1. Introduction
Gastric cancer remains the leading cause of cancer-related deaths despite a steady decline in incidence rates, attributed to improved hygiene and diet and the popularization of therapies against
Helicobacter pylori [
1]. While efficient in reducing the occurrence of non-cardia subtype of gastric adenocarcinoma (GA), they failed to stop the rising incidence of cardia subtype (CA) [
2]. Poor survival rates characteristic for gastric cancer result from the disease being diagnosed at advanced stage, when it is not amenable for curative resection, leaving chemotherapy as a major therapeutic option [
1]. However, chemotherapy is not only highly toxic but its effectiveness is below expectations. Therefore, “a radical shift toward precision medicine” is advocated. However, this requires unraveling the molecular landscape of gastric cancer in order to establish novel therapeutic targets and biomarkers [
2].
Reprogramming of metabolic pathways is a well-recognized hallmark of cancer [
3], going well beyond the Warburg effect [
4]. Therefore, untargeted and targeted metabolic profiling is viewed as a promising tool in precision medicine [
5]. However, recent advances in metabolomics showed that the alterations are neither uniform across cancer types nor constant over time. Rather, they reflect cancer heterogeneity as well as its progression [
6].
L-arginine (Arg) metabolic pathways are among those repeatedly found to be deregulated [
7,
8,
9]. Pharmacological manipulation of pathway enzymes is viewed as an attractive therapeutic approach while monitoring enzyme activity and/or metabolite concentration may aid cancer diagnosis and treatment [
10,
11,
12,
13]. However, a better understanding of pathway status and function in cancer is needed as even the role of arginine, the pathway precursor, is ambiguous [
13]. Arginine is competed for by immune and cancer cells, either contributing to immunosurveillance or supporting tumor growth and metastasis [
13]. Still, the amino acid role in gastric cancer might not be unequivocally tumor-supporting. Uncharacteristically, arginine has been shown to inhibit growth of gastric cancer cells in vitro by upregulating caspase 8 expression and consequently inducing apoptosis [
14].
Synthesis of NO by NO synthases (NOSs) and synthesis of L-ornithine (ornithine; Orn) by arginases (ARGs), as a precursor of polyamines synthesized by ornithine decarboxylase (ODC), are two main competitive ways of Arg utilization [
11]. NO synthesis is inhibited by methylated arginine derivatives such as asymmetric and symmetric dimethylarginines (ADMA and SDMA), products of protein arginine methyltransferases (PRMTs), as they compete with Arg for NOS as well as for amino acid transporters. The ADMA is metabolized to L-citrulline (citrulline; Cit) and dimethylamine (DMA) by dimethylarginine dimethylaminohydrolases (DDAHs). Citrulline can be recycled to arginine in a two-step reaction catalyzed by argininosuccinate synthase-1 (ASS1) and argininosuccinate lyase (ASL) [
10,
15,
16]. A basic overview of key players of arginine metabolic pathways and their interrelationship is depicted in
Figure 1.
Considering the pressing need for new molecular targets and biomarkers for gastric cancer and the growing interest in Arg metabolism in this capacity, our aim was to explore the pathway status in gastric cancer in order to identify aberrations with therapeutic and diagnostic potential. In the present study, a comprehensive analysis of systemic metabolite concentration (Arg, Cit, Orn, ADMA, SDMA, and DMA) and local enzyme (ASL, ARG1, ARG2, ASS1, DDAH1, DDAH2, NOS2, ODC1, PRMT1, PRMT2, and PRMT5) and transporter (ORNT1) expression was conducted. Metabolomic and transcriptomic data were referred to cancer anatomical subsite and pathology. Patterns of interrelationships of pathway players and their correlation with local and systemic immune, inflammatory, and angiogenic mediators and other molecules relevant for cancer were examined.
4. Discussion
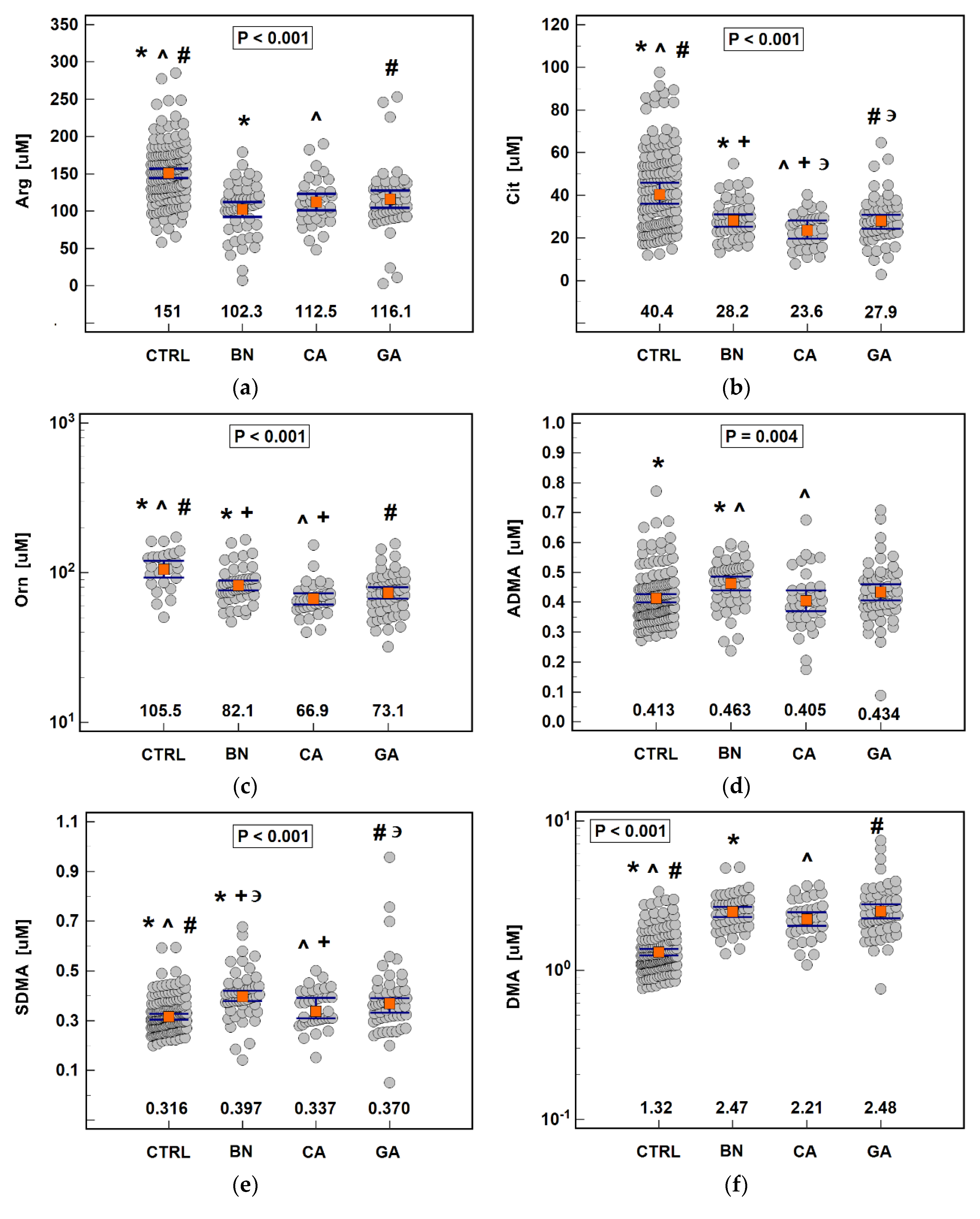
Metabolomic profiling is viewed as a promising approach to discover novel biomarkers facilitating cancer diagnosis and differentiation, monitoring treatment efficacy, and prognostication [
5]. In the present study, the analysis of metabolites associated with arginine metabolism clearly indicated that amino acid deficiency occurs at systemic level already in patients with benign gastric disorders, and is further exacerbated in cardiac cancer. Moreover, depletion of citrulline and ornithine was aggravated in cancer patients with lymph node and distant metastases. Consistently, amino acid concentrations were negatively correlated with a set of immune modulators and proinflammatory cytokines as well as cancer-promoting growth factors. A gastric cancer-related systemic drop in arginine and citrulline, but not ornithine, has previously been shown by Miyagi et al. [
21], and plasma-free amino acid profiling has been successfully explored as a diagnostic tool. Therefore, we assessed the individual discriminative power of arginine, ornithine, and citrulline and found ornithine to be superior cancer marker with overall accuracy exceeding 80%. Citrulline, in turn, was the only metabolite able to discriminate cancer patients by anatomical subsite of primary tumor, although its power was only moderate. Contrary to amino acids, the other evaluated metabolites—dimethylarginines and DMA—were elevated in patients with benign gastric disorders and SDMA and DMA also in cancer patients. Ornithine, ADMA, and SDMA were moderately efficient in discriminating patients with cancer from those with benign gastric disorders. Among assessed metabolites, DMA was the best individual marker of cancer presence. Still, we demonstrated that concomitant quantification of all metabolites was superior to individual determinations in terms of diagnostic power in overall cancer detection, differentiation between benign and cancerous gastric diseases, and distinguishing CA from GA with, respectively, 91%, 70%, and 66% overall accuracy.
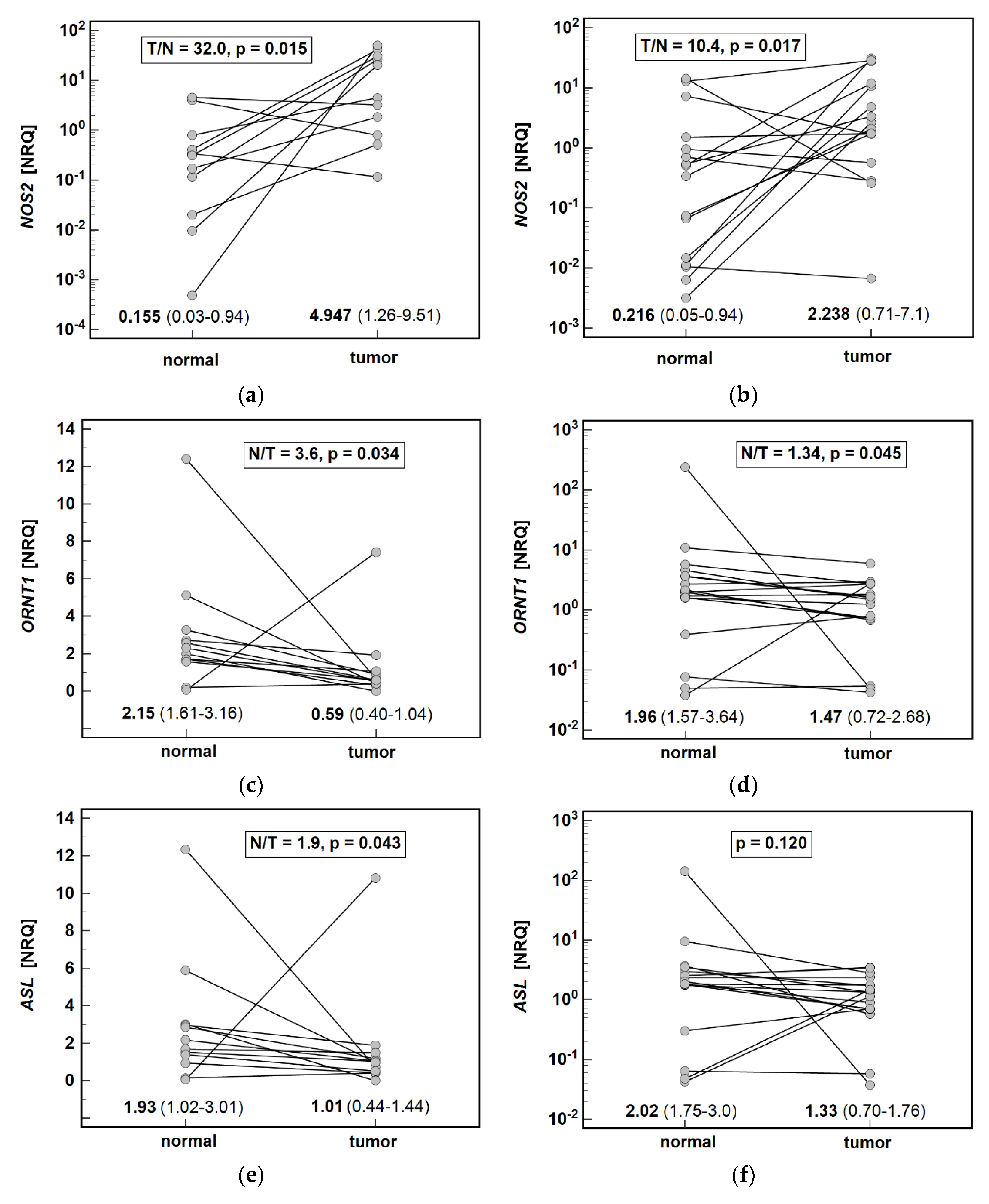
Systemic arginine depletion was accompanied by upregulated local
NOS2 expression while the expression levels of
ARGs were unaltered between tumors and adjacent mucosa. This observation is consistent with that of Wang et al. [
22] who showed upregulated NOS2 in gastric tumors. However, it does not confirm previous findings on the upregulation of ARG in breast [
23] and that of ODC in gastric [
24] tumors. As NOS2 overexpression has been associated with transformed epithelial cells [
22] and that of ARG and ODC with tumor-infiltrating macrophages [
23,
25], possible low content of these immune cells in tumors analyzed in the present study might account for lack of ARG and ODC upregulation. Unlike immunochemistry, the RTqPCR technique is fully quantitative but does not allow for determining the cellular source of expression. Noteworthily, lack of gene upregulation in tumors compared to adjacent tissue may not indicate lack of cancer-related gene upregulation. It has been repeatedly demonstrated that tumor-adjacent tissue might already have upregulated gene expression, even indicating comparative gene downregulation in tumors, despite lack of morphological and histological changes in its architecture [
26,
27,
28,
29]. Such apparent downregulation, resulting from less pronounced upregulation in tumors, has also been noted for
ARG1 expression in the colon [
7]. No normal gastric mucosa was available in the current study to confirm the speculation, but the notion is supported by higher ODC activity in non-transformed mucosa from GC patients than normal mucosa from healthy individuals reported by others [
24].
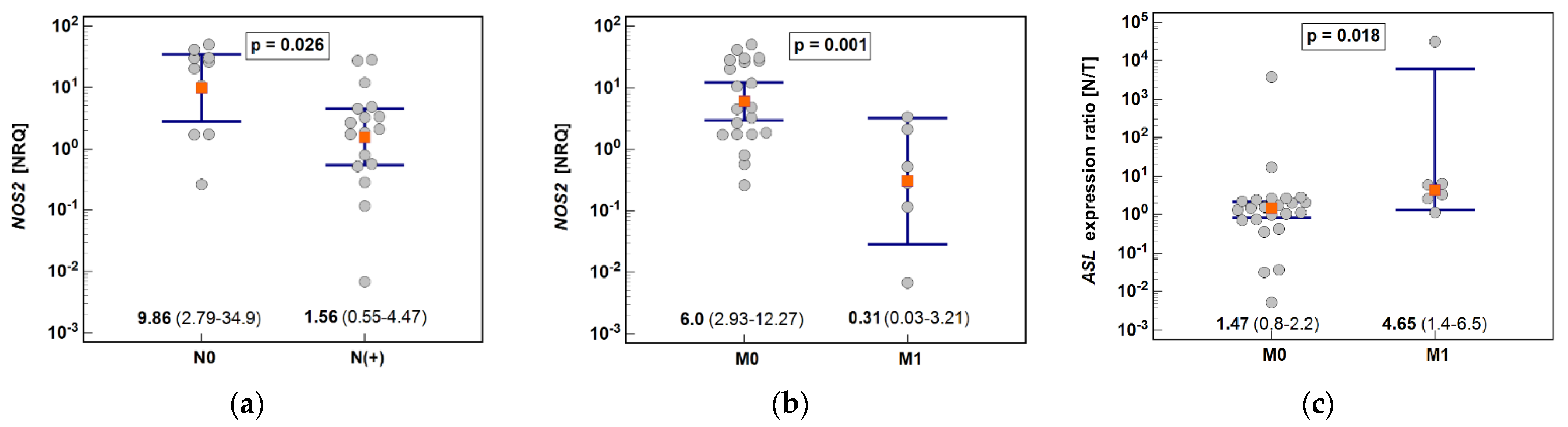
We confirmed, on a larger set of samples, our previous observation [
19] that
NOS2 upregulation is greater in cardia subtype of gastric cancer. The downregulation of
ORNT1,
ASL, and
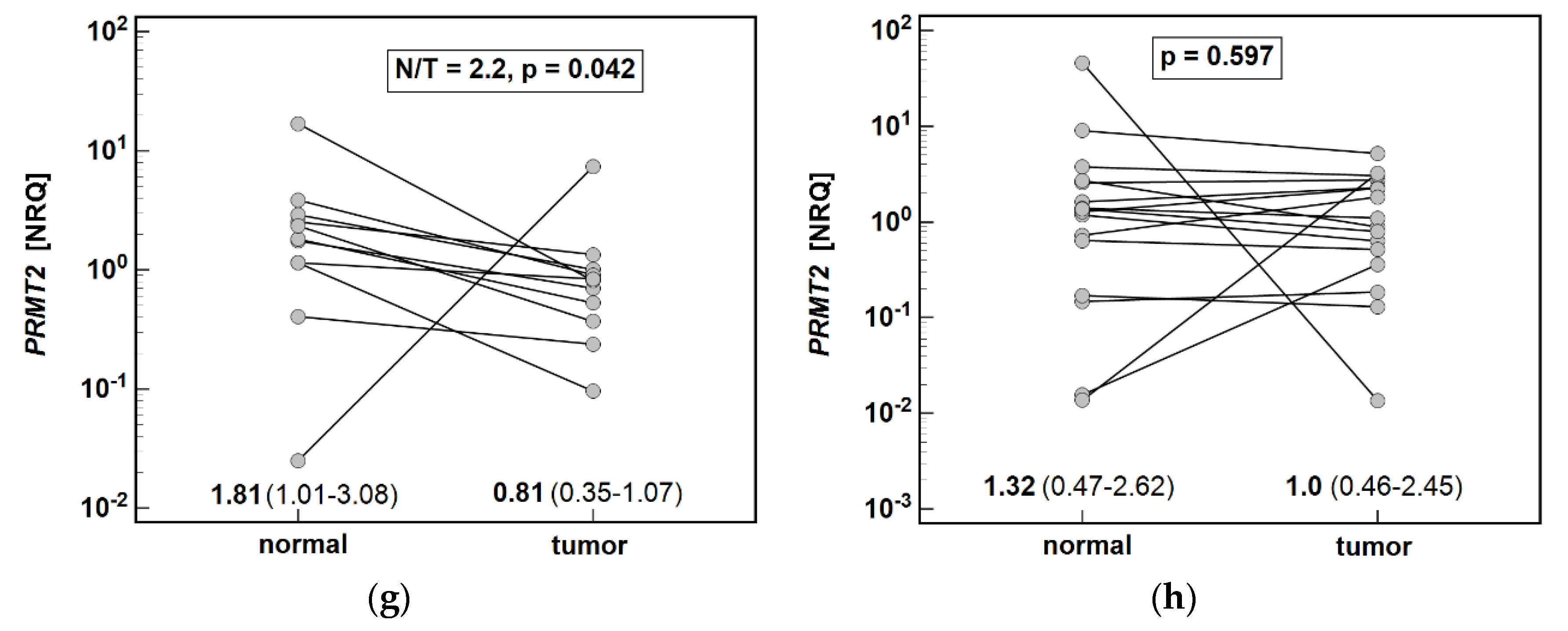
PRMT2 was more evident in cardia subtype as well. Considering the tumor-supporting consequences of enzyme deregulation, this finding might shed some light on molecular background of more aggressive phenotype and worse prognosis of cardia than non-cardia gastric cancer [
30].
Elevated concentration of dimethylarginines, more so in benign disorders than in cancer, is in line with inflammatory character of SDMA and the role attributed to ADMA in gastric injury. ADMA has been shown to induce inflammatory response and oxidative stress in gastric mucosa [
31,
32] and mediate cell migration and invasion via Wnt/β-catenin signaling pathway [
33]. The impact of SDMA accumulation is mostly unknown, but in colorectal tumors, it has been linked with greater metastatic potential [
34]. Here, both ADMA and SDMA were positively correlated with systemic hepatocyte growth factor (HGF), pivotal for gastric cancer development and progression [
35]. SDMA was also correlated with stroma-derived (SCF), the signaling of which is involved in viability and self-renewing properties of cancer stem cells [
36]. The interrelationship between intermediates of arginine metabolic pathways and stemness-promoting cytokines, recurring in various cancers [
9], is intriguing and worth exploration.
Corroborating findings of others [
32,
37], systemic elevation of ADMA was accompanied by
DDAH1 downregulation. A proneoplastic role has generally been attributed to DDAHs [
12] and the only evidence of DDAH1 acting as a tumor suppressor has been shown in gastric cancer [
12,
37]. Still, a co-expression pattern of
DDAH1 observed here might imply a tumor-promoting role.
DDAH1 expression was positively correlated with genes encoding proliferation marker Ki67, anti-apoptotic BCLXL, and mesenchymal marker claudin-2 (
CLDN2), with
BCLXL being an independent predictor of
DDAH1 expression. Similar correlation patterns were observed for
DDAH2, a dominant endothelial isoform.
DDAH activity yields citrulline and DMA. The status and role of DMA in gastric cancer is largely unknown. Nonetheless, its accumulation is likely disadvantageous as DMA is a precursor of nitrosodimethylamine—a suspected carcinogen—and can be effectively transported from blood into gastric fluid [
38]. DMA in our patients was positively correlated with SDMA and with SCGFβ, a recently discovered secreted sulfated glycoprotein of unknown status and role in gastric cancer, which, however, is a marker of drug-resistance in lung and liver cancers [
39].
ASS1 and ASL are involved in intracellular de novo synthesis of arginine from citrulline. Counterintuitively, however, number of cancers downregulate ASS1 and become arginine auxotrophic. The proposed advantage of ASS1 downregulation for tumors is the promotion of proliferation under normal conditions, greater invasiveness under hypoxia, and a buildup of glutamine under acidic conditions [
40,
41]. Herein,
ASS1 expression was not significantly affected but, in line with its proposed pro-survival role in gastric cancer, it was positively correlated with
Ki67 and
BCLXL. Less is known about ASL, which is the only enzyme able to synthesize arginine endogenously and its silencing also results in arginine auxotrophy [
15]. To the best of our knowledge,
ASL expression in gastric cancer has not been investigated. Here, we found it to be downregulated in tumors by two-fold on average. Moreover,
ASL downregulation was more pronounced in cancers metastasizing to distant organs and
ASL expression in tumor was inversely correlated with depth of invasion. Downregulation of
ASL is of clinical relevance as, if confirmed on a larger set of samples, it implies that gastric cancer, particularly the one located in cardia, might still be sensitive to arginine-deprivation therapies, despite
ASS1 overexpression.
Among cancer-related metabolic aberrations, dysregulation of urea cycle enzymes is quite prevalent and linked with worse overall prognosis but better response to immunotherapy based on checkpoint inhibitors [
42]. As the enzymes of the cycle compete for nitrogenous substrates with others, loss-of-function mutations in genes encoding
ASS1 or
ASL or ornithine translocase (
ORNT1) facilitate pyrimidine synthesis by dihydrooratase, and results, as a consequence, in increased cell proliferation [
42]. Therefore, downregulation of
ASL combined with even more accentuated lower expression of
ORNT1 might potentially translate into metabolic rewiring promoting pyrimidine synthesis. In line with Lee et al.’s [
42] observations, determining expression level of
ASL and
ORNT1 might therefore help identify gastric cancer patients more likely to benefit from immune checkpoint inhibitor therapy.
Patients with benign gastric disorders had significantly higher systemic concentrations of dimethylarginines and SDMA was higher also in cancer patients. ADMA and SDMA are products of, respectively, type I and type II PRMTs, but dysregulation of dimethylarginines was not reflected locally and
PRMT1 (prototypical type I enzyme) and
PRMT5 (prototypical type II enzyme) expression levels were unaltered. PRMT upregulation in certain cancer types [
9] has been noted and evoked an interest in PRMTs and their inhibitors as potential antineoplastic strategy [
43,
44]. Still, unaltered PRMT expression agrees well with the housekeeping nature of those enzymes, further underscored by their tight interrelationship observed here. It is worth mentioning, however, that lack of
PRMT1 and
PRMT5 elevation in tumors might not necessarily mean that enzyme expression is not affected by cancer. As we have previously shown in colorectal cancer,
PRMT1 and
PRMT5 expression can be upregulated both in colonic tumors and adjacent tissue [
7].
Unlike main isoforms, tumor PRMT2 expression was clearly downregulated in CA and reflected the depth of tumor invasion. Contrary to PRMT1 and PRMT5, little is known about PRMT2 and its potential substrates. In fact, enzyme has even been suspected of lack of methyltransferase activity [
45]. Nonetheless, it has been shown that PRMT2 may act as a coactivator for various receptors. However, its partners are implicated in opposing activities, either facilitating or inhibiting tumor growth [
45]. While PRMT2 status and role in gastric cancer does not seem to be previously investigated, our observation on diminished PRMT2 expression in cardia tumors is in line with antitumor activity played by peroxisome proliferator-activated receptor γ (PPARγ) in gastric carcinogenesis [
46], even though the receptor is reportedly upregulated in gastric tumors [
47]. Moreover, it also agrees well with growth inhibition exerted by PRMT2 in breast cancer [
48].


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}