
VEGF Maintains Maternal Vascular Space Homeostasis in the Mouse Placenta through Modulation of Trophoblast Giant Cell Functions

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Animals
2.1.1. Endometrium-Specific VEGF Overexpression in Mice during Pregnancy
2.1.2. Placenta-Targeted Expression of sFlt1 shRNA
2.2. Histology
2.3. Alkaline Phosphatase Histochemical Staining
2.4. Image Processing and Morphometric Analysis
2.5. Statistical Analysis
3. Results
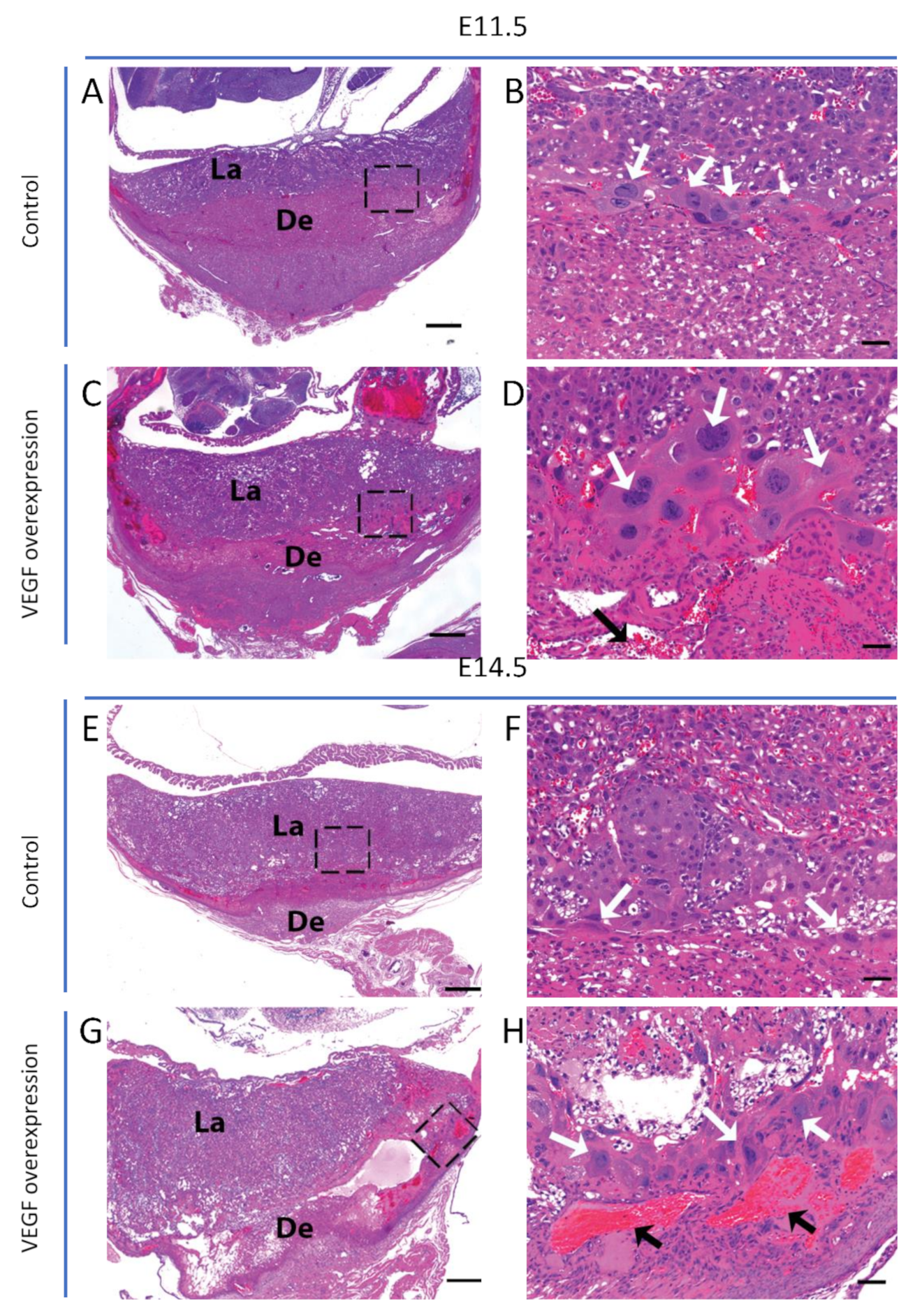
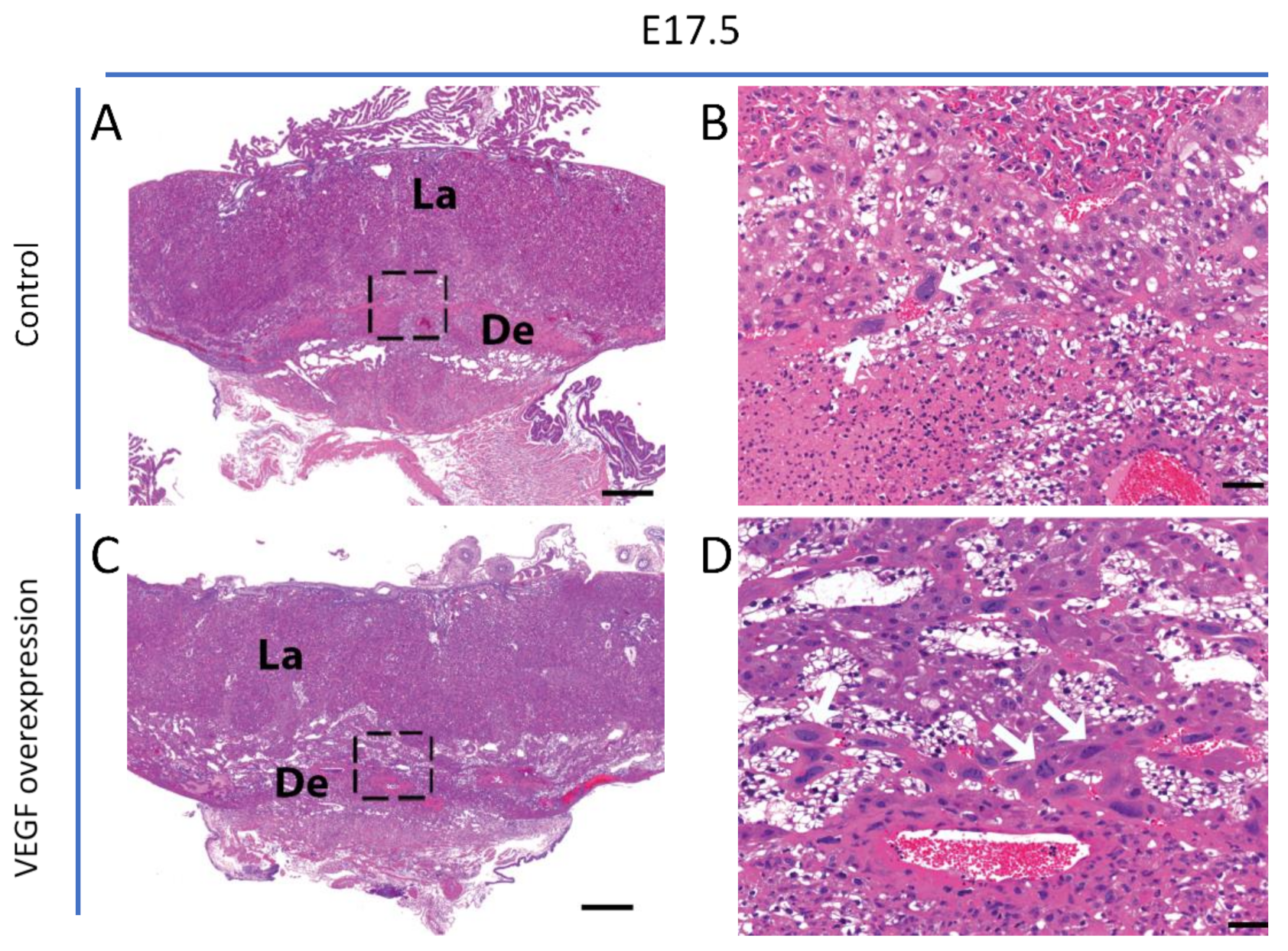
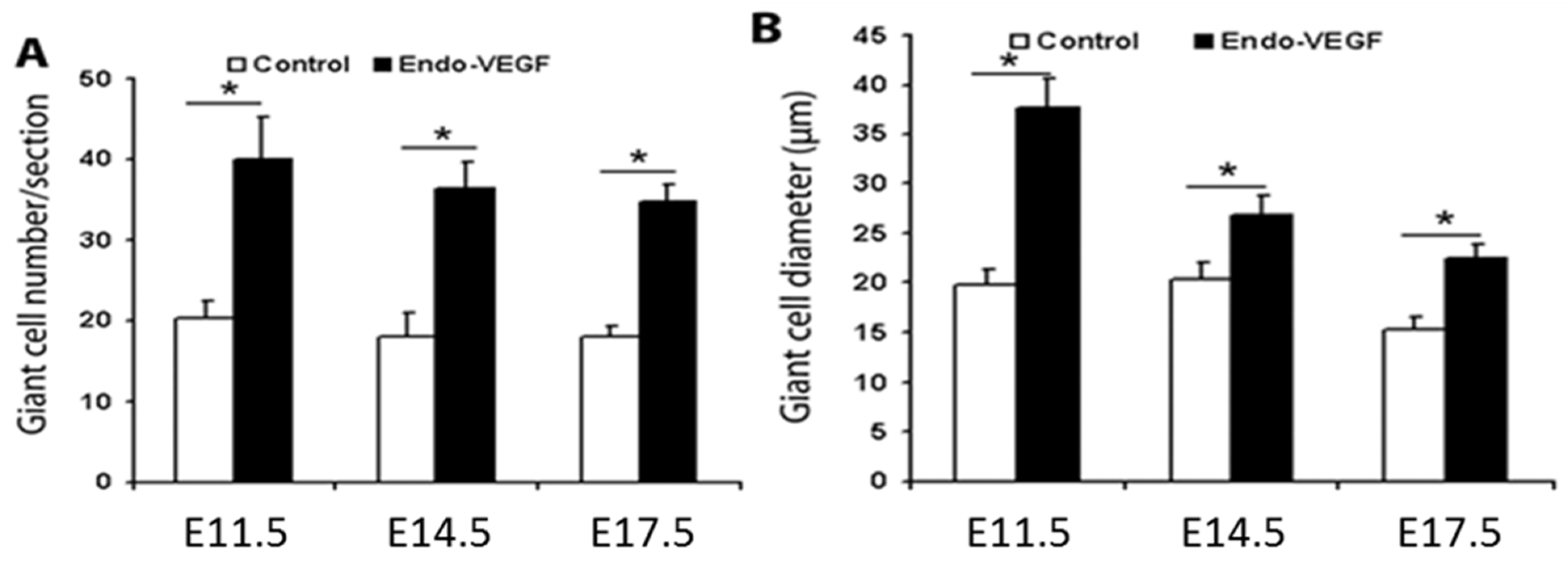
3.1. Endometrial VEGF Overexpression Leads to Dramatic Alteration of Parietal and Sinusoidal Trophoblast Giant Cell Differentiation and Abnormal Enlargement of Maternal Blood Spaces in the Placenta
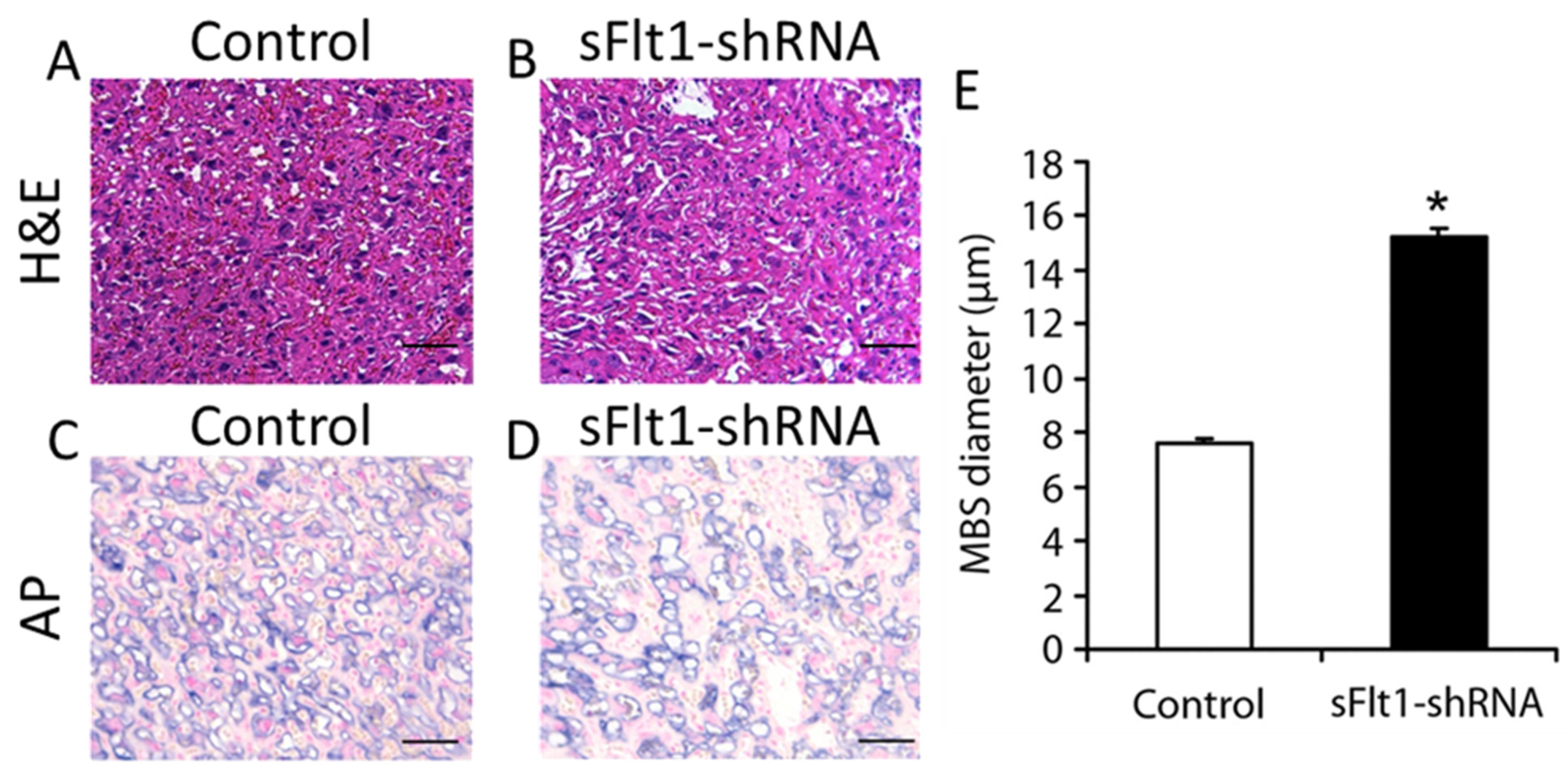
3.2. Placenta-Specific Knockdown of sFlt1 Preferentially Alters Sinusoidal Trophoblast Giant Cell Morphology and Number
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
Abbreviation
| BCIP | 5-Bromo-4-chloro-3-indolyl phosphate |
| Ch-TGCs | Channels TGCs |
| C-TGCs | Canal TGCs |
| E | Embryonic days |
| Flt1 | fms-like tyrosine kinase 1 |
| Fluc | Luciferase |
| GFP | Green Fluorescent Protein |
| H&E | Hematoxylin and Eosin |
| LV | Lentiviruses |
| NBT | Nitro blue tetrazolium |
| NT | Neutralize Tagment buffer |
| NTMT | Alkaline phosphatase buffer |
| P-TGCs | Parietal TGCs |
| sFlt1 | Soluble Flt1 |
| shRNA | A short hairpin RNA or small hairpin RNA |
| SpA-TGCs | Spiral artery-associated TGCs |
| TGCs | Trophoblast giant cells |
| VEGF | Vascular endothelial growth factor |
| VEGFR1 | Vascular endothelial growth factor 1 Receptor |
| VEGFR2 | Vascular endothelial growth factor 2 Receptor |
References
- Ferrara, N.; Carver-Moore, K.; Chen, H.; Dowd, M.; Lu, L.; O’Shea, K.S.; Powell-Braxton, L.; Hillan, K.J.; Moore, M.W. Heterozygous embryonic lethality induced by targeted inactivation of the VEGF gene. Nature 1996, 380, 439–442. [Google Scholar] [CrossRef] [Green Version]
- Carmeliet, P.; Ferreira, V.; Breier, G.; Pollefeyt, S.; Kieckens, L.; Gertsenstein, M.; Fahrig, M.; Vandenhoeck, A.; Harpal, K.; Eberhardt, C.; et al. Abnormal blood vessel development and lethality in embryos lacking a single VEGF allele. Nature 1996, 380, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Karaman, S.; Leppänen, V.M.; Alitalo, K. Vascular endothelial growth factor signaling in development and disease. Development 2018, 145, 151019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; McMaster, M.; Woo, K.; Janatpour, M.; Perry, J.; Karpanen, T.; Alitalo, K.; Damsky, C.; Fisher, S.J. Vascular endothelial growth factor ligands and receptors that regulate human cytotrophoblast survival are dysregulated in severe preeclampsia and hemolysis, elevated liver enzymes, and low platelets syndrome. Am. J. Pathol. 2002, 160, 1405–1423. [Google Scholar] [CrossRef] [Green Version]
- Rai, A.; Cross, J.C. Development of the hemochorial maternal vascular spaces in the placenta through endothelial and vasculogenic mimicry. Dev. Biol. 2014, 387, 131–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, D.G.; Fortier, A.L.; Cross, J.C. Diverse subtypes and developmental origins of trophoblast giant cells in the mouse placenta. Dev. Biol. 2007, 304, 567–578. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.; Cross, J.C. Development and function of trophoblast giant cells in the rodent placenta. Int. J. Dev. Biol. 2010, 54, 341–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jena, M.K.; Sharma, N.R.; Petitt, M.; Maulik, D.; Nayak, N.R. Pathogenesis of Preeclampsia and Therapeutic Approaches Targeting the Placenta. Biomolecules 2020, 10, 953. [Google Scholar] [CrossRef]
- Fan, X.; Rai, A.; Kambham, N.; Sung, J.F.; Singh, N.; Petitt, M.; Dhal, S.; Agrawal, R.; Sutton, R.E.; Druzin, M.L.; et al. Endometrial VEGF induces placental sFLT1 and leads to pregnancy complications. J. Clin. Investig. 2014, 124, 4941–4952. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Smith, S.K.; Day, K.A.; Clark, D.E.; Licence, D.R.; Charnock-Jones, D.S. Alternative splicing of vascular endothelial growth factor (VEGF)-R1 (FLT-1) pre-mRNA is important for the regulation of VEGF activity. Mol. Endocrinol. 1999, 13, 537–545. [Google Scholar] [CrossRef]
- Ferrara, N.; Gerber, H.P.; LeCouter, J. The biology of VEGF and its receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef]
- Furuya, M.; Kurasawa, K.; Nagahama, K.; Kawachi, K.; Nozawa, A.; Takahashi, T.; Aoki, I. Disrupted balance of angiogenic and antiangiogenic signalings in preeclampsia. J. Pregnancy 2011, 2011, 123717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, L.; Brkić, J.; Liu, M.; Fu, G.; Peng, C.; Wang, Y.L. Placental trophoblast cell differentiation: Physiological regulation and pathological relevance to preeclampsia. Mol. Aspects Med. 2013, 34, 981–1023. [Google Scholar] [CrossRef] [PubMed]
- Olsson, A.K.; Dimberg, A.; Kreuger, J.; Claesson-Welsh, L. VEGF receptor signalling—In control of vascular function. Nat. Rev. Mol. Cell Biol. 2006, 7, 359–371. [Google Scholar] [CrossRef]
- Drake, C.J.; LaRue, A.; Ferrara, N.; Little, C.D. VEGF regulates cell behavior during vasculogenesis. Dev. Biol. 2000, 224, 178–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parasar, P.; Guru, N.; Nayak, N.R. Contribution of macrophages to fetomaternal immunological tolerance. Hum. Immunol. 2021, 82, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Dhal, S.; Wu, J.C.; Kuo, C.J.; Druzin, M.L.; Nayak, N.R. A Novel Method of Local Gene Delivery and Noninvasive Imaging of Transgene Expression in the Mouse Endometrium. Biol. Reprod. 2011, 85, 478. [Google Scholar] [CrossRef]
- Fan, X.; Petitt, M.; Gamboa, M.; Huang, M.; Dhal, S.; Druzin, M.L.; Wu, J.C.; Chen-Tsai, Y.; Nayak, N.R. Transient, inducible, placenta-specific gene expression in mice. Endocrinology 2012, 153, 5637–5644. [Google Scholar] [CrossRef] [Green Version]
- Natale, D.; Starovic, M.; Cross, J. Phenotypic analysis of the mouse placenta. Methods Mol. Med. 2006, 121, 275–293. [Google Scholar] [PubMed]
- Tanaka, S.; Kunath, T.; Hadjantonakis, A.K.; Nagy, A.; Rossant, J. Promotion of trophoblast stem cell proliferation by FGF4. Science 1998, 282, 2072–2075. [Google Scholar] [CrossRef]
- Oda, M.; Shiota, K.; Tanaka, S. Trophoblast stem cells. Methods Enzymol. 2006, 419, 387–400. [Google Scholar] [PubMed]
- Rai, A.; Cross, J.C. Three-dimensional cultures of trophoblast stem cells autonomously develop vascular-like spaces lined by trophoblast giant cells. Dev. Biol. 2015, 398, 110–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbert, S.P.; Stainier, D.Y. Molecular control of endothelial cell behaviour during blood vessel morphogenesis. Nat. Rev. Mol. Cell Biol. 2011, 12, 551–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mould, A.; Morgan, M.A.; Li, L.; Bikoff, E.K.; Robertson, E.J. Blimp1/Prdm1 governs terminal differentiation of endovascular trophoblast giant cells and defines multipotent progenitors in the developing placenta. Genes Dev. 2012, 26, 2063–2074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibuya, M. Vascular endothelial growth factor receptor-1 (VEGFR-1/Flt-1): A dual regulator for angiogenesis. Angiogenesis 2006, 9, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Jena, M.K.; Nayak, N.; Chen, K.; Nayak, N.R. Role of Macrophages in Pregnancy and Related Complications. Arch. Immunol. Ther. Exp. 2019, 67, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Hirashima, M.; Lu, Y.; Byers, L.; Rossant, J. Trophoblast expression of fms-like tyrosine kinase 1 is not required for the establishment of the maternal–fetal interface in the mouse placenta. Proc. Natl. Acad. Sci. USA 2003, 100, 15637–15642. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, X.; Muruganandan, S.; Shallie, P.D.; Dhal, S.; Petitt, M.; Nayak, N.R. VEGF Maintains Maternal Vascular Space Homeostasis in the Mouse Placenta through Modulation of Trophoblast Giant Cell Functions. Biomolecules 2021, 11, 1062. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11071062
Fan X, Muruganandan S, Shallie PD, Dhal S, Petitt M, Nayak NR. VEGF Maintains Maternal Vascular Space Homeostasis in the Mouse Placenta through Modulation of Trophoblast Giant Cell Functions. Biomolecules. 2021; 11(7):1062. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11071062
Chicago/Turabian StyleFan, Xiujun, Shanmugam Muruganandan, Philemon D Shallie, Sabita Dhal, Matthew Petitt, and Nihar R Nayak. 2021. "VEGF Maintains Maternal Vascular Space Homeostasis in the Mouse Placenta through Modulation of Trophoblast Giant Cell Functions" Biomolecules 11, no. 7: 1062. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11071062