
L-Proline Activates Mammalian Target of Rapamycin Complex 1 and Modulates Redox Environment in Porcine Trophectoderm Cells

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Cell Viability Assay
2.4. Determination of ROS
2.5. Analysis of GSH and Proline Concentration
2.6. Quantitative PCR Analysis
2.7. Western Blot Analyses
2.8. Statistical Analysis
3. Results
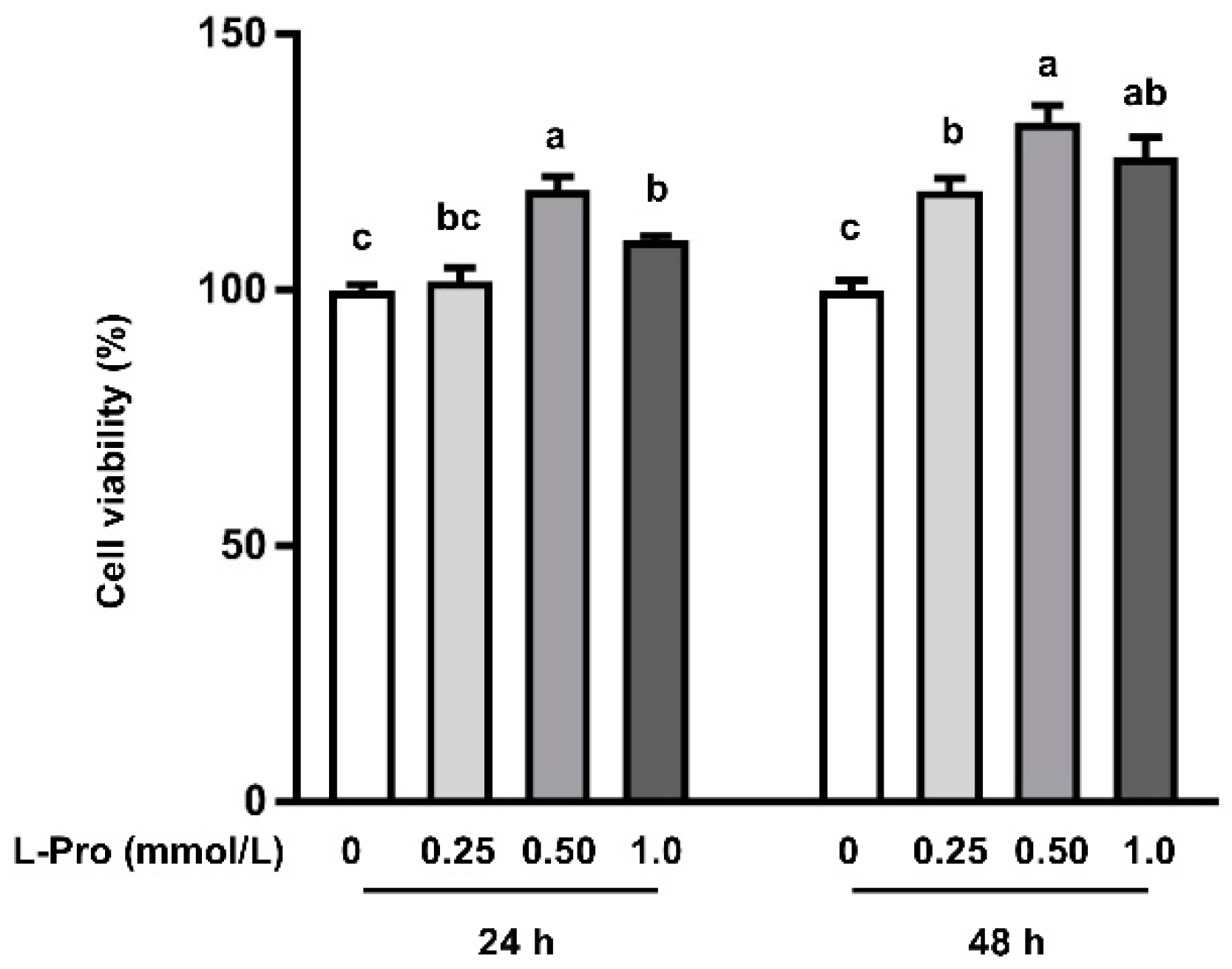
3.1. Proline Enhances Cell Viability in pTr2 Cells
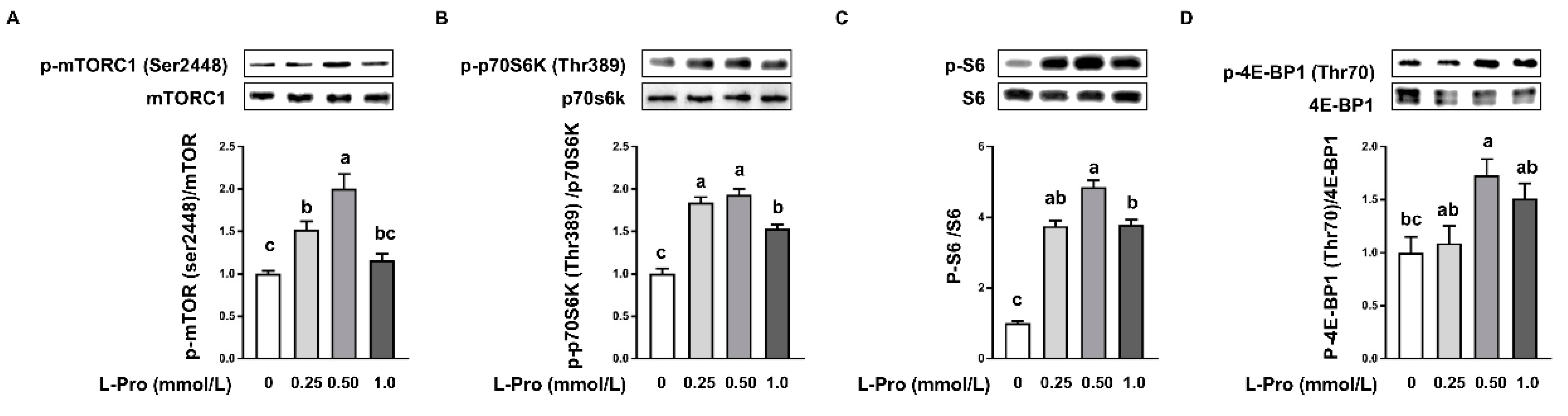
3.2. Proline Activates mTORC1 Cell Signaling Pathway in pTr2 Cells
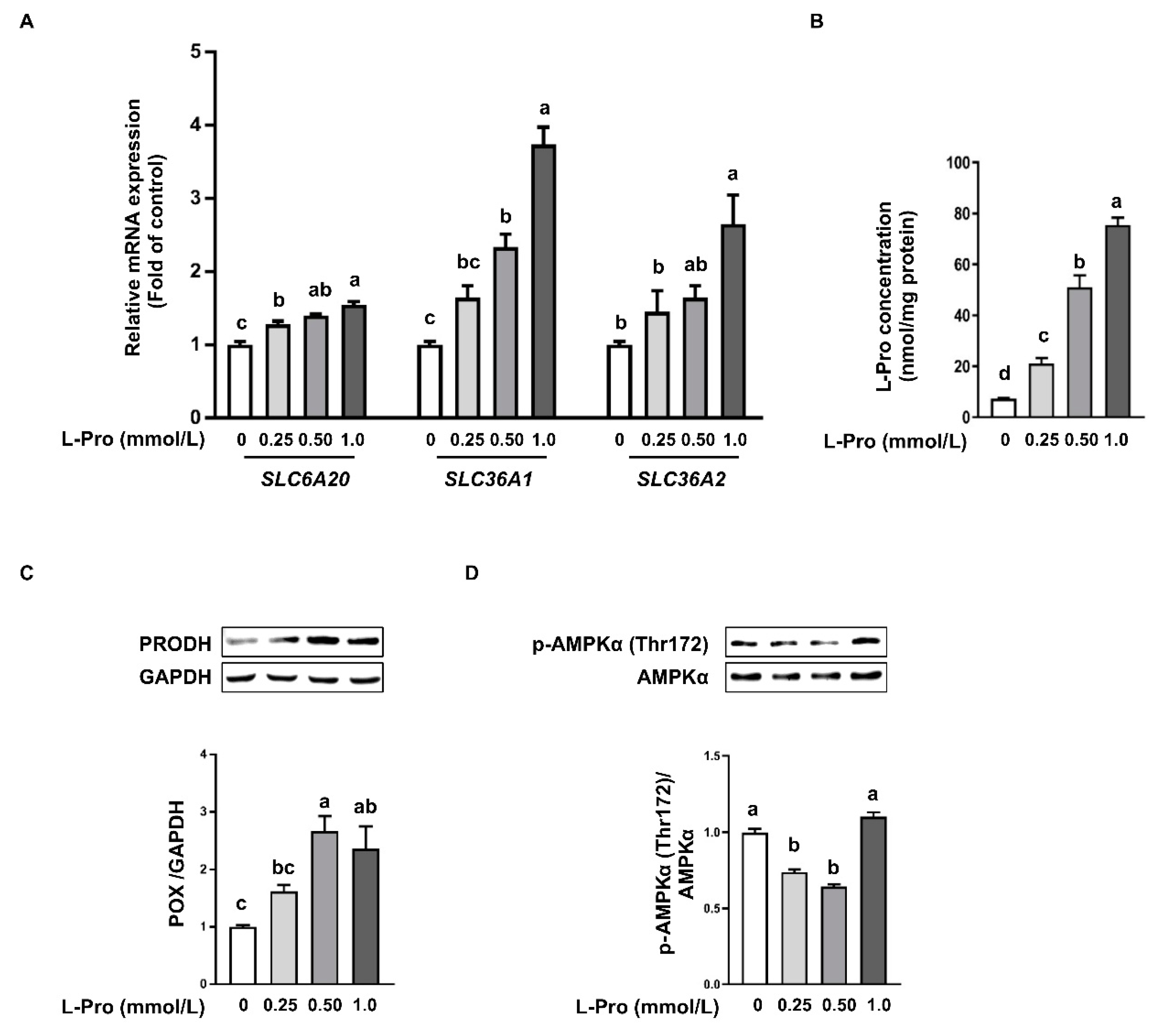
3.3. Proline Enhances Proline Transporter Expression and Catabolism in pTr2 Cells
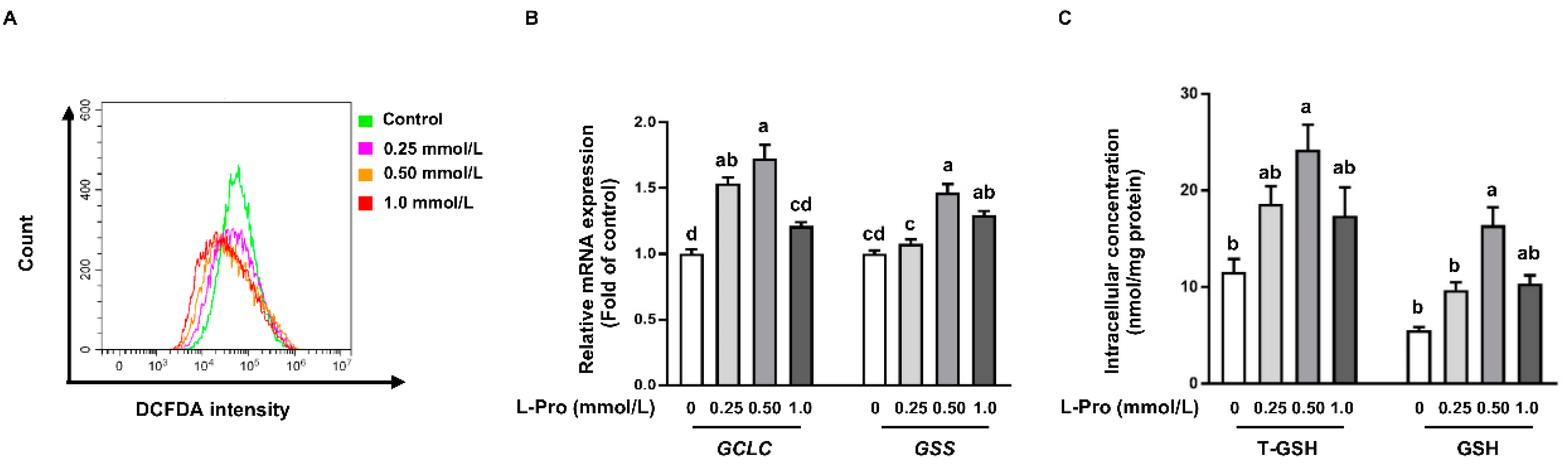
3.4. Proline Reduces ROS Production in pTr2 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Luo, Z.; Xu, X.; Sho, T.; Zhang, J.; Xu, W.; Yao, J.; Xu, J. ROS-induced autophagy regulates porcine trophectoderm cell apoptosis, proliferation, and differentiation. Am. J. Physiol. Physiol. 2019, 316, C198–C209. [Google Scholar] [CrossRef]
- Gao, K.; Jiang, Z.; Lin, Y.; Zheng, C.; Zhou, G.; Chen, F.; Yang, L.; Wu, G. Dietary l-arginine supplementation enhances placental growth and reproductive performance in sows. Amino Acids 2011, 42, 2207–2214. [Google Scholar] [CrossRef]
- E Spencer, T.; Bazer, F.W. Conceptus signals for establishment and maintenance of pregnancy. Reprod. Biol. Endocrinol. 2004, 2, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, W.; Jung, S.; Bazer, F.W.; Kim, J. Stem cell factor-induced AKT cell signaling pathway: Effects on porcine trophectoderm and uterine luminal epithelial cells. Gen. Comp. Endocrinol. 2017, 250, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Bazer, F.W.; Wu, G.; Spencer, T.E.; Johnson, G.A.; Burghardt, R.C.; Bayless, K. Novel pathways for implantation and establishment and maintenance of pregnancy in mammals. Mol. Hum. Reprod. 2009, 16, 135–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazer, F.W.; E Spencer, T.; A Johnson, G.; Burghardt, R.C.; Wu, G. Comparative aspects of implantation. Reproduction 2009, 138, 195–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, W.; Kim, J.; Bazer, F.W.; Song, G. Epidermal growth factor stimulates proliferation and migration of porcine trophectoderm cells through protooncogenic protein kinase 1 and extracellular-signal-regulated kinases 1/2 mitogen-activated protein kinase signal transduction cascades during early pregnancy. Mol. Cell. Endocrinol. 2013, 381, 302–311. [Google Scholar] [CrossRef]
- Gelber, S.E.; Brent, E.; Redecha, P.; Perino, G.; Tomlinson, S.; Davisson, R.L.; Salmon, J.E. Prevention of Defective Placentation and Pregnancy Loss by Blocking Innate Immune Pathways in a Syngeneic Model of Placental Insufficiency. J. Immunol. 2015, 195, 1129–1138. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.; Tan, B.; Yin, Y.; Gao, H.; Li, X.; Jaeger, L.A.; Bazer, F.W.; Wu, G. l-Arginine stimulates the mTOR signaling pathway and protein synthesis in porcine trophectoderm cells. J. Nutr. Biochem. 2012, 23, 1178–1183. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Wang, X.; Yin, Y.; Li, X.; Gao, H.; Bazer, F.W.; Wu, G. Putrescine Stimulates the mTOR Signaling Pathway and Protein Synthesis in Porcine Trophectoderm Cells. Biol. Reprod. 2014, 91, 106. [Google Scholar] [CrossRef]
- Kim, J.; Song, G.; Wu, G.; Gao, H.; Johnson, G.A.; Bazer, F.W. Arginine, Leucine, and Glutamine Stimulate Proliferation of Porcine Trophectoderm Cells Through the MTOR-RPS6K-RPS6-EIF4EBP1 Signal Transduction Pathway. Biol. Reprod. 2013, 88, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.; Luo, W.; Li, S.; Zhao, S.; Sho, T.; Xu, X.; Zhang, J.; Xu, W.; Xu, J. Reactive oxygen species mediated placental oxidative stress, mitochondrial content, and cell cycle progression through mitogen-activated protein kinases in intrauterine growth restricted pigs. Reprod. Biol. 2018, 18, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Chason, R.J.; Csokmay, J.; Segars, J.H.; DeCherney, A.H.; Armant, D.R. Environmental and epigenetic effects upon preimplantation embryo metabolism and development. Trends Endocrinol. Metab. 2011, 22, 412–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, L.P.; Al-Hasan, Y. Impact of Oxidative Stress in Fetal Programming. J. Pregnancy 2012, 2012, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Dennery, P.A. Oxidative stress in development: Nature or nurture? Free Radic. Biol. Med. 2010, 49, 1147–1151. [Google Scholar] [CrossRef] [PubMed]
- Hitchler, M.J.; Domann, F.E. An epigenetic perspective on the free radical theory of development. Free Radic. Biol. Med. 2007, 43, 1023–1036. [Google Scholar] [CrossRef] [Green Version]
- Burton, G.; Jaunaiux, E. Maternal vascularisation of the human placenta: Does the embryo develop in a hypoxic environment? Gynécologie Obs. Fertil. 2001, 29, 503–508. [Google Scholar] [CrossRef]
- Houghton, F.D. Energy metabolism of the inner cell mass and trophectoderm of the mouse blastocyst. Differentiation 2006, 74, 11–18. [Google Scholar] [CrossRef]
- Manes, C. Cyanide-resistant reduction of nitroblue tetrazolium and hydrogen peroxide production by the rabbit blastocyst. Mol. Reprod. Dev. 1992, 31, 114–121. [Google Scholar] [CrossRef]
- Burton, G.J. Oxygen, the Janus gas; its effects on human placental development and function. J. Anat. 2009, 215, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, M. Oxidative Stress and Redox Regulation on In Vitro Development of Mammalian Embryos. J. Reprod. Dev. 2012, 58, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Mun, S.-E.; Sim, B.-W.; Yoon, S.-B.; Jeong, P.-S.; Yang, H.-J.; Choi, S.-A.; Park, Y.-H.; Kim, Y.-H.; Kang, P.; Jeong, K.-J.; et al. Dual effect of fetal bovine serum on early development depends on stage-specific reactive oxygen species demands in pigs. PLoS ONE 2017, 12, e0175427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papis, K.; Poleszczuk, O.; Wenta-Muchalska, E.; Modlinski, J.A. Melatonin effect on bovine embryo development in vitro in relation to oxygen concentration. J. Pineal Res. 2007, 43, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Davis, T.A.; Jaeger, L.A.; Johnson, G.A.; Kim, S.W.; Knabe, D.A.; Meininger, C.J.; Spencer, T.E.; Yin, Y.-L. Important roles for the arginine family of amino acids in swine nutrition and production. Livest. Sci. 2007, 112, 8–22. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Burghardt, R.C.; Johnson, G.A.; Kim, S.W.; Knabe, D.A.; Li, P.; Li, X.; McKnight, J.R.; Satterfield, M.C.; et al. Proline and hydroxyproline metabolism: Implications for animal and human nutrition. Amino Acids 2011, 40, 1053–1063. [Google Scholar] [CrossRef] [Green Version]
- Natarajan, S.K.; Zhu, W.; Liang, X.; Zhang, L.; Demers, A.J.; Zimmerman, M.C.; Simpson, M.A.; Becker, D.F. Proline dehydrogenase is essential for proline protection against hydrogen peroxide-induced cell death. Free Radic. Biol. Med. 2012, 53, 1181–1191. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, N.; Dickman, M.B.; Becker, D.F. Proline modulates the intracellular redox environment and protects mammalian cells against oxidative stress. Free Radic. Biol. Med. 2008, 44, 671–681. [Google Scholar] [CrossRef] [Green Version]
- Popovic, P.J.; Zeh, I.H.J.; Ochoa, J.B. Arginine and Immunity. J. Nutr. 2007, 137, 1681S–1686S. [Google Scholar] [CrossRef] [Green Version]
- Moinard, C.; Cynober, L.; De Bandt, J.-P. Polyamines: Metabolism and implications in human diseases. Clin. Nutr. 2005, 24, 184–197. [Google Scholar] [CrossRef]
- Liu, N.; Dai, Z.; Zhang, Y.; Chen, J.; Yang, Y.; Wu, G.; Tso, P.; Wu, Z. Maternal L-proline supplementation enhances fetal survival, placental development, and nutrient transport in mice. Biol. Reprod. 2018, 100, 1073–1081. [Google Scholar] [CrossRef]
- Zhang, L.; Xue, X.; Yan, J.; Yan, L.-Y.; Jin, X.-H.; Zhu, X.-H.; He, Z.-Z.; Liu, J.; Li, R.; Qiao, J. L-proline: A highly effective cryoprotectant for mouse oocyte vitrification. Sci. Rep. 2016, 6, 26326. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline Mechanisms of Stress Survival. Antioxidants Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wondrak, G.T.; Jacobson, M.K.; Jacobson, E.L. Identification of Quenchers of Photoexcited States as Novel Agents for Skin Photoprotection. J. Pharmacol. Exp. Ther. 2004, 312, 482–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaul, S.; Sharma, S.S.; Mehta, I.K. Free radical scavenging potential of L-proline: Evidence from in vitro assays. Amino Acids 2006, 34, 315–320. [Google Scholar] [CrossRef]
- Liu, N.; Ma, X.; Luo, X.; Zhang, Y.; He, Y.; Dai, Z.; Yang, Y.; Wu, G.; Wu, Z. l-Glutamine Attenuates Apoptosis in Porcine Enterocytes by Regulating Glutathione-Related Redox Homeostasis. J. Nutr. 2018, 148, 526–534. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Meininger, C.J. Analysis of Citrulline, Arginine, and Methylarginines Using High-Performance Liquid Chromatography. Methods Enzymol. 2008, 440, 177–189. [Google Scholar] [CrossRef]
- Wu, G.; Davis, P.K.; Flynn, N.E.; Knabe, D.A.; Davidson, J.T. Endogenous synthesis of arginine plays an important role in maintaining arginine homeostasis in postweaning growing pigs. J. Nutr. 1997, 127, 2342–2349. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Dai, Z.; Zhang, Y.; Jia, H.; Chen, J.; Sun, S.; Wu, G.; Wu, Z. Maternal l-proline supplementation during gestation alters amino acid and polyamine metabolism in the first generation female offspring of C57BL/6J mice. Amino Acids 2019, 51, 805–811. [Google Scholar] [CrossRef]
- Wang, W.; Wu, Z.; Lin, G.; Hu, S.; Wang, B.; Dai, Z.; Wu, G. Glycine Stimulates Protein Synthesis and Inhibits Oxidative Stress in Pig Small Intestinal Epithelial Cells. J. Nutr. 2014, 144, 1540–1548. [Google Scholar] [CrossRef] [Green Version]
- Reddy, D.; Kumavath, R.; Ghosh, P.; Barh, D. Lanatoside C Induces G2/M Cell Cycle Arrest and Suppresses Cancer Cell Growth by Attenuating MAPK, Wnt, JAK-STAT, and PI3K/AKT/mTOR Signaling Pathways. Biomolecules 2019, 9, 792. [Google Scholar] [CrossRef] [Green Version]
- Baek, S.Y.; Hwang, U.W.; Suk, H.Y.; Kim, Y.W. Hemistepsin A Inhibits Cell Proliferation and Induces G0/G1-Phase Arrest, Cellular Senescence and Apoptosis Via the AMPK and p53/p21 Signals in Human Hepatocellular Carcinoma. Biomolecules 2020, 10, 713. [Google Scholar] [CrossRef]
- Bordoloi, D.; Banik, K.; Padmavathi, G.; Vikkurthi, R.; Harsha, C.; Roy, N.K.; Singh, A.K.; Monisha, J.; Wang, H.; Kumar, A.P.; et al. TIPE2 Induced the Proliferation, Survival, and Migration of Lung Cancer Cells Through Modulation of Akt/mTOR/NF-κB Signaling Cascade. Biomolecules 2019, 9, 836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandasamy, P.; Gyimesi, G.; Kanai, Y.; Hediger, M.A. Amino acid transporters revisited: New views in health and disease. Trends Biochem. Sci. 2018, 43, 752–789. [Google Scholar] [CrossRef] [PubMed]
- Brandsch, M. Transport of L-proline, L-proline-containing peptides and related drugs at mammalian epithelial cell membranes. Amino Acids 2006, 31, 119–136. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Johnson, G.A.; Herring, C.; Seo, H.; Dai, Z.; Wang, J.; Wu, Z.; Wang, X. Functional amino acids in the development of the pig placenta. Mol. Reprod. Dev. 2017, 84, 870–882. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Hu, J.; Johnson, G.A.; Spencer, T.E. Polyamine Synthesis from Proline in the Developing Porcine Placenta. Biol. Reprod. 2005, 72, 842–850. [Google Scholar] [CrossRef]
- Rosario, F.J.; Jansson, N.; Kanai, Y.; Prasad, P.D.; Powell, T.L.; Jansson, T. Maternal Protein Restriction in the Rat Inhibits Placental Insulin, mTOR, and STAT3 Signaling and Down-Regulates Placental Amino Acid Transporters. Endocrinology 2011, 152, 1119–1129. [Google Scholar] [CrossRef] [Green Version]
- Kavitha, J.V.; Rosario, F.J.; Nijland, M.J.; McDonald, T.J.; Wu, G.; Kanai, Y.; Powell, T.L.; Nathanielsz, P.W.; Jansson, T. Down-regulation of placental mTOR, insulin/IGF-I signaling, and nutrient transporters in response to maternal nutrient restriction in the baboon. FASEB J. 2014, 28, 1294–1305. [Google Scholar] [CrossRef] [Green Version]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 169, 361–371. [Google Scholar] [CrossRef]
- Lopatynska-Mazurek, M.; Pankowska, A.; Gibula-Tarlowska, E.; Pietura, R.; Kotlinska, J. Rapamycin Improves Recognition Memory and Normalizes Amino-Acids and Amines Levels in the Hippocampal Dentate Gyrus in Adult Rats Exposed to Ethanol during the Neonatal Period. Biomolecules 2021, 11, 362. [Google Scholar] [CrossRef]
- Washington, J.M.; Rathjen, J.; Felquer, F.; Lonic, A.; Bettess, M.D.; Hamra, N.; Semendric, L.; Tan, B.S.N.; Lake, J.-A.; Keough, R.A.; et al. l-Proline induces differentiation of ES cells: A novel role for an amino acid in the regulation of pluripotent cells in culture. Am. J. Physiol. Physiol. 2010, 298, C982–C992. [Google Scholar] [CrossRef] [PubMed]
- Brunton, J.A.; Baldwin, M.P.; Hanna, R.A.; Bertolo, R.F. Proline Supplementation to Parenteral Nutrition Results in Greater Rates of Protein Synthesis in the Muscle, Skin, and Small Intestine in Neonatal Yucatan Miniature Piglets. J. Nutr. 2012, 142, 1004–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyde, R.; Taylor, P.M.; Hundal, H.S. Amino acid transporters: Roles in amino acid sensing and signalling in animal cells. Biochem. J. 2003, 373, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Bazer, F.W.; Datta, S.; Johnson, G.A.; Li, P.; Satterfield, M.C.; Spencer, T.E. Proline metabolism in the conceptus: Implications for fetal growth and development. Amino Acids 2008, 35, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, R.; Gallart, T.; Lluis, C.; Franco, R. Role of glutamate on T-cell mediated immunity. J. Neuroimmunol. 2007, 185, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Zarse, K.; Schmeisser, S.; Groth, M.; Priebe, S.; Beuster, G.; Kuhlow, D.; Guthke, R.; Platzer, M.; Kahn, C.R.; Ristow, M. Impaired Insulin/IGF1 Signaling Extends Life Span by Promoting Mitochondrial L-Proline Catabolism to Induce a Transient ROS Signal. Cell Metab. 2012, 15, 451–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Zhao, L.; Liu, J.; Liu, A.; Jia, C.; Ma, D.; Jiang, Y.; Bai, X. Multi-mechanisms are involved in reactive oxygen species regulation of mTORC1 signaling. Cell. Signal. 2010, 22, 1469–1476. [Google Scholar] [CrossRef]
- Chen, L.; Xu, B.; Liu, L.; Luo, Y.; Yin, J.; Zhou, H.; Chen, W.; Shen, T.; Han, X.; Huang, S. Hydrogen peroxide inhibits mTOR signaling by activation of AMPKα leading to apoptosis of neuronal cells. Lab. Investig. 2010, 90, 762–773. [Google Scholar] [CrossRef]
- Myatt, L.; Cui, X. Oxidative stress in the placenta. Histochem. Cell Biol. 2004, 122, 369–382. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Accession No. | Primers |
|---|---|---|
| GAPDH | NM_001206359.1 | F: 5′-GTCGGAGTGAACGGATTTGG-3′ |
| R: 5′-AGTGGAGGTCAATGAAGGGG-3′ | ||
| GCLC | XM_003482164.4 | F: 5′-GAAAACCAGGCTCTCTGCAC-3′ |
| R: 5′-ATCGCTTCGTCTGGAAAGAA-3′ | ||
| GSS | NM_001244625.1 | F: 5′-GGCTGAAGGACAGTGAGGAG-3′ |
| R: 5′-TTCCCTGCCTGACATAGACC-3′ | ||
| SLC6A20 | XM_021068640.1 | F: 5′-TGGTGGTGTCCTTCTTCCTC-3′ |
| R: 5′-ATTCAGTGGGCAGACAGACC-3′ | ||
| SLC36A1 | XM_021077073.1 | F: 5′-CATCGGCATCTTCTTCACCT-3′ |
| R: 5′-GGTCTATCACCAGCCTCCAA-3′ | ||
| SLC36A2 | XM_021077082.1 | F: 5′-TTGCTAGCCATGGGCTTCAT-3′ |
| R: 5′-AGAAGCTCACGATACGCCTTC-3′ | ||
| SLC38A1 | XM_003355629.4 | F: 5′-GAACACTGGAGCAATGCTGA-3′ |
| R: 5′-ATAGCCGAGATAGCCCAGGT-3′ | ||
| SLC38A2 | XM_005659640.3 | F: 5′-CCTACCTCCTGACAGCTTGC-3′ |
| R: 5′-AACCAAAAGCACCAAACAGG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, N.; Yang, Y.; Si, X.; Jia, H.; Zhang, Y.; Jiang, D.; Dai, Z.; Wu, Z. L-Proline Activates Mammalian Target of Rapamycin Complex 1 and Modulates Redox Environment in Porcine Trophectoderm Cells. Biomolecules 2021, 11, 742. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11050742
Liu N, Yang Y, Si X, Jia H, Zhang Y, Jiang D, Dai Z, Wu Z. L-Proline Activates Mammalian Target of Rapamycin Complex 1 and Modulates Redox Environment in Porcine Trophectoderm Cells. Biomolecules. 2021; 11(5):742. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11050742
Chicago/Turabian StyleLiu, Ning, Ying Yang, Xuemeng Si, Hai Jia, Yunchang Zhang, Da Jiang, Zhaolai Dai, and Zhenlong Wu. 2021. "L-Proline Activates Mammalian Target of Rapamycin Complex 1 and Modulates Redox Environment in Porcine Trophectoderm Cells" Biomolecules 11, no. 5: 742. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11050742