
Lack of Conserved miRNA Deregulation in HPV-Induced Squamous Cell Carcinomas

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Samples
2.2. Sample Processing for NGS
2.3. Analysis of NGS Data
3. Results and Discussion
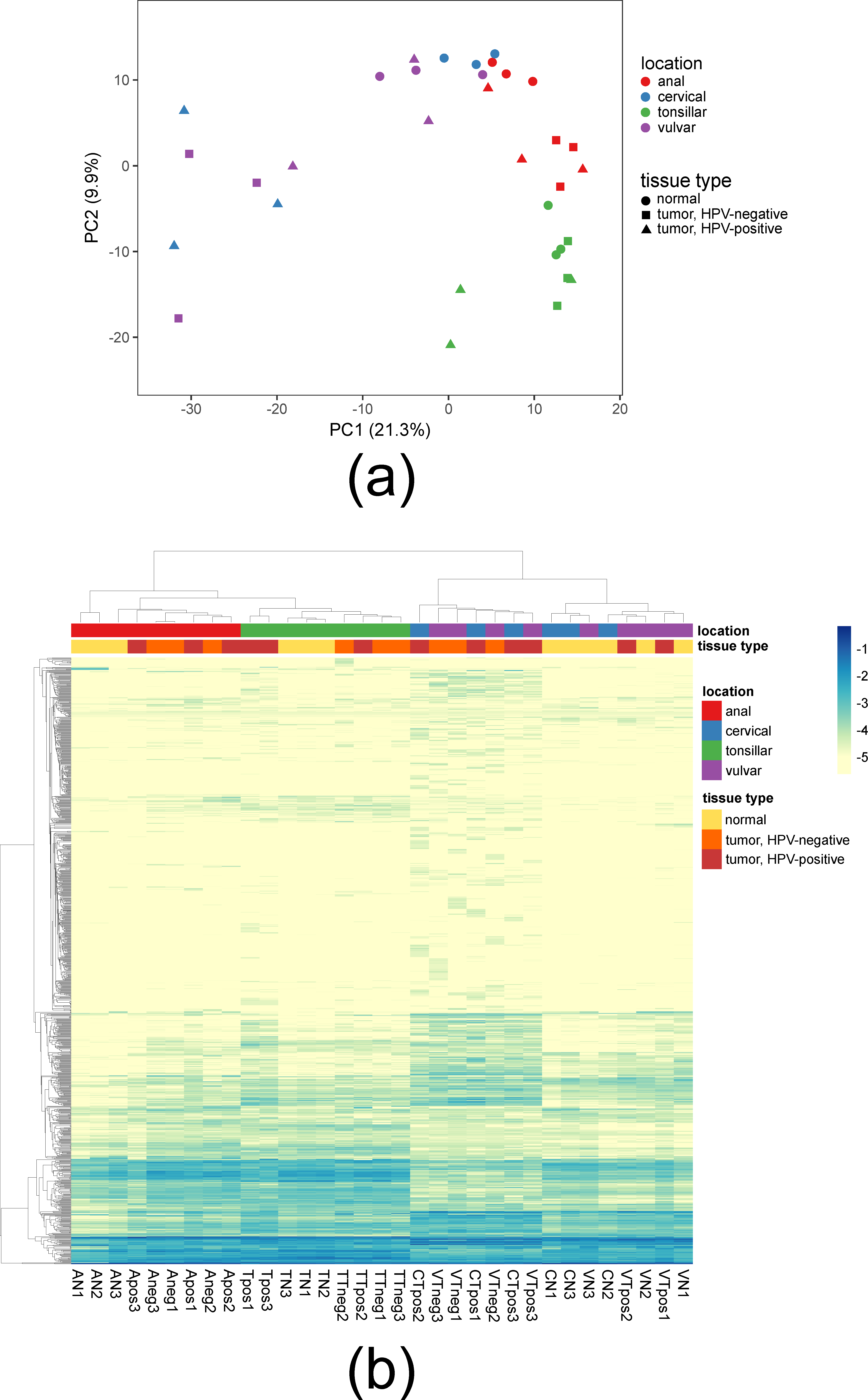
3.1. Global Characterization of miRNA Expression
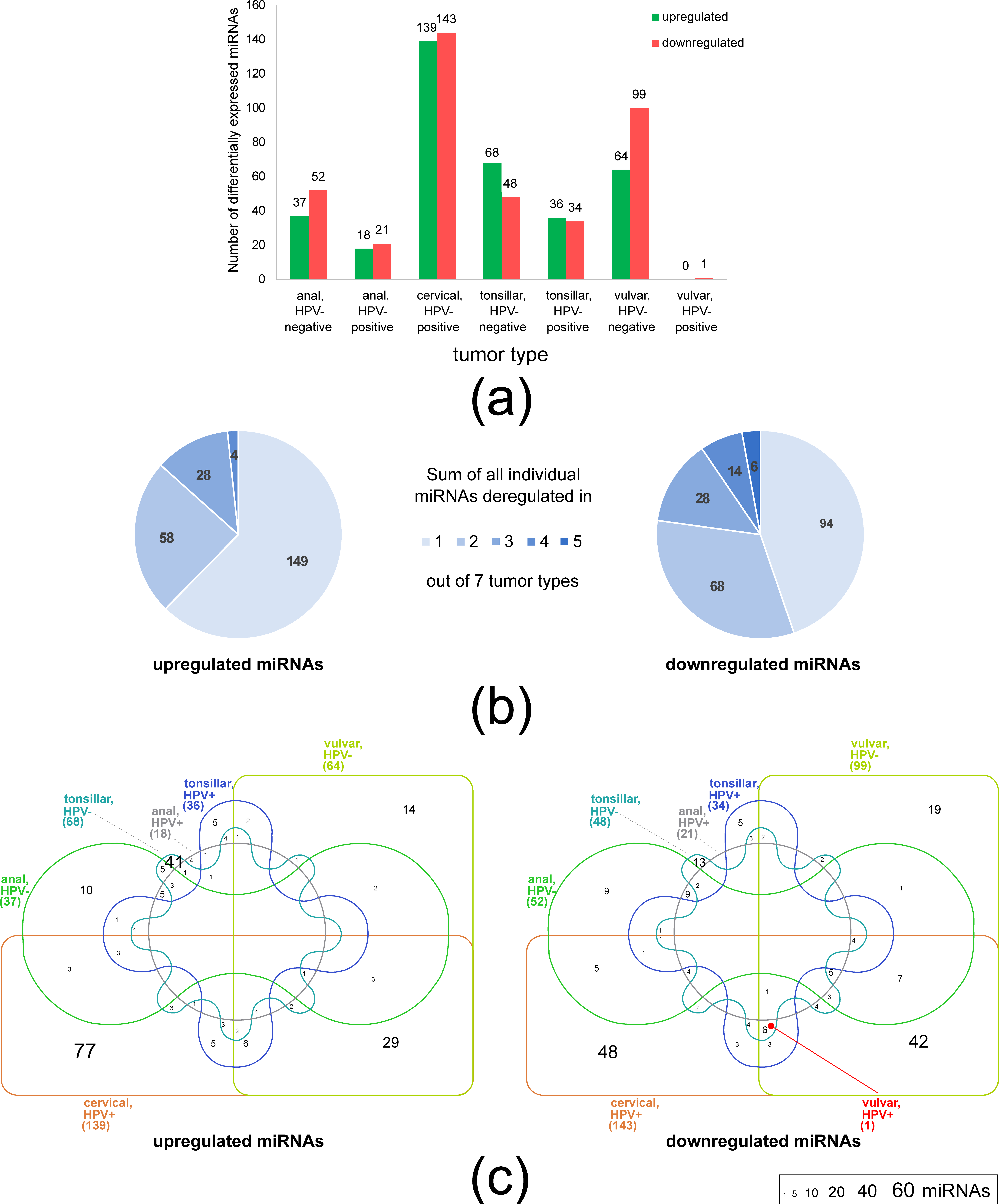
3.2. Differential Expression of miRNA in SCCs

{kind=link}
{kind=link}
| Gene | miRNA | Downregulated in (Location/HPV Status) 1 | Targets in SCCs | References |
|---|---|---|---|---|
| hsa-miR-101-3p | MIR101-1 | anal/−, cervical/+, tonsillar/−, tonsillar/+, vulvar/− | CDK8, COX-2, CXCR7, EZH2, FOS, JAK2, MALAT1, TGFBR1, ZEB1 | [42,43,44,45,46,47,48,49,50,51,52] |
| hsa-miR-10b-5p | MIR10B | anal/−, cervical/+, tonsillar/−, tonsillar/+, vulvar/− | HOXA1, IGF1R, TIAM1 | [53,54,55] |
| hsa-miR-29c-3p | MIR29C | anal/−, cervical/+, tonsillar/−, tonsillar/+, vulvar/− | CCNE, ITGA6, LAMC2 | [56,57] |
| hsa-miR-30a-5p | MIR30A | anal/−, cervical/+, tonsillar/−, tonsillar/+, vulvar/− | FOXD1, FZD2, MEF2D, WNT2 | [58,59,60] |
| hsa-miR-451a | MIR451A | cervical/+, tonsillar/−, tonsillar/+, vulvar/−, vulvar/+ | CDKN2D, ESDN, KIF2A, MAP3K1, PDPK1 | [61,62,63,64] |
| hsa-miR-195-5p | MIR195 | anal/+, cervical/+, tonsillar/−, tonsillar/+, vulvar/− | ARL2, BCL2, CCND1, CCND2, CDC42, DCUN1D1, HDGF, MYB, ROCK1, SMAD3, SMAD7, TRIM14, VEGF, YAP1 | [65,66,67,68,69,70,71,72,73,74,75,76,77,78] |

3.3. Limited Role of HPV in miRNA Deregulation
| Gene | miRNA | Cervical Cancer | Vulvar HPV-Negative Cancer | Function in SCCs | SCC Type 1 | Targets | References | ||
|---|---|---|---|---|---|---|---|---|---|
| FC of Expression | padj | FC of Expression | padj | ||||||
| hsa-miR-103b | MIR103B1 | 16.8 | 1.2 × 10−4 | 11.3 | 4.7 × 10−3 | Tumor suppressor | OSCC | SALL4 | [89] |
| hsa-miR-107 | MIR107 | 15.8 | 8.6 × 10−3 | 12.9 | 1.5 × 10−2 | Tumor suppressor | CC | MCL1 | [90] |
| hsa-miR-125b-5p | MIR125B1 | 6.4 | 1.8 × 10−3 | 14.6 | 3.7 × 10−7 | Tumor suppressor | CC | PIK3CD | [91] |
| hsa-miR-136-3p | MIR136 | 31.6 | 1.0 × 10−7 | 6.5 | 1.1 × 10−2 | Tumor suppressor | CC | E2F1 | [92] |
| hsa-miR-148a-3p | MIR148A | 22.4 | 3.2 × 10−8 | 13.3 | 4.7 × 10−5 | Tumor suppressor | ESCC, OSCC | MAP3K9, IGF1R | [93,94] |
| hsa-miR-148b-3p | MIR148B | 10.4 | 6.7 × 10−3 | 10.4 | 3.2 × 10−2 | Tumor suppressor | CC | CASP3 | [95] |
| hsa-miR-152-3p | MIR152 | 11.8 | 8.8 × 10−5 | 3.7 | 6.8 × 10−2 | Tumor suppressor | CC | KLF5 | [96] |
| hsa-miR-16-5p | MIR16-1 | 7.2 | 3.8 × 10-3 | 10.8 | 1.8 × 10−4 | Tumor suppressor | OSCC | AKT3, BCL2L2 | [97] |
| hsa-miR-17-5p | MIR17 | 22.3 | 6.4 × 10−3 | 11.0 | 3.7 × 10−2 | Tumor suppressor | CC | TP53INP1 | [98] |
| hsa-miR-199a-3p | MIR199A1 | 38.2 | 4.4 × 10−9 | 10.7 | 9.3 × 10−4 | Tumor suppressor | HNSCC | ITGA3 | [99] |
| hsa-miR-199a-5p | MIR199A1 | 4.8 | 2.0 × 10−3 | 4.5 | 1.2 × 10−2 | Tumor suppressor | HNSCC, OSCC | SOX4, IKK2, ITGA3 | [99,100,101] |
| hsa-miR-199b-3p | MIR199B | 34.1 | 7.8 × 10−9 | 10.6 | 9.3 × 10−4 | Tumor suppressor | HNSCC | ITGA3 | [99] |
| hsa-miR-199b-5p | MIR199B | 13.1 | 3.2 × 10−8 | 9.3 | 3.2 × 10−4 | Tumor suppressor | HNSCC | ITGA3 | [99] |
| hsa-miR-211-5p | MIR211 | 15.6 | 3.0 × 10−2 | 58.2 | 2.4 × 10−5 | Tumor suppressor | CC | SPARC | [102] |
| hsa-miR-218-5p | MIR218-1 | 11.4 | 1.2 × 10−3 | 4.3 | 9.3 × 10−2 | Tumor suppressor | CC | IDO1 | [103] |
| hsa-miR-222-3p | MIR222 | 5.7 | 7.8 × 10−4 | 3.8 | 2.0 × 10−2 | Tumor suppressor | CC | ALDH1 | [104] |
| hsa-miR-23a-3p | MIR23A | 3.7 | 1.9 × 10−2 | 3.2 | 6.8 × 10−2 | Tumor suppressor | OSCC | FGF2 | [105] |
| hsa-miR-24-3p | MIR24-1 | 7.5 | 3.0 × 10−4 | 9.9 | 9.3 × 10−4 | Tumor suppressor | LSCC | S100A8, XIAP | [106,107] |
| hsa-miR-27a-3p | MIR27A | 4.0 | 6.0 × 10−3 | 3.2 | 4.5 × 10−2 | Tumor suppressor | CC | TGFBRI | [108] |
| hsa-miR-27b-3p | MIR27B | 5.5 | 2.9 × 10−3 | 6.8 | 1.3 × 10−3 | Tumor suppressor | ESCC | NFE2L2 | [109] |
| hsa-miR-30d-5p | MIR30D | 2.4 | 7.7 × 10−2 | 2.7 | 7.6 × 10−2 | Tumor suppressor | ESCC | EZH2 | [110] |
| hsa-miR-33a-5p | MIR33A | 6.0 | 3.6 × 10−2 | 12.2 | 2.8 × 10−2 | Tumor suppressor | CC | TWIST1 | [111] |
| hsa-miR-376c-3p | MIR376C | 45.4 | 9.0 × 10−6 | 12.8 | 1.8 × 10−2 | Tumor suppressor | CC | BMI1 | [112] |
| hsa-miR-411-5p | MIR411 | 67.3 | 4.1 × 10−5 | 8.9 | 6.6 × 10−2 | Tumor suppressor | CC | STAT3 | [113] |
| hsa-miR-99a-3p | MIR99A | 12.5 | 6.7 × 10−4 | 26.9 | 1.1 × 10−3 | Tumor suppressor | CC | TRIB2 | [114] |
| hsa-miR-99a-5p | MIR99A | 4.2 | 4.4 × 10−2 | 17.7 | 3.7 × 10−7 | Tumor suppressor | CC | TRIB2 | [114] |
| hsa-let-7a-5p | MIRLET7A1 | 9.5 | 2.5 × 10−4 | 9.0 | 9.3 × 10−4 | Tumor suppressor | CC | PKM2, TGFBR1 | [115,116] |
| hsa-let-7b-5p | MIRLET7B | 4.3 | 2.4 × 10−3 | 2.7 | 7.7 × 10−2 | Tumor suppressor | CC | KIAA1377 | [117] |
| hsa-let-7c-5p | MIRLET7C | 14.0 | 4.0 × 10−5 | 11.8 | 2.4 × 10−5 | Tumor suppressor | ESCC, HNSCC | CTHRC1, IGF1R, HMGA2 | [118,119] |
| hsa-let-7e-5p | MIRLET7E | 19.3 | 3.4 × 10−4 | 5.8 | 3.7 × 10−2 | Tumor suppressor | HNSCC | CCR7 | [120] |
| hsa-miR-106b-5p | MIR106B | 6.2 | 3.8 × 10−2 | 8.4 | 3.0 × 10−2 | oncogene | CC | DAB2 | [121] |
| hsa-miR-130a-3p | MIR130A | 37.4 | 1.6 × 10−8 | 8.4 | 6.9 × 10−3 | oncogene | CC | RUNX3 | [122] |
| hsa-miR-141-3p | MIR141 | 3.7 | 3.8 × 10−2 | 3.1 | 6.9 × 10−2 | oncogene | CC | FOXA2 | [123] |
| hsa-miR-4454 | MIR4454 | 7.8 | 3.3 × 10−4 | 5.2 | 2.6 × 10−2 | oncogene | CC | ABHD2, NUDT21 | [124] |
| hsa-miR-191-5p | MIR191 | 8.0 | 5.2 × 10−5 | 5.8 | 2.8 × 10−3 | n.d. | |||
| hsa-miR-3074-5p | MIR3074 | 7.5 | 3.0 × 10−4 | 9.9 | 9.3 × 10−4 | n.d. | |||
| hsa-miR-3195 | MIR3195 | 3.0 | 9.2 × 10−2 | 3.6 | 5.7 × 10−2 | n.d. | |||
| hsa-miR-4286 | MIR4286 | 5.2 | 5.1 × 10−2 | 6.0 | 4.1 × 10−2 | n.d. | |||
| hsa-miR-6510-3p | MIR6510 | 10.7 | 4.3 × 10−2 | 13.4 | 1.9 × 10−2 | n.d. | |||
| hsa-miR-660-5p | MIR660 | 20.0 | 5.7 × 10−4 | 7.1 | 3.4 × 10−2 | n.d. | |||
| hsa-miR-887-3p | MIR887 | 8.8 | 1.7 × 10−3 | 4.2 | 3.5 × 10−2 | n.d. | |||
| hsa-let-7f-5p | MIRLET7F1 | 7.4 | 2.8 × 10−4 | 5.0 | 3.1 × 10−3 | n.d. | |||
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Plummer, M.; de Martel, C.; Vignat, J.; Ferlay, J.; Bray, F.; Franceschi, S. Global burden of cancers attributable to infections in 2012: A synthetic analysis. Lancet Glob. Health 2016, 4, e609–e616. [Google Scholar] [CrossRef] [Green Version]
- de Martel, C.; Plummer, M.; Vignat, J.; Franceschi, S. Worldwide burden of cancer attributable to HPV by site, country and HPV type. Int. J. Cancer 2017, 141, 664–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordfors, C.; Vlastos, A.; Du, J.; Ahrlund-Richter, A.; Tertipis, N.; Grün, N.; Romanitan, M.; Haeggblom, L.; Roosaar, A.; Dahllöf, G.; et al. Human papillomavirus prevalence is high in oral samples of patients with tonsillar and base of tongue cancer. Oral Oncol. 2014, 50, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Marur, S.; D’Souza, G.; Westra, W.H.; Forastiere, A.A. HPV-associated head and neck cancer: A virus-related cancer epidemic. Lancet Oncol. 2010, 11, 781–789. [Google Scholar] [CrossRef] [Green Version]
- Näsman, A.; Attner, P.; Hammarstedt, L.; Du, J.; Eriksson, M.; Giraud, G.; Ahrlund-Richter, S.; Marklund, L.; Romanitan, M.; Lindquist, D.; et al. Incidence of human papillomavirus (HPV) positive tonsillar carcinoma in Stockholm, Sweden: An epidemic of viral-induced carcinoma? Int. J. Cancer 2009, 125, 362–366. [Google Scholar] [CrossRef]
- Tachezy, R.; Klozar, J.; Rubenstein, L.; Smith, E.; Saláková, M.; Smahelová, J.; Ludvíková, V.; Rotnáglová, E.; Kodet, R.; Hamsíková, E. Demographic and risk factors in patients with head and neck tumors. J. Med. Virol. 2009, 81, 878–887. [Google Scholar] [CrossRef]
- Koncar, R.F.; Feldman, R.; Bahassi, E.M.; Hashemi Sadraei, N. Comparative molecular profiling of HPV-induced squamous cell carcinomas. Cancer Med. 2017, 6, 1673–1685. [Google Scholar] [CrossRef] [PubMed]
- Tuna, M.; Amos, C.I. Next generation sequencing and its applications in HPV-associated cancers. Oncotarget 2017, 8, 8877–8889. [Google Scholar] [CrossRef] [Green Version]
- Gillison, M.L.; Akagi, K.; Xiao, W.; Jiang, B.; Pickard, R.K.L.; Li, J.; Swanson, B.J.; Agrawal, A.D.; Zucker, M.; Stache-Crain, B.; et al. Human papillomavirus and the landscape of secondary genetic alterations in oral cancers. Genome Res. 2019, 29, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Haeggblom, L.; Ährlund-Richter, A.; Mirzaie, L.; Farrajota Neves da Silva, P.; Ursu, R.G.; Ramqvist, T.; Näsman, A. Differences in gene expression between high-grade dysplasia and invasive HPV. Cancer Med. 2019, 8, 6221–6232. [Google Scholar] [CrossRef] [Green Version]
- Seiwert, T.Y.; Zuo, Z.; Keck, M.K.; Khattri, A.; Pedamallu, C.S.; Stricker, T.; Brown, C.; Pugh, T.J.; Stojanov, P.; Cho, J.; et al. Integrative and comparative genomic analysis of HPV-positive and HPV-negative head and neck squamous cell carcinomas. Clin. Cancer Res. 2015, 21, 632–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, M.R.; Shin, S.; Park, H.C.; Kim, M.S.; Lee, S.H.; Jung, S.H.; Song, S.Y.; Chung, Y.J. Mutational signatures and chromosome alteration profiles of squamous cell carcinomas of the vulva. Exp. Mol. Med. 2018, 50, e442. [Google Scholar] [CrossRef] [PubMed]
- Prieske, K.; Alawi, M.; Oliveira-Ferrer, L.; Jaeger, A.; Eylmann, K.; Burandt, E.; Schmalfeldt, B.; Joosse, S.A.; Woelber, L. Genomic characterization of vulvar squamous cell carcinoma. Gynecol. Oncol. 2020, 158, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [Green Version]
- Ferracin, M.; Pedriali, M.; Veronese, A.; Zagatti, B.; Gafà, R.; Magri, E.; Lunardi, M.; Munerato, G.; Querzoli, G.; Maestri, I.; et al. MicroRNA profiling for the identification of cancers with unknown primary tissue-of-origin. J. Pathol. 2011, 225, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Iorio, M.V.; Ferracin, M.; Liu, C.G.; Veronese, A.; Spizzo, R.; Sabbioni, S.; Magri, E.; Pedriali, M.; Fabbri, M.; Campiglio, M.; et al. MicroRNA gene expression deregulation in human breast cancer. Cancer Res. 2005, 65, 7065–7070. [Google Scholar] [CrossRef] [Green Version]
- Porkka, K.P.; Pfeiffer, M.J.; Waltering, K.K.; Vessella, R.L.; Tammela, T.L.; Visakorpi, T. MicroRNA expression profiling in prostate cancer. Cancer Res. 2007, 67, 6130–6135. [Google Scholar] [CrossRef] [Green Version]
- Yanaihara, N.; Caplen, N.; Bowman, E.; Seike, M.; Kumamoto, K.; Yi, M.; Stephens, R.M.; Okamoto, A.; Yokota, J.; Tanaka, T.; et al. Unique microRNA molecular profiles in lung cancer diagnosis and prognosis. Cancer Cell 2006, 9, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Lajer, C.B.; Garnæs, E.; Friis-Hansen, L.; Norrild, B.; Therkildsen, M.H.; Glud, M.; Rossing, M.; Lajer, H.; Svane, D.; Skotte, L.; et al. The role of miRNAs in human papilloma virus (HPV)-associated cancers: Bridging between HPV-related head and neck cancer and cervical cancer. Br. J. Cancer 2012, 106, 1526–1534. [Google Scholar] [CrossRef]
- Pereira, P.M.; Marques, J.P.; Soares, A.R.; Carreto, L.; Santos, M.A. MicroRNA expression variability in human cervical tissues. PLoS ONE 2010, 5, e11780. [Google Scholar] [CrossRef] [Green Version]
- Rao, Q.; Shen, Q.; Zhou, H.; Peng, Y.; Li, J.; Lin, Z. Aberrant microRNA expression in human cervical carcinomas. Med. Oncol. 2012, 29, 1242–1248. [Google Scholar] [CrossRef] [PubMed]
- Snoek, B.C.; Verlaat, W.; Babion, I.; Novianti, P.W.; van de Wiel, M.A.; Wilting, S.M.; van Trommel, N.E.; Bleeker, M.C.G.; Massuger, L.F.A.G.; Melchers, W.J.G.; et al. Genome-wide microRNA analysis of HPV-positive self-samples yields novel triage markers for early detection of cervical cancer. Int. J. Cancer 2019, 144, 372–379. [Google Scholar] [CrossRef] [PubMed]
- de Melo Maia, B.; Lavorato-Rocha, A.M.; Rodrigues, L.S.; Coutinho-Camillo, C.M.; Baiocchi, G.; Stiepcich, M.M.; Puga, R.; de A Lima, L.; Soares, F.A.; Rocha, R.M. MicroRNA portraits in human vulvar carcinoma. Cancer Prev. Res. (Phila) 2013, 6, 1231–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Wu, X. MiRNA expression profile of vulvar squamous cell carcinoma and identification of the oncogenic role of miR-590-5p. Oncol. Rep. 2016, 35, 398–408. [Google Scholar] [CrossRef] [Green Version]
- Avissar, M.; Christensen, B.C.; Kelsey, K.T.; Marsit, C.J. MicroRNA expression ratio is predictive of head and neck squamous cell carcinoma. Clin. Cancer Res. 2009, 15, 2850–2855. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.S.; Jiang, W.W.; Smith, I.; Poeta, L.M.; Begum, S.; Glazer, C.; Shan, S.; Westra, W.; Sidransky, D.; Califano, J.A. MicroRNA alterations in head and neck squamous cell carcinoma. Int. J. Cancer 2008, 123, 2791–2797. [Google Scholar] [CrossRef] [Green Version]
- Childs, G.; Fazzari, M.; Kung, G.; Kawachi, N.; Brandwein-Gensler, M.; McLemore, M.; Chen, Q.; Burk, R.D.; Smith, R.V.; Prystowsky, M.B.; et al. Low-level expression of microRNAs let-7d and miR-205 are prognostic markers of head and neck squamous cell carcinoma. Am. J. Pathol. 2009, 174, 736–745. [Google Scholar] [CrossRef] [Green Version]
- Hui, A.B.; Lenarduzzi, M.; Krushel, T.; Waldron, L.; Pintilie, M.; Shi, W.; Perez-Ordonez, B.; Jurisica, I.; O’Sullivan, B.; Waldron, J.; et al. Comprehensive MicroRNA profiling for head and neck squamous cell carcinomas. Clin. Cancer Res. 2010, 16, 1129–1139. [Google Scholar] [CrossRef] [Green Version]
- Ramdas, L.; Giri, U.; Ashorn, C.L.; Coombes, K.R.; El-Naggar, A.; Ang, K.K.; Story, M.D. MiRNA expression profiles in head and neck squamous cell carcinoma and adjacent normal tissue. Head Neck 2009, 31, 642–654. [Google Scholar] [CrossRef]
- Lajer, C.B.; Nielsen, F.C.; Friis-Hansen, L.; Norrild, B.; Borup, R.; Garnæs, E.; Rossing, M.; Specht, L.; Therkildsen, M.H.; Nauntofte, B.; et al. Different miRNA signatures of oral and pharyngeal squamous cell carcinomas: A prospective translational study. Br. J. Cancer 2011, 104, 830–840. [Google Scholar] [CrossRef]
- Miller, D.L.; Davis, J.W.; Taylor, K.H.; Johnson, J.; Shi, Z.; Williams, R.; Atasoy, U.; Lewis, J.S.; Stack, M.S. Identification of a human papillomavirus-associated oncogenic miRNA panel in human oropharyngeal squamous cell carcinoma validated by bioinformatics analysis of the Cancer Genome Atlas. Am. J. Pathol. 2015, 185, 679–692. [Google Scholar] [CrossRef] [Green Version]
- Vojtechova, Z.; Sabol, I.; Salakova, M.; Smahelova, J.; Zavadil, J.; Turek, L.; Grega, M.; Klozar, J.; Prochazka, B.; Tachezy, R. Comparison of the miRNA profiles in HPV-positive and HPV-negative tonsillar tumors and a model system of human keratinocyte clones. BMC Cancer 2016, 16, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotnáglová, E.; Tachezy, R.; Saláková, M.; Procházka, B.; Košl’abová, E.; Veselá, E.; Ludvíková, V.; Hamšíková, E.; Klozar, J. HPV involvement in tonsillar cancer: Prognostic significance and clinically relevant markers. Int. J. Cancer 2011, 129, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.; Simpson, G.; Solymos, P. Vegan: Community Ecology Package. R Package Version 2.5–6. 2019. Available online: https://github.com/vegandevs/vegan (accessed on 1 January 2021).
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef]
- Ludwig, N.; Leidinger, P.; Becker, K.; Backes, C.; Fehlmann, T.; Pallasch, C.; Rheinheimer, S.; Meder, B.; Stähler, C.; Meese, E.; et al. Distribution of miRNA expression across human tissues. Nucleic Acids Res. 2016, 44, 3865–3877. [Google Scholar] [CrossRef] [PubMed]
- Panwar, B.; Omenn, G.S.; Guan, Y. miRmine: A database of human miRNA expression profiles. Bioinformatics 2017, 33, 1554–1560. [Google Scholar] [CrossRef] [Green Version]
- Volinia, S.; Calin, G.A.; Liu, C.G.; Ambs, S.; Cimmino, A.; Petrocca, F.; Visone, R.; Iorio, M.; Roldo, C.; Ferracin, M.; et al. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc. Natl. Acad. Sci. USA 2006, 103, 2257–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Gong, J.; Chu, Y.; Xu, M.; Huo, J.; Lv, L. Esophageal squamous cell carcinoma cell proliferation induced by exposure to low concentration of cigarette smoke extract is mediated via targeting miR-101-3p/COX-2 pathway. Oncol. Rep. 2016, 35, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Lin, C.; Shi, Y.H.; Kuerban, G. MicroRNA-101 inhibits cell proliferation, invasion, and promotes apoptosis by regulating cyclooxygenase-2 in Hela cervical carcinoma cells. Asian Pac. J. Cancer Prev. 2013, 14, 5915–5920. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Tian, L.; Ren, H.; Chen, X.; Wang, Y.; Ge, J.; Wu, S.; Sun, Y.; Liu, M.; Xiao, H. MicroRNA-101 is a potential prognostic indicator of laryngeal squamous cell carcinoma and modulates CDK8. J. Transl. Med. 2015, 13, 271. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Jia, J.; Zang, Y.; Li, J.; Wan, B. MicroRNA-101 regulates autophagy, proliferation and apoptosis via targeting EZH2 in laryngeal squamous cell carcinoma. Neoplasma 2019, 66, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Huang, F.; Shen, G.; Yiming, A. MicroRNA-101 regulates the viability and invasion of cervical cancer cells. Int. J. Clin. Exp. Pathol. 2015, 8, 10148–10155. [Google Scholar] [PubMed]
- Wei, H.; He, W.R.; Chen, K.M.; Wang, X.W.; Yi, C.J. MiR-101 affects proliferation and apoptosis of cervical cancer cells by inhibition of JAK2. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5640–5647. [Google Scholar] [CrossRef]
- Liang, X.; Liu, Y.; Zeng, L.; Yu, C.; Hu, Z.; Zhou, Q.; Yang, Z. MiR-101 inhibits the G1-to-S phase transition of cervical cancer cells by targeting Fos. Int. J. Gynecol. Cancer 2014, 24, 1165–1172. [Google Scholar] [CrossRef]
- Hui, Y.; Li, Y.; Jing, Y.; Feng, J.Q.; Ding, Y. MiRNA-101 acts as a tumor suppressor in oral squamous cell carcinoma by targeting CX chemokine receptor 7. Am. J. Transl. Res. 2016, 8, 4902–4911. [Google Scholar] [PubMed]
- Wu, B.; Lei, D.; Wang, L.; Yang, X.; Jia, S.; Yang, Z.; Shan, C.; Zhang, C.; Lu, B. MiRNA-101 inhibits oral squamous-cell carcinoma growth and metastasis by targeting zinc finger E-box binding homeobox 1. Am. J. Cancer Res. 2016, 6, 1396–1407. [Google Scholar]
- Wang, Y.; Jia, R.Z.; Diao, S.; He, J.; Jia, L. MiRNA-101 targets TGF-βR1 to retard the progression of oral squamous cell carcinoma. Oncol. Res. 2020, 28, 203–212. [Google Scholar] [CrossRef]
- Wang, X.; Li, M.; Wang, Z.; Han, S.; Tang, X.; Ge, Y.; Zhou, L.; Zhou, C.; Yuan, Q.; Yang, M. Silencing of long noncoding RNA MALAT1 by miR-101 and miR-217 inhibits proliferation, migration, and invasion of esophageal squamous cell carcinoma cells. J. Biol. Chem. 2015, 290, 3925–3935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, D.; Zhou, Q.; Wang, D.; Guan, L.; Yuan, L.; Li, S. The downregulation of microRNA-10b and its role in cervical cancer. Oncol. Res. 2016, 24, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Hou, R.; Wang, D.; Lu, J. MicroRNA-10b inhibits proliferation, migration and invasion in cervical cancer cells via direct targeting of insulin-like growth factor-1 receptor. Oncol. Lett. 2017, 13, 5009–5015. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Xu, Y.; Pan, L.; Feng, Y.; Luo, K.; Mu, Q.; Luo, G. MiR-10b downregulated by DNA methylation acts as a tumor suppressor in HPV-positive cervical cancer via targeting Tiam1. Cell Physiol. BioChem. 2018, 51, 1763–1777. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, T.; Nohata, N.; Hanazawa, T.; Kikkawa, N.; Yamamoto, N.; Yoshino, H.; Itesako, T.; Enokida, H.; Nakagawa, M.; Okamoto, Y.; et al. Tumour-suppressive microRNA-29s inhibit cancer cell migration and invasion by targeting laminin-integrin signalling in head and neck squamous cell carcinoma. Br. J. Cancer 2013, 109, 2636–2645. [Google Scholar] [CrossRef] [Green Version]
- Ding, D.P.; Chen, Z.L.; Zhao, X.H.; Wang, J.W.; Sun, J.; Wang, Z.; Tan, F.W.; Tan, X.G.; Li, B.Z.; Zhou, F.; et al. MiR-29c induces cell cycle arrest in esophageal squamous cell carcinoma by modulating cyclin E expression. Carcinogenesis 2011, 32, 1025–1032. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Tang, J.; Xu, S.; Zhang, W.; Jiang, H. MiR-30a-5p inhibits proliferation and migration of lung squamous cell carcinoma cells by targeting FOXD1. Biomed. Res. Int. 2020, 2020, 2547902. [Google Scholar] [CrossRef] [Green Version]
- Qi, B.; Wang, Y.; Chen, Z.J.; Li, X.N.; Qi, Y.; Yang, Y.; Cui, G.H.; Guo, H.Z.; Li, W.H.; Zhao, S. Down-regulation of miR-30a-3p/5p promotes esophageal squamous cell carcinoma cell proliferation by activating the Wnt signaling pathway. World J. Gastroenterol. 2017, 23, 7965–7977. [Google Scholar] [CrossRef]
- Zhao, J.; Li, B.; Shu, C.; Ma, Y.; Gong, Y. Downregulation of miR-30a is associated with proliferation and invasion via targeting MEF2D in cervical cancer. Oncol. Lett. 2017, 14, 7437–7442. [Google Scholar] [CrossRef] [Green Version]
- Fukumoto, I.; Kinoshita, T.; Hanazawa, T.; Kikkawa, N.; Chiyomaru, T.; Enokida, H.; Yamamoto, N.; Goto, Y.; Nishikawa, R.; Nakagawa, M.; et al. Identification of tumour suppressive microRNA-451a in hypopharyngeal squamous cell carcinoma based on microRNA expression signature. Br. J. Cancer 2014, 111, 386–394. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Zhao, J.; Zhang, H.; Fan, X.; Geng, W.; Qiao, S. MicroRNA-451a prevents cutaneous squamous cell carcinoma progression via the 3-phosphoinositide-dependent protein kinase-1-mediated PI3K/AKT signaling pathway. Exp. Ther. Med. 2021, 21, 116. [Google Scholar] [CrossRef] [PubMed]
- Zang, W.Q.; Yang, X.; Wang, T.; Wang, Y.Y.; Du, Y.W.; Chen, X.N.; Li, M.; Zhao, G.Q. MiR-451 inhibits proliferation of esophageal carcinoma cell line EC9706 by targeting CDKN2D and MAP3K1. World J. Gastroenterol. 2015, 21, 5867–5876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchida, A.; Seki, N.; Mizuno, K.; Yamada, Y.; Misono, S.; Sanada, H.; Kikkawa, N.; Kumamoto, T.; Suetsugu, T.; Inoue, H. Regulation of KIF2A by antitumor miR-451a inhibits cancer cell aggressiveness features in lung squamous cell carcinoma. Cancers 2019, 11, 258. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Liu, J.; Wang, L.; Yang, X.; Liu, X. MicroRNA-195 inhibits cell proliferation, migration and invasion in laryngeal squamous cell carcinoma by targeting ROCK1. Mol. Med. Rep. 2017, 16, 7154–7162. [Google Scholar] [CrossRef] [PubMed]
- Shuang, Y.; Li, C.; Zhou, X.; Huang, Y.; Zhang, L. MicroRNA-195 inhibits growth and invasion of laryngeal carcinoma cells by directly targeting DCUN1D1. Oncol. Rep. 2017, 38, 2155–2165. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wei, H.; Yin, D.; Lu, Y.; Zhang, Y.; Zhang, Q.; Ma, X.; Zhang, S. MicroRNA-195 inhibits proliferation of cervical cancer cells by targeting cyclin D1a. Tumor Biol. 2016, 37, 4711–4720. [Google Scholar] [CrossRef]
- Song, R.; Cong, L.; Ni, G.; Chen, M.; Sun, H.; Sun, Y. MicroRNA-195 inhibits the behavior of cervical cancer tumors by directly targeting HDGF. Oncol. Lett. 2017, 14, 767–775. [Google Scholar] [CrossRef] [Green Version]
- DU, X.; Lin, L.I.; Zhang, L.; Jiang, J. MicroRNA-195 inhibits the proliferation, migration and invasion of cervical cancer cells via the inhibition of CCND2 and MYB expression. Oncol. Lett. 2015, 10, 2639–2643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, X.; Jia, Q.; Yuan, J.; Shi, X.; Guo, H.; Gao, J.; Guo, Y. MicroRNA-195 suppresses cell proliferation, migration and invasion in epithelial ovarian carcinoma via inhibition of the CDC42/CCND1 pathway. Int. J. Mol. Med. 2020, 46, 1862–1872. [Google Scholar] [CrossRef]
- Sun, N.; Ye, L.; Chang, T.; Li, X. MicroRNA-195-Cdc42 axis acts as a prognostic factor of esophageal squamous cell carcinoma. Int. J. Clin. Exp. Pathol. 2014, 7, 6871–6879. [Google Scholar]
- Liu, W.; Wang, J.; Meng, N.; Wang, X.; Ge, D. MiR-195 inhibits proliferation of oral squamous cell carcinoma cells through regulating Smad7. Panminerva Med. 2020. [Google Scholar] [CrossRef]
- Zhou, Q.; Han, L.R.; Zhou, Y.X.; Li, Y. MiR-195 suppresses cervical cancer migration and invasion through targeting Smad3. Int. J. Gynecol. Cancer 2016, 26, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Chen, Y.; Li, Y.; Li, C.; Qin, T.; Bai, M.; Zhang, Z.; Jia, R.; Su, Y.; Wang, C. MiR-195 suppresses metastasis and angiogenesis of squamous cell lung cancer by inhibiting the expression of VEGF. Mol. Med. Rep. 2019, 20, 2625–2632. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, Y.; Ning, Y.E.; Gu, H.; Tong, Y.; Wang, N. MiR-195-5p inhibits malignant progression of cervical cancer by targeting YAP1. Onco Targets Ther. 2020, 13, 931–944. [Google Scholar] [CrossRef] [Green Version]
- Pan, S.S.; Zhou, H.E.; Yu, H.Y.; Xu, L.H. MiR-195-5p inhibits the cell migration and invasion of cervical carcinoma through suppressing ARL2. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 10664–10671. [Google Scholar] [CrossRef]
- Wang, T.; Ren, Y.; Liu, R.; Ma, J.; Shi, Y.; Zhang, L.; Bu, R. MiR-195-5p suppresses the proliferation, migration, and invasion of oral squamous cell carcinoma by targeting TRIM14. Biomed. Res. Int. 2017, 2017, 7378148. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.F.; Wei, S.B.; Gong, K.; Gan, Y.H.; Yu, G.Y. Prognostic implications of micoRNA miR-195 expression in human tongue squamous cell carcinoma. PLoS ONE 2013, 8, e56634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- Iacona, J.R.; Lutz, C.S. miR-146a-5p: Expression, regulation, and functions in cancer. Wiley Interdiscip. Rev. RNA 2019, 10, e1533. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Song, J.; Ding, B.; Cui, Y.; Liang, J.; Han, S. MiR-146a promotes cervical cancer cell viability via targeting IRAK1 and TRAF6. Oncol. Rep. 2018, 39, 3015–3024. [Google Scholar] [CrossRef] [Green Version]
- Gee, H.E.; Camps, C.; Buffa, F.M.; Patiar, S.; Winter, S.C.; Betts, G.; Homer, J.; Corbridge, R.; Cox, G.; West, C.M.; et al. hsa-mir-210 is a marker of tumor hypoxia and a prognostic factor in head and neck cancer. Cancer 2010, 116, 2148–2158. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Guo, H.; Yin, S.; Du, H. miR-139-5p functions as a tumor suppressor in cervical cancer by targeting TCF4 and inhibiting Wnt/β-catenin signaling. Onco Targets Ther. 2019, 12, 7739–7748. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Jin, J.; Ma, T.; Zhai, H. MiR-139-5p inhibits the tumorigenesis and progression of oral squamous carcinoma cells by targeting HOXA9. J. Cell Mol. Med. 2017, 21, 3730–3740. [Google Scholar] [CrossRef]
- Rasmussen, C.L.; Sand, F.L.; Hoffmann Frederiksen, M.; Kaae Andersen, K.; Kjaer, S.K. Does HPV status influence survival after vulvar cancer? Int. J. Cancer 2018, 142, 1158–1165. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhang, Y.; Zhang, Z. Prevalence of human papillomavirus and its prognostic value in vulvar cancer: A systematic review and meta-analysis. PLoS ONE 2018, 13, e0204162. [Google Scholar] [CrossRef] [PubMed]
- Hinten, F.; Molijn, A.; Eckhardt, L.; Massuger, L.F.A.G.; Quint, W.; Bult, P.; Bulten, J.; Melchers, W.J.G.; de Hullu, J.A. Vulvar cancer: Two pathways with different localization and prognosis. Gynecol. Oncol. 2018, 149, 310–317. [Google Scholar] [CrossRef]
- Williams, A.; Syed, S.; Velangi, S.; Ganesan, R. New directions in vulvar cancer pathology. Curr. Oncol. Rep. 2019, 21, 88. [Google Scholar] [CrossRef]
- Liu, X.; Cao, Y.; Zhang, Y.; Zhou, H.; Li, H. Regulatory effect of MiR103 on proliferation, EMT and invasion of oral squamous carcinoma cell through SALL4. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 9931–9938. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Li, G.; Zhou, J.; Han, N.; Liu, Z.; Yin, J. MiR-107 activates ATR/Chk1 pathway and suppress cervical cancer invasion by targeting MCL1. PLoS ONE 2014, 9, e111860. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Li, X.; Zhu, X.; Huang, L.; Huang, Y.; Mao, C.; Yan, Q.; Zhu, J.; Zhao, W.; Shi, H. MiR-125b inhibits tumor growth and promotes apoptosis of cervical cancer cells by targeting phosphoinositide 3-kinase catalytic subunit delta. Cell Physiol. BioChem. 2012, 30, 1310–1318. [Google Scholar] [CrossRef]
- Lu, H.J.; Jin, P.Y.; Tang, Y.; Fan, S.H.; Zhang, Z.F.; Wang, F.; Wu, D.M.; Lu, J.; Zheng, Y.L. MicroRNA-136 inhibits proliferation and promotes apoptosis and radiosensitivity of cervical carcinoma through the NF-κB pathway by targeting E2F1. Life Sci. 2018, 199, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.X.; Yu, T.; Yu, Z.; Yang, X.G. MicroRNA-148a regulates the MAPK/ERK signaling pathway and suppresses the development of esophagus squamous cell carcinoma via targeting MAP3K9. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 6497–6504. [Google Scholar] [CrossRef] [PubMed]
- Jia, T.; Ren, Y.; Wang, F.; Zhao, R.; Qiao, B.; Xing, L.; Ou, L.; Guo, B. MiR-148a inhibits oral squamous cell carcinoma progression through ERK/MAPK pathway via targeting IGF-IR. BioSci. Rep. 2020, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mou, Z.; Xu, X.; Dong, M.; Xu, J. MicroRNA-148b acts as a tumor suppressor in cervical cancer by inducing G1/S-phase cell cycle arrest and apoptosis in a caspase-3-dependent manner. Med. Sci. Monit. 2016, 22, 2809–2815. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Lu, Y.; Wang, S.; Sheng, X.; Zhang, S. MicroRNA-152 acts as a tumor suppressor microRNA by inhibiting Krüppel-like factor 5 in human cervical cancer. Oncol. Res. 2019, 27, 335–340. [Google Scholar] [CrossRef]
- Wang, X.; Li, G.H. MicroRNA-16 functions as a tumor-suppressor gene in oral squamous cell carcinoma by targeting AKT3 and BCL2L2. J. Cell Physiol. 2018, 233, 9447–9457. [Google Scholar] [CrossRef] [Green Version]
- Wei, Q.; Li, Y.X.; Liu, M.; Li, X.; Tang, H. MiR-17-5p targets TP53INP1 and regulates cell proliferation and apoptosis of cervical cancer cells. IUBMB Life 2012, 64, 697–704. [Google Scholar] [CrossRef] [Green Version]
- Koshizuka, K.; Hanazawa, T.; Kikkawa, N.; Arai, T.; Okato, A.; Kurozumi, A.; Kato, M.; Katada, K.; Okamoto, Y.; Seki, N. Regulation of ITGA3 by the anti-tumor miR-199 family inhibits cancer cell migration and invasion in head and neck cancer. Cancer Sci. 2017, 108, 1681–1692. [Google Scholar] [CrossRef] [Green Version]
- Wei, D.; Shen, B.; Wang, W.; Zhou, Y.; Yang, X.; Lu, G.; Yang, J.; Shao, Y. MicroRNA-199a-5p functions as a tumor suppressor in oral squamous cell carcinoma via targeting the IKKβ/NF-κB signaling pathway. Int. J. Mol. Med. 2019, 43, 1585–1596. [Google Scholar] [CrossRef] [Green Version]
- Wei, D.; Wang, W.; Shen, B.; Zhou, Y.; Yang, X.; Lu, G.; Yang, J.; Shao, Y. MicroRNA-199a-5p suppresses migration and invasion in oral squamous cell carcinoma through inhibiting the EMT-related transcription factor SOX4. Int. J. Mol. Med. 2019, 44, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Qu, X.; Gao, D.; Ren, Q.; Jiang, X.; Bai, J.; Sheng, L. MiR-211 inhibits proliferation, invasion and migration of cervical cancer via targeting SPARC. Oncol. Lett. 2018, 16, 853–860. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Tu, H.; Liang, Y.; Tang, D. MiR-218 produces anti-tumor effects on cervical cancer cells in vitro. World J. Surg. Oncol. 2018, 16, 204. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhang, Y.; Liang, S.; Ying, Y. Aldehyde dehydrogenase 1, a target of miR-222, is expressed at elevated levels in cervical cancer. Exp. Ther. Med. 2020, 19, 1673–1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Qi, S.; Zhang, X.; Wu, J.; Yang, X.; Wang, R. MiR-23a-3p suppresses cell proliferation in oral squamous cell carcinomas by targeting FGF2 and correlates with a better prognosis: miR-23a-3p inhibits OSCC growth by targeting FGF2. Pathol. Res. Pract. 2019, 215, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Chen, Z.; Xue, F.; Chen, W.; Ma, R.; Cheng, S.; Cui, P. MicroRNA-24 inhibits growth, induces apoptosis, and reverses radioresistance in laryngeal squamous cell carcinoma by targeting X-linked inhibitor of apoptosis protein. Cancer Cell Int. 2015, 15, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Fu, W.; Chen, H.; Shang, C.; Zhong, M. MiR-24 functions as a tumor suppressor in Hep2 laryngeal carcinoma cells partly through down-regulation of the S100A8 protein. Oncol. Rep. 2012, 27, 1097–1103. [Google Scholar] [CrossRef] [Green Version]
- Fang, F.; Huang, B.; Sun, S.; Xiao, M.; Guo, J.; Yi, X.; Cai, J.; Wang, Z. MiR-27a inhibits cervical adenocarcinoma progression by downregulating the TGF-βRI signaling pathway. Cell Death Dis. 2018, 9, 395. [Google Scholar] [CrossRef]
- Han, M.; Li, N.; Li, F.; Wang, H.; Ma, L. MiR-27b-3p exerts tumor suppressor effects in esophageal squamous cell carcinoma by targeting Nrf2. Hum. Cell 2020, 33, 641–651. [Google Scholar] [CrossRef]
- Xie, R.; Wu, S.N.; Gao, C.C.; Yang, X.Z.; Wang, H.G.; Zhang, J.L.; Yan, W.; Ma, T.H. MicroRNA-30d inhibits the migration and invasion of human esophageal squamous cell carcinoma cells via the post-transcriptional regulation of enhancer of zeste homolog 2. Oncol. Rep. 2017, 37, 1682–1690. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Gui, Y.; Xie, P.; Li, G. MicroRNA-33a regulates the invasion of cervical cancer cells via targeting Twist1. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2015, 40, 1060–1067. [Google Scholar] [CrossRef]
- Deng, Y.; Xiong, Y.; Liu, Y. MiR-376c inhibits cervical cancer cell proliferation and invasion by targeting BMI1. Int. J. Exp. Pathol. 2016, 97, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Shan, D.; Shang, Y.; Hu, T. MicroRNA-411 inhibits cervical cancer progression by directly targeting STAT3. Oncol. Res. 2019, 27, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Xin, J.X.; Yue, Z.; Zhang, S.; Jiang, Z.H.; Wang, P.Y.; Li, Y.J.; Pang, M.; Xie, S.Y. MiR-99 inhibits cervical carcinoma cell proliferation by targeting TRIB2. Oncol. Lett. 2013, 6, 1025–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.; Chen, X.; Peng, R.; Liu, H.; Yin, P.; Peng, H.; Zhou, Y.; Sun, Y.; Wen, L.; Yi, H.; et al. Let-7a suppresses cell proliferation via the TGF-β/SMAD signaling pathway in cervical cancer. Oncol. Rep. 2016, 36, 3275–3282. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Zhao, X.; Yuan, X.; Jiang, J.; Li, P. MiR-let-7a inhibits cell proliferation, migration, and invasion by down-regulating PKM2 in cervical cancer. Oncotarget 2017, 8, 28226–28236. [Google Scholar] [CrossRef] [Green Version]
- Zheng, S.; Liu, Q.; Ma, R.; Tan, D.; Shen, T.; Zhang, X.; Lu, X. Let-7b-5p inhibits proliferation and motility in squamous cell carcinoma cells through negative modulation of KIAA1377. Cell Biol. Int. 2019, 43, 634–641. [Google Scholar] [CrossRef] [PubMed]
- Hou, B.; Ishinaga, H.; Midorikawa, K.; Nakamura, S.; Hiraku, Y.; Oikawa, S.; Ma, N.; Takeuchi, K.; Murata, M. Let-7c inhibits migration and epithelial-mesenchymal transition in head and neck squamous cell carcinoma by targeting IGF1R and HMGA2. Oncotarget 2018, 9, 8927–8940. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Luo, M.; Lü, M.; Zhou, T.; Liu, F.; Guo, X.; Zhang, J.; Kang, M. Let-7c-5p inhibits cell proliferation and migration and promotes apoptosis via the CTHRC1/AKT/ERK pathway in esophageal squamous cell carcinoma. Onco Targets Ther. 2020, 13, 11193–11209. [Google Scholar] [CrossRef]
- Wang, S.; Jin, S.; Liu, M.D.; Pang, P.; Wu, H.; Qi, Z.Z.; Liu, F.Y.; Sun, C.F. hsa-let-7e-5p inhibits the proliferation and metastasis of head and neck squamous cell carcinoma cells by targeting chemokine receptor 7. J. Cancer 2019, 10, 1941–1948. [Google Scholar] [CrossRef]
- Cheng, Y.; Guo, Y.; Zhang, Y.; You, K.; Li, Z.; Geng, L. MicroRNA-106b is involved in transforming growth factor β1-induced cell migration by targeting disabled homolog 2 in cervical carcinoma. J. Exp. Clin. Cancer Res. 2016, 35, 11. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Wang, X.; Liu, W. MicroRNA-130a-3p promotes the proliferation and inhibits the apoptosis of cervical cancer cells via negative regulation of RUNX3. Mol. Med. Rep. 2020, 22, 2990–3000. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Zhang, Z.; Du, M.Z.; Guan, Y.C.; Yao, J.N.; Yu, H.Y.; Wang, B.J.; Wang, X.L.; Wu, S.L.; Li, Z. MicroRNA-141-3p fosters the growth, invasion, and tumorigenesis of cervical cancer cells by targeting FOXA2. Arch. BioChem. Biophys. 2018, 657, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hu, H.; Luo, Z.; Liu, S.; Wu, W.; Zhu, M.; Wang, J.; Liu, Y.; Lu, Z. MiR-4454 up-regulated by HPV16 E6/E7 promotes invasion and migration by targeting ABHD2/NUDT21 in cervical cancer. BioSci. Rep. 2020, 40. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nunvar, J.; Pagacova, L.; Vojtechova, Z.; Azevedo, N.T.D.d.; Smahelova, J.; Salakova, M.; Tachezy, R. Lack of Conserved miRNA Deregulation in HPV-Induced Squamous Cell Carcinomas. Biomolecules 2021, 11, 764. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11050764
Nunvar J, Pagacova L, Vojtechova Z, Azevedo NTDd, Smahelova J, Salakova M, Tachezy R. Lack of Conserved miRNA Deregulation in HPV-Induced Squamous Cell Carcinomas. Biomolecules. 2021; 11(5):764. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11050764
Chicago/Turabian StyleNunvar, Jaroslav, Lucie Pagacova, Zuzana Vojtechova, Nayara Trevisan Doimo de Azevedo, Jana Smahelova, Martina Salakova, and Ruth Tachezy. 2021. "Lack of Conserved miRNA Deregulation in HPV-Induced Squamous Cell Carcinomas" Biomolecules 11, no. 5: 764. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11050764