
In Vitro Compression Model for Orthodontic Tooth Movement Modulates Human Periodontal Ligament Fibroblast Proliferation, Apoptosis and Cell Cycle

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Antibodies
2.2. Cell Culture and In Vitro Compressive Stimulation Model
2.3. Trypan Blue Staining—Quantification of Cell Numbers
2.4. Phalloidin/DAPI Staining
2.5. Combined Cell Proliferation and Apoptosis Assay
2.6. Isolation of RNA, Purification and cDNA Synthesis
2.7. Real-Time Quantitative PCR
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
3.1. Actin Filaments of hPDLFs Changed under Compressive Force

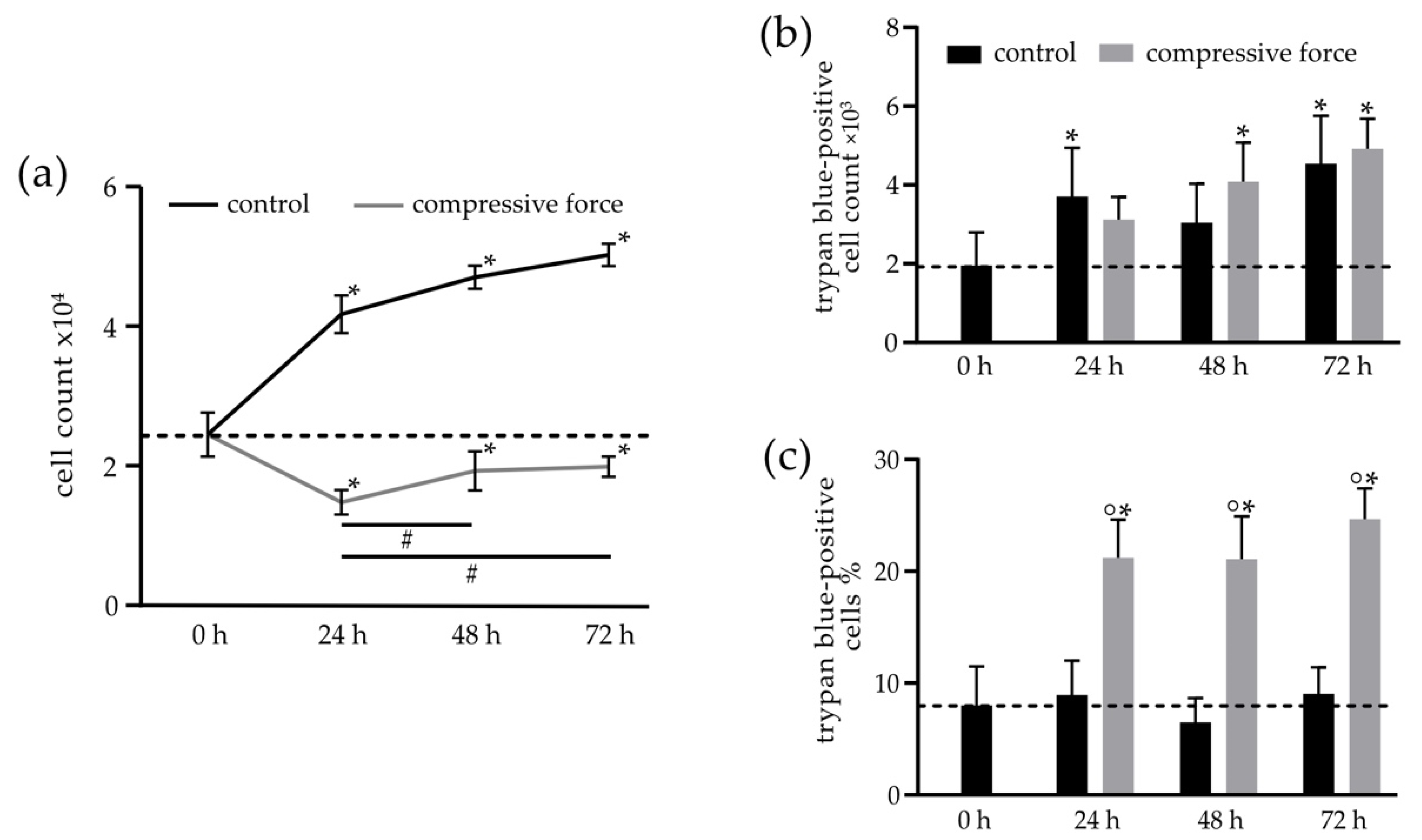
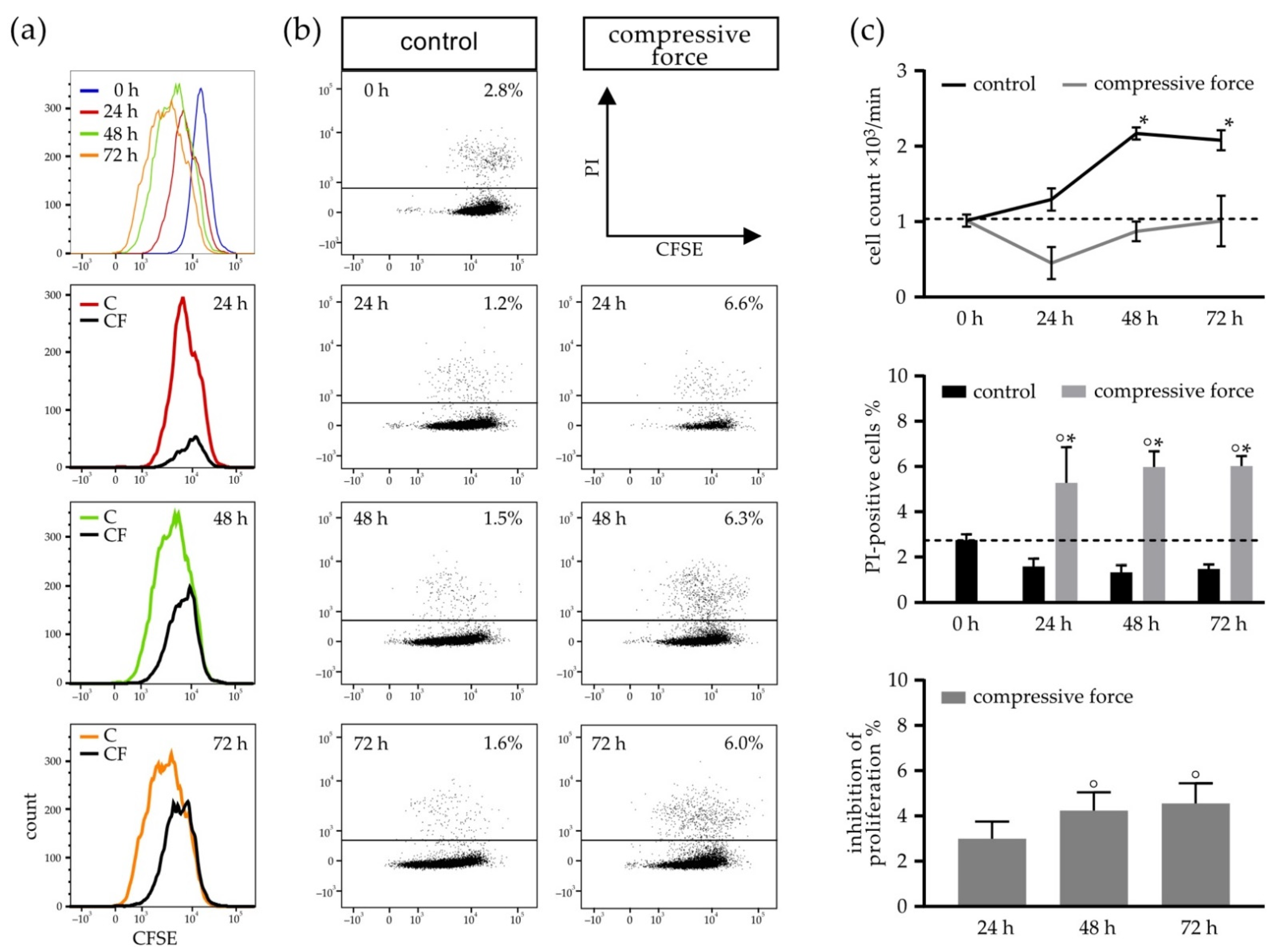
3.2. Mechanical Stimulation of hPDLF Affects Cell Number and Proliferation

3.3. Compressive Force Stimulated hPDLFs Showed Inhibition of Proliferation and Reduction of Viability

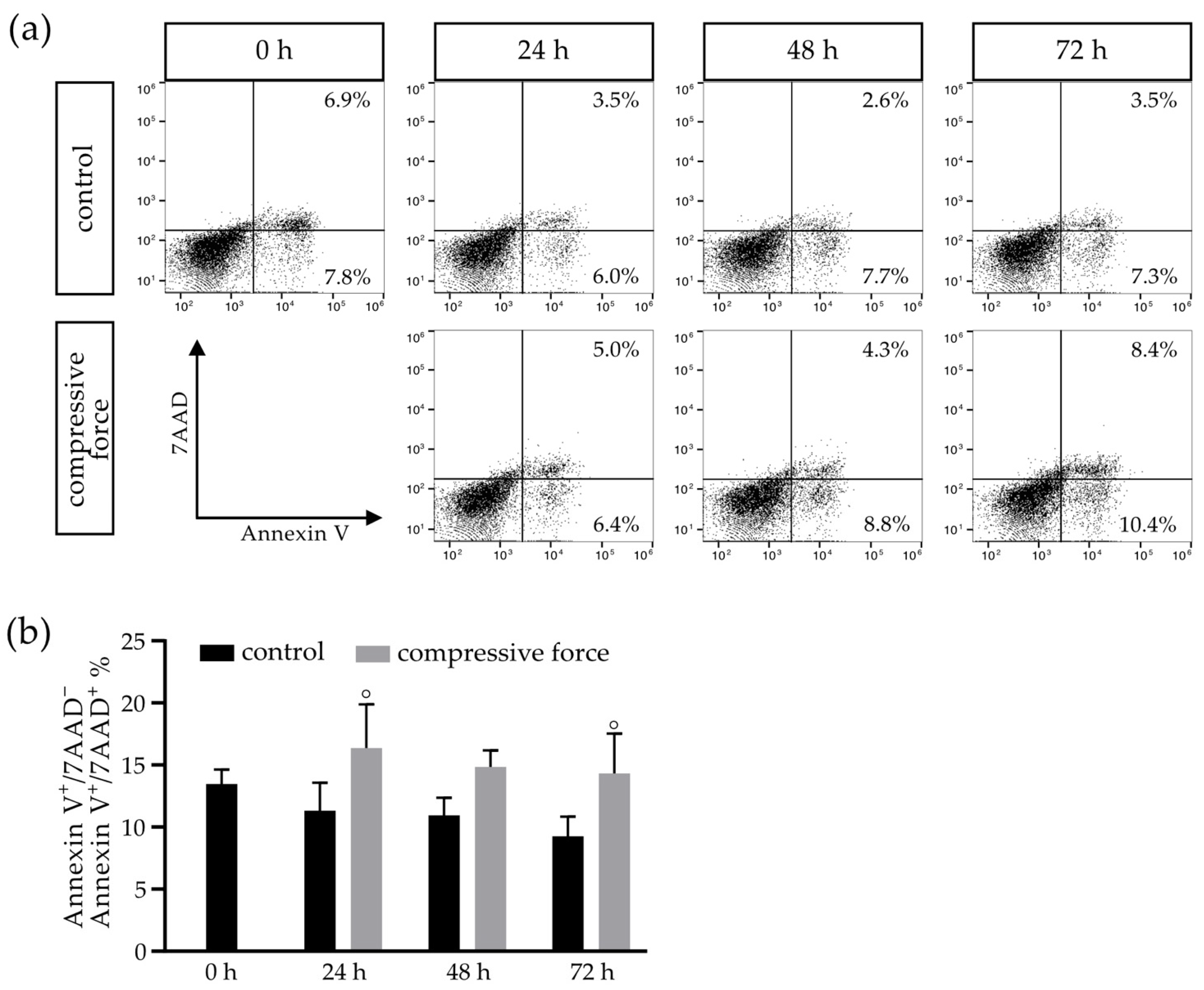
3.4. Compressive Force Affects hPDLFs in a Proapoptotic Manner

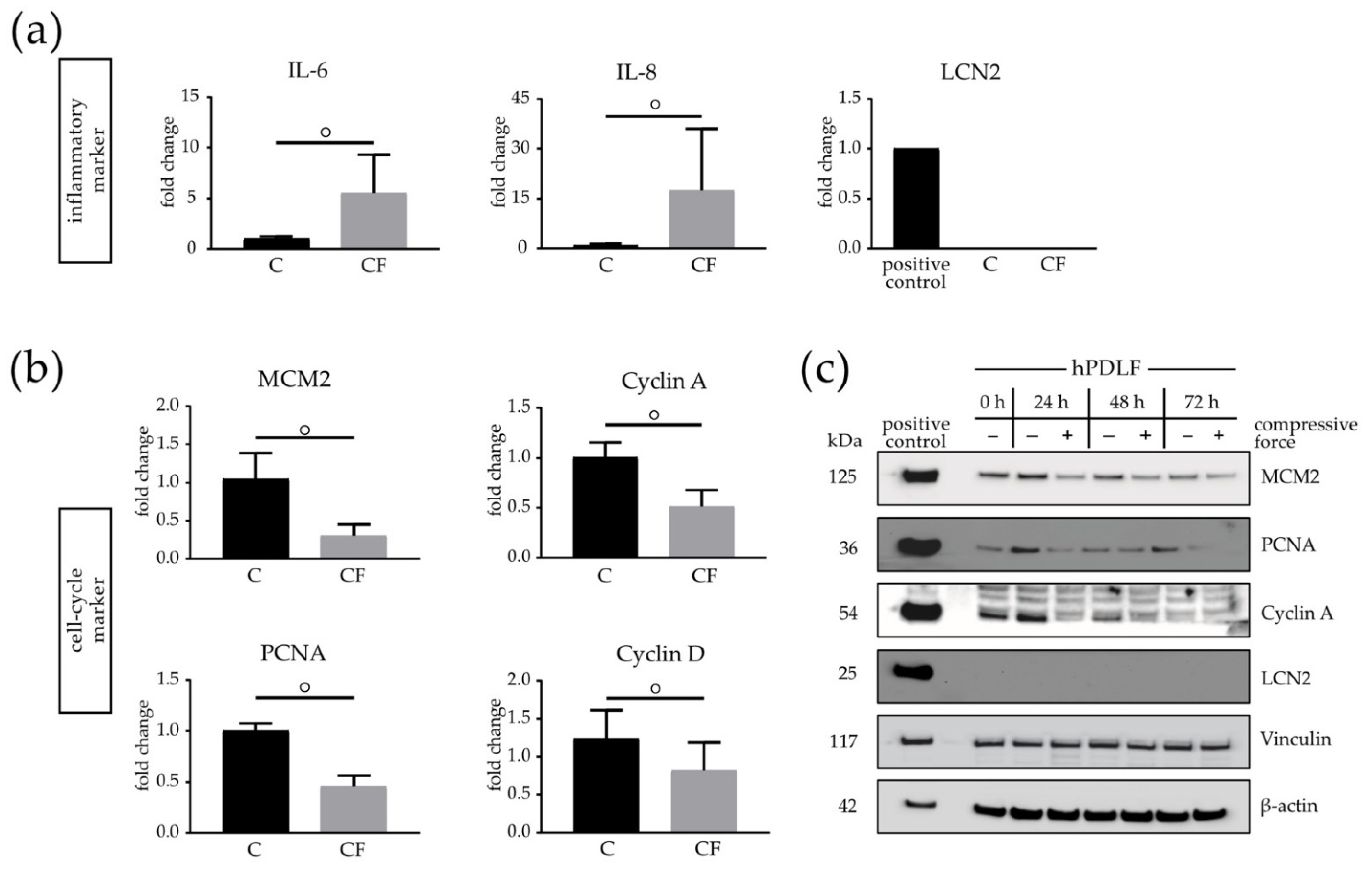
3.5. Cell Cycle and Restructuring Affected on mRNA and Protein Level in Mechanically Stimulated hPDLFs

4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nanci, A.; Bosshardt, D. Structure of periodontal tissues in health and disease. Periodontology 2000 2006, 40, 11–28. [Google Scholar] [CrossRef]
- Ho, S.P.; Marshall, S.J.; Ryder, M.I.; Marshall, G.W. The tooth attachment mechanism defined by structure, chemical composition and mechanical properties of collagen fibers in the periodontium. Biomaterials 2007, 28, 5238–5245. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; Hasegawa, T.; Yamamoto, T.; Hongo, H.; Amizuka, N. Histology of human cementum: Its structure, function, and development. Jpn. Dent. Sci. Rev. 2016, 52, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Linkous, E.R.; Trojan, T.M.; Harris, E.F. External apical root resorption and vectors of orthodontic tooth movement. Am. J. Orthod. Dentofac. Orthop. 2020, 158, 700–709. [Google Scholar] [CrossRef] [PubMed]
- Årtun, J.; Van’t Hullenaar, R.; Doppel, D.; Kuijpers-Jagtman, A.M. Identification of orthodontic patients at risk of severe apical root resorption. Am. J. Orthod. Dentofac. Orthop. 2009, 135, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Jäger, F.; Mah, J.K.; Bumann, A. Peridental bone changes after orthodontic tooth movement with fixed appliances: A cone-beam computed tomographic study. Angle Orthod. 2017, 87, 672–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meikle, M.C. The tissue, cellular, and molecular regulation of orthodontic tooth movement: 100 Years after Carl Sandstedt. Eur. J. Orthod. 2006, 28, 221–240. [Google Scholar] [CrossRef] [PubMed]
- Melsen, B. Biological reaction of alveolar bone to orthodontic tooth movement. Angle Orthod. 1999, 69, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, V.; Davidovitch, Z. Cellular, molecular, and tissue-level reactions to orthodontic force. Am. J. Orthod. Dentofac. Orthop. 2006, 129, 469.e1–469.e32. [Google Scholar] [CrossRef]
- McCulloch, C. Origins and functions of cells essential for periodontal repair: The role of fibroblasts in tissue homeostasis. Oral Dis. 1995, 1, 271–278. [Google Scholar] [CrossRef]
- Deschner, B.; Rath, B.; Jäger, A.; Deschner, J.; Denecke, B.; Memmert, S.; Götz, W. Gene analysis of signal transduction factors and transcription factors in periodontal ligament cells following application of dynamic strain. J. Orofac. Orthop. 2012, 73, 486–496. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, N.; Schröder, A.; Jantsch, J.; Spanier, G.; Proff, P.; Kirschneck, C. The role of mechanotransduction versus hypoxia during simulated orthodontic compressive strain—an in vitro study of human periodontal ligament fibroblasts. Int. J. Oral Sci. 2019, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hlaing, E.E.H.; Ishihara, Y.; Wang, Z.; Odagaki, N.; Kamioka, H. Role of intracellular Ca2+–based mechanotransduction of human periodontal ligament fibroblasts. FASEB J. 2019, 33, 10409–10424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madureira, D.F.; Taddei, S.D.A.; Abreu, M.H.N.G.; Pretti, H.; Lages, E.M.B.; Da Silva, T.A. Kinetics of interleukin-6 and chemokine ligands 2 and 3 expression of periodontal tissues during orthodontic tooth movement. Am. J. Orthod. Dentofac. Orthop. 2012, 142, 494–500. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Fukasawa, S. Is inflammation a friend or foe for orthodontic treatment?: Inflammation in orthodontically induced inflammatory root resorption and accelerating tooth movement. Int. J. Mol. Sci. 2021, 22, 2388. [Google Scholar] [CrossRef]
- Schröder, A.; Bauer, K.; Spanier, G.; Proff, P.; Wolf, M.; Kirschneck, C. Expression kinetics of human periodontal ligament fibroblasts in the early phases of orthodontic tooth movement. J. Orofac. Orthop. 2018, 79, 337–351. [Google Scholar] [CrossRef]
- Ichioka, H.; Yamamoto, T.; Yamamoto, K.; Honjo, K.-I.; Adachi, T.; Oseko, F.; Mazda, O.; Kanamura, N.; Kita, M. Biomechanical force induces the growth factor production in human periodontal ligament-derived cells. Odontology 2016, 104, 27–34. [Google Scholar] [CrossRef]
- Pagano, S.; Lombardo, G.; Costanzi, E.; Balloni, S.; Bruscoli, S.; Flamini, S.; Coniglio, M.; Valenti, C.; Cianetti, S.; Marinucci, L. Morpho-functional effects of different universal dental adhesives on human gingival fibroblasts: An in vitro study. Odontology 2021, 109, 524–539. [Google Scholar] [CrossRef]
- Shah, A.A.; Sandler, J. Limiting factors in orthodontic treatment: 1. Factors related to patient, operator and orthodontic appliances. Dent. Update 2006, 33, 43–52. [Google Scholar] [CrossRef]
- Li, M.; Zhang, C.; Yang, Y. Effects of mechanical forces on osteogenesis and osteoclastogenesis in human periodontal ligament fibroblasts. Bone Jt. Res. 2019, 8, 19–31. [Google Scholar] [CrossRef]
- Kanzaki, H.; Chiba, M.; Shimizu, Y.; Mitani, H. Periodontal ligament cells under mechanical stress induce osteoclastogenesis by receptor activator of nuclear factor κB ligand up-regulation via prostaglandin E2 synthesis. J. Bone Miner. Res. 2002, 17, 210–220. [Google Scholar] [CrossRef] [Green Version]
- Jingwen, B.; Yaochen, L.; Guojun, Z. Cell cycle regulation and anticancer drug discovery. Cancer Biol. Med. 2017, 14, 348. [Google Scholar] [CrossRef] [PubMed]
- Janjic, M.; Docheva, D.; Trickovic Janjic, O.; Wichelhaus, A.; Baumert, U. In Vitro Weight-Loaded Cell Models for Understanding Mechanodependent Molecular Pathways Involved in Orthodontic Tooth Movement: A Systematic Review. Stem Cells Int. 2018, 2018, 3208285. [Google Scholar] [CrossRef] [Green Version]
- Niederau, C.; Craveiro, R.B.; Azraq, I.; Brockhaus, J.; Bastian, A.; Kirschneck, C.; Wolf, M. Selection and validation of reference genes by RT-qPCR for murine cementoblasts in mechanical loading experiments simulating orthodontic forces in vitro. Sci. Rep. 2020, 10, 10893. [Google Scholar] [CrossRef]
- Kaighn, M.; Narayan, K.S.; Ohnuki, Y.; Lechner, J.F.; Jones, L. Establishment and characterization of a human prostatic carcinoma cell line (PC-3). Investig. Urol. 1979, 17, 16–23. [Google Scholar]
- Schröder, S.K.; Asimakopoulou, A.; Tillmann, S.; Koschmieder, S.; Weiskirchen, R. TNF-α controls Lipocalin-2 expression in PC-3 prostate cancer cells. Cytokine 2020, 135. [Google Scholar] [CrossRef]
- Weider, M.; Schröder, A.; Docheva, D.; Rodrian, G.; Enderle, I.; Seidel, C.L.; Andreev, D.; Wegner, M.; Bozec, A.; Deschner, J.; et al. A human periodontal ligament fibroblast cell line as a new model to study periodontal stress. Int. J. Mol. Sci. 2020, 21, 7961. [Google Scholar] [CrossRef] [PubMed]
- Wolf, M.; Lossdörfer, S.; Craveiro, R.; Jäger, A. High-mobility group box protein-1 released by human-periodontal ligament cells modulates macrophage migration and activity in vitro. Innate Immun. 2014, 20, 688–696. [Google Scholar] [CrossRef] [Green Version]
- Nayak, B.; Galil, K.; Wiltshire, W.; PC, L. Molecular Biology of Orthodontic Tooth Movement Orthodontic Tooth Movement and ECM Remodeling Role of Cytokines, Growth Factors and. J. Dent. Oral Health 2013, 1, 13–15. [Google Scholar] [CrossRef]
- Marciniak, J.; Lossdörfer, S.; Knaup, I.; Bastian, A.; Craveiro, R.B.; Jäger, A.; Wolf, M. Orthodontic cell stress modifies proinflammatory cytokine expression in human PDL cells and induces immunomodulatory effects via TLR-4 signaling in vitro. Clin. Oral Investig. 2020, 24, 1411–1419. [Google Scholar] [CrossRef]
- Wu, J.; Li, Y.; Fan, X.; Zhang, C.; Wang, Y.; Zhao, Z. Analysis of gene expression profile of periodontal ligament cells subjected to cyclic compressive force. DNA Cell Biol. 2011, 30, 865–873. [Google Scholar] [CrossRef] [Green Version]
- Lombardi, M.L.; Jaalouk, D.E.; Shanahan, C.M.; Burke, B.; Roux, K.J.; Lammerding, J. The interaction between nesprins and sun proteins at the nuclear envelope is critical for force transmission between the nucleus and cytoskeleton. J. Biol. Chem. 2011, 286, 26743–26753. [Google Scholar] [CrossRef] [Green Version]
- Sawada, Y.; Tamada, M.; Dubin-Thaler, B.J.; Cherniavskaya, O.; Sakai, R.; Tanaka, S.; Sheetz, M.P. Force Sensing by Mechanical Extension of the Src Family Kinase Substrate p130Cas. Cell 2006, 127, 1015–1026. [Google Scholar] [CrossRef] [Green Version]
- Yoshigi, M.; Hoffman, L.M.; Jensen, C.C.; Yost, H.J.; Beckerle, M.C. Mechanical force mobilizes zyxin from focal adhesions to actin filaments and regulates cytoskeletal reinforcement. J. Cell Biol. 2005, 171, 209–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, L.M.; Jensen, C.C.; Chaturvedi, A.; Yoshigi, M.; Beckerle, M.C. Stretch-induced actin remodeling requires targeting of zyxin to stress fibers and recruitment of actin regulators. Mol. Biol. Cell 2012, 23, 1846–1859. [Google Scholar] [CrossRef]
- Blawat, K.; Mayr, A.; Hardt, M.; Kirschneck, C.; Nokhbehsaim, M.; Behl, C.; Deschner, J.; Jäger, A.; Memmert, S. Regulation of autophagic signaling by mechanical loading and inflammation in human PDL fibroblasts. Int. J. Mol. Sci. 2020, 21, 9446. [Google Scholar] [CrossRef]
- Memmert, S.; Nogueira, A.V.B.; Damanaki, A.; Nokhbehsaim, M.; Eick, S.; Divnic-Resnik, T.; Spahr, A.; Rath-Deschner, B.; Till, A.; Götz, W.; et al. Damage-regulated autophagy modulator 1 in oral inflammation and infection. Clin. Oral Investig. 2018, 22, 2933–2941. [Google Scholar] [CrossRef] [Green Version]
- Wolf, M.; Marciniak, J.; Lossdörfer, S.; Kirschneck, C.; Brauner, I.; Götz, W.; Jäger, A. Role of HSP70 protein in human periodontal ligament cell function and physiology. Ann. Anat. 2019, 221, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Wolf, M.; Lossdörfer, S.; Römer, P.; Kirschneck, C.; Küpper, K.; Deschner, J.; Jäger, A. Short-term heat pre-treatment modulates the release of HMGB1 and pro-inflammatory cytokines in hPDL cells following mechanical loading and affects monocyte behavior. Clin. Oral Investig. 2016, 20, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Kido, R.; Hiroshima, Y.; Kido, J.I.; Ikuta, T.; Sakamoto, E.; Inagaki, Y.; Naruishi, K.; Yumoto, H. Advanced glycation end-products increase lipocalin 2 expression in human oral epithelial cells. J. Periodontal Res. 2020, 55, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.; Gürbüz, N.; Özbalci, F.İ.; Koşkan, Ö.; Yetkin Ay, Z. Increase in serum and salivary neutrophil gelatinase-associated lipocalin levels with increased periodontal inflammation. J. Appl. Oral Sci. 2020, 28, e20200276. [Google Scholar] [CrossRef]
- Asimakopoulou, A.; Borkham-Kamphorst, E.; Tacke, F.; Weiskirchen, R. Lipocalin-2 (NGAL/LCN2), a “Help-Me” Signal in Organ Inflammation. Hepatology 2016, 63, 668–669. [Google Scholar] [CrossRef]
- Sunil, V.R.; Patel, K.J.; Nilsen-Hamilton, M.; Heck, D.E.; Laskin, J.D.; Laskin, D.L. Acute endotoxemia is associated with upregulation of lipocalin 24p3/Lcn2 in lung and liver. Exp. Mol. Pathol. 2007, 83, 177–187. [Google Scholar] [CrossRef] [Green Version]
- Sheng, C.; Mendler, I.H.; Rieke, S.; Snyder, P.; Jentsch, M.; Friedrich, D.; Drossel, B.; Loewer, A. PCNA-Mediated Degradation of p21 Coordinates the DNA Damage Response and Cell Cycle Regulation in Individual Cells. Cell Rep. 2019, 27, 48–58.e7. [Google Scholar] [CrossRef] [Green Version]
- Szelachowska, J.; Dziegiel, P.; Jelen-Krzeszewska, J.; Jelen, M.; Matkowski, R.; Pomiecko, A.; Spytkowska, B.; Jagas, M.; Gisterek, I.; Kornafel, J. Mcm-2 protein expression predicts prognosis better than Ki-67 antigen in oral cavity squamocellular carcinoma. Anticancer Res. 2006, 26, 2473–2478. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Name (Homo Sapiens) | 5′-Forward Primer-3′ (Length/Tm/%GC) | 5′-Reverse Primer-3′ (Length/Tm/%GC) |
|---|---|---|---|
| MCM2 | Minichromosome maintenance complex component 2 | gtggtactgctatggcggaat (21 bp/59.9 °C/52%GC) | tgagaggatcattgcctcgc (20 bp/59.4 °C/55%GC) |
| IL-6 | Interleukin 6 | catcctcgacggcatctcag (20 bp/60.32 °C/60%GC) | tcaccaggcaagtctcctca (20 bp/60.47 °C/55%GC) |
| IL-8 | Interleukin 8 | catactccaaacctttccacc (21 bp/57.9 °C/47,6%GC) | cttcaaaaacttctccacaacc (22 bp/56.9 °C/40.9%GC) |
| PCNA | Proliferating cell nuclear antigen | tggagaacttggaaatggaaac (22 bp/56.5 °C/40%GC) | gaactggttcattcatctctatgg (24 bp/59.3 °C/41%GC) |
| CCNA1 | Cyclin A1 | cccaagcaagggtttgacatc (21 bp/59.73 °C, 52%GC) | taccagcataggggaaactgtg (22 bp/59.76 °C/50%GC) |
| CCND1 | Cyclin D1 | gatgccaacctcctcaacga (20 bp/59.4 °C/55%GC) | gttcctcgcagacctccag (19 bp/61 °C/63%GC) |
| LCN2/NGAL | Lipocalin-2; Oncogene 24p3; Neutrophil gelatinase-associated lipocalin | ctccacctcagacctgatcc (20 bp/59 °C/60%GC) | acataccacttcccctggaat (21 bp/59 °C/48%GC) |
| RPL22 | Ribosomal protein L22 | tgattgcacccaccctgtag (20 bp/59.67 °C/55%GC) | ggttcccagcttttccgttc (20 bp/59.4 °C/55%GC) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brockhaus, J.; Craveiro, R.B.; Azraq, I.; Niederau, C.; Schröder, S.K.; Weiskirchen, R.; Jankowski, J.; Wolf, M. In Vitro Compression Model for Orthodontic Tooth Movement Modulates Human Periodontal Ligament Fibroblast Proliferation, Apoptosis and Cell Cycle. Biomolecules 2021, 11, 932. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11070932
Brockhaus J, Craveiro RB, Azraq I, Niederau C, Schröder SK, Weiskirchen R, Jankowski J, Wolf M. In Vitro Compression Model for Orthodontic Tooth Movement Modulates Human Periodontal Ligament Fibroblast Proliferation, Apoptosis and Cell Cycle. Biomolecules. 2021; 11(7):932. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11070932
Chicago/Turabian StyleBrockhaus, Julia, Rogerio B. Craveiro, Irma Azraq, Christian Niederau, Sarah K. Schröder, Ralf Weiskirchen, Joachim Jankowski, and Michael Wolf. 2021. "In Vitro Compression Model for Orthodontic Tooth Movement Modulates Human Periodontal Ligament Fibroblast Proliferation, Apoptosis and Cell Cycle" Biomolecules 11, no. 7: 932. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11070932