
Identification of HSP47 Binding Site on Native Collagen and Its Implications for the Development of HSP47 Inhibitors

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Expression and Purification of HSP47
2.2. Binding of HSP47 to Toolkit Peptides
2.3. Virtual Screening of Small Inhibitors Targeting Human the HSP47/Collagen Binding Interface
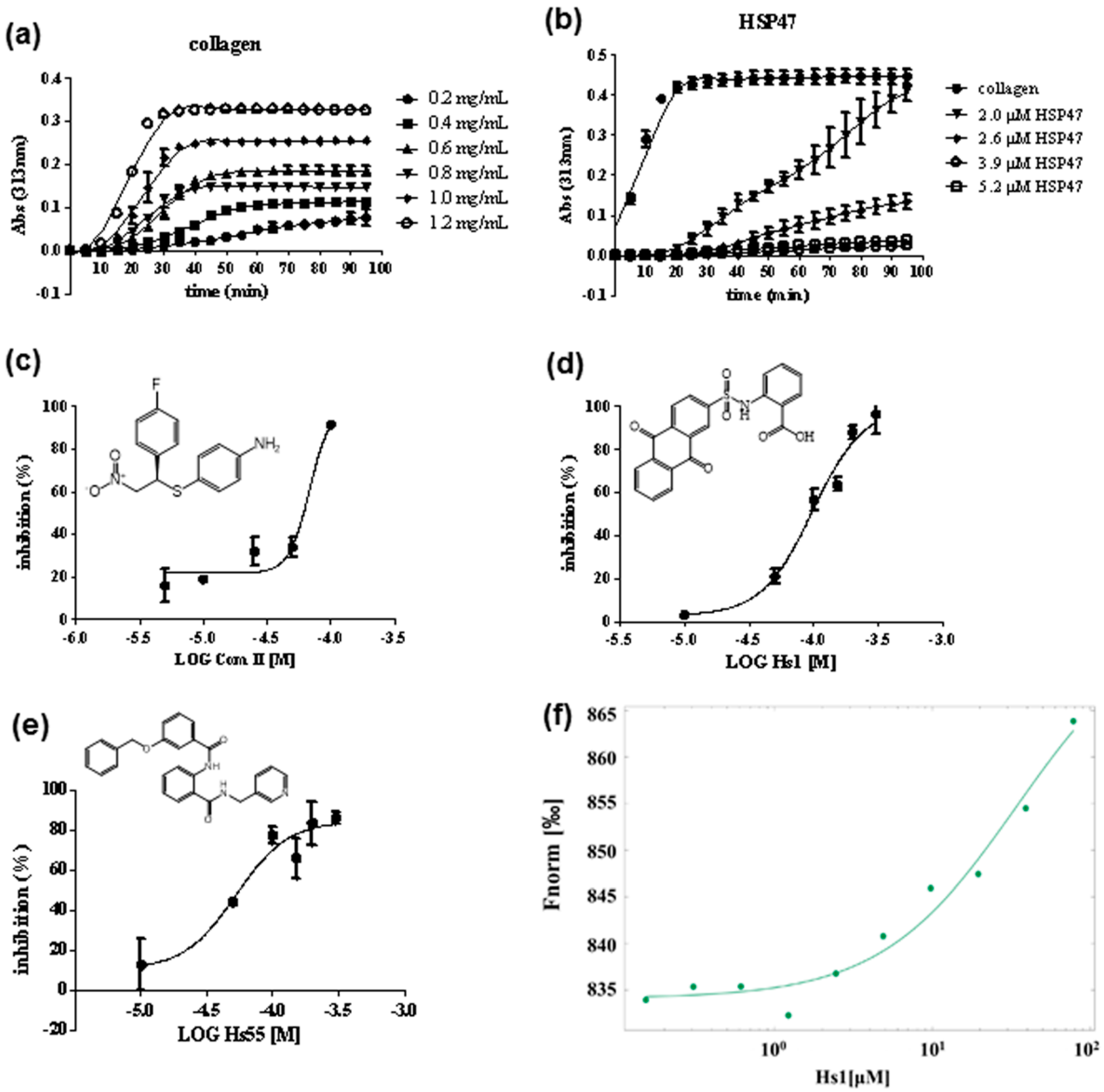
2.4. Turbidity Assay
2.5. Binding Affinity Measurement of HSP47 with Small Molecular Inhibitor
2.6. Molecular Docking of Small Inhibitors on HSP47
3. Results and Discussion
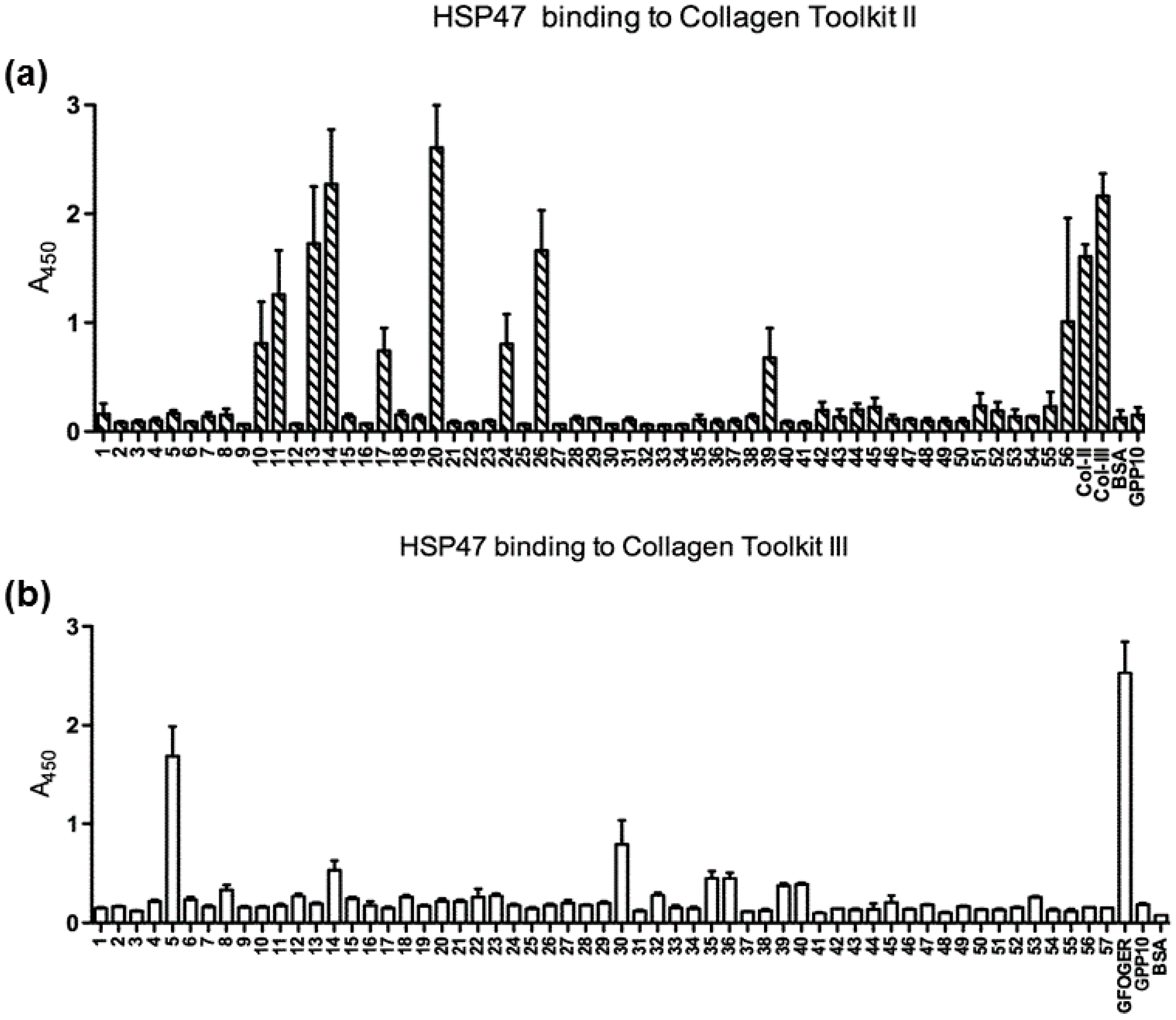
3.1. Binding of HSP47 to the Collagen Toolkit Peptides
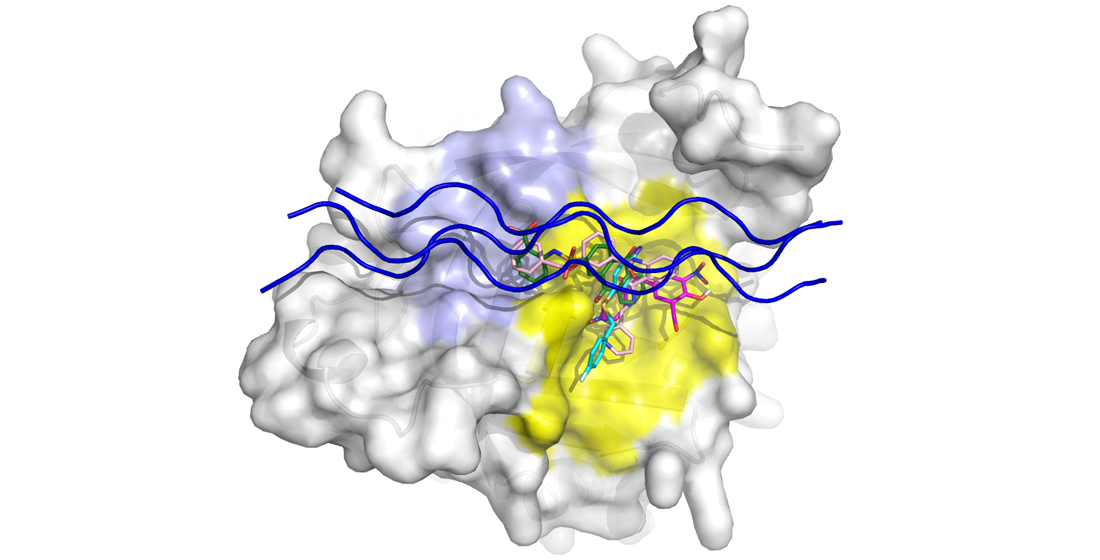
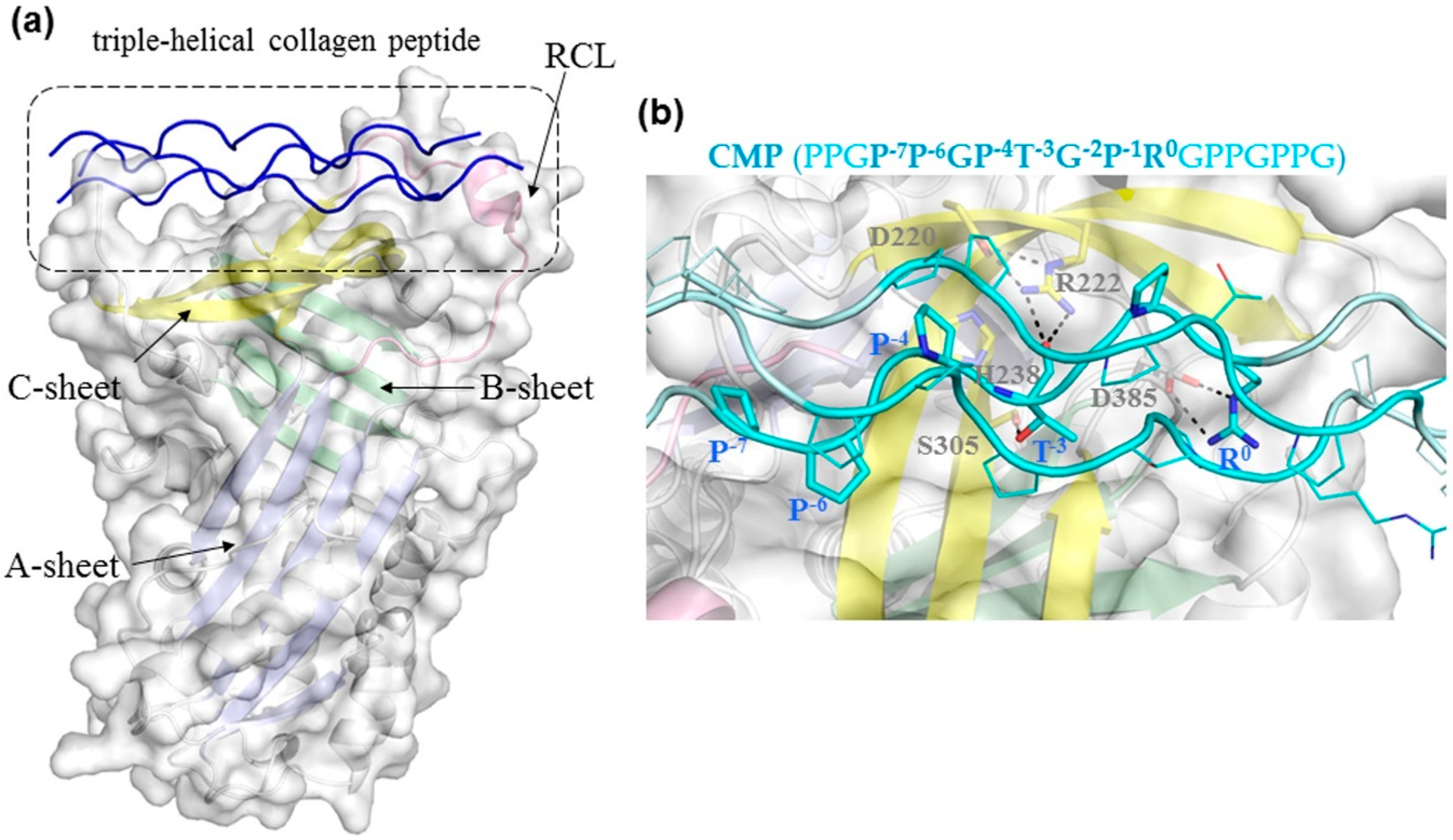
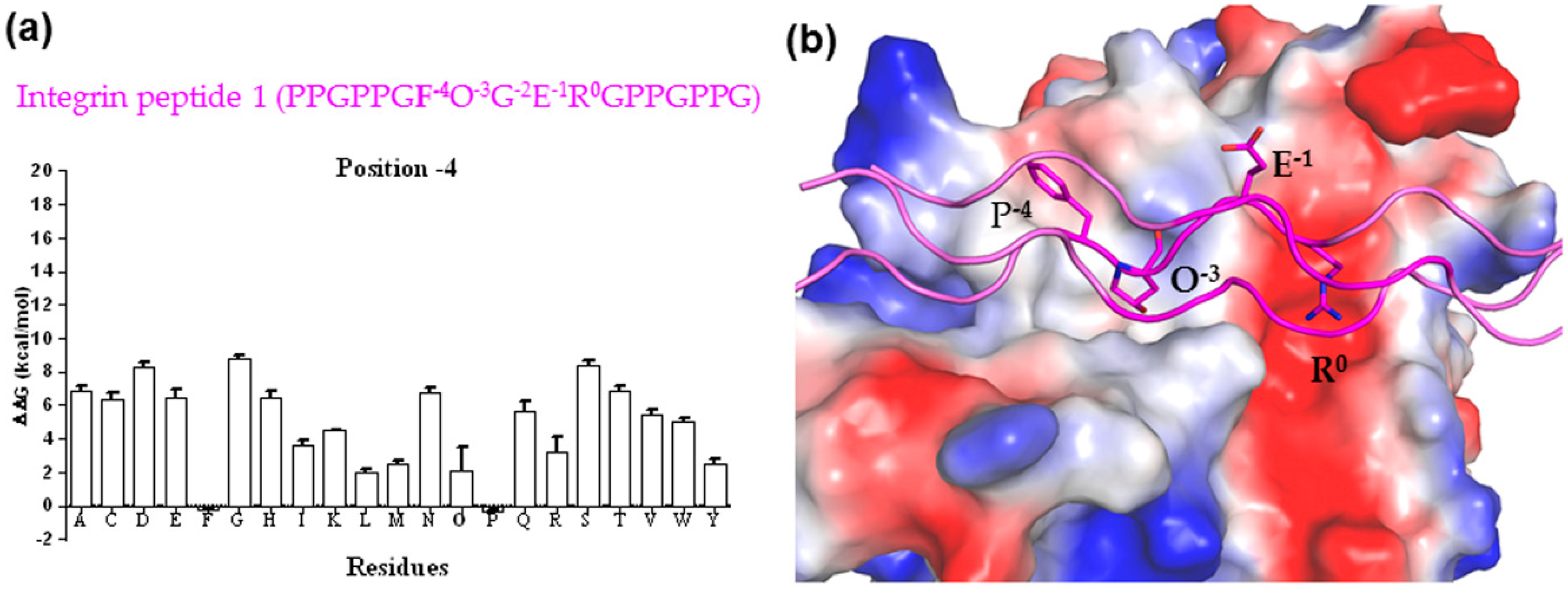
3.2. Binding Mechanism Analysis between HSP47 and Native Collagen Peptides
3.3. Screening and Identification of HSP47 Small Molecular Inhibitors
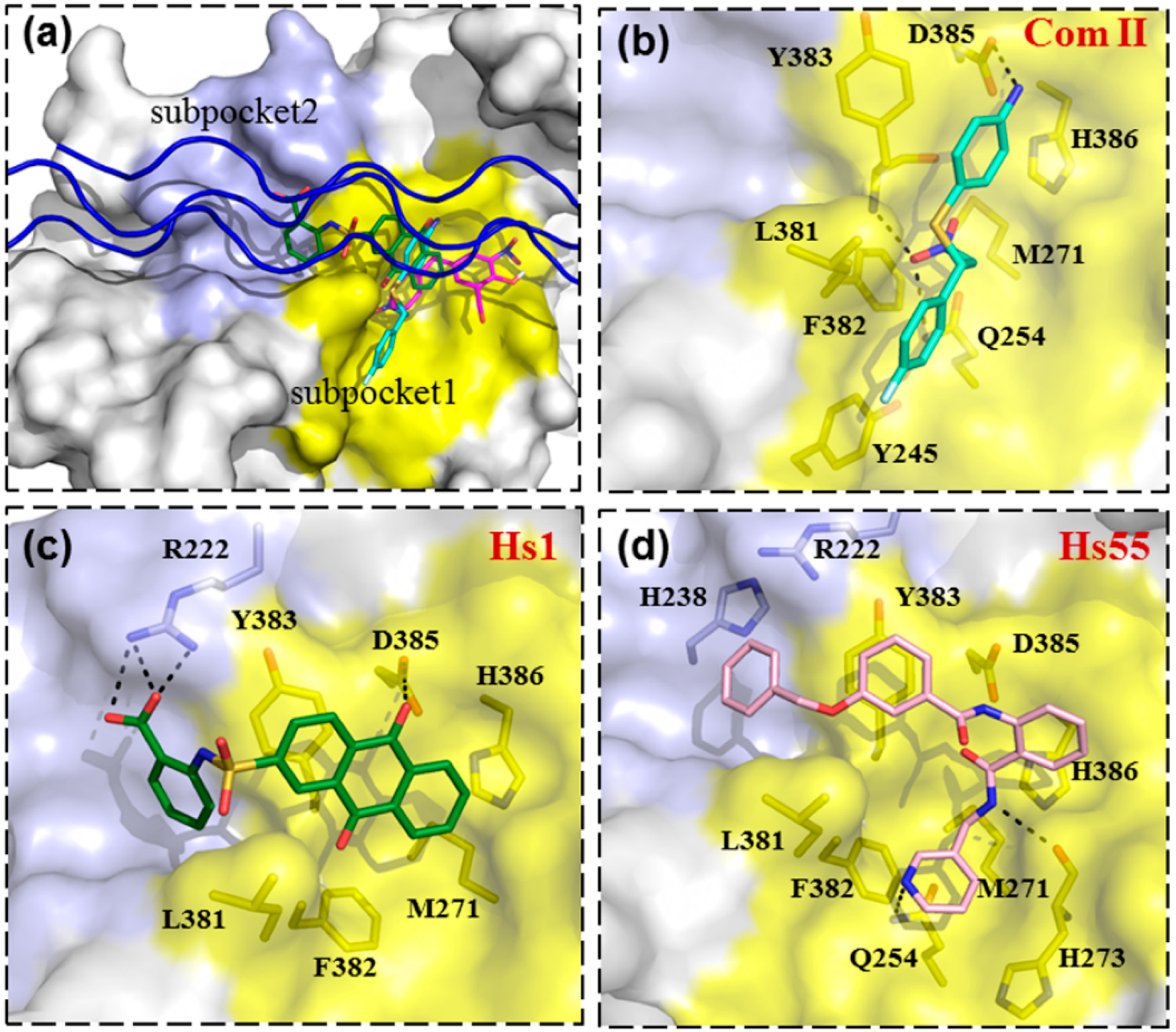
3.4. Binding Modes of Small Molecular Inhibitors on HSP47
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Prockop, D.J.; Kivirikko, K.I. Collagens: Molecular Biology, Diseases, and Potentials for Therapy. Annu. Rev. Biochem. 1995, 64, 403–434. [Google Scholar] [CrossRef] [PubMed]
- Lamande, S.R.; Bateman, J.F. Procollagen folding and assembly: The role of endoplasmic reticulum enzymes and molecular chaperones. Semin. Cell Dev. Biol. 1999, 10, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Hirayoshi, K.; Kudo, H.; Takechi, H.; Nakai, A.; Iwamatsu, A.; Yamada, K.M.; Nagata, K. HSP47: A tissue-specific, transformation-sensitive, collagen-binding heat shock protein of chicken embryo fibroblasts. Mol. Cell. Biol. 1991, 11, 4036–4044. [Google Scholar] [CrossRef] [Green Version]
- Ito, S.; Nagata, K. Biology of Hsp47 (Serpin H1), a collagen-specific molecular chaperone. Semin. Cell Dev. Biol. 2017, 62, 142–151. [Google Scholar] [CrossRef]
- Nagata, K.; Saga, S.; Yamada, K.M. A major collagen-binding protein of chick embryo fibroblasts is a novel heat shock protein. J. Cell Biol. 1986, 103, 223–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, C.; Ananthanarayanan, V.S. Structure-function studies on hsp47: pH-dependent inhibition of collagen fibril formation in vitro. Biochem. J. 2000, 349, 877–883. [Google Scholar] [CrossRef] [Green Version]
- Koide, T.; Takahara, Y.; Asada, S.; Nagata, K. Xaa-Arg-Gly triplets in the collagen triple helix are dominant binding sites for the molecular chaperone HSP47. J. Biol. Chem. 2002, 277, 6178–6182. [Google Scholar] [CrossRef] [Green Version]
- Nagai, N.; Hosokawa, M.; Itohara, S.; Adachi, E.; Matsushita, T.; Hosokawa, N.; Nagata, K. Embryonic lethality of molecular chaperone Hsp47 knockout mice is associated with defects in collagen biosynthesis. J. Cell Biol. 2000, 150, 1499–1506. [Google Scholar] [CrossRef] [Green Version]
- Masuda, H.; Fukumoto, M.; Hirayoshi, K.; Nagata, K. Coexpression of the collagen-binding stress protein HSP47 gene and the α1 (I) and α1 (III) collagen genes in carbon tetrachloride-induced rat liver fibrosis. J. Clin. Investig. 1994, 94, 2481–2488. [Google Scholar] [CrossRef]
- Honzawa, Y.; Nakase, H.; Shiokawa, M.; Yoshino, T.; Imaeda, H.; Matsuura, M.; Kodama, Y.; Ikeuchi, H.; Andoh, A.; Sakai, Y.; et al. Involvement of interleukin-17A-induced expression of heat shock protein 47 in intestinal fibrosis in Crohn’s disease. Gut 2014, 63, 1902–1912. [Google Scholar] [CrossRef]
- Friedman, S.L.; Sheppard, D.; Duffield, J.S.; Violette, S. Therapy for fibrotic diseases: Nearing the starting line. Sci. Transl. Med. 2013, 5, 167sr1. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Murase, K.; Kato, J.; Kobune, M.; Sato, T.; Kawano, Y.; Takimoto, R.; Takada, K.; Miyanishi, K.; Matsunaga, T.; et al. Resolution of liver cirrhosis using vitamin A-coupled liposomes to deliver siRNA against a collagen-specific chaperone. Nat. Biotechnol. 2008, 26, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Ishiwatari, H.; Sato, Y.; Murase, K.; Yoneda, A.; Fujita, R.; Nishita, H.; Birukawa, N.K.; Hayashi, T.; Sato, T.; Miyanishi, K.; et al. Treatment of pancreatic fibrosis with siRNA against a collagen-specific chaperone in vitamin A-coupled liposomes. Gut 2013, 62, 1328–1339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirai, K.; Kikuchi, S.; Kurita, A.; Ohashi, S.; Adachi, E.; Matsuoka, Y.; Nagata, K.; Watanabe, M. Immunohistochemical distribution of heat shock protein 47 (HSP47) in scirrhous carcinoma of the stomach. Anticancer Res. 2006, 26, 71–78. [Google Scholar]
- Zhu, J.; Xiong, G.; Fu, H.; Evers, B.M.; Zhou, B.P.; Xu, R. Chaperone Hsp47 drives malignant growth and invasion by modulating an ECM gene network. Cancer Res. 2015, 75, 1580–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasikumar, P.; AlOuda, K.S.; Kaiser, W.J.; Holbrook, L.M.; Kriek, N.; Unsworth, A.J.; Bye, A.P.; Sage, T.; Ushioda, R.; Nagata, K. The chaperone protein HSP47: A platelet collagen binding protein that contributes to thrombosis and hemostasis. J. Thromb. Haemost. 2018, 16, 946–959. [Google Scholar] [CrossRef] [Green Version]
- Thomson, C.A.; Atkinson, H.M.; Ananthanarayanan, V.S. Identification of small molecule chemical inhibitors of the collagen-specific chaperone Hsp47. J. Med. Chem. 2005, 48, 1680–1684. [Google Scholar] [CrossRef] [PubMed]
- Okano-Kosugi, H.; Matsushita, O.; Asada, S.; Herr, A.B.; Kitagawa, K.; Koide, T. Development of a high-throughput screening system for the compounds that inhibit collagen-protein interactions. Anal. Biochem. 2009, 394, 125–131. [Google Scholar] [CrossRef]
- Katarkar, A.; Haldar, P.K.; Chaudhuri, K. De novo design based pharmacophore query generation and virtual screening for the discovery of Hsp-47 inhibitors. Biochem. Biophys. Res. Commun. 2015, 456, 707–713. [Google Scholar] [CrossRef]
- Ito, S.; Ogawa, K.; Takeuchi, K.; Takagi, M.; Yoshida, M.; Hirokawa, T.; Hirayama, S.; Shinya, K.; Shimada, I.; Doi, T.; et al. A small-molecule compound inhibits a collagen-specific molecular chaperone and could represent a potential remedy for fibrosis. J. Biol. Chem. 2017, 292, 20076–20085. [Google Scholar] [CrossRef] [Green Version]
- Miyamura, T.; Sakamoto, N.; Kakugawa, T.; Taniguchi, H.; Akiyama, Y.; Okuno, D.; Moriyama, S.; Hara, A.; Kido, T.; Ishimoto, H.; et al. Small molecule inhibitor of HSP47 prevents pro-fibrotic mechanisms of fibroblasts in vitro. Biochem. Biophys. Res. Commun. 2020, 530, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Raynal, N.; Hamaia, S.W.; Siljander, P.R.M.; Maddox, B.; Peachey, A.R.; Fernandez, R.; Foley, L.J.; Slatter, D.A.; Jarvis, G.E.; Farndale, R.W. Use of synthetic peptides to locate novel integrin α2β1-binding motifs in human collagen III. J. Biol. Chem. 2006, 281, 3821–3831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koide, T.; Nishikawa, Y.; Asada, S.; Yamazaki, C.M.; Takahara, Y.; Homma, D.L.; Otaka, A.; Ohtani, K.; Wakamiya, N.; Nagata, K.; et al. Specific recognition of the collagen triple helix by chaperone HSP47: II. The HSP47-binding structural motif in collagens and related proteins. J. Biol. Chem. 2006, 281, 11177–11185. [Google Scholar] [CrossRef] [Green Version]
- Widmer, C.; Gebauer, J.M.; Brunstein, E.; Rosenbaum, S.; Zaucke, F.; Drogemuller, C.; Leeb, T.; Baumann, U. Molecular basis for the action of the collagen-specific chaperone Hsp47/SERPINH1 and its structure-specific client recognition. Proc. Natl. Acad. Sci. USA 2012, 109, 13243–13247. [Google Scholar] [CrossRef] [Green Version]
- Farndale, R.W.; Lisman, T.; Bihan, D.; Hamaia, S.; Smerling, C.S.; Pugh, N.; Konitsiotis, A.; Leitinger, B.; de Groot, P.G.; Jarvis, G.E.; et al. Cell-collagen interactions: The use of peptide toolkits to investigate collagen-receptor interactions. Biochem. Soc. Trans. 2008, 36, 241–250. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; De Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic. Acids. Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schymkowitz, J.; Borg, J.; Stricher, F.; Nys, R.; Rousseau, F.; Serrano, L. The FoldX web server: An online force field. Nucleic Acids Res. 2005, 33, W382–W388. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.; Huey, R. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Oleg, T.; Arthur, J.O. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar]
- Hamaia, S.W.; Pugh, N.; Raynal, N.; Némoz, B.; Stone, R.; Gullberg, D.; Bihan, D.; Farndale, R.W. Mapping of potent and specific binding motifs, GLOGEN and GVOGEA, for integrin 1αβ1 using collagen Toolkits II and III. J. Biol. Chem. 2012, 287, 26019–26028. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, Y.; Takahara, Y.; Asada, S.; Shigenaga, A.; Otaka, A.; Kitagawa, K.; Koide, T. A structure-activity relationship study elucidating the mechanism of sequence-specific collagen recognition by the chaperone HSP47. Bioorg. Med. Chem. 2010, 18, 3767–3775. [Google Scholar] [CrossRef] [PubMed]
- Farndale, R.W. Collagen-binding proteins: Insights from the Collagen toolkits. Essays Biochem. 2019, 63, 337–348. [Google Scholar] [PubMed]
- Knight, C.G.; Morton, L.F.; Peachey, A.R.; Tuckwell, D.S.; Farndale, R.W.; Barnes, M.J. The collagen-binding A-domains of integrin α1β1 and α2β1 recognize the same specific amino acid sequence, GFOGER, in native (triple-helical) collagens. J. Biol. Chem. 2000, 275, 35–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siljander, P.R.; Hamaia, S.; Peachey, A.R.; Slatter, D.A.; Smethurst, P.A.; Ouwehand, W.H.; Knight, C.G.; Farndale, R.W. Integrin activation state determines selectivity for novel recognition sites in fibrillar collagens. J. Biol. Chem. 2004, 279, 47763–47772. [Google Scholar] [CrossRef] [Green Version]
- Williams, B.R.; Gelman, R.A.; Poppke, D.C.; Piez, K. Collagen fibril formation. Optimal in vitro conditions and preliminary kinetic results. J. Biol. Chem. 1978, 253, 6578–6585. [Google Scholar] [CrossRef]
- Yoshida, M.; Saito, M.; Ito, S.; Ogawa, K.; Goshima, N.; Nagata, K.; Doi, T. Structure-Activity Relationship Study on Col-003, a Protein-Protein Interaction Inhibitor between Collagen and Hsp47. Chem. Pharm. Bull. 2020, 68, 220–226. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Name | Sequence | Mean A450 |
|---|---|---|
| Toolkit II | ||
| 14 | ---------------------------------------GPPGPRGPOGPQGATGPLGPKGQTGEOGIA GPP | >2 |
| 20 | ------GPPGANGDOGROGEOGLOGARGLTGROGDAGPP | >2 |
| 13 | -GPPGAKGSAGAOGIAGAOGFOGPRGPOGPQGPP | 1.5–2 |
| 26 | ---------------------------------------GPPGERGEQGAOGPSGFQGLOGPOGPOGEGGPP | 1.5–2 |
| 11 | --------------------GPPGARGPEGAQGPRGEOGTOGSOGPAGASGPP | 1–1.5 |
| 10 | GPPGGOGFOGAOGAKGEAGPTGARGPEGAQGPP | 0.5–1 |
| 17 | - GPPGKRGARGEOGGVGPIGPOGERGAOGNRGPP | 0.5–1 |
| 24 | -------------GPPGKAGEKGLOGAOGLRGLOGKDGETGAAGPP | 0.5–1 |
| 39 | ---------------------------------------GPPGARGAQGPOGATGFOGAAGRVGPOGSNGPP | 0.5–1 |
| Toolkit III | ||
| 5 | ---------------------------------------GPPGER GLOGPOGIKGPAGIOGFOGMKGHRGPP | 1.5–2 |
| 30 | --------------------------------GPPGAOGLRGGAGPOGPEGGKGAAGPOGPOGPP | 0.5–1 |
| 14 | --------------------GPPGIOGAOGLMGARGPOGPAGANGAOGLRGPP | 0.5–1 |
| Integrin Peptide | ||
| 1 | ---------------------------------GPPGFOGERGPP | >2 |
| 2 | --------------------------------GPPGMOGERGPP | 1.5–2 |
| 3 | ---------------------------------GPPGAOGERGPP | 1.5–2 |
| 4 | ---------------------------------GPPGLOGERGPP | 1.5–2 |
| 5 | ---------------------------------GPPGROGERGPP | 0–0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, H.; Sasikumar, P.; Little, G.; Bihan, D.; Hamaia, S.W.; Zhou, A.; Gibbins, J.M.; Farndale, R.W. Identification of HSP47 Binding Site on Native Collagen and Its Implications for the Development of HSP47 Inhibitors. Biomolecules 2021, 11, 983. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11070983
Cai H, Sasikumar P, Little G, Bihan D, Hamaia SW, Zhou A, Gibbins JM, Farndale RW. Identification of HSP47 Binding Site on Native Collagen and Its Implications for the Development of HSP47 Inhibitors. Biomolecules. 2021; 11(7):983. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11070983
Chicago/Turabian StyleCai, Haiyan, Parvathy Sasikumar, Gemma Little, Dominique Bihan, Samir W. Hamaia, Aiwu Zhou, Jonathan M. Gibbins, and Richard W. Farndale. 2021. "Identification of HSP47 Binding Site on Native Collagen and Its Implications for the Development of HSP47 Inhibitors" Biomolecules 11, no. 7: 983. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11070983