
Luteolin-Fabricated ZnO Nanostructures Showed PLK-1 Mediated Anti-Breast Cancer Activity

 ,
, 
 , ,
, , 
Abstract
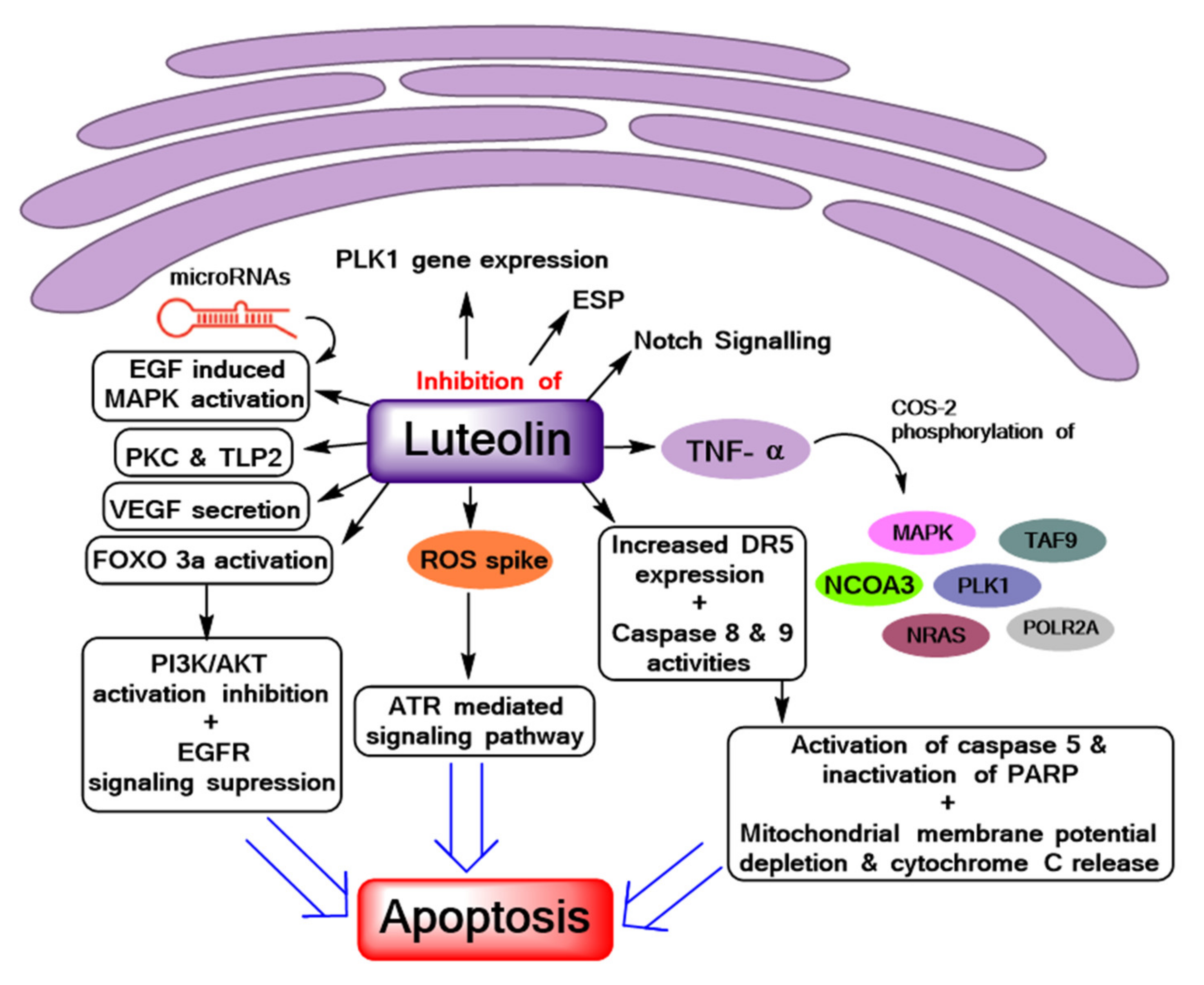
:1. Introduction
2. Materials and Methods
2.1. Plant Material Collection and Extraction of Eclipta alba Phytochemicals
2.2. Isolation of Bioactive Compound
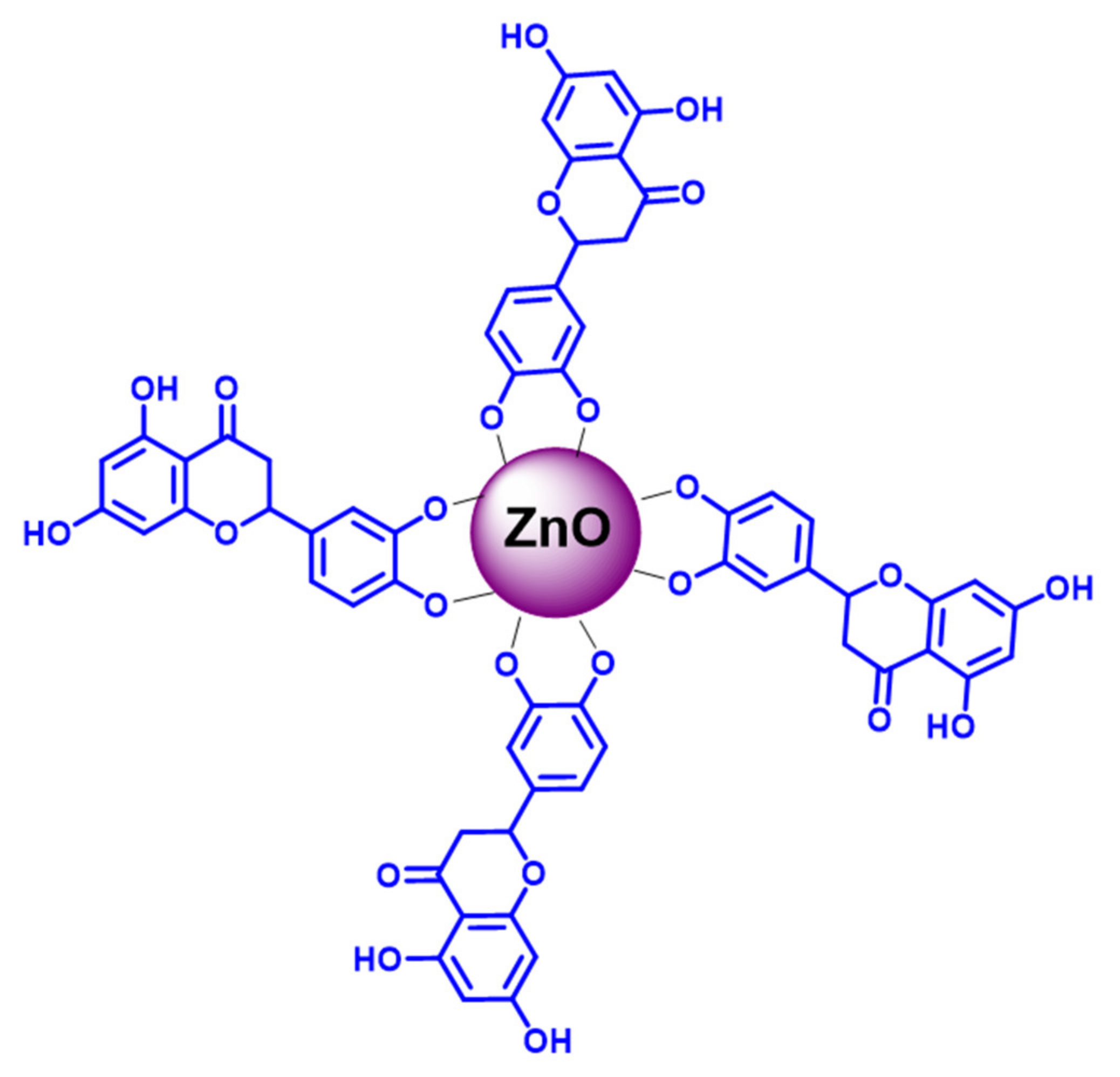
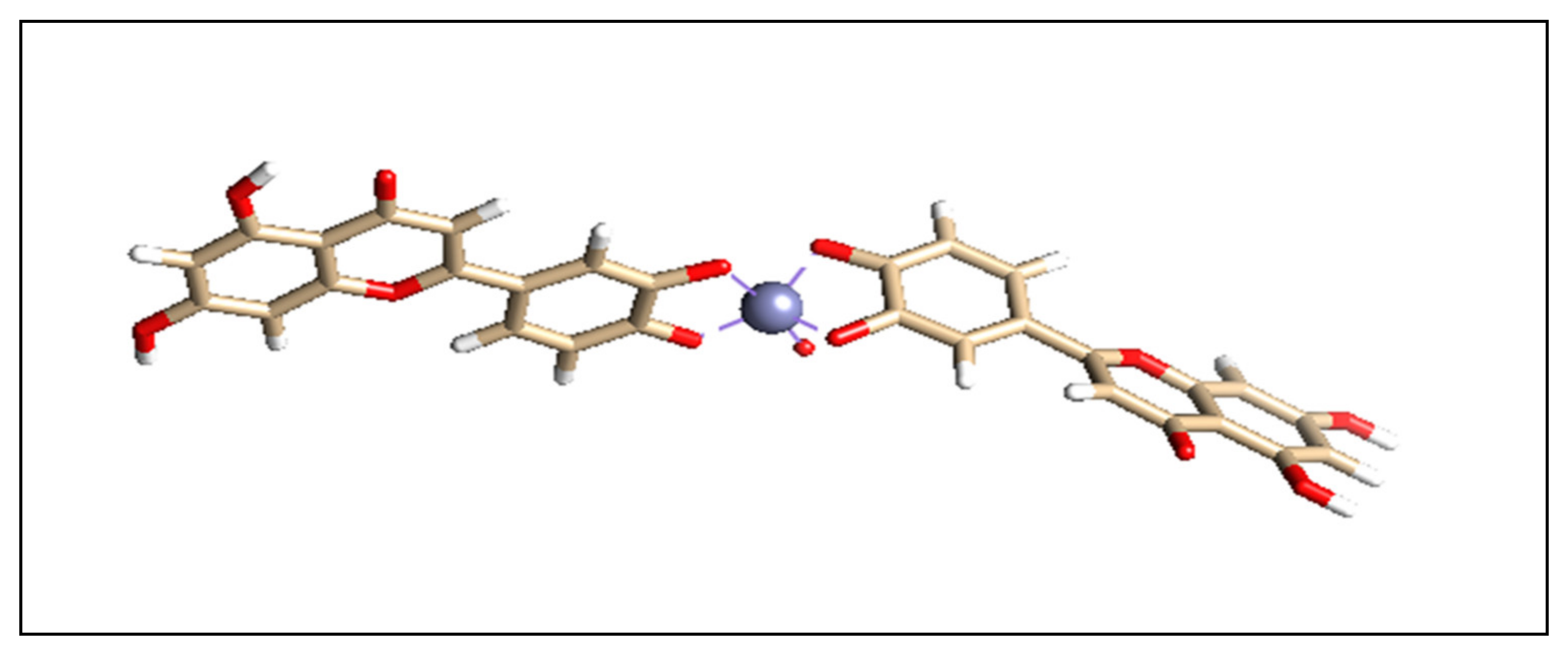
2.3. Preparation of L-ZnONPs
2.4. Determination of Anticancer Activity of As-Synthesized L-ZnONPs
2.5. Measurement of Cell Viability Using MTT Assay
2.6. In Silico Anticancer Study
2.6.1. Ligand Optimization Using Bioinformatics Software
2.6.2. Protein Modeling, Validation, and Preparation Using Bioinformatics Software and Tools
2.6.3. Binding Site Residues
2.6.4. Molecular Interaction Studies
2.6.5. Statistical Analysis
3. Results and Discussion
3.1. XRD Studies
3.2. SEM Analysis
3.3. TEM Analysis
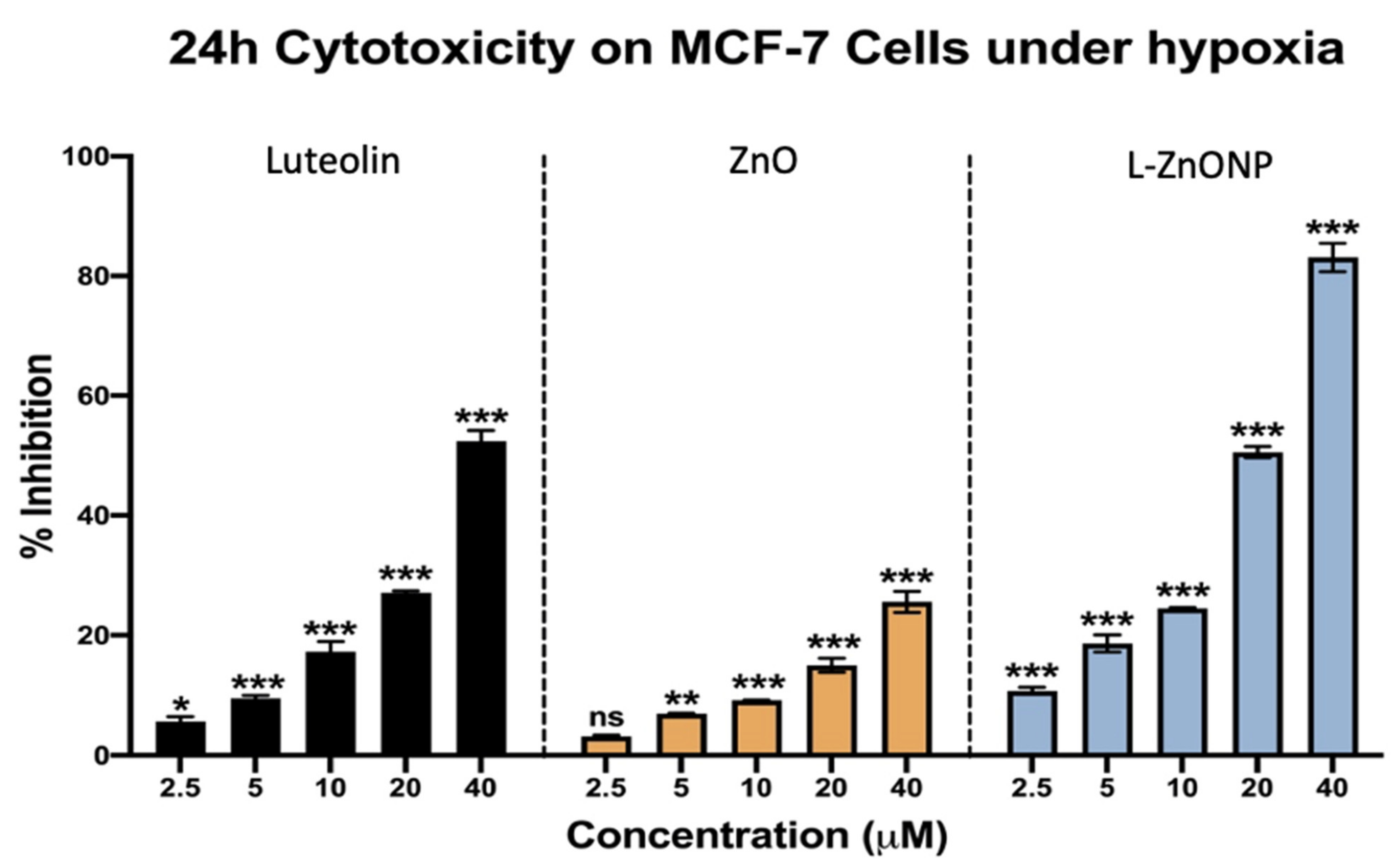
3.4. L-ZnONPs Showed Greater Cytotoxicity in the MCF-7 Cell Line
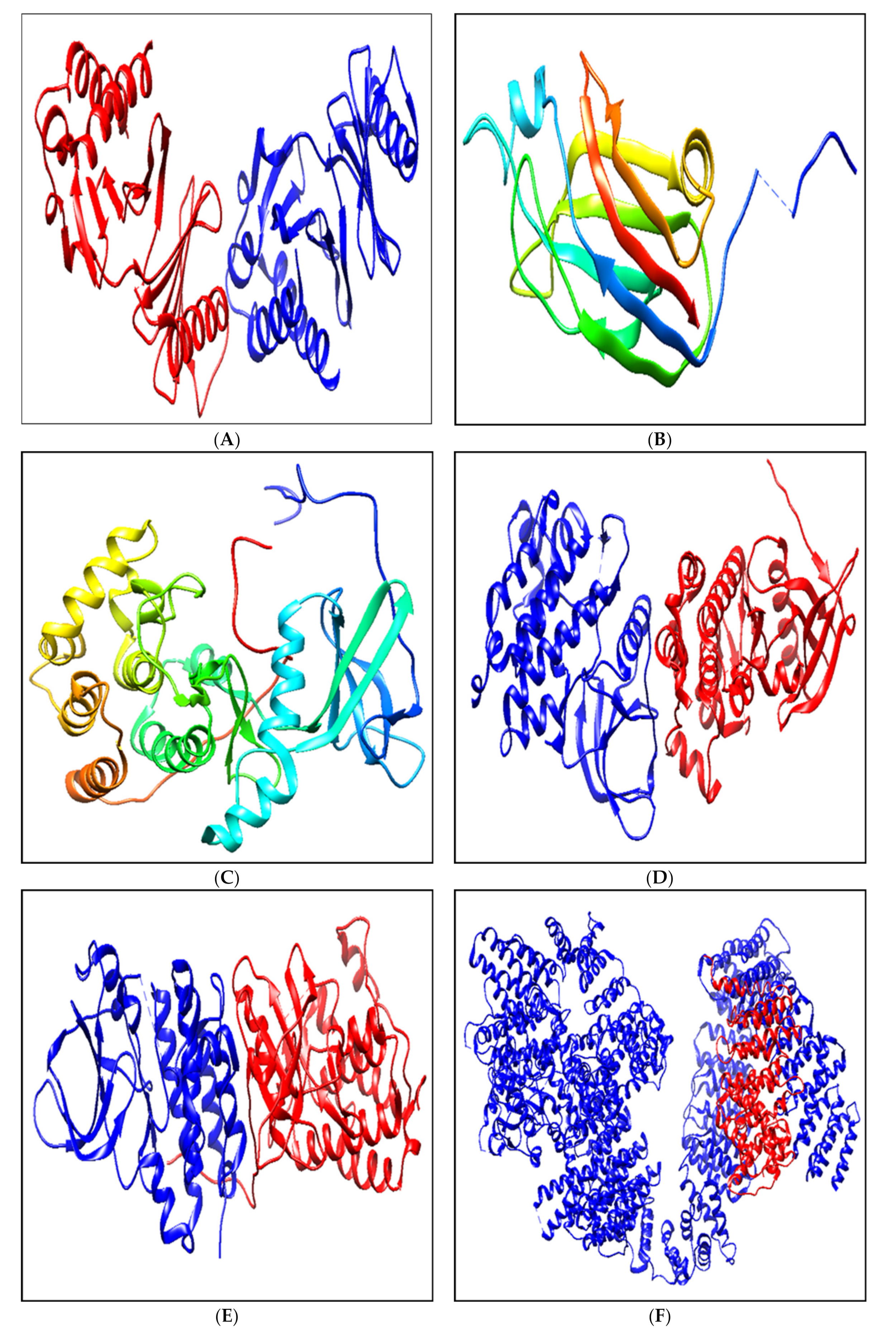
3.5. In Silico Protein Validation
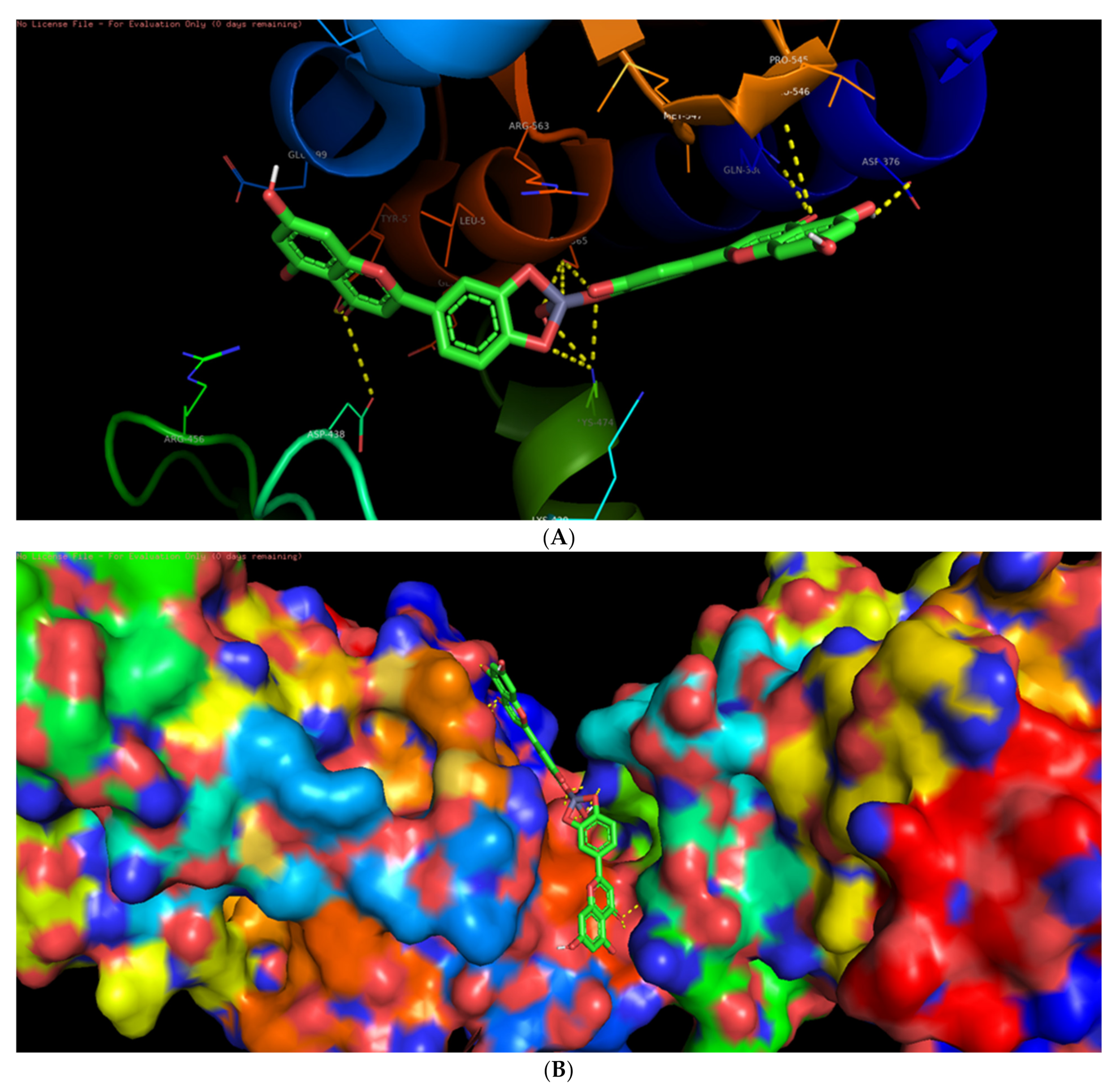
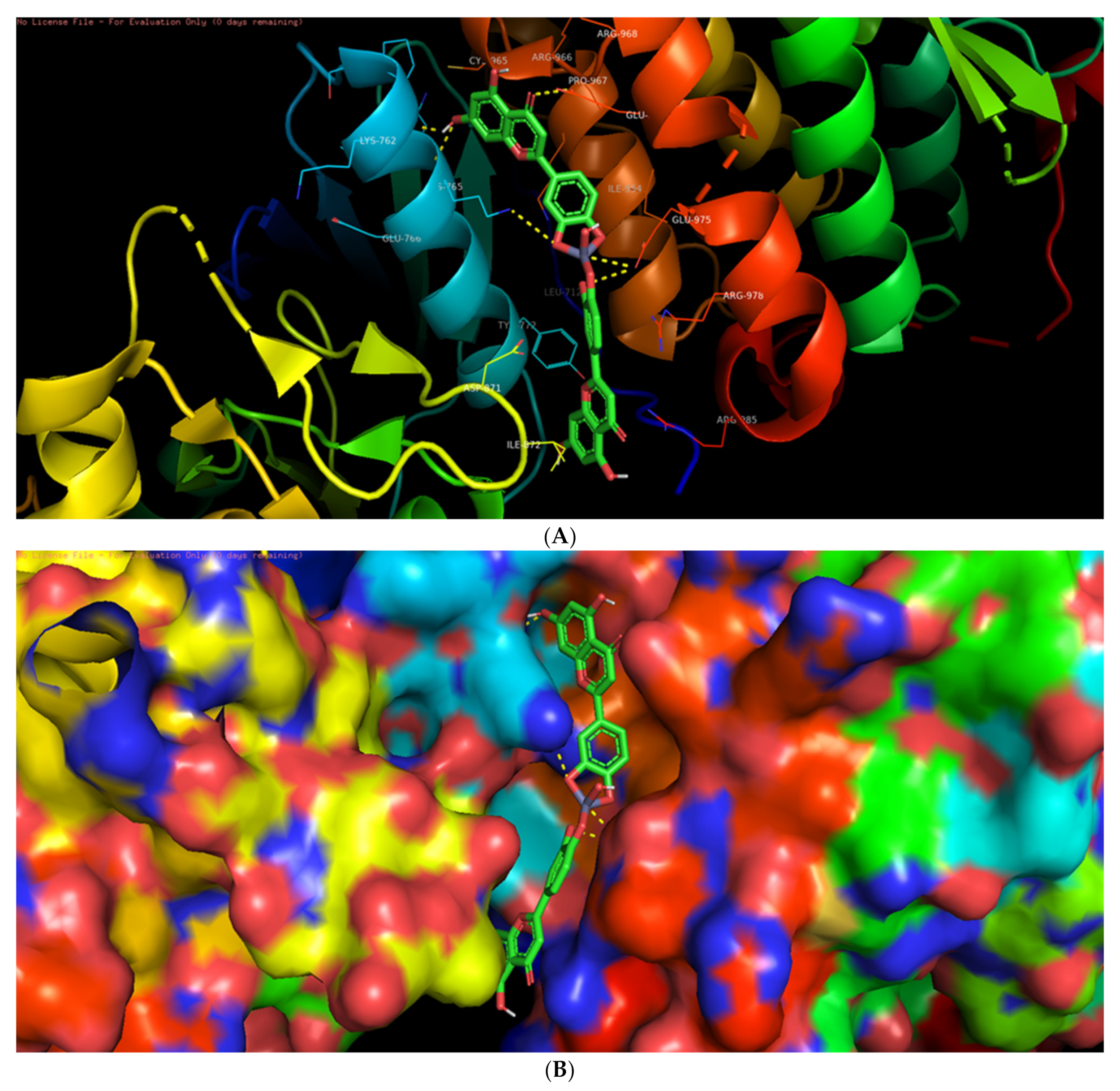
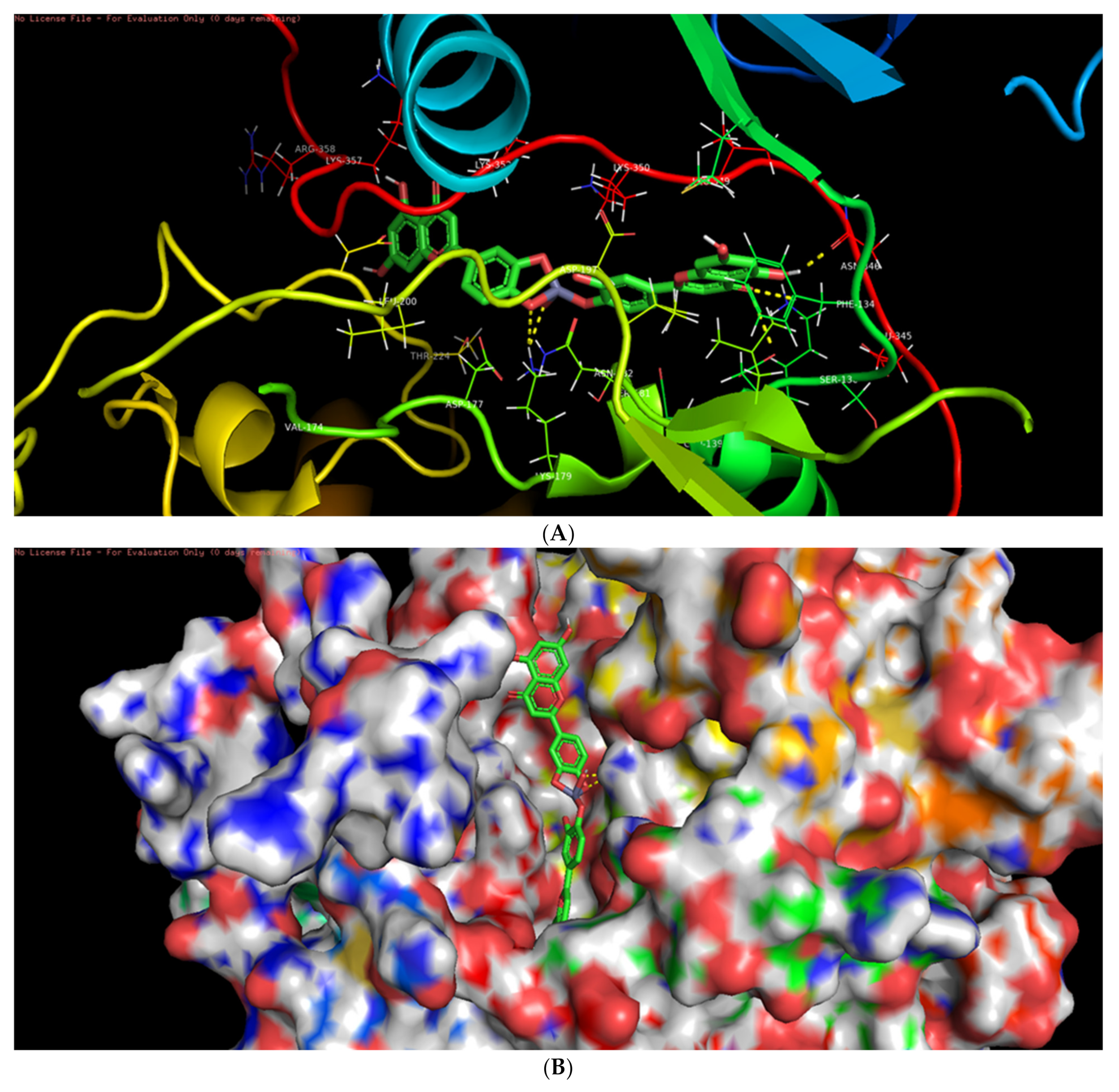
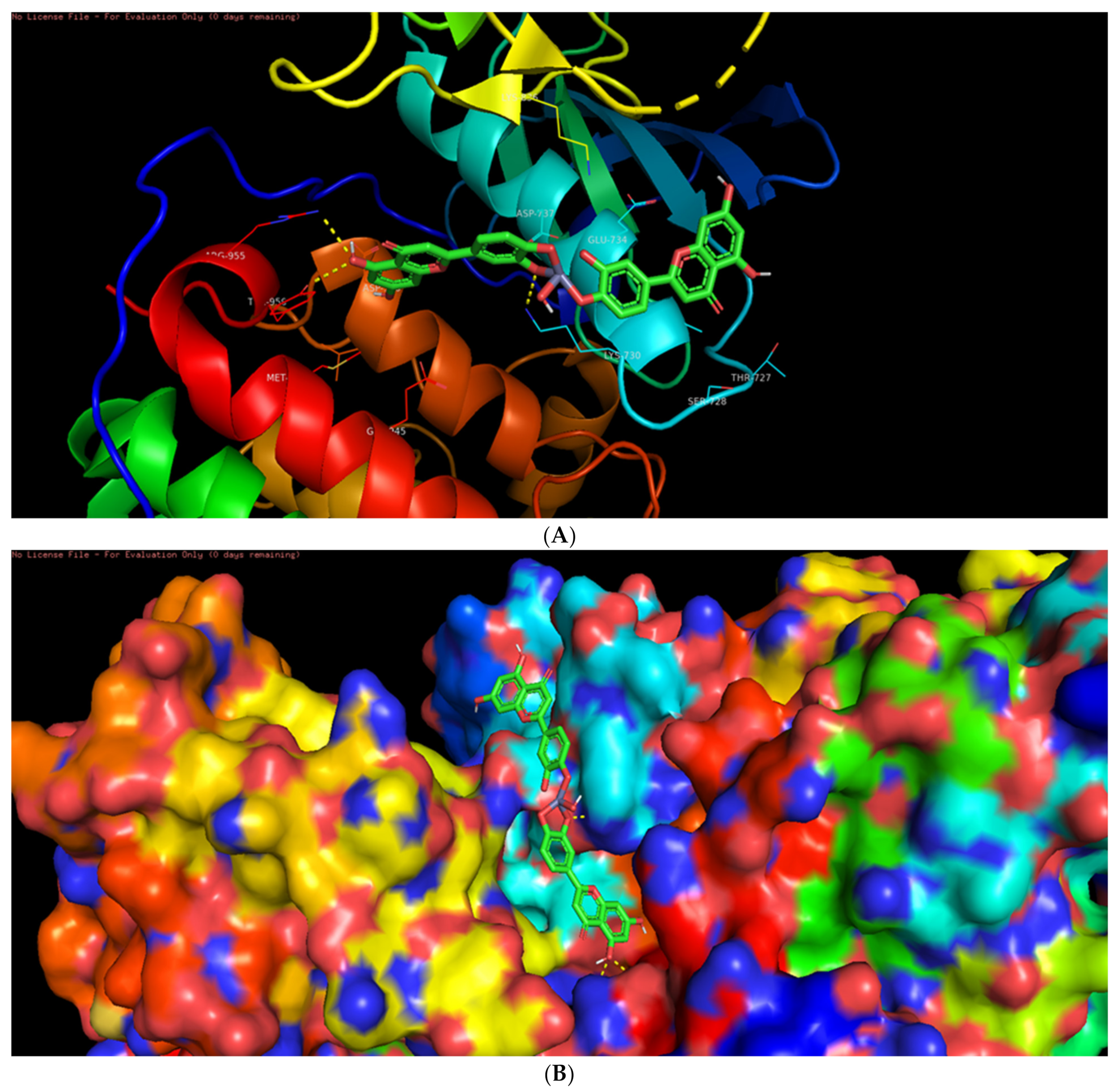
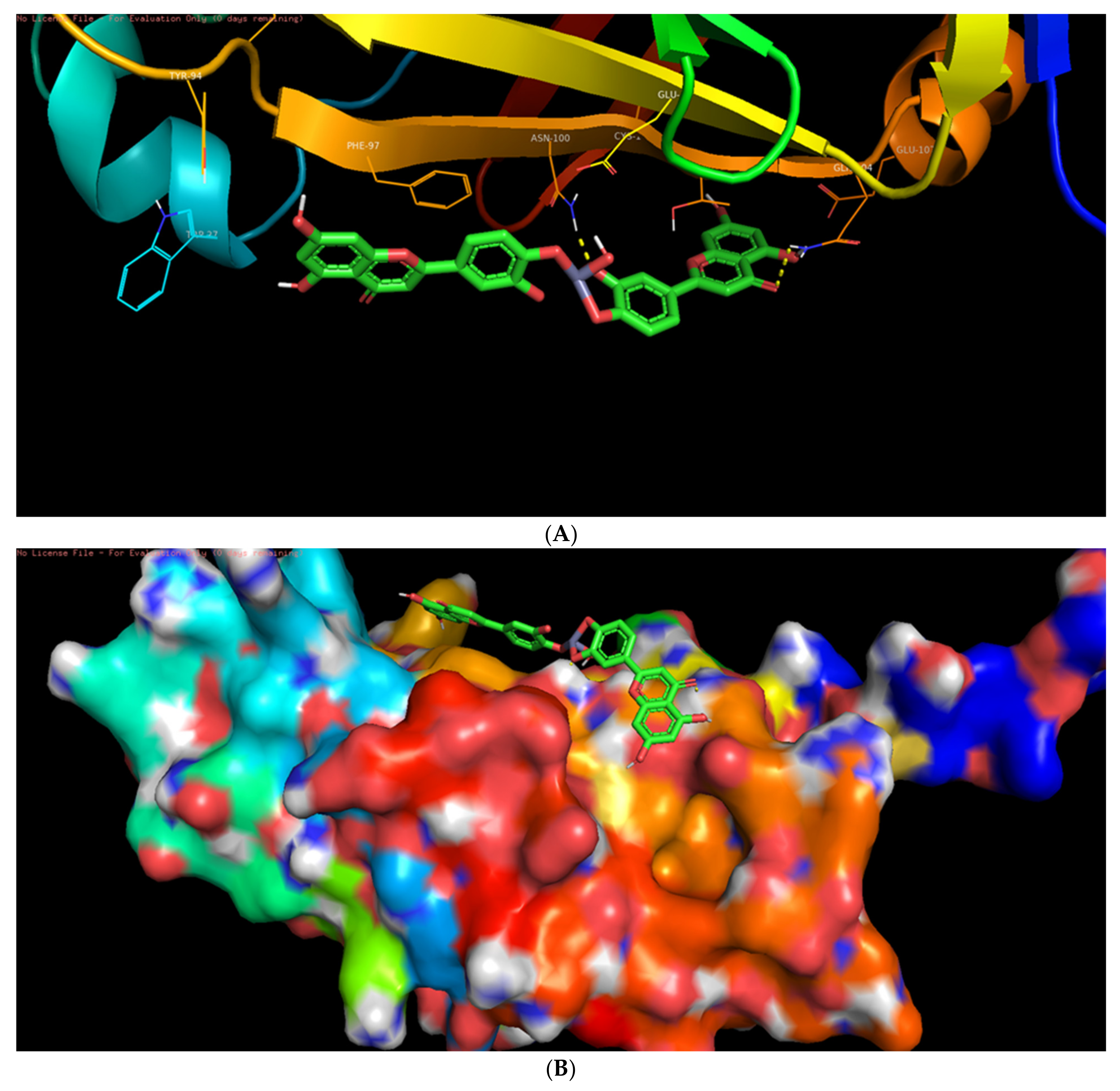
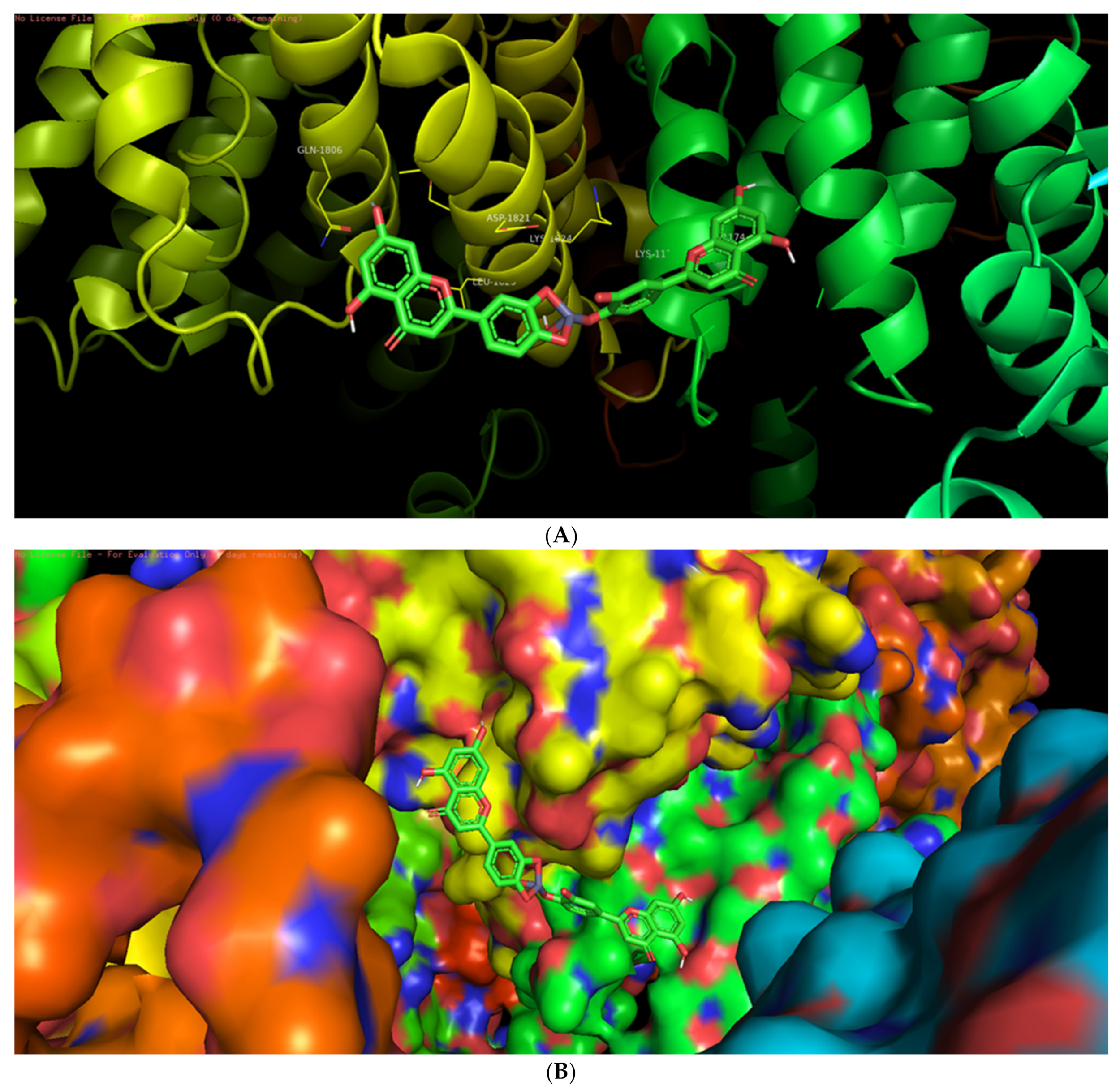
3.6. Molecular Docking Studies
3.7. Interaction of L-ZnONPs with Proteins
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AKT | Protein kinase B |
| DMEM | Dulbecco’s Modified Eagle Medium |
| DR5 | Death receptor 5 |
| EGFR/HER | Endothelial growth factor receptor |
| ER | Estrogen receptor |
| ESP | Estrogen signaling pathway |
| ERK | Extracellular signal-regulated kinase |
| JNK | Jun N-terminal kinase |
| MAPK | Mitogen-activated protein kinase |
| NF-kB | Nuclear factor kappa B |
| PARP | Poly ADP-ribose polymerase |
| PI3K | Phosphatidylinositol 3-kinase |
| PKC | Protein kinase C |
| PLK-1 | Polo-like kinase 1 |
| PR | Progesterone receptor |
| ROS | Reactive oxygen species |
| TNFa | Tumor necrosis factor alpha |
| TLP2 | Tumor progression locus 2 |
| VEGF | Vascular endothelial growth factor |
References
- Sivakumar, P.; Lee, M.; Kim, Y.S.; Shim, M.S. Photo-triggered antibacterial and anticancer activities of zinc oxide nanoparticles. J. Mater. Chem. B 2018, 6, 4852–4871. [Google Scholar] [CrossRef]
- Joshi, S.S.; Patil, P.R.; Nimase, M.S.; Bakare, P.P. Role of ligands in the formation, phase stabilization, structural and magneticproperties of α-Fe2O3 nanoparticles. J. Nanoparticle Res. 2006, 8, 635–643. [Google Scholar] [CrossRef]
- Cheng, X.L.; Zhao, H.; Huo, L.H.; Gao, S.; Zhao, J.G. ZnO nanoparticulate thin film: Preparation, characterization and gas-sensing property. Sens. Actuators B Chem. 2004, 102, 248–252. [Google Scholar] [CrossRef]
- Lee, S.Y.; Shim, E.S.; Kang, H.S.; Pang, S.S.; Kang, J.S. Fabrication of ZnO thin film diode using laser annealing. Thin Solid Films 2005, 473, 31–34. [Google Scholar] [CrossRef]
- Wang, Z.L.; Kong, X.Y.; Ding, Y.; Gao, P.; Hughes, W.L.; Yang, R.; Zhang, Y. Semiconducting and piezoelectric oxide nanostructures induced by polar surfaces. Adv. Funct. Mater. 2004, 14, 943–956. [Google Scholar] [CrossRef]
- Huang, Y.H.; Zhang, Y.; Liu, L.; Fan, S.S.; Wei, Y.; He, J. Controlled synthesis and field emission properties of ZnO nanostructures with different morphologies. J. Nanosci. Nanotechnol. 2006, 6, 787–790. [Google Scholar] [CrossRef] [PubMed]
- Rao, C.N.R.; Müller, A.; Cheetham, A.K. The Chemistry of Nanomaterials: Synthesis, Properties and Applications; John Wiley & Sons: Hoboken, NJ, USA, 2006. [Google Scholar]
- Theodore, L. Nanotechnology: Basic Calculations for Engineers and Scientists; Wiley-VCH: Weinheim, Germany, 2005; p. 446. ISBN 0-471-73951-0. [Google Scholar]
- Wang, X.; Yang, W.; Yang, X. A study on the antibacterial activity of one-dimensional ZnO nanowire arrays: Effects of the orientation and plane surface. Chem. Commun. 2007, 42, 4419–4421. [Google Scholar] [CrossRef] [PubMed]
- McDonald, M.; Mila, I.; Scalbert, A. Precipitation of metal ions by plant polyphenols: Optimal conditions and origin of precipitation. J. Agric. Food Chem. 1996, 44, 599–606. [Google Scholar] [CrossRef]
- Ravi, S.S.; Christena, L.R.; SaiSubramanian, N.; Anthony, S.P. Green synthesized silver nanoparticles for selective colorimetric sensing of Hg2+ in aqueous solution at wide pH range. Analyst 2013, 138, 4370–4377. [Google Scholar] [CrossRef] [PubMed]
- Devatha, C.P.; Thalla, A.K. Chapter 7—Green Synthesis of Nanomaterials. In Synthesis of Inorganic Nanomaterials; Mohan Bhagyaraj, S., Oluwafemi, O.S., Kalarikkal, N., Thomas, S., Eds.; Woodhead Publishing: Cambridge, UK; Shaston, UK, 2018; pp. 169–184. [Google Scholar]
- Selim, Y.; Adel, M.; Ragab, I.; Abd, E.A.M. Green Synthesis of Zinc Oxide Nanoparticles Using Aqueous Extract of Deverra tortuosa and their Cytotoxic Activities. Sci. Rep. 2020, 10, 3445. [Google Scholar] [CrossRef] [Green Version]
- Inamuddin; Shakeel, N.; Imran, A.M.; Kanchi, S.; Abbas, K.H. Green synthesis of ZnO nanoparticles decorated on polyindole functionalized-MCNTs and used as anode material for enzymatic biofuel cell applications. Sci. Rep. 2020, 10, 5052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, K.; Singh, A.; Mukherjee, A.; Bhat, B.; Ramesh, B.; Burman, A. Eclipta alba extract with potential for hair growth promoting activity. J. Ethnopharmacol. 2009, 124, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Shabeer, M.; Khan, G.; Ali, A.; Ullah, Z. Standardization of Eclipta alba (L). Asian J. Res. Chem. 2012, 4, 1825–1828. [Google Scholar]
- Balakrishnan, P.; Sekar, G.; Ramalingam, P.; Nagarasan, S.; Murugesan, V.; Shanmugam, K. DISTINCTIVE PHARMACOLOGICAL ACTIVITIES OF Eclipta alba AND IT’S COUMESTAN WEDELOLACTONE. Indo Am. J. Pharm. Sci. 2018, 5, 2996–3002. [Google Scholar]
- Jadhav, V.M.; Thorat, R.M.; Salaskar, V.J. Chemical composition, pharmacological activities of Eclipta alba. J. Pharm. Res. 2009, 2, 1229–1231. [Google Scholar]
- Seelinger, G.; Merfort, I.; Wölfle, U.; Schempp, C.M. Anti-carcinogenic effects of the flavonoid luteolin. Molecules 2008, 13, 2628–2651. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Rauf, A.; Abu-Izneid, T.; Nadeem, M.; Shariati, M.A.; Khan, I.A.; Imran, A.; Orhan, I.E.; Rizwan, M.; Atif, M.; et al. Luteolin, a flavonoid, as an anticancer agent: A review. Biomed. Pharmacother. 2019, 112, 108612. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Ge, R.; Li, Y.; Liu, S. Luteolin exhibits anti-breast cancer property through up-regulating miR-203. Artif. Cells Nanomed. Biotechnol. 2019, 47, 326532–326571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, W.; Jing, W.; Xing, G.; Xinxuan, W.; Gu, X. Luteolin: Anti-breast Cancer Effects and Mechanisms. J. Explor. Res. Pharmacol. 2018, 3, 85–90. [Google Scholar]
- Song, S.; Su, Z.; Xu, H.; Niu, M.; Chen, X.; Min, H.; Zhang, B.; Sun, G.; Xie, S.; Wang, H.; et al. Luteolin selectively kills STAT3 highly activated gastric cancer cells through enhancing the binding of STAT3 to SHP-1. Cell Death Dis. 2017, 8, e2612. [Google Scholar] [CrossRef]
- Kittiratphatthana, N.; Kukongviriyapan, V.; Prawan, A.; Senggunprai, L. Luteolin induces cholangiocarcinoma cell apoptosis through the mitochondrial-dependent pathway mediated by reactive oxygen species. J. Pharm. Pharmacol. 2016, 68, 1184–1192. [Google Scholar] [CrossRef] [PubMed]
- Dia, V.P.; Pangloli, P. Epithelial-to-Mesenchymal Transition in Paclitaxel-Resistant Ovarian Cancer Cells Is Downregulated by Luteolin. J. Cell Physiol. 2017, 232, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Palko-Labuz, A.; Sroda-Pomianek, K.; Uryga, A.; Kostrzewa-Suslow, E.; Michalak, K. Anticancer activity of baicalein and luteolin studied in colorectal adenocarcinoma LoVo cells and in drug-resistant LoVo/Dx cells. Biomed. Pharmacother. 2017, 88, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Lim, T.; Han, M.S.; Lee, S.H.; Baek, S.H.; Nan, H.Y.; Lee, C. Anticancer effect of luteolin is mediated by downregulation of TAM receptor tyrosine kinases, but not interleukin-8, in non-small cell lung cancer cells. Oncol. Rep. 2017, 37, 1219–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Yang, J.; Wang, J. Modulatory effect of luteolin on redox homeostasis and inflammatory cytokines in a mouse model of liver cancer. Oncol. Lett. 2016, 12, 4767–4772. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.; Byun, S.; Son, J.E.; Oh, M.; Lee, J.; Kang, M.; Heo, Y.-S.; Lee, K.W.; Lee, H.J. Raf and PI3K are the Molecular Targets for the Anti-metastatic Effect of Luteolin. Phytother. Res. 2012, 27, 1481–1488. [Google Scholar]
- Wang, L.M.; Kp, X.; Hn, H.; F, S.; W, Z.; Xie, M.J. Luteolin inhibits proliferation induced by IGF-1 pathway dependent ERalpha in human breast cancer MCF-7 cells. Asian Pac. J. Cancer Prev. APJCP 2012, 13, 1431–1437. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Sasaki, N.; Saito, M.; M, S.; Endo, N.; Kugawa, F.; Ueno, A. Luteolin attenuates doxorubicin-induced cytotoxicity to MCF-7 human breast cancer cells. Biol. Pharm. Bull. 2015, 38, 703–709. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.W.; Zhang, H.D.; Mao, L.; Mao, C.F.; Chen, W.; Cui, M.; Ma, R.; Cao, H.-X.; Jing, C.-W.; Wang, Z.; et al. Luteolin Inhibits Breast Cancer Development and Progression In Vitro and by Suppressing Notch Signaling and Regulating MiRNAs. Cell. Physiol. Biochem. 2015, 37, 1693–1711. [Google Scholar] [CrossRef] [Green Version]
- Brusselmans, K.; Vrolix, R.; Verhoeven, G.; Swinnen, J.V. Induction of cancer cell apoptosis by flavonoids is associated with their ability to inhibit fatty acid synthase activity. J. Biol. Chem. 2005, 280, 5636–5645. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.J.; Oh, S.Y.; Sung, M.K. Luteolin exerts anti-tumor activity through the suppression of epidermal growth factor receptor-mediated pathway in MDA-MB-231 ER-negative breast cancer cells. Food Chem. Toxicol. 2012, 50, 4136–4143. [Google Scholar] [CrossRef]
- Lin, C.H.; Chang, C.Y.; Lee, K.R.; Lin, H.J.; Chen, T.H.; Wan, L. Flavones inhibit breast cancer proliferation through the Akt/FOXO3a signaling pathway. BMC Cancer 2015, 15, 958. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Cheng, H.; Bai, Z.; Li, J. Breast Cancer Cell Line Classification and Its Relevance with Breast Tumor Subtyping. J. Cancer 2017, 8, 3131–3141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Xie, K.P.; Huo, H.N.; Wang, L.M.; Zou, W.; Xie, M. [Inhibitory effect of luteolin on the angiogenesis of chick chorioallantoic membrane and invasion of breast cancer cells via downregulation of AEG-1 and MMP-2]. Sheng Li Xue Bao 2013, 65, 513–518. [Google Scholar]
- Lin, D.; Kuang, G.; Wan, J.; Zhang, X.; Li, H.; Gong, X.; Li, H. Luteolin suppresses the metastasis of triple-negative breast cancer by reversing epithelial-to-mesenchymal transition via downregulation of beta-catenin expression. Oncol. Rep. 2017, 37, 895–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, P.S.; Satelli, A.; Moridani, M.; Jenkins, M.; Rao, U.S. Luteolin induces apoptosis in multidrug resistant cancer cells without affecting the drug transporter function: Involvement of cell line-specific apoptotic mechanisms. Int. J. Cancer 2012, 130, 2703–2714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.H.; Ham, S.; Kwon, T.H.; Kim, M.S.; Lee, D.H.; Kang, J.W.; Oh, S.-R.; Yoon, D.-Y. Luteolin Induces Cell Cycle Arrest and Apoptosis Through Extrinsic and Intrinsic Signaling Pathways in MCF-7 Breast Cancer Cells. J. Environ. Pathol. Toxicol. Oncol. 2014, 33, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Sui, J.Q.; Xie, K.P.; Xie, M.J. Inhibitory effect of luteolin on the proliferation of human breast cancer cell lines induced by epidermal growth factor. Acta Physiol. Sin. 2016, 68, 27–34. [Google Scholar]
- Markaverich, B.; Shoulars, K.; Rodriguez, M. Luteolin Regulation of Estrogen Signaling and Cell Cycle Pathway Genes in MCF-7 Human Breast Cancer Cells. Int. J. Biomed. Sci. IJBS 2011, 7, 101–111. [Google Scholar] [PubMed]
- Nordeen, S.K.; Bona, B.J.; Jones, D.N.; Lambert, J.R.; Jackson, T.A. Endocrine Disrupting Activities of the Flavonoid Nutraceuticals Luteolin and Quercetin. Horm. Cancer 2013, 4, 293–300. [Google Scholar] [CrossRef]
- Kim, J.E.; Son, J.E.; Jang, Y.J.; Lee, D.E.; Kang, N.J.; Jung, S.K.; Heo, J.-S.; Lee, K.W.; Lee, H.J. Luteolin, a Novel Natural Inhibitor of Tumor Progression Locus 2 Serine/Threonine Kinase, Inhibits Tumor Necrosis Factor-α-Induced Cyclooxygenase-2 Expression in JB6 Mouse Epidermis Cells. J. Pharmacol. Exp. Ther. 2011, 338, 1013. [Google Scholar] [CrossRef] [PubMed]
- Ferriola, P.C.; Cody, V.; Middleton, E. Protein kinase C inhibition by plant flavonoids: Kinetic mechanisms and structure-activity relationships. Biochem. Pharmacol. 1989, 38, 1617–1624. [Google Scholar] [CrossRef]
- Sourvinos, G.; Tsatsanis, C.; Spandidos, D.A. Overexpression of the Tpl-2/Cot oncogene in human breast cancer. Oncogene 1999, 18, 4968–4973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, M.T.; Liang, Y.; Besch-Williford, C.; Hyder, S.M. Luteolin inhibits lung metastasis, cell migration, and viability of triple-negative breast cancer cells. Breast Cancer Targets Ther. 2017, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Du, G.J.; Song, Z.H.; Lin, H.H.; Han, X.F.; Zhang, S.; Yang, Y.M. Luteolin as a glycolysis inhibitor offers superior efficacy and lesser toxicity of doxorubicin in breast cancer cells. Biochem. Biophys. Res. Commun. 2008, 372, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.W.; Suh, Y.J. Synergistic apoptotic effect of celecoxib and luteolin on breast cancer cells. Oncol. Rep. 2012, 29, 819–825. [Google Scholar] [CrossRef] [Green Version]
- Jeon, Y.W.; Ahn, Y.E.; Chung, W.S.; Choi, H.J.; Suh, Y.J. Synergistic effect between celecoxib and luteolin is dependent on estrogen receptor in human breast cancer cells. Tumor Biol. 2015, 36, 6349–6359. [Google Scholar] [CrossRef]
- Tanino, R.; Amano, Y.; Tong, X.; Sun, R.; Tsubata, Y.; Harada, M.; Fujita, Y.; Isobe, T. Anticancer Activity of ZnO Nanoparticles against Human Small-Cell Lung Cancer in an Orthotopic Mouse Model. Mol. Cancer Ther. 2020, 19, 502. [Google Scholar] [CrossRef] [Green Version]
- Bisht, G.; Rayamajhi, S. ZnO Nanoparticles: A Promising Anticancer Agent. Nanobiomedicine 2016, 3, 1. [Google Scholar] [CrossRef]
- Rasmussen, J.W.; Martinez, E.; Louka, P.; Wingett, D.G. Zinc oxide nanoparticles for selective destruction of tumor cells and potential for drug delivery applications. Expert Opin. Drug Deliv. 2010, 7, 1063–1077. [Google Scholar] [CrossRef] [Green Version]
- Kadhem, H.A.; Ibraheem, S.A.; Jabir, M.S.; Kadhim, A.A.; Taqi, Z.J.; Florin, M.D. Zinc Oxide Nanoparticles Induces Apoptosis in Human Breast Cancer Cells via Caspase-8 and P53 Pathway. Nano Biomed. Eng. 2019, 11, 35–43. [Google Scholar] [CrossRef]
- Food for Human Consumption—Substances Generally Recognized as Safe; 21CFR182.8991; FDA: Silver Spring, MD, USA, 2019.
- Prasad, K.S.; Shashanka, K.P.; Ravindra, V.; Ghada, L.; Ashwini, P.; Prasad, M.N.N.; Sandeep, K.S.; Najat, M.; Asad, S.; Shivamallu, C. Antitumor Potential of Green Synthesized ZnONPs Using Root Extract ofWithania somnifera against Human Breast Cancer Cell Line. Separations 2021, 8, 8. [Google Scholar] [CrossRef]
- Denizot, F.; Lang, R. Rapid colorimetric assay for cell growth and survival. Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Methods 1986, 89, 271–277. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Naz, A.; Bano, K.; Akhtar, N. Conformational analysis (geometry optimization) of nucleosidic antitumor antibiotic showdomycin by Arguslab 4 software. Pak. J. Pharm. Sci. 2009, 22, 78–82. [Google Scholar] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Xia, M.; Chen, J.; Deng, F.; Yuan, R.; Zhang, X.; Shen, F. Data set for phylogenetic tree and RAMPAGE Ramachandran plot analysis of SODs in Gossypium raimondii and G. arboreum. Data Brief 2016, 9, 345–348. [Google Scholar] [CrossRef] [Green Version]
- Junsu, K.; Hahnbeom, P.; Heo, L.; Seok, C. GalaxyWEB server for protein structure prediction and refinement. Nucleic Acids Res. 2012, 40, W294–W297. [Google Scholar]
- Dallakyan, S.; Olson, A.J. Small-molecule library screening by docking with PyRx. Methods Mol. Biol. 2015, 1263, 243–250. [Google Scholar]
- Majumdar, D.; Jung, K.H.; Zhang, H.; Nannapaneni, S.; Wang, X.; Amin, A.; Chen, Z.; Chen, Z.G.; Shin, D.M. Luteolin nanoparticle in chemoprevention: In vitro and in vivo anticancer activity. Cancer Prev. Res. 2014, 7, 65–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhari, R.; Li, Z. PyMine: A PyMOL plugin to integrate and visualize data for drug discovery. BMC Res. Notes 2015, 8, 517. [Google Scholar] [CrossRef] [PubMed] [Green Version]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl No. | Protein PDB ID | Favored Region % | Allowed Region % | Outlier Region % |
|---|---|---|---|---|
| 1 | 1Q4O | 96.6 | 2.7 | 0.7 |
| 2 | 2FK9 | 96.6 | 3.4 | 0.0 |
| 3 | 2LAV | 98.7 | 1.3 | 0.0 |
| 4 | 3PP0 | 97.8 | 2.2 | 0.0 |
| 5 | 4RIW | 98.5 | 1.5 | 0.0 |
| 6 | 5YZ0 | 98.1 | 1.9 | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kollur, S.P.; Prasad, S.K.; Pradeep, S.; Veerapur, R.; Patil, S.S.; Amachawadi, R.G.; S, R.P.; Lamraoui, G.; Al-Kheraif, A.A.; Elgorban, A.M.; et al. Luteolin-Fabricated ZnO Nanostructures Showed PLK-1 Mediated Anti-Breast Cancer Activity. Biomolecules 2021, 11, 385. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11030385
Kollur SP, Prasad SK, Pradeep S, Veerapur R, Patil SS, Amachawadi RG, S RP, Lamraoui G, Al-Kheraif AA, Elgorban AM, et al. Luteolin-Fabricated ZnO Nanostructures Showed PLK-1 Mediated Anti-Breast Cancer Activity. Biomolecules. 2021; 11(3):385. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11030385
Chicago/Turabian StyleKollur, Shiva Prasad, Shashanka K. Prasad, Sushma Pradeep, Ravindra Veerapur, Sharanagouda S. Patil, Raghavendra G. Amachawadi, Rajendra Prasad S, Ghada Lamraoui, Abdulaziz A. Al-Kheraif, Abdallah M. Elgorban, and et al. 2021. "Luteolin-Fabricated ZnO Nanostructures Showed PLK-1 Mediated Anti-Breast Cancer Activity" Biomolecules 11, no. 3: 385. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11030385