
Targeting Nrf2 with Probiotics and Postbiotics in the Treatment of Periodontitis


1
Department of Periodontology, Institute of Dentistry, University of Turku, 20520 Turku, Finland
2
Department of Biology, Faculty of Science, Ankara University, Ankara 06100, Turkey
3
Department of Periodontology, Faculty of Dentistry, Biruni University, Istanbul 34010, Turkey
*
Authors to whom correspondence should be addressed.
Biomolecules 2022, 12(5), 729; https://0-doi-org.brum.beds.ac.uk/10.3390/biom12050729
Submission received: 12 April 2022
/
Revised: 15 May 2022
/
Accepted: 19 May 2022
/
Published: 22 May 2022
(This article belongs to the Special Issue Role of Nrf2 in Disease: Novel Molecular Mechanisms and Therapeutic Approaches II)
Abstract
:Periodontitis is a destructive disease of the tooth-surrounding tissues. Infection is the etiological cause of the disease, but its extent and severity depend on the immune–inflammatory response of the host. Immune cells use reactive oxygen species to suppress infections, and there is homeostasis between oxidative and antioxidant mechanisms during periodontal health. During periodontitis, however, increased oxidative stress triggers tissue damage, either directly by activating apoptosis and DNA damage or indirectly by activating proteolytic cascades. Periodontal treatment aims to maintain an infection and inflammation-free zone and, in some cases, regenerate lost tissues. Although mechanical disruption of the oral biofilm is an indispensable part of periodontal treatment, adjunctive measures, such as antibiotics or anti-inflammatory medications, are also frequently used, especially in patients with suppressed immune responses. Recent studies have shown that probiotics activate antioxidant mechanisms and can suppress extensive oxidative stress via their ability to activate nuclear factor erythroid 2-related factor 2 (Nrf2). The aim of this narrative review is to describe the essential role of Nrf2 in the maintenance of periodontal health and to propose possible mechanisms to restore the impaired Nrf2 response in periodontitis, with the aid of probiotic and postbiotics.
Periodontitis is a highly prevalent inflammatory disease of an infectious origin, and it is the major cause of tooth loss globally. Increased oxidative stress and diminished antioxidant capacity are associated with periodontal pathogenesis. While conventional periodontal treatment is effective in many individuals, some unresponsive patients can theoretically benefit from adjunctive therapies that may have both direct effects on biofilm control and indirect effects, such as modulation of the host response. Regulation of antioxidant capacity with adjunctive therapy models is an emerging topic in various medical fields, particularly via nuclear factor erythroid 2-related factor 2 (Nrf2) activation. Nrf2 is a transcription factor that has been the subject of numerous studies, revealing the mechanism behind the oxidative-stress-induced cellular damage. This narrative review aimed to describe the Nrf2 activating abilities of postbiotics and address the potential benefit of postbiotic use as an adjunct to conventional periodontal treatment.
1. Periodontal Health and Disease
The periodontium consists of four specialized tissues that work together harmoniously: Alveolar process (bone that forms the walls of the tooth socket), periodontal ligament (connective tissue that joins the tooth to the alveolar bone), cementum (thin hard tissue that lines the root surface where the fibers of the periodontal ligament are inserted), and gingiva (oral mucosa that abuts the tooth and seals the underlying tissues). Periodontal health is defined by the absence of clinical signs of gingival inflammation and alveolar bone loss [1]. However, even in clinically healthy gingiva, periodontal tissues resist constant microbial challenges through innate and adaptive defense mechanisms [2]. In susceptible individuals, the balance between the subgingival microflora and the host response is disturbed, resulting in uncontrolled, persistent inflammation and consequent loss of alveolar bone and periodontium, namely periodontitis [3,4].
Periodontitis is an infectious inflammatory disease. Clinical signs of periodontitis include gingival inflammatory changes, such as redness, swelling, bleeding, pocket formation and/or gingival recession, bone loss, increased tooth mobility, pathologic tooth migration, suppuration, and abscess formation, and it can lead to tooth loss (Figure 1). In addition to oral consequences, periodontitis is also associated with various systemic diseases and conditions, such as hyperglycemia, chronic pulmonary diseases, cardiovascular disease, and obstetric complications, which may be related to periodontitis and the systemic consequences of chronic, low-grade inflammation [5].
Periodontal inflammation begins in response to oral pathogens by recruiting polymorphonuclear leukocytes (PMNs), which form a barrier to prevent the migration of microbes into the deep tissues [6]. PMNs attempt to clear out pathogens with their distinct functions, such as chemotaxis, degranulation, extracellular trap formation, phagocytosis, and release of defensins, antimicrobial peptides and reactive oxygen species (ROS) [7]. As the lesion progresses, a dense and more complex inflammatory infiltrate forms, including macrophages, lymphocytes, and plasma cells [8]. The ecological changes caused by bacterial and host-derived products and the manipulation of the immune system by some pathogens such as Porphyromonas gingivalis lead to a dysbiotic relationship between subgingival bacteria and the host, resulting in chronic, excessive, and dysregulated inflammation and disruption of homeostasis [9].
2. Oxidative Stress in Pathogenesis of Periodontal Disease
Reactive oxygen species are crucial for host tissue metabolism, e.g., cell signaling, proliferation, and renewal. However, excessive accumulation of ROS in tissues causes damage to lipids, carbohydrates, proteins, and deoxyribonucleic acids, eventually triggering a chronic inflammatory process. Various autoimmune or inflammatory oral conditions have been associated with cellular damage as a result of increased oxidative stress, such as lichen planus, lichenoid reactions, leukoplakia, pemphigus vulgaris, recurrent aphthous ulcers, oral squamous cell carcinoma, and periodontitis [10,11,12,13,14]. Current evidence suggests a close relationship between the pathogenesis of periodontitis and oxidative stress caused by oxygen-derived free radicals. Biomarkers associated with oxidative stress are elevated in periodontitis compared to a healthy periodontium [14,15]. PMNs play a central role in this mechanism as first responders. These cells migrate into periodontal tissues by responding to mediators, such as interleukin (IL)-8 or interferon (INF)-α, and releasing large amounts of ROS, triggering a cytotoxic respiratory burst catalyzed by nicotinamide adenine dinucleotide phosphate (NADPH) oxidase [16]. Essentially, ROS are protective in nature; however, their overexpression is cytotoxic to host cells [15]. Indeed, periodontitis patients demonstrate a hyper-responsive PMN phenotype, and their blood plasma contains a considerable number of pro-inflammatory cytokines that stimulate the production of ROS by neutrophils [16].
In periodontal tissues, PMNs generate ROS by responding to Fcγ receptors, but also to nonopsonized pathogens, such as Fusobacterium nucleatum and P. gingivalis [17,18]. The release of superoxide by neutrophils in the presence or absence of these nonopsonized pathogens suggests that extracellular ROS production is constitutive in periodontitis, whereas intracellular ROS are produced as a secondary response to periodontal inflammation [18].
The oxidative stress caused by PMNs damages not only the anaerobic periodontal pathogens but also the neutrophils themselves and plays an important role in periodontal destruction [19]. Tissue damage caused by ROS can occur in several forms: Lipid peroxidation, DNA damage, protein damage, oxidation of protective enzymes, and stimulation of proinflammatory cytokines [20]. Active PMNs can severely injure gingival epithelial cells, while their oxidants and proteases together damage periodontal tissues [21]. In addition, bone remodeling gets disrupted due to the increase in the receptor activator of nuclear factor kappa B ligand (RANKL)/osteoprotegerin ratio due to ROS, leading to osteoclastogenesis and alveolar bone loss [15,22] (Figure 2). These hyper-active and hyper-reactive PMN phenotypes are thought to be primarily associated with disrupted redox potential, particularly impaired regulation of antioxidant mechanisms by nuclear factor erythroid-2-related factor 2 (Nrf2) [23]. Nrf2 is a basic cap-n-collar leucine zipper transcription factor that is crucial in maintaining the periodontal tissue homeostasis by regulating anti-oxidative response element genes [24].
3. Anti-Oxidative Mechanisms in Periodontal Tissues
In a healthy state, the activities of ROS are balanced by various antioxidant mechanisms. When the antioxidant capacity is overwhelmed, oxidative stress occurs. This shift in the balance between ROS and antioxidants can be caused either by increased production and activity of ROS or by a disturbance in antioxidant defenses [17]. Antioxidant concentrations in the GCF of periodontitis patients have been found to be reduced compared to healthy individuals [25,26].
Nrf2 and its inhibitor Keap1 are important regulators of antioxidant defense mechanisms. Nrf2 controls the gene transcription of many antioxidant enzymes such as catalase, superoxide dismutase, NADPH, heme oxygenase, and glutathione peroxidase, which have been widely studied as antioxidants in periodontitis [27,28]. The Nrf2 pathway is suppressed in severe periodontitis, which is responsible for the elevated PMN levels and associated high oxidative damage in periodontitis. Nrf2-/- mice exhibit low catalase production by PMNs and increased periodontal tissue breakdown, suggesting a central role in periodontal pathogenesis [28]. This is also consistent with the promotion of RANKL-induced activation of kinases and nuclear factor activated T cells in Nrf2 deficiency, leading to osteoclast differentiation and consequent severe alveolar bone loss [28,29]. Nuclear Nrf2 is more frequently present and heme oxygenase-1 expression and luciferase activity are increased in experimental periodontitis when compared to healthy individuals [30]. Moreover, apoptosis of periodontal ligament stem cells, which are highly efficient in periodontal repair, is increased by oxidative stress, which also induces the antioxidant effect of Nrf2. Overexpression of Nrf2 increases antioxidants, such as NADPH-quinone oxidoreductase 1, heme oxygenase, and γ-glutamylcysteine synthetase, and induces cell proliferation while limiting apoptosis [31]. This suggests the importance of Nrf2 in maintaining the integrity of periodontal ligament cells and the antioxidant capacity of these cells. However, the Nrf2 response in gingival keratinocytes does not appear to be induced by P. gingivalis lipopolysaccharide, although it is stimulated by nicotine-induced oxidative stress [32]. Therefore, other antioxidant pathways besides Nrf2, such as forkhead box-O or sirtuins, should be considered in the pathogenesis of periodontitis, although there are still many unknowns in oxidative damage and antioxidant mechanisms in periodontal tissues [33,34,35,36].
Anti-oxidative mechanisms in peri-implant diseases also need to be briefly mentioned, considering that peri-implantitis is an increasing oral health problem. Peri-implant diseases share many common threads with periodontal diseases, but also exhibit different histopathologic characteristics due to anatomical and functional differences [37]. A scarce number of studies demonstrated increased oxidative stress specified with myeloperoxidase levels and nitric oxide metabolism in peri-implant disease, resembling inflammation in natural teeth [38,39,40]. To the best of our knowledge, only one study evaluated Nrf2 levels in dental peri-implantitis, suggesting an increase in Parkinsonism-associated deglycase 7, which regulates Nrf2, and increased oxidative stress measured with 8-hydroxy-deoxyguanosine, although Nrf2 expression was not altered in peri-implantitis mucosa [41].
4. Probiotics and Postbiotics
According to the Food and Agriculture Organization (FAO), “probiotics are live microorganisms which when administered in adequate amounts confer a health benefit on the host”. According to this consensus definition, probiotics should be administered or consumed alive. Apart from their use for oral health, probiotics may have many beneficial properties, such as competing with pathogens for adhesion, producing antimicrobial components, contributing to immunomodulation, and strengthening mucosal barrier function [42].
Postbiotics are all components produced and released by microorganisms as a result of metabolic activities. The components of postbiotics are intracellular or extracellular soluble byproducts of metabolism, as well as cell wall and cell membrane components, released as a result of bacterial cell lysis [43] (Figure 3). The term postbiotics is still very current, but its use beyond probiotics has led to its use in many different areas, such as the food industry, human health, nutrition, functional foods, and pharmaceuticals [44]. Postbiotics are cell-produced metabolic cocktails and do not contain microorganisms, making them more stable and user-friendly than probiotics. Unlike probiotics, postbiotics have the advantage of being more stable and longer lasting, posing fewer risks than live probiotics in immunocompromised individuals, and not spreading antibiotic resistance factors. Finally, publications investigating the potential immunomodulatory, antioxidant, antimicrobial, antibiofilm, antioxidant, and anticancer effects of postbiotics are increasing exponentially in the literature [45].
5. Antioxidant Abilities of Probiotics and Postbiotics
Oxidative stress damages macromolecules, such as proteins and nucleic acids, by increasing the level of oxygen radicals in the cell. Organisms have enzymes that respond to ROS by neutralizing them and preventing the damage they cause. Superoxide dismutase (SOD), glutathione peroxidase (GPx), glutathione reductase (GR), and non-enzymatic antioxidants are the main responses that protect the organism from oxidative stress [46]. The use of antioxidants from biological sources is becoming increasingly popular as supportive antioxidants [47].
Probiotics can exert their potential antioxidant effects in several ways. One of these ways is that some lactic acid bacteria show the ability to chelate metal ions, although the mechanism of action is unknown [22,47]. Redox-active metals, such as copper (Cu), iron (Fe), and cobalt (Co), which lead to redox reactions, stimulate the formation of ROS [48]. Probiotics may also have their own antioxidant enzymes and promote the production of antioxidant enzymes in the host. For example, Kullisaar et al. [49] demonstrated the presence of the enzyme superoxide dismutase (SOD) in Lactobacillus fermentum. Bacterial SODs offer promising results in the treatment of Crohn’s disease in studies using mouse models [50]. The products produced by probiotics as a result of their metabolic activities, such as short-chain fatty acids, vitamins, and folic acid, may also exhibit antioxidant activity on their own. The antioxidants of probiotics and postbiotics are summarized in Table 1. Although there are many studies in the literature on the antioxidant capacity of probiotics, a general overview of their antioxidant properties is provided here.
6. Probiotics and Nrf2 Activation
It is also known that probiotics can activate various pathways, such as Nrf2-Keap1-ARE, NFκB, MAPK, and PKC, which are responsible for the response to oxidative stress in the host [47]. In the light of all this information, it is clear that probiotics have the potential to exert antioxidant activity directly or indirectly. The Nrf2 activating capabilities of probiotics are summarized in Table 2.
7. Postbiotics and Nrf2 Activation
In this section, we review the effects of probiotic components on Nrf2 activation in different systems. Although the concept of postbiotics has not yet come to the forefront in many of the studies mentioned, we have considered cellular components and byproducts of probiotics as postbiotics because they now fall within the definition of postbiotics.
7.1. Exopolysaccharides (EPS)
Many probiotic bacteria are capable of producing exopolysaccharides. The best-known benefits of exopolysaccharides to the host are in enhancing immune modulation and barrier function [69]. Some EPS extracted from probiotics have the ability to scavenge radicals directly and chelate metals [70,71]. For example, exopolysaccharides from strain L. rhamnosus GG, one of the best-studied probiotic strains, reduce H2O2-induced oxidative damage in intestinal epithelial cells and enhance the antioxidant response by activating Kelch-like ECH-associated protein 1 (Keap1)/Nrf2 signaling pathways [72].
7.2. Short-Chain Fatty Acids (SCFA)
Short-chain fatty acids, such as propionate, acetate, and butyrate, are functional for epithelial integrity, barrier function, and modulation of gut microbiota composition and immune response [73]. Butyric acid is one of the best-studied short-chain fatty acids and can be defined as a postbiotic that can be produced by probiotic bacteria [74]. Butyrate, for example, can trigger the antioxidant glutathione, leading to an oxidative stress response in colon cells [75]. Butyrate can also induce Nrf2 by histone acetylation in the liver [76]. Although butyrate has beneficial effects primarily in the gastrointestinal tract, butyrate produced by butyrogenic bacteria in the oral cavity plays a key role in the progression of periodontitis. Butyrate also promotes a proinflammatory response in fibroblasts of the periodontal ligament. Therefore, the role of butyrate in oral health can be considered a double-edged sword [77].
7.3. Carotenoids
7.4. Bioconverted Metabolites as Postbiotics
The nature of postbiotics may vary depending on the content of the medium in which the producing bacteria grow. Various bioactive metabolites can be produced by the bioconversion capabilities of probiotics. Lee et al. [81] published an important review on the use of bioactive metabolites obtained by probiotics-mediated bioconversion as therapeutic agents in periodontitis. Based on the concept of bioconversion mentioned in this study, it is possible to include all metabolites that probiotics can produce in the category of postbiotics, depending on the diversity of culture media in which they are cultivated. Thanks to their rich enzymatic arsenal, probiotic bacteria are capable of biotransforming many sources added to the culture media. For example, probiotics are able to produce fatty acids with different structures from fatty acids added to the culture media, bioactive peptides from milk and soy milk, and various flavonoid and phenolic compounds [82,83,84,85]. For example, L. rhamnosus GG and L. plantarum 299 v strains can ferment phenolic compounds to ferulic acid and caffeic acid [86]. Ferulic acid can induce heme oxygenase-1 via activation of ERK and Nrf2 in lymphocytes [87]. Cell proliferation and healing in oral tissues can be promoted by ferulic acid [88]. Caffeic acid appears to be an effective chemoprotective agent against oxidative damage, as it has been shown to stimulate the expression of detoxification enzymes via the ERK/Nrf2 pathway [89]. The increase in total antioxidant status (TAS) and decrease in total oxidative status (TOS) levels in inflamed gingival tissue suggest that caffeic acid has antioxidant therapeutic potential in periodontitis [90]. A natural compound known as urolithin A (UA) is derived from gut bacteria by the process of digested ellagitannins (ETs) and ellagic acid (EA), polyphenols abundant in fruits, nuts, and pomegranates [91]. Urolithin can enhance barrier functions through activation of aryl hydrocarbon receptor (AhR)-Nrf2-dependent pathways. By reversing barrier dysfunction, it reduces colitis in preclinical models, in addition to its anti-inflammatory and antioxidant properties [92]. Catechin is another metabolite that can be produced by probiotics through the fermentation of polyphenols [93]. Catechin can protect the gastric mucosa from oxidative damage by activating Nrf2 [94]. Casein-derived antioxidant peptides produced by microbial proteases are also gaining popularity for their potential Nrf2 activation. Casein-derived peptides obtained by microbial proteases can act as Keap1-Nrf2 signaling activators [95].
It should be noted that the above postbiotics have not yet been supported by direct in vitro or preclinical studies to regulate the Nrf2 response in periodontitis. However, postbiotics that have been associated with the Nrf2 response in other studies are likely to be candidates for activating periodontitis-related antioxidant response mechanisms in the future.
8. Postbiotics as Nrf2 Activators in Periodontitis: Scientific Evidence
Currently, there is limited evidence that postbiotics can be used in the oral cavity as modulators of host response, inthe form of in vitro studies. Some Lactobacillus strains are able to metabolize linoleic acid into various isomeric intermediates, i.e., fatty acids. Among these fatty acids, 10-oxo-trans-11-octadecenoic acid (KetoC) mediates the antioxidant response via the Nfr2-ARE pathway. KetoC induces the expression of antioxidant-related genes in gingival epithelial cells and administration of KetoC reduces ROS levels by promoting phosphorylation of signal-regulated kinase (ERK), leading to nuclear translocation of Nrf2 [96]. Nrf2 regulates the expression of heme oxygenase-1 (HO-1), which has been extensively studied for its immunomodulatory and cryoprotective roles [97]. Vo et al. [98] showed that a Bacillus subtilis-derived surfactin has the potential to induce Nrf2 activation and HO-1 expression. Surfactin inhibited particulate-matter-induced VCAM-1 expression by increasing Nrf2-dependent HO-1 activation in human gingival fibroblasts.
9. Postbiotics as Nrf2 Activators and Their Potential Use as Antioxidants in Periodontitis—Future Perspective
Anti-infective periodontal treatment (nonsurgical periodontal treatment/initial treatment/cause-related therapy) conventionally aims to control inflammation by reducing the microbial burden and prevent re-colonization by removing deposits on hard surfaces and by instructing proper oral hygiene measures (Figure 4). Following anti-infective treatment, periodontal surgery can be implemented to regenerate lost tissues and/or to obtain physiological contours. Various current nonsurgical and surgical approaches in periodontal treatment yield similar outcomes, but aimed clinical endpoints of therapy are not fully achieved in a significant number of sites or individuals [99,100]. While mild–moderate periodontitis cases can be effectively treated with nonsurgical treatment, more advanced cases require the inclusion of periodontal surgical treatments into the treatment plan. During the last two decades, the benefit of adjunctive applications, such as the use of laser or photodynamic therapy or host-modifying agents (anti-inflammatories, matrix metalloproteinase inhibitors, bisphosphonates, anti-oxidants etc.), as part of periodontal therapy, has been discussed very widely [101,102]. In fact, host modulation has been widely studied, since our understanding of periodontal treatment is shifting towards controlling the infection by reducing inflammation [101]. Indeed, animal studies have demonstrated that oxidative stress can be decreased with certain agents, such as etanercept, Stemodia maritima L. extract, gliclazide, Calendula officinalis, and atorvastatin [103,104,105,106,107]. On the other hand, scientific evidence is yet scarce to consider many host modulators for daily clinical use [108].
According to the European Federation of Periodontology treatment guideline, the scientific evidence for the complementary use of lasers, photodynamic therapy, probiotics, doxycycline in subantimicrobial doses, local or systemic bisphosphonates or nonsteroidal anti-inflammatory drugs, and local administration of statin or metformin gels is currently insufficient [109]. Therefore, exploration of different complementary treatment models, such as systemic or local administration of postbiotics, might be beneficial. Hypothetically, postbiotics could be used as an adjunct to periodontal treatment, especially in patients who do not respond to treatment, to reduce the inflammatory burden and, indirectly, infection by reducing oxidative stress. In addition, they could modulate the host response and prevent disease progression or recurrence in susceptible individuals.
10. Conclusions
Postbiotics can be considered as candidate host-modifying agents, since they are thought to be more reliable than probiotics regarding their clinical safety. Various postbiotics have been proven to have the potential to activate the Nrf2 pathway, which, in return, can reduce oxidative damage and inflammatory burden in periodontitis. The adjunctive use of postbiotics as Nrf2 activators can be particularly beneficial in non-responsive or high-risk periodontitis patients.
Author Contributions
Conceptualization, review and editing, U.K.G. Writing—original draft preparation, B.K. and M.Y. All authors have read and agreed to the published version of the manuscript.
Funding
The authors Basar Karaca and Mustafa Yilmaz are currently being supported by The Scientific and Technical Research Council of Turkey (TÜBİTAK) with the project numbers 1059B192001325 and 1059B192000842, respectively.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The study did not report any data.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study or in the writing of the manuscript.
References
- Chapple, I.L.C.; Mealey, B.L.; Van Dyke, T.E.; Bartold, P.M.; Dommisch, H.; Eickholz, P.; Geisinger, M.L.; Genco, R.J.; Glogauer, M.; Goldstein, M.; et al. Periodontal Health and Gingival Diseases and Conditions on an Intact and a Reduced Periodontium: Consensus Report of Workgroup 1 of the 2017 World Workshop on the Classification of Periodontal and Peri-Implant Diseases and Conditions. J. Periodontol. 2018, 89, S74–S84. [Google Scholar] [CrossRef] [PubMed]
- Silva, N.; Abusleme, L.; Bravo, D.; Dutzan, N.; Garcia-Sesnich, J.; Vernal, R.; Hernández, M.; Gamonal, J. Host Response Mechanisms in Periodontal Diseases. J. Appl. Oral Sci. 2015, 23, 329–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dyke, T.E.; Sheilesh, D. Risk Factors for Periodontitis. J. Int. Acad. Periodontol. 2005, 7, 3–7. [Google Scholar] [PubMed]
- Cekici, A.; Kantarci, A.; Hasturk, H.; Van Dyke, T.E. Inflammatory and Immune Pathways in the Pathogenesis of Periodontal Disease. Periodontol. 2000 2014, 64, 57–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-García, M.; Hernández-Lemus, E. Periodontal Inflammation and Systemic Diseases: An Overview. Front. Physiol. 2021, 12, 709438. [Google Scholar] [CrossRef]
- Landzberg, M.; Doering, H.; Aboodi, G.M.; Tenenbaum, H.C.; Glogauer, M. Quantifying Oral Inflammatory Load: Oral Neutrophil Counts in Periodontal Health and Disease. J. Periodontal Res. 2015, 50, 330–336. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Chavakis, T.; Hajishengallis, E.; Lambris, J.D. Neutrophil Homeostasis and Inflammation: Novel Paradigms from Studying Periodontitis. J. Leukoc. Biol. 2015, 98, 539–548. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Korostoff, J.M. Revisiting the Page & Schroeder Model: The Good, the Bad and the Unknowns in the Periodontal Host Response 40 Years Later. Periodontol. 2000 2017, 75, 116–151. [Google Scholar] [CrossRef]
- Hajishengallis, G. Immunomicrobial Pathogenesis of Periodontitis: Keystones, Pathobionts, and Host Response. Trends Immunol. 2014, 35, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Bagan, J.; Saez, G.; Tormos, C.; Gavalda, C.; Sanchis, J.M.; Bagan, L.; Scully, C. Oxidative stress and recurrent aphthous stomatitis. Clin. Oral Investig. 2014, 18, 1919–1923. [Google Scholar] [CrossRef]
- Tvarijonaviciute, A.; Aznar-Cayuela, C.; Rubio, C.P.; Ceron, J.J.; López-Jornet, P. Evaluation of salivary oxidate stress biomarkers, nitric oxide and C-reactive protein in patients with oral lichen planus and burning mouth syndrome. J. Oral Pathol. Med. 2017, 46, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Sardaro, N.; Della Vella, F.; Incalza, M.A.; Di Stasio, D.; Lucchese, A.; Contaldo, M.; Laudadio, C.; Petruzzi, M. Oxidative stress and oral mucosal diseases: An overview. In Vivo 2019, 33, 289–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyay, R.B.; Carnelio, S.; Shenoy, R.P.; Gyawali, P.; Mukherjee, M. Oxidative stress and antioxidant defense in oral lichen planus and oral lichenoid reaction. Scand. J. Clin. Lab. 2010, 70, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Cai, W.; Zhao, S.; Shi, L.; Chen, Y.; Li, X.; Sun, X.; Mao, Y.; He, B.; Hou, Y.; et al. Oxidative Stress-Related Biomarkers in Saliva and Gingival Crevicular Fluid Associated with Chronic Periodontitis: A Systematic Review and Meta-Analysis. J. Clin. Periodontol. 2019, 46, 608–622. [Google Scholar] [CrossRef] [PubMed]
- Sczepanik, S.F.C.; Grossi, M.L.; Casati, M.; Goldberg, M.; Glogauer, M.; Fine, N.; Tenenbaum, H.C. Periodontitis Is an Inflammatory Disease of Oxidative Stress: We Should Treat It That Way. Periodontol. 2000 2020, 84, 45–68. [Google Scholar] [CrossRef]
- Dias, I.H.; Matthews, J.B.; Chapple, I.L.; Wright, H.J.; Dunston, C.R.; Griffiths, H.R. Activation of the neutrophil respiratory burst by plasma from periodontitis patients is mediated by pro-inflammatory cytokines. J. Clin. Periodontol. 2011, 38, 1–7. [Google Scholar] [CrossRef]
- Chapple, I.L.C.; Matthews, J.B. The Role of Reactive Oxygen and Antioxidant Species in Periodontal Tissue Destruction. Periodontol. 2000 2007, 43, 160–232. [Google Scholar] [CrossRef]
- Ling, M.R.; Chapple, I.L.C.; Matthews, J.B. Neutrophil superoxide release and plasma C-reactive protein levels pre- and post-periodontal therapy. J. Clin. Periodontol. 2016, 43, 652–658. [Google Scholar] [CrossRef]
- Katsuragi, H.; Ohtake, M.; Kurasawa, I.; Saito, K. Intracellular production and extracellular release of oxygen radicals by PMNs and oxidative stress on PMNs during phagocytosis of periodontopathic bacteria. Odontology 2003, 91, 13–18. [Google Scholar] [CrossRef]
- Chapple, I.L.C. Role of Free Radicals and Antioxidants in the Pathogenesis of the Inflammatory Periodontal Diseases. Clin. Mol. Pathol. 1996, 49, M247. [Google Scholar] [CrossRef] [Green Version]
- AItman, L.C.; Baker, C.; Fleckman, P.; Luchtel, D.; Oda, D. Neutrophil-Mediated Damage to Human Gingival Epithelial Cells. J. Periodontal Res. 1992, 27, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K.; Choi, Y.G.; Baik, J.Y.; Han, S.Y.; Jeong, D.W.; Bae, Y.S.; Kim, N.; Lee, S.Y. A Crucial Role for Reactive Oxygen Species in RANKL-Induced Osteoclast Differentiation. Blood 2005, 106, 852–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, I.H.K.; Chapple, I.L.C.; Milward, M.; Grant, M.M.; Hill, E.; Brown, J.; Griffiths, H.R. Sulforaphane Restores Cellular Glutathione Levels and Reduces Chronic Periodontitis Neutrophil Hyperactivity In Vitro. PLoS ONE 2013, 8, e66407. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, L.; Mesgarzadeh, J.; Xu, I.; Powers, E.T.; Wiseman, R.L.; Bollong, M.J. Defining the functional targets of Cap ‘n’collar transcription factors NRF1, NRF2, and NRF3. Antioxidants 2020, 9, 1025. [Google Scholar] [CrossRef]
- Chapple, I.L.C.; Brock, G.; Eftimiadi, C.; Matthews, J.B. Glutathione in Gingival Crevicular Fluid and Its Relation to Local Antioxidant Capacity in Periodontal Health and Disease. Mol. Pathol. 2002, 55, 367. [Google Scholar] [CrossRef] [Green Version]
- Brock, G.R.; Butterworth, C.J.; Matthews, J.B.; Chapple, I.L.C. Local and Systemic Total Antioxidant Capacity in Periodontitis and Health. J. Clin. Periodontol. 2004, 31, 515–521. [Google Scholar] [CrossRef]
- Niture, S.K.; Khatri, R.; Jaiswal, A.K. Regulation of Nrf2—An Update. Free Radic. Biol. Med. 2014, 66, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Sima, C.; Aboodi, G.M.; Lakschevitz, F.S.; Sun, C.; Goldberg, M.B.; Glogauer, M. Nuclear Factor Erythroid 2-Related Factor 2 Down-Regulation in Oral Neutrophils Is Associated with Periodontal Oxidative Damage and Severe Chronic Periodontitis. Am. J. Pathol. 2016, 186, 1417–1426. [Google Scholar] [CrossRef] [Green Version]
- Hyeon, S.; Lee, H.; Yang, Y.; Jeong, W. Nrf2 Deficiency Induces Oxidative Stress and Promotes RANKL-Induced Osteoclast Differentiation. Free Radic. Biol. Med. 2013, 65, 789–799. [Google Scholar] [CrossRef]
- Kataoka, K.; Ekuni, D.; Tomofuji, T.; Irie, K.; Kunitomo, M.; Uchida, Y.; Fukuhara, D.; Morita, M. Visualization of Oxidative Stress Induced by Experimental Periodontitis in Keap1-Dependent Oxidative Stress Detector-Luciferase Mice. Int. J. Mol. Sci. Artic. 2016, 17, 1907. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yang, H.; Wen, Y.; Li, B.; Zhao, Y.; Xing, J.; Zhang, M.; Chen, Y. Nrf2 Inhibits Periodontal Ligament Stem Cell Apoptosis under Excessive Oxidative Stress. Int. J. Mol. Sci. Artic. Int. J. Mol. Sci. 2017, 18, 1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasnak, G.; Könönen, E.; Syrjänen, S.; Gürsoy, M.; Zeidán-Chuliá, F.; Firatli, E.; Gürsoy, U.K. NFE2L2/NRF2, OGG1, and Cytokine Responses of Human Gingival Keratinocytes against Oxidative Insults of Various Origin. Mol. Cell. Biochem. 2019, 452, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Dong, G.; Guo, L.; Graves, D.T. The function of dendritic cells in modulating the host response. Mol. Oral Microbiol. 2018, 33, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Sztukowska, M.; Ojo, A.; Scott, D.A.; Wang, H.; Lamont, R.J. FOXO Responses to Porphyromonas gingivalis in Epithelial Cells. Cell. Microbiol. 2015, 17, 1605–1617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michelle, P.; Caribé, V.; Cunha Villar, C.; Romito, G.A.; Pacanaro, A.P.; Cassaro Strunz, C.M.; Yoshio Takada, J.; Luiz, A.; Machado, C.; De Padua Mansur, A. Influence of the Treatment of Periodontal Disease in Serum Concentration of Sirtuin 1 and Mannose-Binding Lectin. J. Periodontol. 2020, 91, 900–905. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, Y.; Gao, J.; Li, T.; Gan, X.; Yu, H.; Xueqi Gan, C.; Key, S. Sirtuin 3 Deficiency Exacerbates Age-Related Periodontal Disease. J. Periodont. Res. 2021, 56, 1163–1173. [Google Scholar] [CrossRef]
- Carcuac, O.; Berglundh, T. Composition of human peri-implantitis and periodontitis lesions. J. Dent. Res. 2014, 93, 1083–1088. [Google Scholar] [CrossRef] [Green Version]
- Liskmann, S.; Zilmer, M.; Vihalemm, T.; Salum, O.; Fischer, K. Correlation of Peri-Implant Health and Myeloperoxidase Levels: A Cross-Sectional Clinical Study. Clin. Oral Implants Res. 2004, 15, 546–552. [Google Scholar] [CrossRef]
- Tözüm, T.F.; Akman, A.C.; Yamalik, N.; Tulunoglu, I.; Turkyilmaz, I.; Karabulut, E.; Kilinc, K.; Cehreli, M.C. Analysis of the Inflammatory Process Around Endosseous Dental Implants and Natural Teeth: Myeloperoxidase Level and Nitric Oxide Metabolism. Int. J. Oral Maxillofac. Implants. 2007, 22, 969–979. [Google Scholar]
- Güncü, G.N.; Tözüm, T.F.; Güncü, B.M.; Yamalik, N.; Tümer, C.; Karabulut, E.; Kilinç, K. Myeloperoxidase as a Measure of Polymorphonuclear Leukocyte Response in Inflammatory Status Around Immediately and Delayed Loaded Dental Implants: A Randomized Controlled Clinical Trial. Clin. Implant Dent. Relat. Res. 2007, 10, 30–39. [Google Scholar] [CrossRef]
- Kasnak, G.; Firatli, E.; Könönen, E.; Olgac, V.; Zeidán-Chuliá, F.; Gursoy, U.K. Elevated Levels of 8-OHdG and PARK7/DJ-1 in Peri-Implantitis Mucosa. Clin. Implant Dent. Relat. Res. 2018, 20, 574–582. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar-Toalá, J.E.; Garcia-Varela, R.; Garcia, H.S.; Mata-Haro, V.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An Evolving Term within the Functional Foods Field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Cuevas-González, P.F.; Liceaga, A.M.; Aguilar-Toalá, J.E. Postbiotics and Paraprobiotics: From Concepts to Applications. Food Res. Int. 2020, 136, 109502. [Google Scholar] [CrossRef] [PubMed]
- Żółkiewicz, J.; Marzec, A.; Ruszczyński, M.; Feleszko, W. Postbiotics—A Step Beyond Pre- and Probiotics. Nutrients 2020, 12, 2189. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 416763. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, Y.; Wang, Y.; Xu, H.; Mei, X.; Yu, D.; Wang, Y.; Li, W. Antioxidant Properties of Probiotic Bacteria. Nutrients 2017, 9, 521. [Google Scholar] [CrossRef]
- Valdes-Lopez, O.; Ntatsi, G.; Fotopoulos, V.; Georgiadou, E.C.; Kowalska, E.; Patla, K.; Kulbat, K.; Smolí Nska, B.; Leszczy’nska, J.; Leszczy’nska, L. Influence of Heavy Metals (Ni, Cu, and Zn) on Nitro-Oxidative Stress Responses, Proteome Regulation and Allergen Production in Basil (Ocimum basilicum L.) Plants. Front. Plant Sci. 2018, 9, 862. [Google Scholar] [CrossRef] [Green Version]
- Kullisaar, T.; Zilmer, M.; Mikelsaar, M.; Vihalemm, T.; Annuk, H.; Kairane, C.; Kilk, A. Two Antioxidative Lactobacilli Strains as Promising Probiotics. Int. J. Food Microbiol. 2002, 72, 215–224. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; del Carmen, S.; Miyoshi, A.; Azevedo, V.; Sesma, F.; Langella, P.; Bermúdez-Humarán, L.G.; Watterlot, L.; Perdigon, G.; de Moreno de LeBlanc, A. Use of Superoxide Dismutase and Catalase Producing Lactic Acid Bacteria in TNBS Induced Crohn’s Disease in Mice. J. Biotechnol. 2011, 151, 287–293. [Google Scholar] [CrossRef]
- Amaretti, A.; di Nunzio, M.; Pompei, A.; Raimondi, S.; Rossi, M.; Bordoni, A. Antioxidant Properties of Potentially Probiotic Bacteria: In Vitro and in Vivo Activities. Appl. Microb. Cell Physiol. 2013, 97, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Ejtahed, H.S.; Mohtadi-Nia, J.; Homayouni-Rad, A.; Niafar, M.; Asghari-Jafarabadi, M.; Mofid, V. Probiotic Yogurt Improves Antioxidant Status in Type 2 Diabetic Patients. Nutrition 2012, 28, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Kodali, V.P.; Sen, R. Antioxidant and free radical scavenging activities of an exopolysaccharide from a probiotic bacterium. Biotech. J. 2008, 3, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Afify, A.E.M.R.; Romeilah, R.M.; Sultan, S.I.M.; Hussein, M.M.; Afify, A.E.M.R.; Romeilah, R.M.; Shaimaa, I.M.; Hussein, M.M. Antioxidant Activity And Biological Evaluations Of Probiotic Bacteria Strains. Int. J. Acad. Res. 2012, 4, 131–139. [Google Scholar] [CrossRef]
- Fakruddin, M.; Ahmed, M.M. Antimicrobial and Antioxidant Activities of Saccharomyces cerevisiae IFST062013, a Potential Probiotic. BMC Complement Altern. Med. 2017, 17, 64. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Kim, J.S.; Kim, Y.; Jeong, Y.; Kim, J.E.; Paek, N.S.; Kang, C.H. Antioxidant and Probiotic Properties of Lactobacilli and Bifidobacteria of Human Origins. Biotechnol. Bioprocess Eng. 2020, 25, 421–430. [Google Scholar] [CrossRef]
- Gomes, A.C.; de Sousa, R.G.M.; Botelho, P.B.; Gomes, T.L.N.; Prada, P.O.; Mota, J.F. The Additional Effects of a Probiotic Mix on Abdominal Adiposity and Antioxidant Status: A Double-Blind, Randomized Trial. Obesity 2017, 25, 30–38. [Google Scholar] [CrossRef]
- Yang, S.J.; Lee, J.E.; Lim, S.M.; Kim, Y.J.; Lee, N.K.; Paik, H.D. Antioxidant and Immune-Enhancing Effects of Probiotic Lactobacillus plantarum 200655 Isolated from Kimchi. Food Sci. Biotechnol. 2017, 25, 30–38. [Google Scholar] [CrossRef]
- Bonfili, L.; Cecarini, V.; Cuccioloni, M.; Angeletti, M.; Berardi, S.; Scarpona, S.; Rossi, G.; Eleuteri, A.M. SLAB51 Probiotic Formulation Activates SIRT1 Pathway Promoting Antioxidant and Neuroprotective Effects in an AD Mouse Model. Mol. Neurobiol. 2018, 55, 7987–8000. [Google Scholar] [CrossRef] [Green Version]
- Aboulgheit, A.; Karbasiafshar, C.; Zhang, Z.; Sabra, M.; Shi, G.; Tucker, A.; Sodha, N.; Abid, M.R.; Sellke, F.W. Lactobacillus plantarum Probiotic Induces Nrf2-Mediated Antioxidant Signaling and ENOS Expression Resulting in Improvement of Myocardial Diastolic Function. Am. J. Physiol. Heart Circ. Physiol. 2021, 321, H839–H849. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.; Yang, Z.; Yang, W.; Huang, L.; Xu, C.; Liu, M.; Ge, J.; Wang, Y.; Jiang, S. Illicium verum Extracts and Probiotics with Added Glucose Oxidase Promote Antioxidant Capacity through Upregulating Hepatic and Jejunal Nrf2/Keap1 of Weaned Piglets. J. Anim. Sci. 2020, 98, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Maherinia, H.; Peeri, M.; Azarbayjani, M.A.; Delfan, M. Aerobic Exercise Training Combined with Probiotic Supplement Improves Antioxidant Defence of Cardiomyocytes by Regulating Nrf2 and Caspase3 Gene Expression in Type 2 Diabetic Rats. Comp. Exerc. Physiol. 2022, 0, 1–10. [Google Scholar] [CrossRef]
- Xu, H.; Wang, J.; Cai, J.; Feng, W.; Wang, Y.; Liu, Q.; Cai, L. Protective Effect of Lactobacillus rhamnosus GG and Its Supernatant against Myocardial Dysfunction in Obese Mice Exposed to Intermittent Hypoxia Is Associated with the Activation of Nrf2 Pathway. Int. J. Biol. Sci. 2019, 15, 2471. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, B.J.; Liu, K.H.; Owens, J.A.; Hunter-Chang, S.; Camacho, M.C.; Eboka, R.U.; Chandrasekharan, B.; Baker, N.F.; Darby, T.M.; Robinson, B.S.; et al. Gut-Resident Lactobacilli Activate Hepatic Nrf2 and Protect Against Oxidative Liver Injury. Cell Metab. 2020, 31, 956–968. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Wang, C.; Zhang, L.; Zhao, Y.; Duan, C.; Zhang, X.; Gao, L.; Li, S. Lactobacillus plantarum NA136 Improves the Non-Alcoholic Fatty Liver Disease by Modulating the AMPK/Nrf2 Pathway. Appl. Microbiol. Biotechnol. 2019, 103, 5843–5850. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, B.J.; Robinson, B.S.; Owens, J.; Liu, K.; Eboka, R.; Darby, T.; Luo, L.; Jones, D.; Jones, R.; Neish, A. Regulation of the Hepatic Antioxidant Response by the Probiotic Lactobacillus rhamnosus GG. FASEB J. 2019, 33, 369.5. [Google Scholar] [CrossRef]
- Endo, H.; Niioka, M.; Kobayashi, N.; Tanaka, M.; Watanabe, T. Butyrate-Producing Probiotics Reduce Nonalcoholic Fatty Liver Disease Progression in Rats: New Insight into the Probiotics for the Gut-Liver Axis. PLoS ONE 2013, 8, e63388. [Google Scholar] [CrossRef] [Green Version]
- Feng, T.; Wang, J. Oxidative Stress Tolerance and Antioxidant Capacity of Lactic Acid Bacteria as Probiotic: A Systematic Review. Gut Microbes 2020, 12, 1801944. [Google Scholar] [CrossRef]
- Lebeer, S.; Claes, I.J.J.; Verhoeven, T.L.A.; Vanderleyden, J.; De Keersmaecker, S.C.J. Exopolysaccharides of Lactobacillus rhamnosus GG form a protective shield against innate immune factors in the intestine. Microb. Biotechnol. 2011, 4, 368–374. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Ji, J.; Chen, X.; Jiang, M.; Rui, X.; Dong, M. Structural Elucidation and Antioxidant Activities of Exopolysaccharides from Lactobacillus helveticus MB2-1. Carbohydr. Polym. 2014, 102, 351–359. [Google Scholar] [CrossRef]
- Rani, R.P.; Anandharaj, M.; David Ravindran, A. Characterization of a Novel Exopolysaccharide Produced by Lactobacillus gasseri FR4 and Demonstration of Its in Vitro Biological Properties. Int. J. Biol. Macromol. 2018, 109, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, Q.; Gao, N.; Wang, Z.; Li, F.; Li, J.; Shan, A. Exopolysaccharides Produced by Lactobacillus rhamnosus GG Alleviate Hydrogen Peroxide-Induced Intestinal Oxidative Damage and Apoptosis through the Keap1/Nrf2 and Bax/Bcl-2 Pathways in vitro. Food Func. 2021, 12, 9632–9641. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Duncan, S.H.; Scott, K.P.; Louis, P. Links between diet, gut microbiota composition and gut metabolism. Proc. Nutr. Soc. 2015, 74, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef] [PubMed]
- Hamer, H.M.; Jonkers, D.M.A.E.; Bast, A.; Vanhoutvin, S.A.L.W.; Fischer, M.A.J.G.; Kodde, A.; Troost, F.J.; Venema, K.; Brummer, R.J.M. Butyrate Modulates Oxidative Stress in the Colonic Mucosa of Healthy Humans. Clin. Nutr. 2009, 28, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Jia, Y.; Yang, S.; Zhao, N.; Hu, Y.; Hong, J.; Gao, S.; Zhao, R. Sodium Butyrate Protects against High-Fat Diet-Induced Oxidative Stress in Rat Liver by Promoting Expression of Nuclear Factor E2-Related Factor 2. Br. J. Nutr. 2019, 122, 400–410. [Google Scholar] [CrossRef]
- Guan, X.; Li, W.; Meng, H. A double-edged sword: Role of butyrate in the oral cavity and the gut. Mol. Oral Microbiol. 2021, 36, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Ben-Dor, A.; Steiner, M.; Gheber, L.; Danilenko, M.; Dubi, N.; Linnewiel, K.; Zick, A.; Sharoni, Y.; Levy, J. Carotenoids Activate the Antioxidant Response Element Transcription System. Mol. Cancer Ther. 2005, 4, 177–186. [Google Scholar] [CrossRef]
- Hagi, T.; Kobayashi, M.; Nomura, M. Aerobic Condition Increases Carotenoid Production Associated with Oxidative Stress Tolerance in Enterococcus gilvus. FEMS Microbiol. Lett. 2014, 350, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Seo, D.H.; Park, Y.S.; Cha, I.T.; Seo, M.J. Isolation of Lactobacillus plantarum subsp. plantarum Producing C30 Carotenoid 4,4′-Diaponeurosporene and the Assessment of Its Antioxidant Activity. J. Microbiol. Biotechnol. 2019, 29, 1925–1930. [Google Scholar] [CrossRef]
- Lee, Y.; Yoon, Y.; Choi, K.H. Probiotics-Mediated Bioconversion and Periodontitis. Food Sci. Anim. 2021, 41, 905–922. [Google Scholar] [CrossRef] [PubMed]
- Muro Urista, C.; Lvarez Fernández, R.A.; Riera Rodriguez, F.; Cuenca, A.A.; Jurado, A.T. Review: Production and Functionality of Active Peptides from Milk. Food Sci. Technol. Int. 2011, 17, 293–317. [Google Scholar] [CrossRef] [PubMed]
- Suthanthangjai, W.; Kilmartin, P.A.; Phillips, A.R.J.; Davies, K.; Ansell, J. Bioconversion of Pinot Noir Anthocyanins into Bioactive Phenolic Compounds by Lactic Acid Bacteria. Nutr. Aging. 2014, 2, 145–149. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Hong, G.E.; Zhang, H.; Yang, C.Y.; Han, K.H.; Mandal, K.; Lee, C.H. Production of the Isoflavone Aglycone and Antioxidant Activities in Black Soymilk Using Fermentation with Streptococcus thermophilus S10. Food Sci. Biotechnol. 2015, 24, 537–544. [Google Scholar] [CrossRef]
- Lee, J.; Kim, M.; Kim, D.; Yu, C.; Kim, B.S.; Lee, J.S.; Kang, S.S. Anti-Biofilm Effect of Bioconversion of Whey by Lactic Acid Bacteria against Foodborne Pathogenic Bacteria. Curr. Top. Lact. Acid Bact. Probiotics 2020, 6, 25–31. [Google Scholar] [CrossRef]
- Rogozinska, M.; Korsak, D.; Mroczek, J.; Biesaga, M. Catabolism of Hydroxycinnamic Acids in Contact with Probiotic Lactobacillus. J. Appl. Microbiol. 2021, 131, 1464–1473. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Hong, Q.; Wang, Y.; Liang, Q.; Tan, H.; Xiao, C.; Tang, X.; Shao, S.; Zhou, S.; Gao, Y. Ferulic Acid Induces Heme Oxygenase-1 via Activation of ERK and Nrf2. Drug Discov. Ther. 2011, 5, 299–305. [Google Scholar] [CrossRef] [Green Version]
- San Miguel, S.M.; Opperman, L.A.; Allen, E.P.; Zielinski, J.; Svoboda, K.K.H. Bioactive Antioxidant Mixtures Promote Proliferation and Migration on Human Oral Fibroblasts. Arch. Oral Biol. 2011, 56, 812–822. [Google Scholar] [CrossRef]
- Yang, S.Y.; Cheol Pyo, M.; Nam, M.H.; Lee, K.W. ERK/Nrf2 Pathway Activation by Caffeic Acid in HepG2 Cells Alleviates Its Hepatocellular Damage Caused by t-Butylhydroperoxide-Induced Oxidative Stress. BMC Complement. Altern. Med. 2019, 19, 139. [Google Scholar] [CrossRef] [Green Version]
- Özden, F.O.; Lütfioğlu, M.; Demir, E.; Bilgici, B. Antioxidant Effect of Caffeic Acid Phenethyl Ester in Experimentally Induced Periodontitis. Clin. Oral Investig. 2021, 25, 4959–4966. [Google Scholar] [CrossRef]
- D’Amico, D.; Andreux, P.A.; Valdés, P.; Singh, A.; Rinsch, C.; Auwerx, J. Impact of the Natural Compound Urolithin A on Health, Disease, and Aging. Trends Mol. Med. 2021, 27, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Chandrashekharappa, S.; Bodduluri, S.R.; Baby, B.V.; Hegde, B.; Kotla, N.G.; Hiwale, A.A.; Saiyed, T.; Patel, P.; Vijay-Kumar, M.; et al. Enhancement of the Gut Barrier Integrity by a Microbial Metabolite through the Nrf2 Pathway. Nat. Commun. 2019, 10, 89. [Google Scholar] [CrossRef] [Green Version]
- Tabasco, R.; Sánchez-Patán, F.; Monagas, M.; Bartolomé, B.; Victoria Moreno-Arribas, M.; Peláez, C.; Requena, T. Effect of Grape Polyphenols on Lactic Acid Bacteria and Bifidobacteria Growth: Resistance and Metabolism. Food Microbiol. 2011, 28, 1345–1352. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.T.; Wu, C.H.; Ho, C.Y.; Yen, G.C. Catechin Protects against Ketoprofen-Induced Oxidative Damage of the Gastric Mucosa by up-Regulating Nrf2 in vitro and in vivo. J. Nutr. Biochem. 2013, 24, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Cui, Y.J.; Bai, S.S.; Yang, Z.J.; Miao-Cai; Megrous, S.; Aziz, T.; Sarwar, A.; Li, D.; Yang, Z.N. Antioxidant Activity of Novel Casein-Derived Peptides with Microbial Proteases as Characterized via Keap1-Nrf2 Pathway in HepG2 Cells. J. Microbiol. Biotechnol. 2021, 31, 1163–1174. [Google Scholar] [CrossRef]
- Yokoji-Takeuchi, M.; Takahashi, N.; Yamada-Hara, M.; Sulijaya, B.; Tsuzuno, T.; Aoki-Nonaka, Y.; Tabeta, K.; Kishino, S.; Ogawa, J.; Yamazaki, K. A Bacterial Metabolite Induces Nrf2-Mediated Anti-Oxidative Responses in Gingival Epithelial Cells by Activating the MAPK Signaling Pathway. Arch. Oral Biol. 2020, 110, 104602. [Google Scholar] [CrossRef]
- Campbell, N.K.; Fitzgerald, H.K.; Dunne, A. Regulation of Inflammation by the Antioxidant Haem Oxygenase 1. Nat. Rev. Immunol. 2021, 21, 411–425. [Google Scholar] [CrossRef]
- Vo, T.T.T.; Huang, H.W.; Wee, Y.; Feng, S.W.; Cheng, H.C.; Tuan, V.P.; Lee, I.T. Surfactin reduces particulate matter–induced VCAM-1–dependent monocyte adhesion in human gingival fibroblasts by increasing Nrf2-dependent HO-1 expression. J. Periodont. Res. 2022, 57, 115–130. [Google Scholar] [CrossRef]
- Graziani, F.; Karapetsa, D.; Alonso, B.; Herrera, D. Nonsurgical and surgical treatment of periodontitis: How many options for one disease? Periodontol. 2000 2017, 75, 152–188. [Google Scholar] [CrossRef]
- Van der Weijden, G.A.; Dekkers, G.J.; Slot, D.E. Success of non-surgical periodontal therapy in adult periodontitis patients: A retrospective analysis. Int. J. Dent. Hyg. 2019, 17, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Bartold, P.M.; Van Dyke, T.E. Host modulation: Controlling the inflammation to control the infection. Periodontol. 2000 2017, 75, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Preshaw, P.M. Host modulation therapy with anti-inflammatory agents. Periodontol. 2000 2018, 76, 131–149. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, R.; Mazzon, E.; Muià, C.; Crisafulli, C.; Terrana, D.; Greco, S.; Britti, D.; Santori, D.; Oteri, G.; Cordasco, G.; et al. Effects of Etanercept, a Tumour Necrosis Factor-a Antagonist, in an Experimental Model of Periodontitis in Rats. Br. J. Pharmacol. 2007, 150, 286–297. [Google Scholar] [CrossRef] [Green Version]
- Araújo, R.F.D.; Souza, T.O.; Moura, L.M.D.; Torres, K.P.; Souza, L.B.D.; Alves, M.D.S.C.F.; Rocha, H.O.; de Araújo, A.A. Atorvastatin decreases bone loss, inflammation and oxidative stress in experimental periodontitis. PLoS ONE 2013, 8, e75322. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.D.R.; Lopes, A.P.; Martins, C.; Brito, G.A.; Carneiro, V.C.; Goes, P. The effect of Calendula offiinalis on oxidative stress and bone loss in experimental periodontitis. Front. Physiol. 2017, 1, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, A.H.; Freire, J.M.D.O.; de Sousa, L.H.T.; Parente, A.T.; de Sousa, N.A.; Arriaga, A.M.C.; da Silva, F.R.L.; Melo, I.M.; da Silva, I.I.C.; Pereira, K.M.A.; et al. Stemodia maritima L. extract decreases inflammation, oxidative stress, and alveolar bone loss in an experimental periodontitis rat model. Front. Physiol. 2017, 8, 988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Araújo, A.A.; De Morais, H.B.; De Medeiros, C.A.C.X.; De Castro Brito, G.A.; Guedes, P.M.M.; Hiyari, S.; Pirih, F.Q.; De Araújo Júnior, R.F. Gliclazide Reduced Oxidative Stress, Inflammation, and Bone Loss in an Experimental Periodontal Disease Model. J. Appl. Oral Sci. 2019, 27, 1–14. [Google Scholar] [CrossRef]
- Donos, N.; Calciolari, E.; Brusselaers, N.; Goldoni, M.; Bostanci, N.; Belibasakis, G.N. The Adjunctive Use of Host Modulators in Non-Surgical Periodontal Therapy. A Systematic Review of Randomized, Placebo-Controlled Clinical Studies. J. Clin. Periodontol. 2020, 47, 199–238. [Google Scholar] [CrossRef] [Green Version]
- Sanz, M.; Herrera, D.; Kebschull, M.; Chapple, I.; Jepsen, S.; Berglundh, T.; Sculean, A.; Tonetti, M.S. Treatment of stage I–III periodontitis—The EFP S3 level clinical practice guideline. J. Clin. Periodontol. 2020, 47, 4–60. [Google Scholar] [CrossRef]
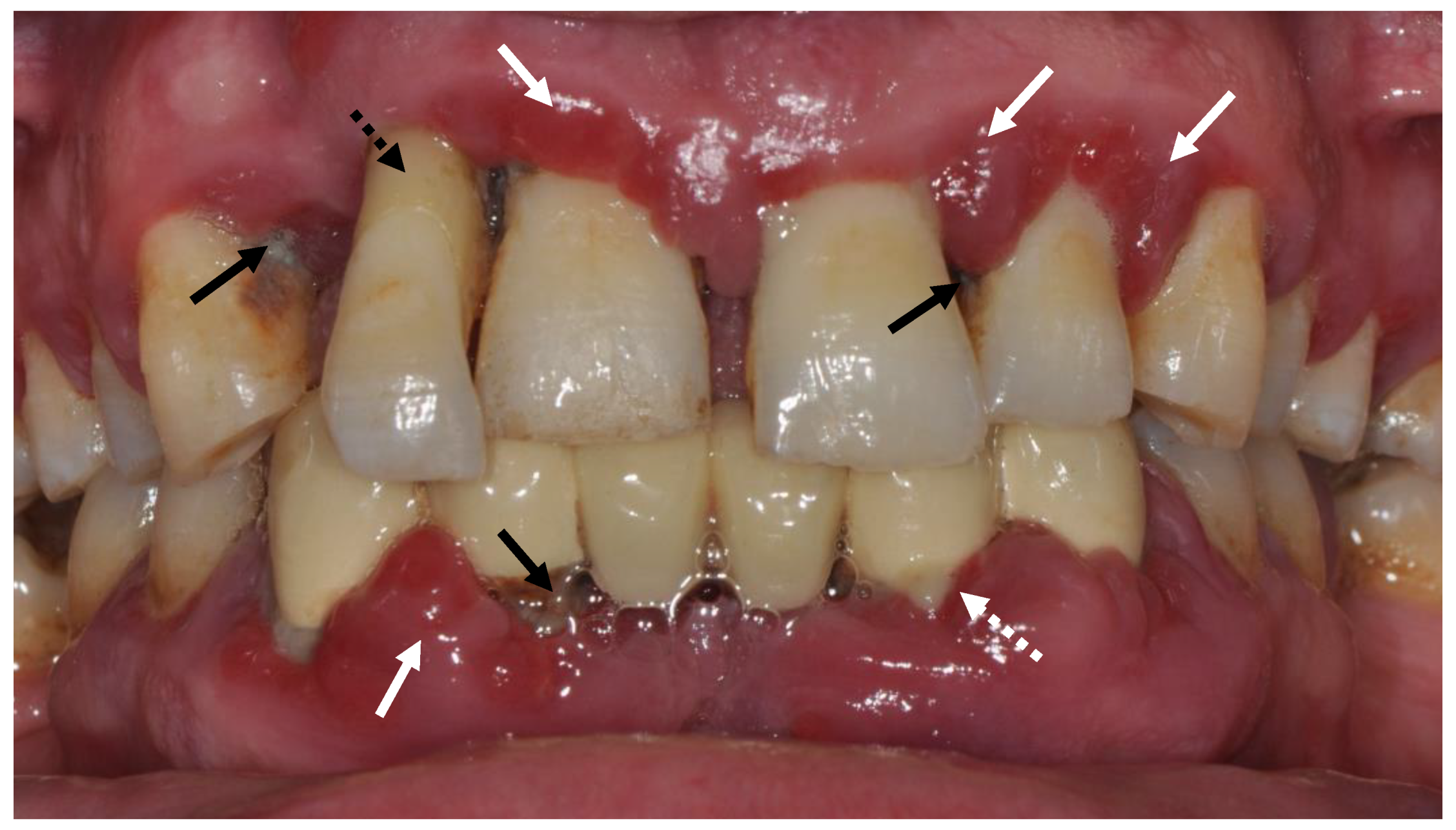
Figure 1.
Clinical signs of severe periodontitis. White arrows: Signs of gingival inflammation (erythema, swelling and texture change). White dashed arrow: Suppuration. Black arrows: Dental deposits. Dashed black arrow: Signs of Gingival recession and pathologic tooth migration.
Figure 1.
Clinical signs of severe periodontitis. White arrows: Signs of gingival inflammation (erythema, swelling and texture change). White dashed arrow: Suppuration. Black arrows: Dental deposits. Dashed black arrow: Signs of Gingival recession and pathologic tooth migration.

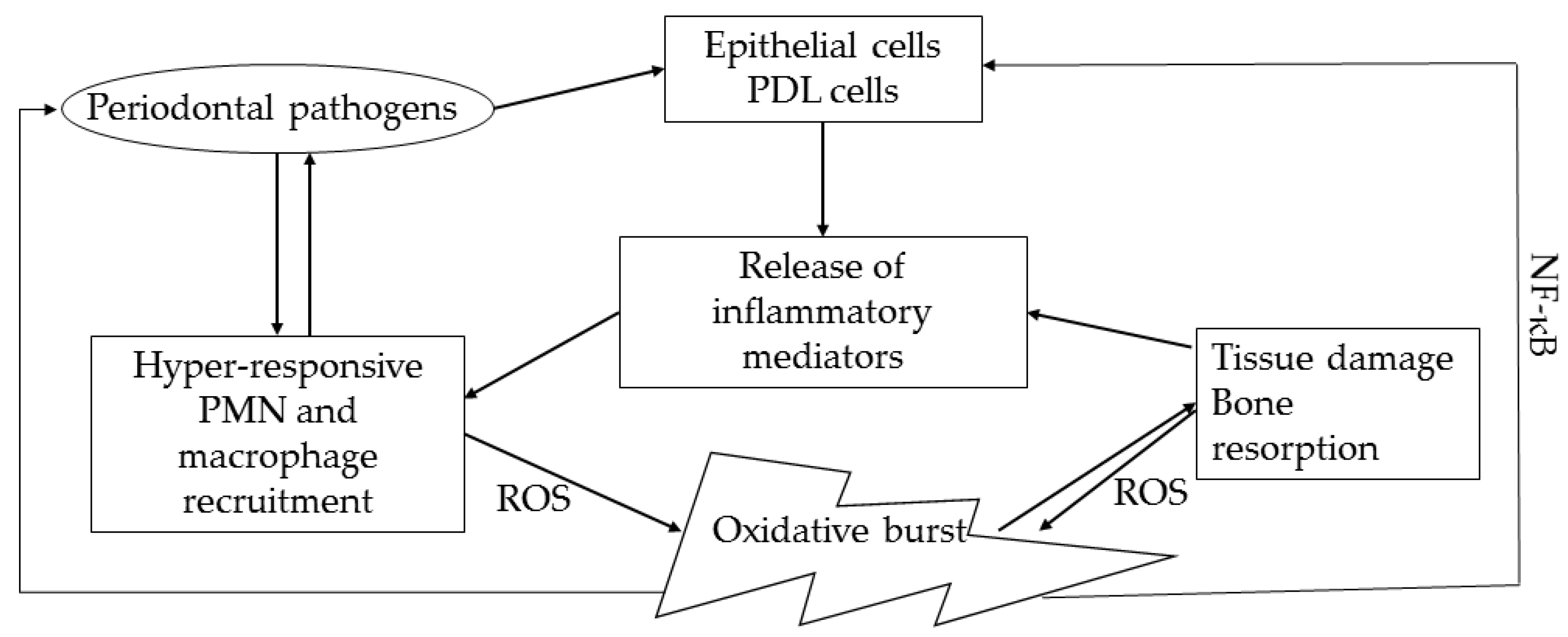
Figure 2.
Oxidative stress in periodontal disease pathogenesis. PDL: Periodontal Ligament; ROS: Reactive Oxygen Species; NF-κB: Nuclear Factor kappa B.
Figure 2.
Oxidative stress in periodontal disease pathogenesis. PDL: Periodontal Ligament; ROS: Reactive Oxygen Species; NF-κB: Nuclear Factor kappa B.

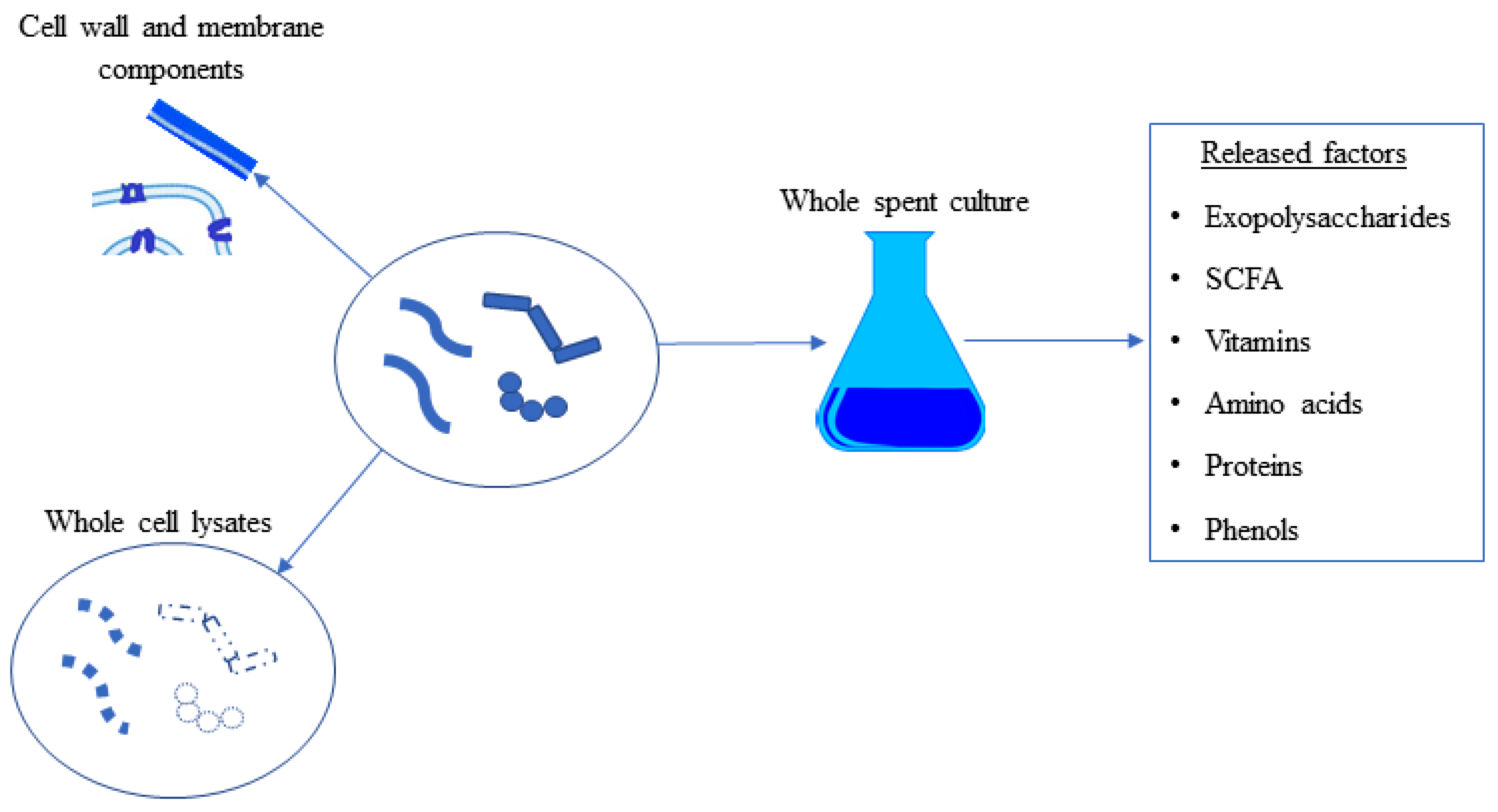
Figure 3.
The contents that can be classified as postbiotics. Whole cell lysates include all intracellular and cell membrane/cell wall components. Only cell wall and membrane components can be used as postbiotics. Factors secreted into the extracellular environment, fermentation broth, or spent culture media can be considered postbiotics. SCFA: Short-Chain Fatty Acids.
Figure 3.
The contents that can be classified as postbiotics. Whole cell lysates include all intracellular and cell membrane/cell wall components. Only cell wall and membrane components can be used as postbiotics. Factors secreted into the extracellular environment, fermentation broth, or spent culture media can be considered postbiotics. SCFA: Short-Chain Fatty Acids.

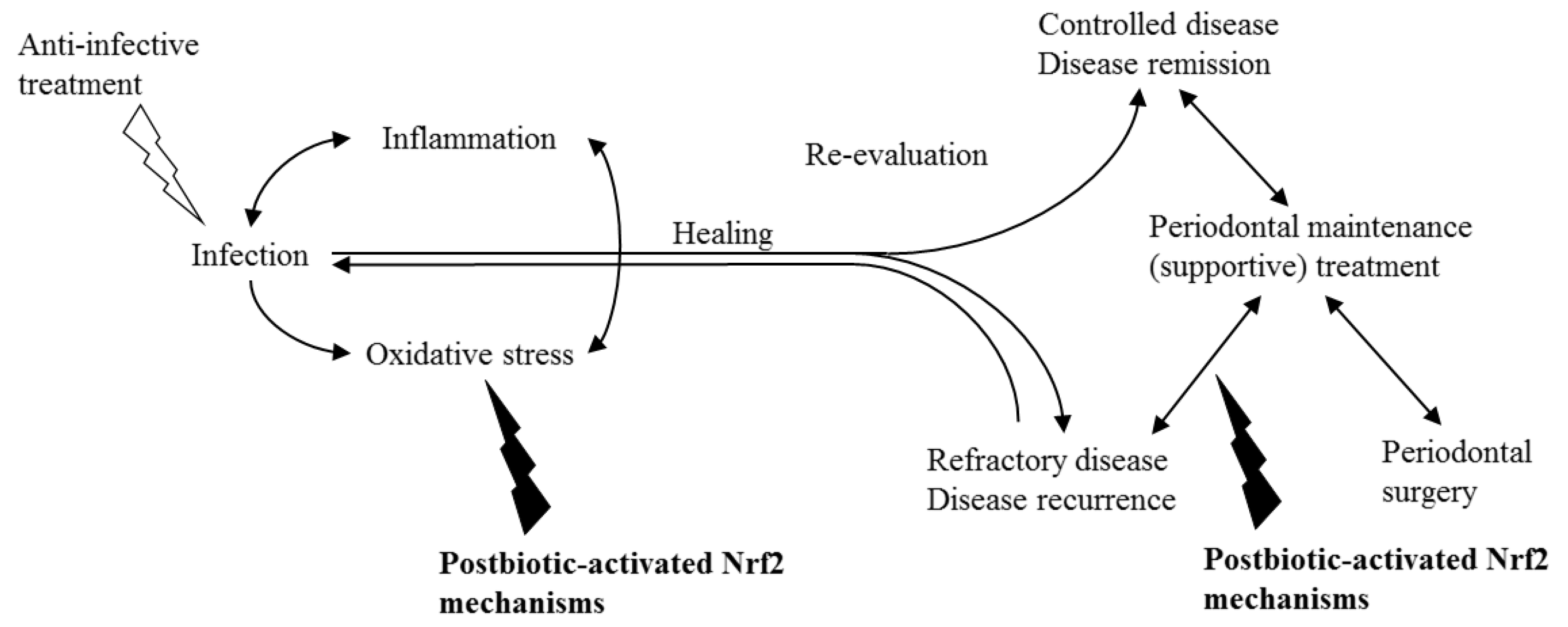
Figure 4.
The primary aim of conventional periodontal treatment is to control inflammation and oxidative stress by reducing the infection. Yet, host-modifying agents can be beneficial in unresponsive and/or susceptible individuals. Postbiotic-activated Nrf2 mechanisms can be considered as potential agents to control the inflammation by reducing oxidative stress as an adjunct to anti-infective periodontal treatment or during supportive therapy to prevent recurrence in high-risk individuals.
Figure 4.
The primary aim of conventional periodontal treatment is to control inflammation and oxidative stress by reducing the infection. Yet, host-modifying agents can be beneficial in unresponsive and/or susceptible individuals. Postbiotic-activated Nrf2 mechanisms can be considered as potential agents to control the inflammation by reducing oxidative stress as an adjunct to anti-infective periodontal treatment or during supportive therapy to prevent recurrence in high-risk individuals.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Probiotic bacteria and their postbiotic mediators as antioxidant agents.
| Probiotics | Antioxidant Source | Mode of Action | Reference |
|---|---|---|---|
| Bifidobacterium, Lactobacillus, Lactococcus, Streptococcus thermophilus | Intact cells and spent culture media * | Scavenging activity on radicals | [51] |
| Lactobacillus bulgaricus, Streptococcus thermophilus, Bifidobacterium lactis Bb12 and Lactobacillus acidophilus La5 strains | Yogurt including indicated probiotics | Increased erythrocyte superoxide dismutase and glutathione peroxidase activities and total antioxidant status in Type 2 diabetes | [52] |
| Bacillus coagulans RK-02 | B. coagulans derived exopolysaccharides * | Superoxide radical scavenging activity | [53] |
| Lactobacillus rhamnosus GG, Lactobacillus retueria (ATCC 20016), Bifidobacterium breve (ATCC 15700), Probionebacterium freudenreichii ssp. | Cell free culture extract * | DPPH scavenging activity | [54] |
| Saccharomyces cerevisiae IFST062013 | Cell extract and autolysate * | Hydroxyl radical and nitric oxide scavencing activity | [55] |
| Human originated Lactobacilli and Bifidobacteria strains | Living cells | DPPH and ABTS scavenging activity | [56] |
| Lactobacillus acidophilus LA-14, Lactobacillus casei LC-11, Lactococcus lactis LL-23, Bifidobacterium bifidum BB-06, and Bifidobacterium lactis BL-4 | Commercial probiotic Danisco® | Increased activity of glutathione peroxidase | [57] |
| Lactobacillus plantarum 200655 | Living cells isolated from kimchi | DPPH and ABTS scavenging activity | [58] |
| SLAB51 Probiotic Formulation | Living cells | Activates SIRT1 pathway promoting antioxidant effects | [59] |
* can be considered as postbiotics. ATCC: American Type Culture Collection. DPPH: 2,2-diphenyl-1-picrylhydrazyl. ABTS: 2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid). SIRT1: Sirtuin I.
Table 2.
Nrf2 activating abilities of probiotic bacteria.
| Source | Antioxidant Activity | Reference |
|---|---|---|
| Lactobacillus plantarum | Increased production of caffeic acid by L. plantarum in combination with apple juice promotes Nrf2 activation, resulting in improved diastolic function in chronic ischemic myocardium. | [60] |
| Commercial probiotic | Probiotics combined with Illicium verum extract and glucose oxidase enzyme upregulate hepatic and jejunal Nrf2/Keap1 pathway. | [61] |
| Commercial probiotic | Probiotic supplement improves antioxidant defence of cardiomyocytes by regulating Nrf2 and caspase3 gene expression in type 2 diabetic rats. | [62] |
| Lactobacillus rhamnosus GG and its spent culture supernatant | Improvement of myocardial dysfunction in obese mice exposed to intermittent hypoxia by activating Nrf2 Pathway | [63] |
| Lactobacillus rhamnosus GG | Significant induction of Nrf2 target transcripts in liver tissue via situmulation of xenobiotic | [64] |
| Lactobacillus plantarum NA136 | Increased the nuclear translocation of Nrf2 leading to improved antioxidant response in case of non-alcoholic fatty liver disease | [65] |
| Gut-resident Lactobacilli | Activation of Nrf2 response against oxidative liver injury through gut-liver axis | [66] |
| MIYAIRI 588–a butyrate-producing probiotic strain | Reduced nonalcoholic fatty liver disease progression via Nrf2 pathway | [67] |
Keap1: Kelch-like ECH-associated protein 1.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Karaca, B.; Yilmaz, M.; Gursoy, U.K. Targeting Nrf2 with Probiotics and Postbiotics in the Treatment of Periodontitis. Biomolecules 2022, 12, 729. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12050729
AMA Style
Karaca B, Yilmaz M, Gursoy UK. Targeting Nrf2 with Probiotics and Postbiotics in the Treatment of Periodontitis. Biomolecules. 2022; 12(5):729. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12050729
Chicago/Turabian StyleKaraca, Basar, Mustafa Yilmaz, and Ulvi Kahraman Gursoy. 2022. "Targeting Nrf2 with Probiotics and Postbiotics in the Treatment of Periodontitis" Biomolecules 12, no. 5: 729. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12050729
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.