
Altered Expression of Protamine-like and Their DNA Binding Induced by Cr(VI): A Possible Risk to Spermatogenesis?

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Mussels Sampling and Exposure to Cr(VI)
2.3. Gonad Sampling and PL Proteins Extraction
2.4. Plasmid DNA Preparation
2.5. Extraction of Sperm DNA
2.6. Electrophoretic Analysis
2.7. Analysis of the Effect of M. galloprovincialis PL Proteins on DNA Electrophoretic Mobility
2.8. DNA Protection Analysis
2.9. RNA Extraction, cDNA Synthesis and RT-qPCR
2.10. Determination of the Sperm Motility
2.11. Fluorescence Spectroscopy of PL Proteins
3. Results
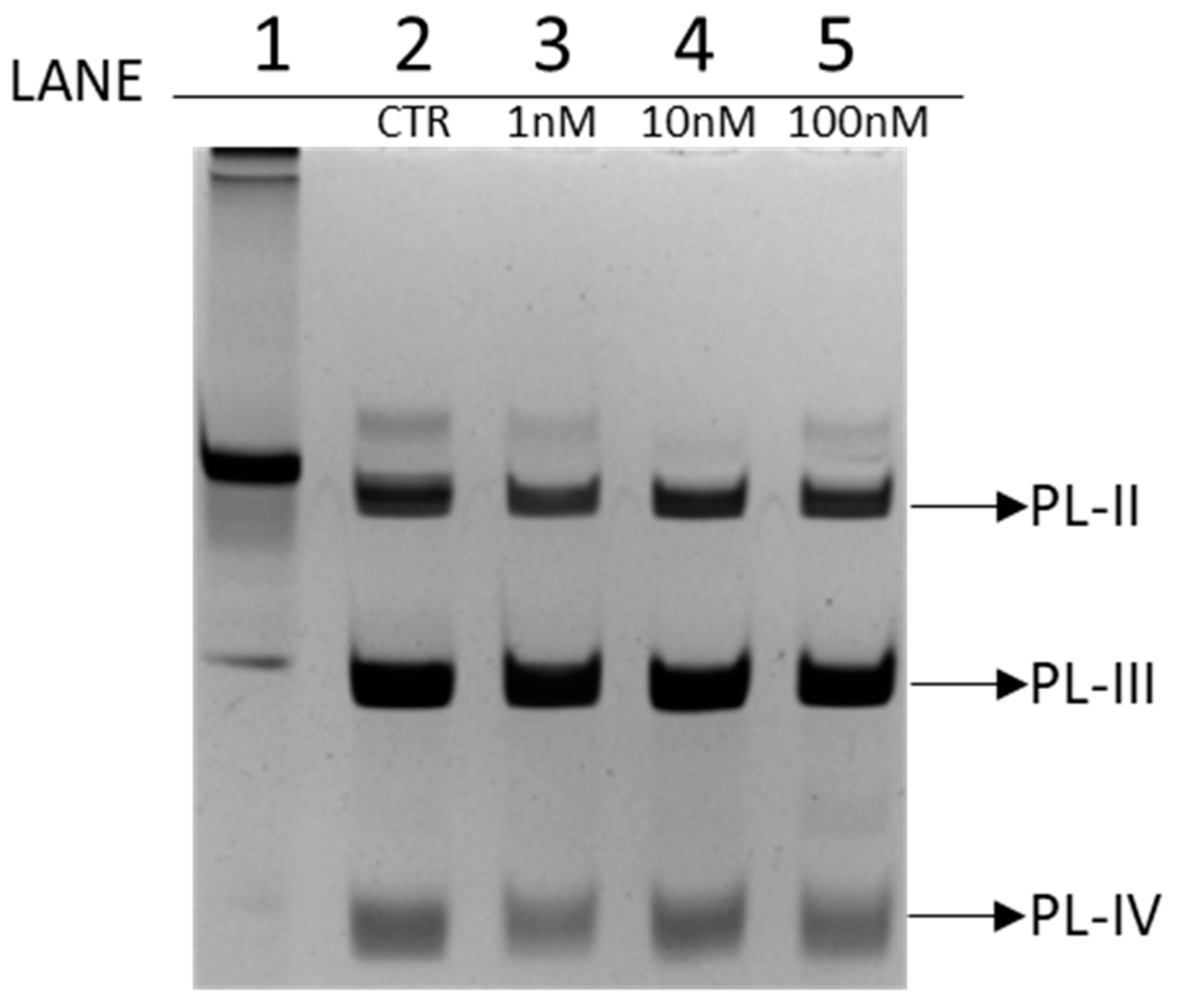
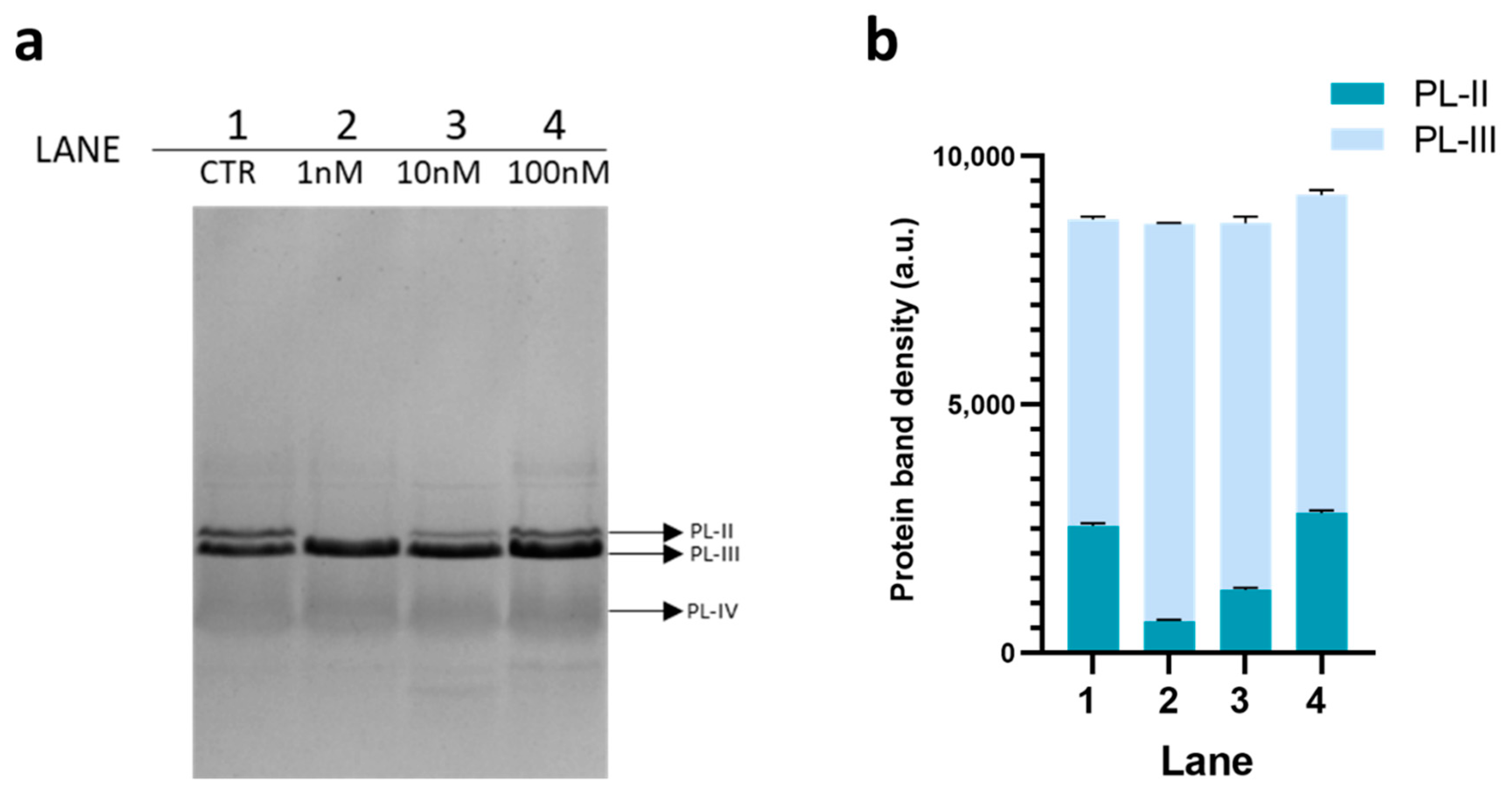
3.1. Electrophoretic Analysis of PL Proteins
3.2. Analysis of PL-DNA Interaction by EMSA
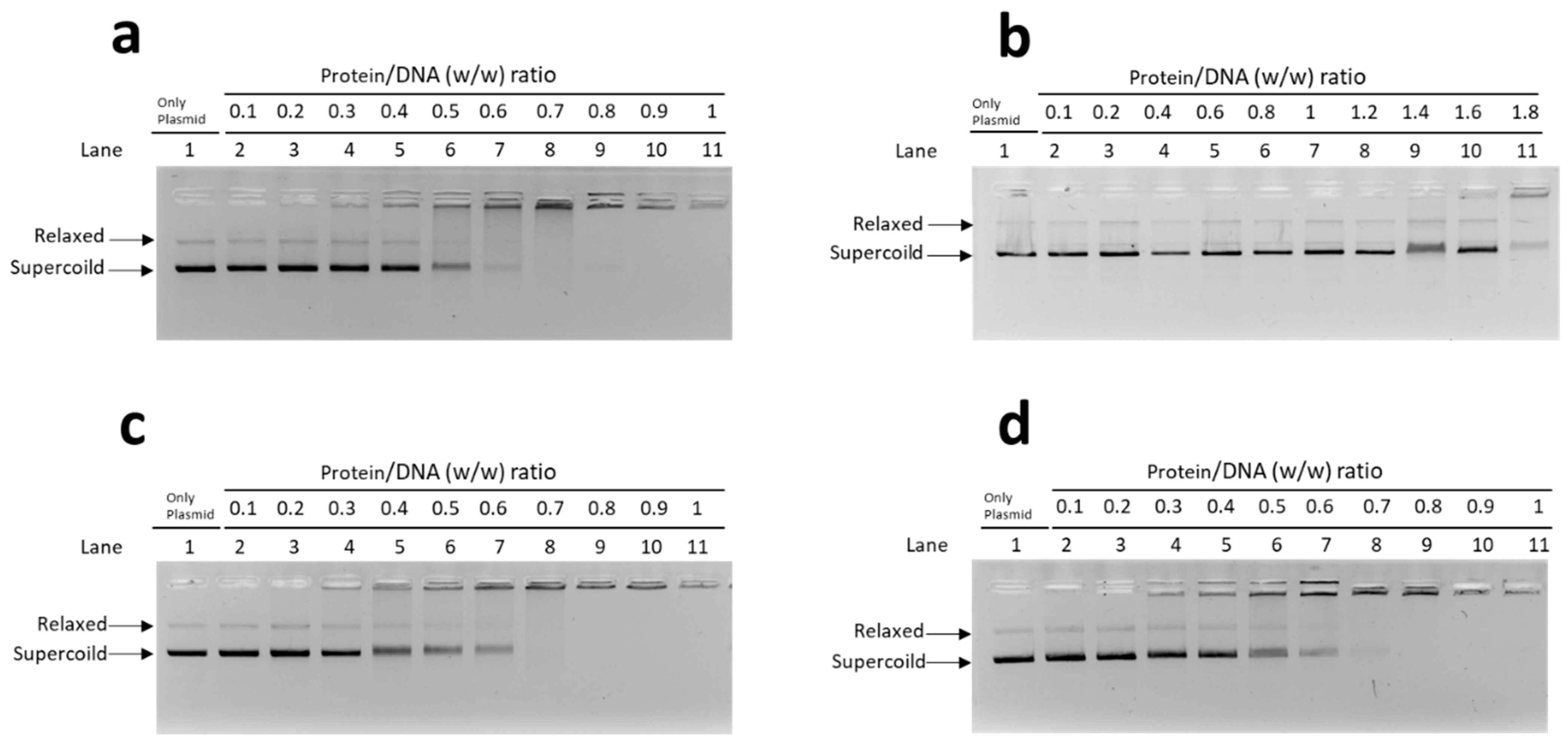
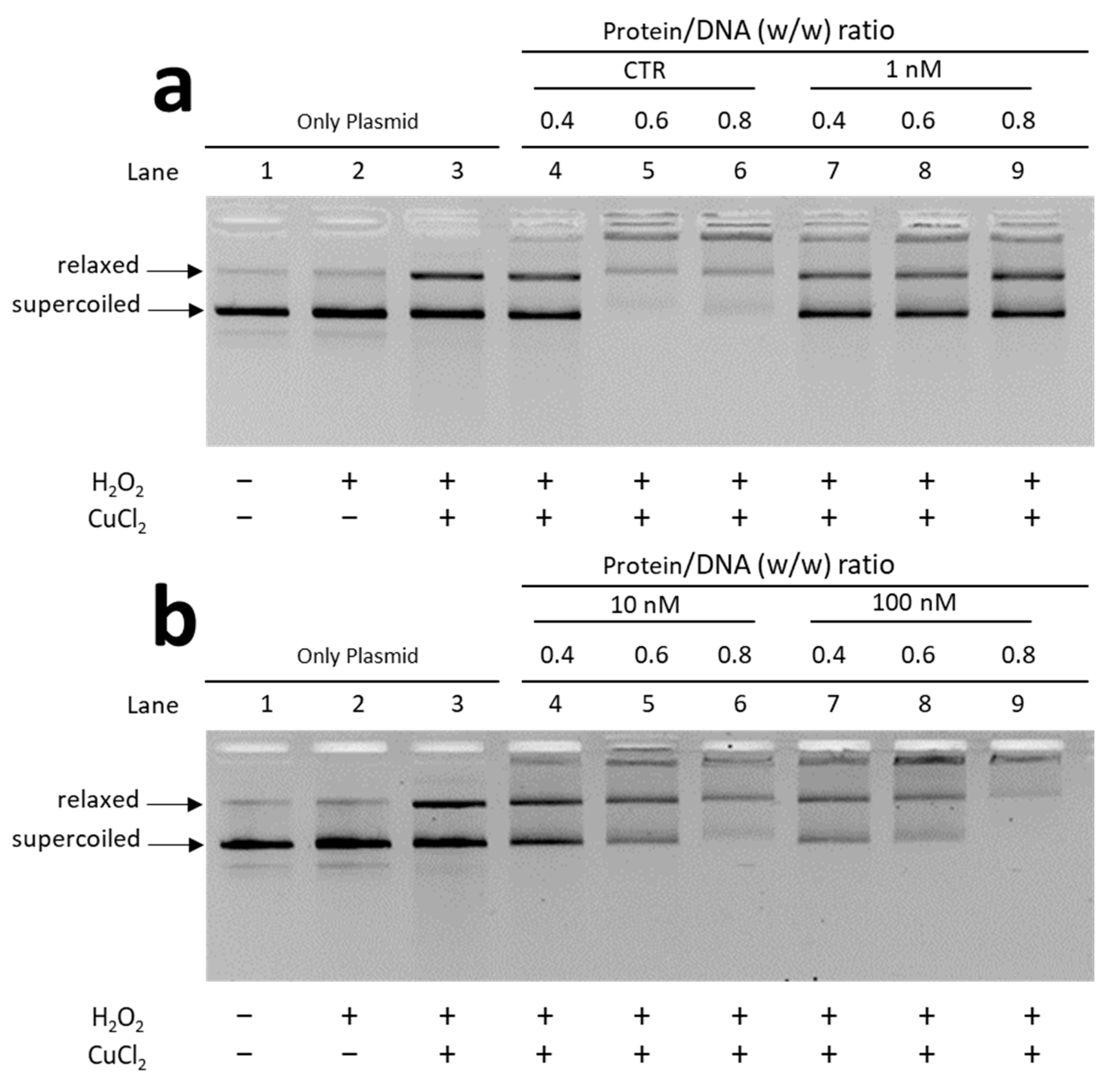
3.2.1. EMSA with Plasmid DNA
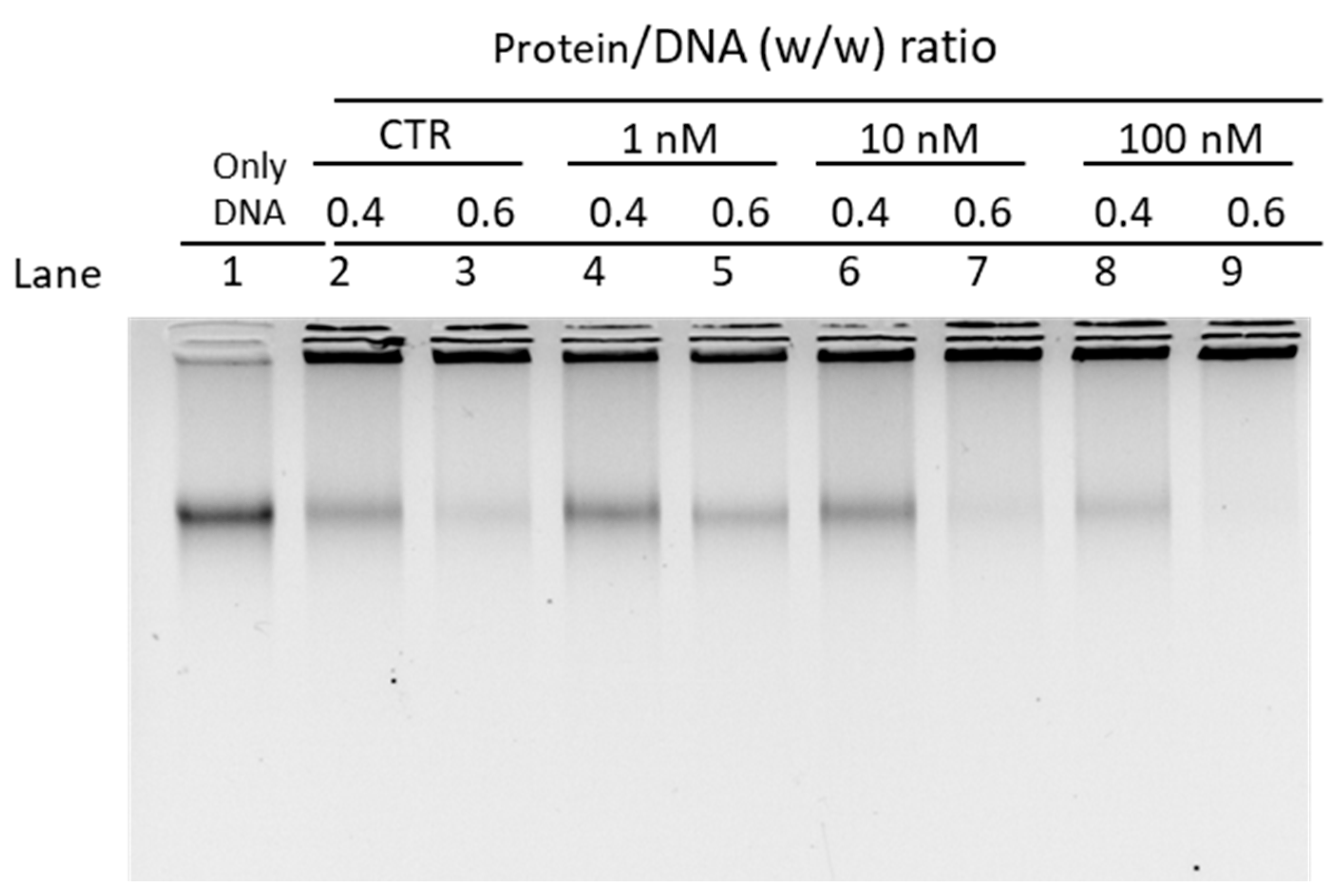
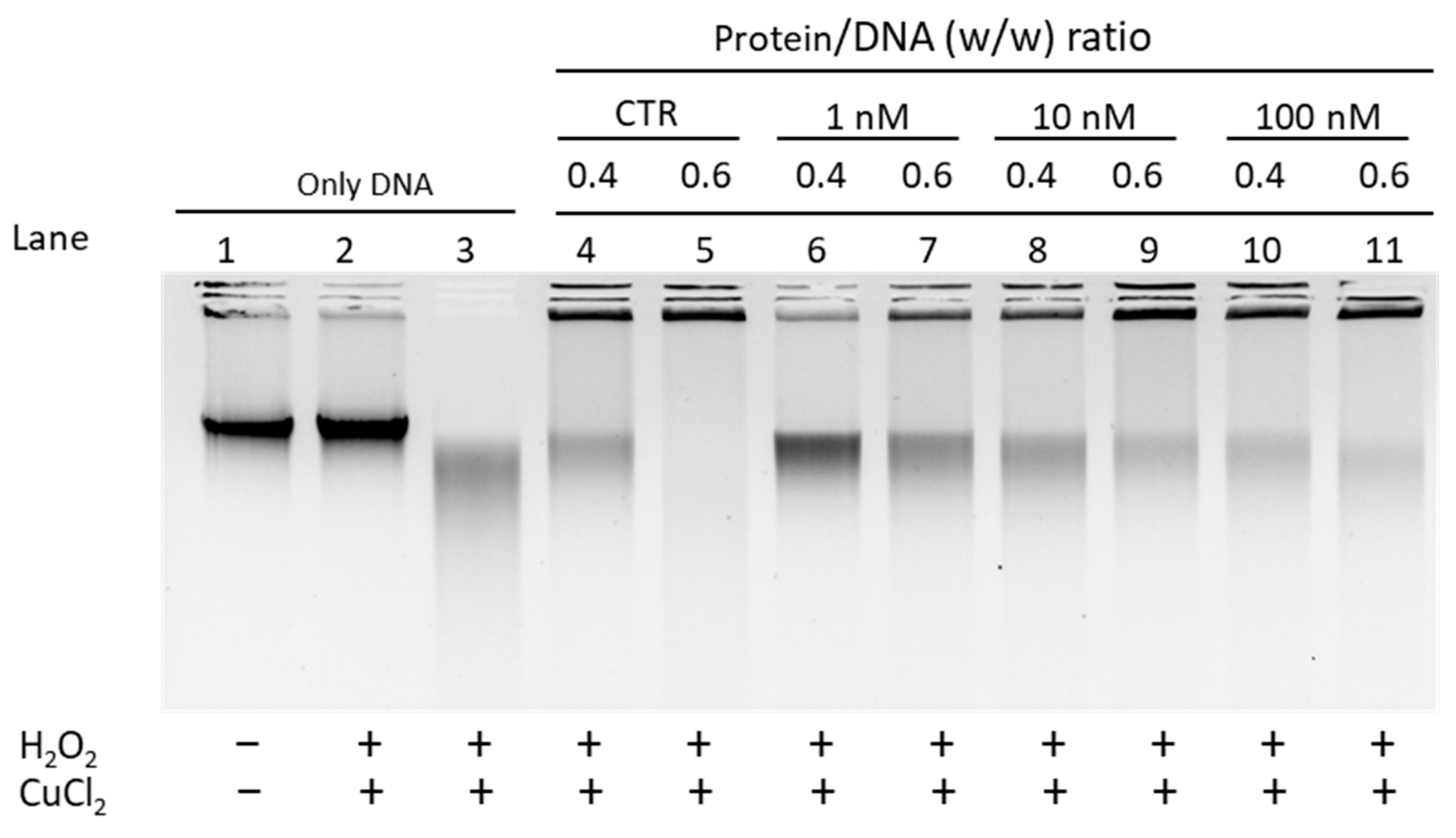
3.2.2. EMSA with Sperm DNA
3.3. Analysis of the Ability of PLs to Protect DNA from Oxidative Damage
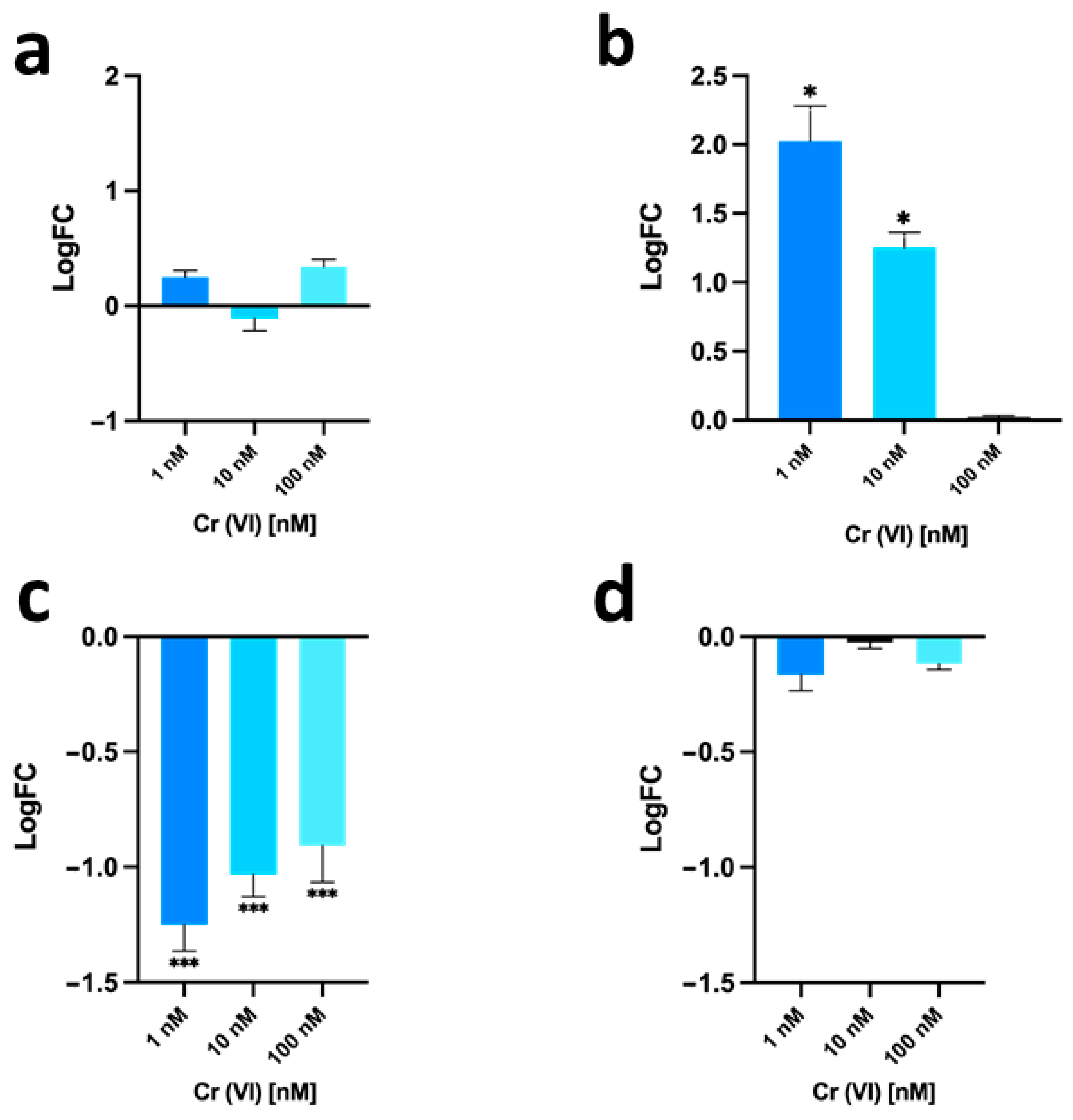
3.4. Gonadal mt10, hsp70 and PL-Proteins Genes Expression
3.5. Evaluation of Sperm Motility
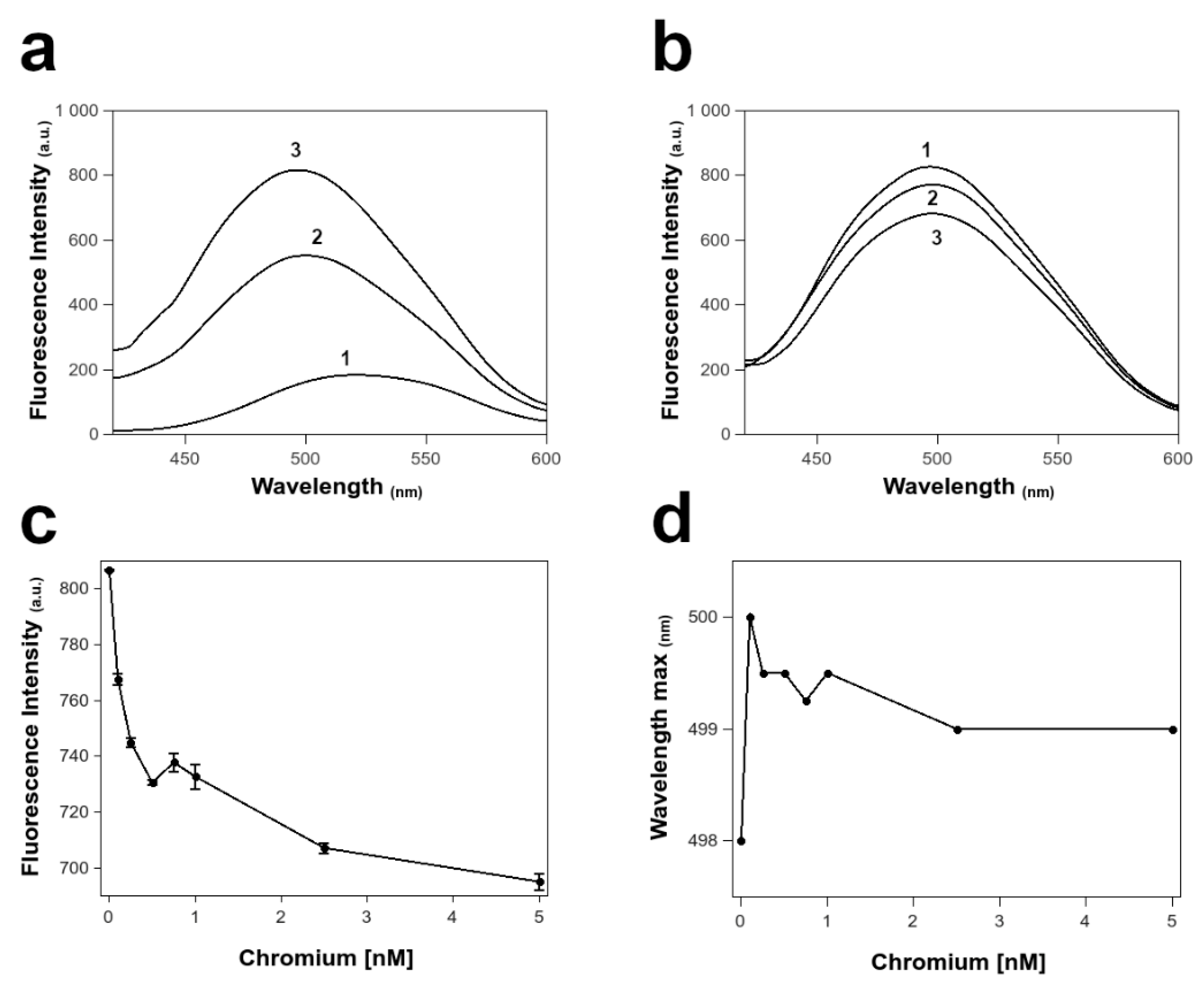
3.6. Fluorescence Spectroscopy of PL Proteins in ANS
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| APS | Ammonium persulfate |
| ANS | 8-anilino-1-naphthalenesulfonic acid |
| ASW | Artificial sea water |
| AU-PAGE | Acetic acid-urea polyacrylamide gel electrophoresis |
| SDS-PAGE | Sodium dodecyl sulphate–polyacrylamide gel electrophoresis |
| EDTA | Ethylenediaminetetraacetic acid |
| EMSA | Electrophoretic mobility shift assay |
| PCA | Perchloric acid |
| PL | Protamine-like |
| RT-qPCR | Reverse transcript quantitative polymerase chain reaction |
| C. variopedatus | Chaetopterus variopedatus |
| ROS | reactive oxygen species |
References
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2014, 7, 60–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.; Bharagava, R.N. Toxic and genotoxic effects of hexavalent chromium in environment and its bioremediation strategies. J. Environ. Sci. Health Part C 2016, 34, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Shock, E.L.; Sassani, D.C.; Willis, M.; Sverjensky, D.A. Inorganic species in geologic fluids: Correlations among standard molal thermodynamic properties of aqueous ions and hydroxide complexes. Geochim. Cosmochim. Acta 1997, 61, 907–950. [Google Scholar] [CrossRef]
- Dabizha, A.; Kersten, M. Exothermic adsorption of chromate by goethite. Appl. Geochem. 2020, 123, 104785. [Google Scholar] [CrossRef]
- Velma, V.; Tchounwou, P.B. Hexavalent chromium-induced multiple biomarker responses in liver and kidney of goldfish, Carassius auratus. Environ. Toxicol. 2011, 26, 649–656. [Google Scholar] [CrossRef] [Green Version]
- DesMarias, T.L.; Costa, M. Mechanisms of chromium-induced toxicity. Curr. Opin. Toxicol. 2019, 14, 1–7. [Google Scholar] [CrossRef]
- Shi, X.; Mao, Y.; Knapton, A.D.; Ding, M.; Rojanasakul, Y.; Gannett, P.M.; Dalal, N.; Liu, K. Reaction of Cr(VI) with ascorbate and hydrogen peroxide generates hydroxyl radicals and causes DNA damage: Role of a Cr(IV)-mediated Fenton-like reaction. Carcinogenesis 1994, 15, 2475–2478. [Google Scholar] [CrossRef]
- De Mattia, G.; Bravi, M.C.; Laurenti, O.; De Luca, O.; Palmeri, A.; Sabatucci, A.; Mendico, G.; Ghiselli, A. Impairment of cell and plasma redox state in subjects professionally exposed to chromium. Am. J. Ind. Med. 2004, 46, 120–125. [Google Scholar] [CrossRef]
- Pereira, S.C.; Oliveira, P.F.; Oliveira, S.R.; Pereira, M.D.L.; Alves, M.G. Impact of Environmental and Lifestyle Use of Chromium on Male Fertility: Focus on Antioxidant Activity and Oxidative Stress. Antioxidants 2021, 10, 1365. [Google Scholar] [CrossRef]
- Zhen, L.; Wang, L.; Fu, J.; Li, Y.; Zhao, N.; Li, X. Hexavalent chromium affects sperm motility by influencing protein tyrosine phosphorylation in the midpiece of boar spermatozoa. Reprod. Toxicol. 2016, 59, 66–79. [Google Scholar] [CrossRef]
- Das, J.; Kang, M.-H.; Kim, E.; Kwon, D.-N.; Choi, Y.-J.; Kim, J.-H. Hexavalent chromium induces apoptosis in male somatic and spermatogonial stem cells via redox imbalance. Sci. Rep. 2015, 5, 13921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhuri, D.; Narayan, R.; Saxena, D. Effect of lead and chromium on nucleic acid and protein synthesis during sperm-zona binding in mice. Toxicol. Vitr. 2001, 15, 605–613. [Google Scholar] [CrossRef]
- Elderfield, H. Chromium speciation in sea water. Earth Planet. Sci. Lett. 1970, 9, 10–16. [Google Scholar] [CrossRef]
- Grimaud, D.; Michard, G. Concentration du chrome dans deux profils de l'ocean pacifique. Mar. Chem. 1974, 2, 229–237. [Google Scholar] [CrossRef]
- Cranston, R.E.; Murray, J.W. Chromium species in the Columbia River and estuary1. Limnol. Oceanogr. 1980, 25, 1104–1112. [Google Scholar] [CrossRef]
- Jeandel, C.; Minster, J.-F. Isotope dilution measurement of inorganic chromium(III) and total chromium in seawater. Mar. Chem. 1984, 14, 347–364. [Google Scholar] [CrossRef]
- Connelly, D.P.; Statham, P.J.; Knap, A.H. Seasonal changes in speciation of dissolved chromium in the surface Sargasso Sea. Deep Sea Res. Part I Oceanogr. Res. Pap. 2006, 53, 1975–1988. [Google Scholar] [CrossRef]
- Eliopoulos, I.-P.; Eliopoulos, G.; Economou-Eliopoulos, M. The Cr(VI) Stability in Contaminated Coastal Groundwater: Salinity as a Driving Force. Minerals 2021, 11, 160. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, Y.; Huang, B. The acute toxicity effects of hexavalent chromium in antioxidant system and gonad development to male clam Geloina coaxans. Eur. Zool. J. 2020, 87, 325–335. [Google Scholar] [CrossRef]
- Chen, H.; Cao, J.; Li, L.; Wu, X.; Bi, R.; Klerks, P.; Xie, L. Maternal transfer and reproductive effects of Cr(VI) in Japanese medaka (Oryzias latipes) under acute and chronic exposures. Aquat. Toxicol. 2016, 171, 59–68. [Google Scholar] [CrossRef]
- Li, L.; Chen, H.; Bi, R.; Xie, L. Bioaccumulation, subcellular distribution, and acute effects of chromium in Japanese medaka (Oryzias latipes). Environ. Toxicol. Chem. 2015, 34, 2611–2617. [Google Scholar] [CrossRef] [PubMed]
- Pagano, G.; Esposito, A.; Bove, P.; de Angelis, M.; Rota, A.; Giordano, G.G. The effects of hexavalent and trivalent chromium on fertilization and development in sea urchins. Environ. Res. 1983, 30, 442–452. [Google Scholar] [CrossRef]
- Viarengo, A.; Canesi, L. Mussels as biological indicators of pollution. Aquaculture 1991, 94, 225–243. [Google Scholar] [CrossRef]
- Vassalli, Q.A.; Caccavale, F.; Avagnano, S.; Murolo, A.; Guerriero, G.; Fucci, L.; Ausió, J.; Piscopo, M. New Insights into Protamine-Like Component Organization in Mytilus galloprovincialis' Sperm Chromatin. DNA Cell Biol. 2015, 34, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Lettieri, G.; Mollo, V.; Ambrosino, A.; Caccavale, F.; Troisi, J.; Febbraio, F.; Piscopo, M. Molecular effects of copper on the reproductive system of Mytilus galloprovincialis. Mol. Reprod. Dev. 2019, 86, 1357–1368. [Google Scholar] [CrossRef]
- Lettieri, G.; Maione, M.; Ranauda, M.A.; Mele, E.; Piscopo, M. Molecular effects on spermatozoa of Mytilus galloprovincialis exposed to hyposaline conditions. Mol. Reprod. Dev. 2019, 86, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Piscopo, M. Seasonal dependence of cadmium molecular effects on Mytilus galloprovincialis (Lamarck, 1819) protamine-like protein properties. Mol. Reprod. Dev. 2019, 86, 1418–1429. [Google Scholar] [CrossRef] [PubMed]
- Piscopo, M.; Notariale, R.; Rabbito, D.; Ausió, J.; Olanrewaju, O.S.; Guerriero, G. Mytilus galloprovincialis (Lamarck, 1819) spermatozoa: hsp70 expression and protamine-like protein property studies. Environ. Sci. Pollut. Res. Int. 2018, 25, 12957–12966. [Google Scholar] [CrossRef]
- De Guglielmo, V.; Puoti, R.; Notariale, R.; Maresca, V.; Ausió, J.; Troisi, J.; Verrillo, M.; Basile, A.; Febbraio, F.; Piscopo, M. Alterations in the properties of sperm protamine-like II protein after exposure of Mytilus galloprovincialis (Lamarck 1819) to sub-toxic doses of cadmium. Ecotoxicol. Environ. Saf. 2019, 169, 600–606. [Google Scholar] [CrossRef]
- Lettieri, G.; Notariale, R.; Ambrosino, A.; Di Bonito, A.; Giarra, A.; Trifuoggi, M.; Manna, C.; Piscopo, M. Spermatozoa Transcriptional Response and Alterations in PL Proteins Properties after Exposure of Mytilus galloprovincialis to Mercury. Int. J. Mol. Sci. 2021, 22, 1618. [Google Scholar] [CrossRef]
- Lettieri, G.; Notariale, R.; Carusone, N.; Giarra, A.; Trifuoggi, M.; Manna, C.; Piscopo, M. New Insights into Alterations in PL Proteins Affecting Their Binding to DNA after Exposure of Mytilus galloprovincialis to Mercury—A Possible Risk to Sperm Chromatin Structure? Int. J. Mol. Sci. 2021, 22, 5893. [Google Scholar] [CrossRef] [PubMed]
- Piscopo, M.; Trifuoggi, M.; Notariale, R.; Labar, S.; Troisi, J.; Giarra, A.; Rabbito, D.; Puoti, R.; Brundo, M.V.; Basile, A.; et al. Protamine-like proteins analyses as emerging biotechnique for cadmium impact assessment on male mollusk Mytilus galloprovincialis (Lamarck 1819). Acta Biochim. Pol. 2018, 65, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Yılmaz, A.B.; Yanar, A.; Alkan, E.N. Review of heavy metal accumulation on aquatic environment in Northern East Mediterrenean Sea part I: Some essential metals. Rev. Environ. Health 2017, 32, 119–163. [Google Scholar] [CrossRef] [PubMed]
- Lettieri, G.; Carusone, N.; Notariale, R.; Prisco, M.; Ambrosino, A.; Perrella, S.; Manna, C.; Piscopo, M. Morphological, Gene, and Hormonal Changes in Gonads and In-Creased Micrococcal Nuclease Accessibility of Sperm Chromatin Induced by Mercury. Biomolecules 2022, 12, 87. [Google Scholar] [CrossRef]
- Notariale, R.; Basile, A.; Montana, E.; Romano, N.C.; Cacciapuoti, M.G.; Aliberti, F.; Gesuele, R.; De Ruberto, F.; Sorbo, S.; Tenore, G.C.; et al. Protamine-like proteins have bactericidal activity. The first evidence in Mytilus galloprovincialis. Acta Biochim. Pol. 2018, 65, 585–594. [Google Scholar] [CrossRef]
- Carbone, A.; Fioretti, F.M.; Fucci, L.; Ausió, J.; Piscopo, M. High efficiency method to obtain supercoiled DNA with a commercial plasmid purification kit. Acta Biochim. Pol. 2012, 59, 275–278. [Google Scholar] [CrossRef] [Green Version]
- Piscopo, M.; Trifuoggi, M.; Scarano, C.; Gori, C.; Giarra, A.; Febbraio, F. Relevance of arginine residues in Cu(II)-induced DNA breakage and Proteinase K resistance of H1 histones. Sci. Rep. 2018, 8, 7414. [Google Scholar] [CrossRef] [Green Version]
- Piscopo, M.; Tenore, G.C.; Notariale, R.; Maresca, V.; Maisto, M.; De Ruberto, F.; Heydari, M.; Sorbo, S.; Basile, A. Antimicrobial and antioxidant activity of proteins from Feijoa sellowiana Berg. fruit before and after in vitro gastrointestinal digestion. Nat. Prod. Res. 2020, 34, 2607–2611. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Di Matteo, O.; Langellotti, A.L.; Masullo, P.; Sansone, G. Cryopreservation of the Mediterranean mussel (Mytilus galloprovincialis) spermatozoa. Cryobiology 2009, 58, 145–150. [Google Scholar] [CrossRef]
- Boni, R.; Gallo, A.; Montanino, M.; Macina, A.; Tosti, E. Dynamic changes in the sperm quality of Mytilus galloprovincialis under continuous thermal stress. Mol. Reprod. Dev. 2016, 83, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Tosti, E. Reprotoxicity of the Antifoulant Chlorothalonil in Ascidians: An Ecological Risk Assessment. PLoS ONE 2015, 10, e0123074. [Google Scholar] [CrossRef] [PubMed]
- Wildt, D.E.; Comizzoli, P.; Pukazhenthi, B.; Songsasen, N. Lessons from biodiversity-the value of nontraditional species to advance reproductive science, conservation, and human health. Mol. Reprod. Dev. 2010, 77, 397–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maresca, V.; Fusaro, L.; Sorbo, S.; Siciliano, A.; Loppi, S.; Paoli, L.; Monaci, F.; Karam, E.A.; Piscopo, M.; Guida, M.; et al. Functional and structural biomarkers to monitor heavy metal pollution of one of the most contaminated freshwater sites in Southern Europe. Ecotoxicol. Environ. Saf. 2018, 163, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Basile, A.; Loppi, S.; Piscopo, M.; Paoli, L.; Vannini, A.; Monaci, F.; Sorbo, S.; Lentini, M.; Esposito, S. The biological response chain to pollution: A case study from the “Italian Triangle of Death” assessed with the liverwort Lunularia cruciata. Environ. Sci. Pollut. Res. Int. 2017, 24, 26185–26193. [Google Scholar] [CrossRef]
- Tumolo, M.; Ancona, V.; De Paola, D.; Losacco, D.; Campanale, C.; Massarelli, C.; Uricchio, V.F. Chromium Pollution in European Water, Sources, Health Risk, and Remediation Strategies: An Overview. Int. J. Environ. Res. Public Health 2020, 17, 5438. [Google Scholar] [CrossRef]
- Velma, V.; Vutukuru, S.S.; Tchounwou, P.B. Ecotoxicology of Hexavalent Chromium in Freshwater Fish: A Critical Review. Rev. Environ. Health 2009, 24, 129–146. [Google Scholar] [CrossRef] [Green Version]
- Bakshi, A.; Panigrahi, A.K. A comprehensive review on chromium induced alterations in fresh water fishes. Toxicol. Rep. 2018, 5, 440–447. [Google Scholar] [CrossRef]
- Shelake, R.M.; Ito, Y.; Masumoto, J.; Morita, E.H.; Hayashi, H. A novel mechanism of “metal gel-shift” by histidine-rich Ni2+-binding Hpn protein from Helicobacter pylori strain SS1. PLoS ONE 2017, 12, e0172182. [Google Scholar] [CrossRef] [Green Version]
- Piscopo, M.; Conte, M.; Di Paola, F.; Conforti, S.; Rana, G.; De Petrocellis, L.; Fucci, L.; Geraci, G. Relevance of Arginines in the Mode of Binding of H1 Histones to DNA. DNA Cell Biol. 2010, 29, 339–347. [Google Scholar] [CrossRef]
- Fioretti, F.M.; Febbraio, F.; Carbone, A.; Branno, M.; Carratore, V.; Fucci, L.; Ausió, J.; Piscopo, M. A Sperm Nuclear Basic Protein from the Sperm of the Marine Worm Chaetopterus variopedatus with Sequence Similarity to the Arginine-Rich C-Termini of Chordate Protamine-Likes. DNA Cell Biol. 2012, 31, 1392–1402. [Google Scholar] [CrossRef] [PubMed]
- Salvati, D.; Conforti, S.; Conte, M.; Matassa, D.S.; Fucci, L.; Piscopo, M. Self-association of Chaetopterus variopedatus sperm histone H1-like. Relevance of arginine content and possible physiological role. Acta Biochim. Pol. 2008, 55, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Lettieri, G.; D’Agostino, G.; Mele, E.; Cardito, C.; Esposito, R.; Cimmino, A.; Giarra, A.; Trifuoggi, M.; Raimondo, S.; Notari, T.; et al. Discovery of the Involvement in DNA Oxidative Damage of Human Sperm Nuclear Basic Proteins of Healthy Young Men Living in Polluted Areas. Int. J. Mol. Sci. 2020, 21, 4198. [Google Scholar] [CrossRef] [PubMed]
- Lettieri, G.; Marra, F.; Moriello, C.; Prisco, M.; Notari, T.; Trifuoggi, M.; Giarra, A.; Bosco, L.; Montano, L.; Piscopo, M. Molecular Alterations in Spermatozoa of a Family Case Living in the Land of Fires. A First Look at Possible Transgenerational Effects of Pollutants. Int. J. Mol. Sci. 2020, 21, 6710. [Google Scholar] [CrossRef] [PubMed]
- Slavík, J. Anilinonaphthalene sulfonate as a probe of membrane composition and function. Biochim. Biophys. Acta 1982, 694, 1–25. [Google Scholar] [CrossRef]
- Peng, W.; Ding, F.; Peng, Y.-K.; Jiang, Y.-T.; Zhang, L. Binding Patterns and Structure–Affinity Relationships of Food Azo Dyes with Lysozyme: A Multitechnique Approach. J. Agric. Food Chem. 2013, 61, 12415–12428. [Google Scholar] [CrossRef] [PubMed]
- Dondero, F.; Piacentini, L.; Banni, M.; Rebelo, M.; Burlando, B.; Viarengo, A. Quantitative PCR analysis of two molluscan metallothionein genes unveils differential expression and regulation. Gene 2005, 345, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E. Hormesis: Why it is Important to Toxicology and Toxicologists. Environ. Toxicol. Chem. 2008, 27, 1451–1474. [Google Scholar] [CrossRef]
- Piscopo, M.; Ricciardiello, M.; Palumbo, G.; Troisi, J. Selectivity of metal bioaccumulation and its relationship with glutathione S-transferase levels in gonadal and gill tissues of Mytilus galloprovincialis exposed to Ni (II), Cu (II) and Cd (II). Rendiconti Lincei 2016, 27, 737–748. [Google Scholar] [CrossRef] [Green Version]
- Jia, L.; Liu, Z.; Chen, W.; Ye, Y.; Yu, S.; He, X. Hormesis Effects Induced by Cadmium on Growth and Photosynthetic Performance in a Hyperaccumulator, Lonicera japonica Thunb. J. Plant Growth Regul. 2015, 34, 13–21. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | F-Primer | F-Primer Length | R-Primer | R-Primer Length | Accession Number |
|---|---|---|---|---|---|
| GAPDH | CTGCACCACCAACTGCTT | 18 | TTCTGGGTGGCAGTGATG | 18 | SY171038758-018/019 |
| Hsp70 | CGCGATGCCAAACTAGACAA | 20 | TCACCTGACAAAATGGCTGC | 20 | AY861684 |
| Mt10 | GCCTGCACCTTGTAACTGTAT | 21 | CTGTACACCCTGCTTCACAC | 20 | AY566248 |
| PL-III | CACCCAACAAGAAGGATGCC | 20 | CCTTGCCCTTTTCTTTCCCC | 20 | SY140930274 |
| PL-II/IV | AAGCCCAAGTAGACGTTCCA | 20 | TCCGAGGTGTGATGTGTTGA | 20 | SY140930274 |
| Score | |
|---|---|
| Unexposed | 5 |
| 1 nM Cr(VI) exposed | 1 |
| 10 and 100 nM Cr(VI) exposed | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moriello, C.; Costabile, M.; Spinelli, M.; Amoresano, A.; Palumbo, G.; Febbraio, F.; Piscopo, M. Altered Expression of Protamine-like and Their DNA Binding Induced by Cr(VI): A Possible Risk to Spermatogenesis? Biomolecules 2022, 12, 700. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12050700
Moriello C, Costabile M, Spinelli M, Amoresano A, Palumbo G, Febbraio F, Piscopo M. Altered Expression of Protamine-like and Their DNA Binding Induced by Cr(VI): A Possible Risk to Spermatogenesis? Biomolecules. 2022; 12(5):700. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12050700
Chicago/Turabian StyleMoriello, Claudia, Martina Costabile, Michele Spinelli, Angela Amoresano, Giancarlo Palumbo, Ferdinando Febbraio, and Marina Piscopo. 2022. "Altered Expression of Protamine-like and Their DNA Binding Induced by Cr(VI): A Possible Risk to Spermatogenesis?" Biomolecules 12, no. 5: 700. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12050700