammond, R.S.; Tull, L.E.; Stackman, R.W. On the delay-dependent involvement of the hippocampus in object recognition memory. Neurobiol. Learn Mem. 2004, 82, 26–34. [Google Scholar] [CrossRef]
- Koltunowska, D.; Gibula-Bruzda, E.; Kotlinska, J.H. The influence of ionotropic and metabotropic glutamate receptor ligands on anxiety-like effect of amphetamine withdrawal in rats. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 45, 242–249. [Google Scholar] [CrossRef]
- Gomez, J.L.; Lewis, M.J.; Sebastian, V.; Serrano, P.; Luine, V.N. Alcohol administration blocks stress-induced impairments in memory and anxiety and alters hippocampal neurotransmitter receptor expression in male rats. Horm. Behav. 2013, 63, 659–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, H.C.; Feldman, J.J.; Streissguth, A.P.; Sampson, P.D.; Bookstein, F.L. Neuropsychological deficits in adolescents with fetal alcohol syndrome: Clinical findings. Alcohol. Clin. Exp. Res. 1998, 22, 1998–2012. [Google Scholar] [CrossRef]
- Burden, M.J.; Westerlund, A.; Muckle, G.; Dodge, N.; Dewailly, E.; Nelson, C.A.; Jacobson, S.W.; Jacobson, J.L. The effects of maternal binge drinking during pregnancy on neural correlates of response inhibition and memory in childhood. Alcohol. Clin. Exp. Res. 2011, 35, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Röskam, S.; Koch, M. Effects of neonatal and peripubertal ethanol treatment on various aspects of adult rat behavior and brain anatomy. Int. J. Dev. Neurosci. 2009, 27, 249–256. [Google Scholar] [CrossRef]
- Kim, C.K.; Kalynchuk, L.E.; Kornecook, T.J.; Mumby, D.G.; Dadgar, N.A.; Pinel, J.P.; Weinberg, J. Object-recognition and spatial learning and memory in rats prenatally exposed to ethanol. Behav. Neurosci. 1997, 111, 985–995. [Google Scholar] [CrossRef]
- Summers, B.L.; Henry, C.M.; Rofe, A.M.; Coyle, P. Dietary zinc supplementation during pregnancy prevents spatial and object recognition memory impairments caused by early prenatal ethanol exposure. Behav. Brain Res. 2008, 186, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.A.; Peterson, J.; Basavaraj, B.S.; Saito, M. Local and regional network function in behaviorally relevant cortical circuits of adult mice following postnatal alcohol exposure. Alcohol. Clin. Exp. Res. 2011, 35, 1974–1984. [Google Scholar] [CrossRef] [Green Version]
- Terasaki, L.S.; Gomez, J.; Schwarz, J.M. An examination of sex differences in the effects of early-life opiate and alcohol exposure. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150123. [Google Scholar] [CrossRef] [Green Version]
- Kelly, S.J.; Leggett, D.C.; Cronise, K. Sexually dimorphic effects of alcohol exposure during development on the processing of social cues. Alcohol Alcohol. 2009, 44, 555–560. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Hawkey, A.B.; Li, H.; Dai, L.; Brim, H.H.; Frank, J.A.; Luo, J.; Barron, S.; Chen, G. Neonatal Ethanol Exposure Causes Behavioral Deficits in Young Mice. Alcohol. Clin. Exp. Res. 2018, 42, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.J.; Munchow, A.H.; Rios, L.M.; Zhang, G.; Asgeirsdóttir, H.N.; Stackman, R.W., Jr. The rodent hippocampus is essential for nonspatial object memory. Curr. Biol. 2013, 23, 1685–1690. [Google Scholar] [CrossRef] [Green Version]
- Bayer, S.A.; Altman, J.; Russo, R.J.; Zhang, X. Timetables of neurogenesis in the human brain based on experimentally determined patterns in the rat. Neurotoxicology 1993, 14, 83–144. [Google Scholar] [PubMed]
- Goodlett, C.R.; Johnson, T.B. Neonatal binge ethanol exposure using intubation: Timing and dose effects on place learning. Neurotoxicol. Teratol. 1997, 19, 435–446. [Google Scholar] [CrossRef]
- Bonthius, D.J.; Pantazis, N.J.; Karacay, B.; Bonthius, N.E.; Taggard, D.A.; Lothman, E.W. Alcohol exposure during the brain growth spurt promotes hippocampal seizures, rapid kindling, and spreading depression. Alcohol. Clin. Exp. Res. 2001, 25, 734–745. [Google Scholar] [CrossRef]
- Hamilton, G.F.; Whitcher, L.T.; Klintsova, A.Y. Postnatal binge-like alcohol exposure decreases dendritic complexity while increasing the density of mature spines in mPFC Layer II/III pyramidal neurons. Synapse 2010, 64, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Klintsova, A.Y.; Helfer, J.L.; Calizo, L.H.; Dong, W.K.; Goodlett, C.R.; Greenough, W.T. Persistent impairment of hippocampal neurogenesis in young adult rats following early postnatal alcohol exposure. Alcohol. Clin. Exp. Res. 2007, 31, 2073–2082. [Google Scholar] [CrossRef] [PubMed]
- Puglia, M.P.; Valenzuela, C.F. Repeated third trimester-equivalent ethanol exposure inhibits long-term potentiation in the hippocampal CA1 region of neonatal rats. Alcohol 2010, 44, 283–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayer, S.A. Development of the hippocampal region in the rat I. Neurogenesis examined with 3H-thymidine autoradiography. J. Comp. Neurol. 1980, 190, 87–114. [Google Scholar] [CrossRef]
- Bayer, S.A. Changes in the total number of dentate granule cells in juvenile and adult rats: A correlated volumetric and 3 H-thymidine autoradiographic study. Exp. Brain Res. 1982, 46, 315–323. [Google Scholar] [CrossRef]
- Beason-Held, L.L.; Rosene, D.L.; Killiany, R.J.; Moss, M.B. Hippocampal formation lesions produce memory impairment in the rhesus monkey. Hippocampus 1999, 9, 562–574. [Google Scholar] [CrossRef]
- Cave, C.B.; Squire, L.R. Equivalent impairment of spatial and nonspatial memory following damage to the human hippocampus. Hippocampus 1991, 1, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.M.; Squire, L.R. Impaired recognition memory in patients with lesions limited to the hippocampal formation. Behav. Neurosci. 1997, 111, 667–675. [Google Scholar] [CrossRef]
- Zola, S.M.; Squire, L.R.; Teng, E.; Stefanacci, L.; Buffalo, E.A.; Clark, R.E. Impaired recognition memory in monkeys after damage limited to the hippocampal region. J. Neurosci. 2000, 20, 451–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, R.E.; Zola, S.M.; Squire, L.R. Impaired recognition memory in rats after damage to the hippocampus. J. Neurosci. 2000, 20, 8853–8860. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, C.J.; Pinar, C.; Yang, W.; Pang, A.F.; Suesser, K.E.; Choi, J.S.J.; Christie, B.R. Impaired Bidirectional Synaptic Plasticity in Juvenile Offspring Following Prenatal Ethanol Exposure. Alcohol. Clin. Exp. Res. 2019, 43, 2153–2166. [Google Scholar] [CrossRef]
- Pierrefiche, O. Long Term Depression in Rat Hippocampus and the Effect of Ethanol during Fetal Life. Brain Sci. 2017, 7, 157. [Google Scholar] [CrossRef] [PubMed]
- Kervern, M.; Silvestre de Ferron, B.; Alaux-Cantin, S.; Fedorenko, O.; Antol, J.; Naassila, M.; Pierrefiche, O. Aberrant NMDA-dependent LTD after perinatal ethanol exposure in young adult rat hippocampus. Hippocampus 2015, 25, 912–923. [Google Scholar] [CrossRef]
- Dowling, R.J.; Topisirovic, I.; Fonseca, B.D.; Sonenberg, N. Dissecting the role of mTOR: Lessons from mTOR inhibitors. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2010, 1804, 433–439. [Google Scholar] [CrossRef]
- Tapiero, H.; Mathé, G.; Couvreur, P.; Tew, K.D., II. Glutamine and glutamate. Biomed. Pharmacother. 2002, 56, 446–457. [Google Scholar] [CrossRef]
- Waagepetersen, H.S.; Sonnewald, U.; Schousboe, A. Glutamine, Glutamate, and GABA: Metabolic aspects. In Handbook of Neurochemistry and Molecular Neurobiology: Amino Acids and Peptides in the Nervous System; Springer: New York, NY, USA, 2007; pp. 1–21. [Google Scholar]
- Alasmari, F.; Goodwani, S.; McCullumsmith, R.E.; Sari, Y. Role of glutamatergic system and mesocorticolimbic circuits in alcohol dependence. Prog. Neurobiol. 2018, 171, 32–49. [Google Scholar] [CrossRef] [PubMed]
- Ruppel, R.A.; Kochanek, P.M.; Adelson, P.D.; Rose, M.E.; Wisniewski, S.R.; Bell, M.J.; Clark, R.S.; Marion, D.W.; Graham, S.H. Excitatory amino acid concentrations in ventricular cerebrospinal fluid after severe traumatic brain injury in infants and children: The role of child abuse. J. Pediatr. 2001, 138, 18–25. [Google Scholar] [CrossRef]
- Silverstein, F.S.; Naik, B.; Simpson, J. Hypoxia-ischemia stimulates hippocampal glutamate efflux in perinatal rat brain: An in vivo microdialysis study. Pediatric Res. 1991, 30, 587–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, D.A. Maternal stress increases fetal brain and neonatal cerebral cortex 5-hydroxytryptamine synthesis in rats: A possible mechanism by which stress influences brain development. Pharmacol. Biochem. Behav. 1990, 35, 943–947. [Google Scholar] [CrossRef]
- Herlenius, E.; Lagercrantz, H. Development of neurotransmitter systems during critical periods. Exp. Neurol. 2004, 190, 8–21. [Google Scholar] [CrossRef]
- Chefer, V.; Meis, J.; Wang, G.; Kuzmin, A.; Bakalkin, G.; Shippenberg, T. Repeated exposure to moderate doses of ethanol augments hippocampal glutamate neurotransmission by increasing release. Addict. Biol. 2011, 16, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Spanagel, R. Are metabotropic glutamate receptors promising targets for the treatment of alcoholism? Biol. Psychiatry 2010, 67, 798–799. [Google Scholar] [CrossRef]
- Ikonomidou, C.; Bosch, F.; Miksa, M.; Bittigau, P.; Vöckler, J.; Dikranian, K.; Tenkova, T.I.; Stefovska, V.; Turski, L.; Olney, J.W. Blockade of NMDA receptors and apoptotic neurodegeneration in the developing brain. Science 1999, 283, 70–74. [Google Scholar] [CrossRef]
- Wozniak, D.F.; Hartman, R.E.; Boyle, M.P.; Vogt, S.K.; Brooks, A.R.; Tenkova, T.; Young, C.; Olney, J.W.; Muglia, L.J. Apoptotic neurodegeneration induced by ethanol in neonatal mice is associated with profound learning/memory deficits in juveniles followed by progressive functional recovery in adults. Neurobiol. Dis. 2004, 17, 403–414. [Google Scholar] [CrossRef]
- Collingridge, G.L.; Kehl, S.J.; McLennan, H. Excitatory amino acids in synaptic transmission in the Schaffer collateral-commissural pathway of the rat hippocampus. J. Physiol. 1983, 334, 33–46. [Google Scholar] [CrossRef]
- Baker, K.B.; Kim, J.J. Effects of stress and hippocampal NMDA receptor antagonism on recognition memory in rats. Learn Mem. 2002, 9, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.P.; Shimizu, E.; Dube, G.R.; Rampon, C.; Kerchner, G.A.; Zhuo, M.; Liu, G.; Tsien, J.Z. Genetic enhancement of learning and memory in mice. Nature 1999, 401, 63–69. [Google Scholar] [CrossRef]
- Zeisel, S.H. The fetal origins of memory: The role of dietary choline in optimal brain development. J. Pediatr. 2006, 149, 131–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berman, R.F.; Hannigan, J.H. Effects of prenatal alcohol exposure on the hippocampus: Spatial behavior, electrophysiology, and neuroanatomy. Hippocampus 2000, 10, 94–110. [Google Scholar] [CrossRef]
- De Jaeger, X.; Cammarota, M.; Prado, M.A.; Izquierdo, I.; Prado, V.F.; Pereira, G.S. Decreased acetylcholine release delays the consolidation of object recognition memory. Behav. Brain Res. 2013, 238, 62–68. [Google Scholar] [CrossRef]
- Lunde-Young, R.; Davis-Anderson, K.; Naik, V.; Nemec, M.; Wu, G.; Ramadoss, J. Regional dysregulation of taurine and related amino acids in the fetal rat brain following gestational alcohol exposure. Alcohol 2018, 66, 27–33. [Google Scholar] [CrossRef]
- Hamilton, D.A.; Petropoulos, H.; Kodituwakku, P.; Tesche, C.D.; Sutherland, R.J.; Savage, D.D. Functional morphological and spectroscopic measurements of the hippocampus in fetal alcohol syndrome. Alcohol. Clin. Exp. Res. 2003, 27, 217. [Google Scholar]
- O’Leary-Moore, S.K.; McMechan, A.P.; Galloway, M.P.; Hannigan, J.H. Neonatal alcohol-induced region-dependent changes in rat brain neurochemistry measured by high-resolution magnetic resonance spectroscopy. Alcohol. Clin. Exp. Res. 2008, 32, 1697–1707. [Google Scholar] [CrossRef]
- Lee, D.W.; Kim, S.Y.; Kim, J.H.; Lee, T.; Yoo, C.; Nam, Y.K.; Jung, J.Y.; Shin, H.C.; Kim, H.Y.; Kim, D.J.; et al. Quantitative assessment of neurochemical changes in a rat model of long-term alcohol consumption as detected by in vivo and ex vivo proton nuclear magnetic resonance spectroscopy. Neurochem. Int. 2013, 62, 502–509. [Google Scholar] [CrossRef]
- Li, J.; Kim, S.G.; Blenis, J. Rapamycin: One drug, many effects. Cell Metab. 2014, 19, 373–379. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Ke, Z.; Xu, M.; Liao, M.; Wang, X.; Qi, Y.; Zhang, T.; Frank, J.A.; Bower, K.A.; Shi, X.; et al. Autophagy is a protective response to ethanol neurotoxicity. Autophagy 2012, 8, 1577–1589. [Google Scholar] [CrossRef] [Green Version]
- Miller, R.A.; Harrison, D.E.; Astle, C.M.; Fernandez, E.; Flurkey, K.; Han, M.; Javors, M.A.; Li, X.; Nadon, N.L.; Nelson, J.F.; et al. Rapamycin-mediated lifespan increase in mice is dose and sex dependent and metabolically distinct from dietary restriction. Aging Cell 2014, 13, 468–477. [Google Scholar] [CrossRef]
- Rodriguez, K.A.; Dodds, S.G.; Strong, R.; Galvan, V.; Sharp, Z.D.; Buffenstein, R. Divergent tissue and sex effects of rapamycin on the proteasome-chaperone network of old mice. Front. Mol. Neurosci. 2014, 7, 83. [Google Scholar] [CrossRef] [Green Version]
- Earasi, K.; Myc, L.; Obaida, Z.; Zhang, Z.; Song, C.; Macleod, J.; Shim, Y. Effect of gender on the treatment responses to rapamycin on the progression of emphysema in cigarette smoke-exposed mice. Chest 2020, 158, 1813–1814. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | R10 | EtOH | EtOH + R10 | |
|---|---|---|---|---|
| Glu | 7.423833 | 7.531667 | 8.771667 ** | 7.3698333 ## |
| Gln | 2.897333 | 3.029833 | 3.8481667 * | 2.9535 # |
| Tau | 5.569167 | 5.584333 | 5.656333 ### | 6.386167 *** |
| Naa | 3.661 | 3.257 | 3.191333 | 3.3311667 |
| Cho | 1.124667 | 1.189667 | 1.1365 | 1.3233333 |
| Ins | 5.357667 | 5.606667 | 5.573 | 5.2195 |
| Cr | 6.986333 | 6.860833 | 7.175 | 7.033667 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopatynska-Mazurek, M.; Pankowska, A.; Gibula-Tarlowska, E.; Pietura, R.; Kotlinska, J.H. Rapamycin Improves Recognition Memory and Normalizes Amino-Acids and Amines Levels in the Hippocampal Dentate Gyrus in Adult Rats Exposed to Ethanol during the Neonatal Period. Biomolecules 2021, 11, 362. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11030362
Lopatynska-Mazurek M, Pankowska A, Gibula-Tarlowska E, Pietura R, Kotlinska JH. Rapamycin Improves Recognition Memory and Normalizes Amino-Acids and Amines Levels in the Hippocampal Dentate Gyrus in Adult Rats Exposed to Ethanol during the Neonatal Period. Biomolecules. 2021; 11(3):362. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11030362
Chicago/Turabian StyleLopatynska-Mazurek, Malgorzata, Anna Pankowska, Ewa Gibula-Tarlowska, Radoslaw Pietura, and Jolanta H. Kotlinska. 2021. "Rapamycin Improves Recognition Memory and Normalizes Amino-Acids and Amines Levels in the Hippocampal Dentate Gyrus in Adult Rats Exposed to Ethanol during the Neonatal Period" Biomolecules 11, no. 3: 362. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11030362