
Hyperhomocysteinemia Increases Cortical Excitability and Aggravates Mechanical Hyperalgesia and Anxiety in a Nitroglycerine-Induced Migraine Model in Rats

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Model of Prenatal Hyperhomocysteinemia
2.2. Behavioral Assessment
2.2.1. Open Field Test
2.2.2. Von Frey Test
2.2.3. Light/Dark Transition Test
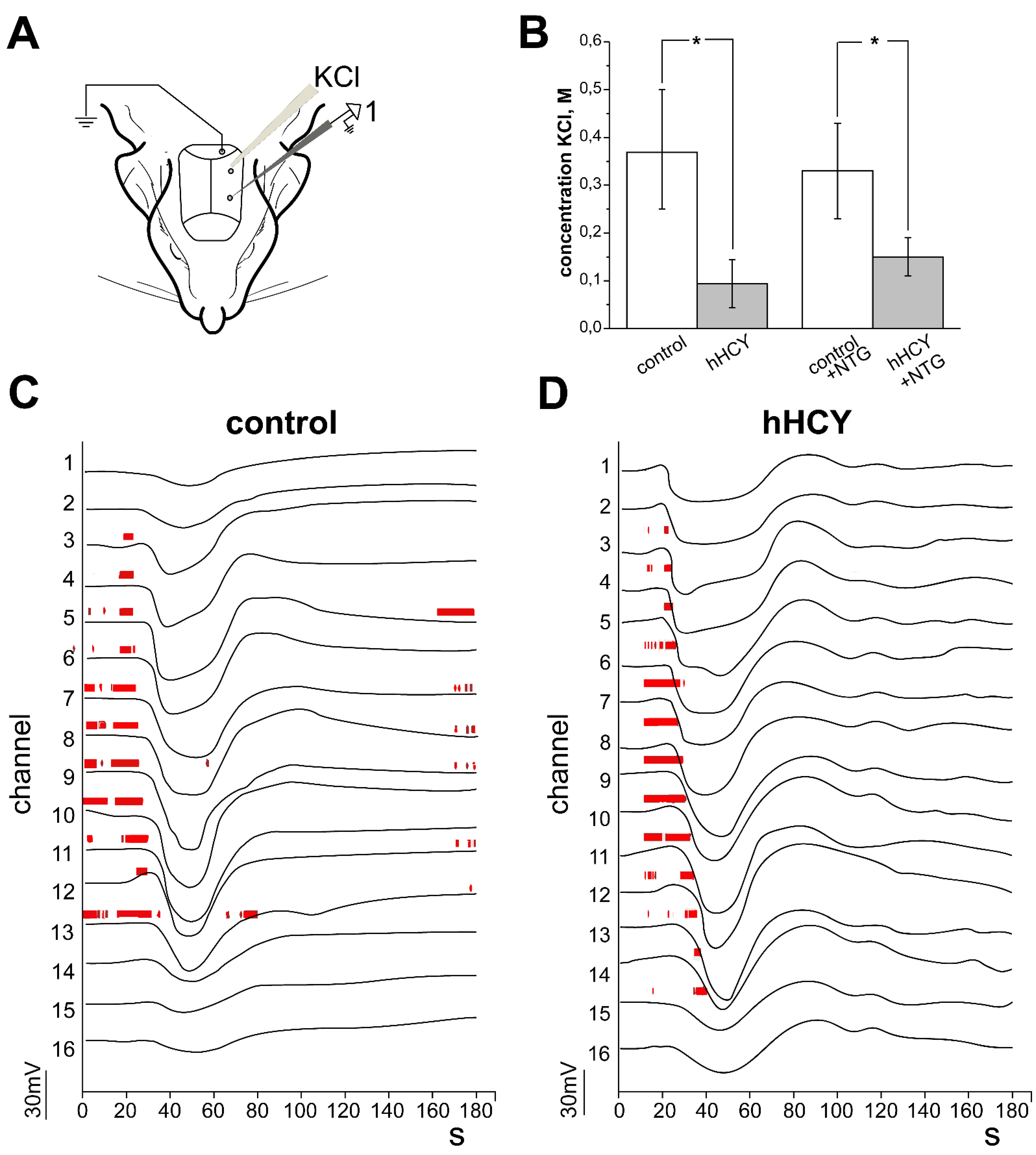
2.3. Electrophysiological Recordings of Cortical Spreading Depression
2.4. Statistical Analysis
3. Results
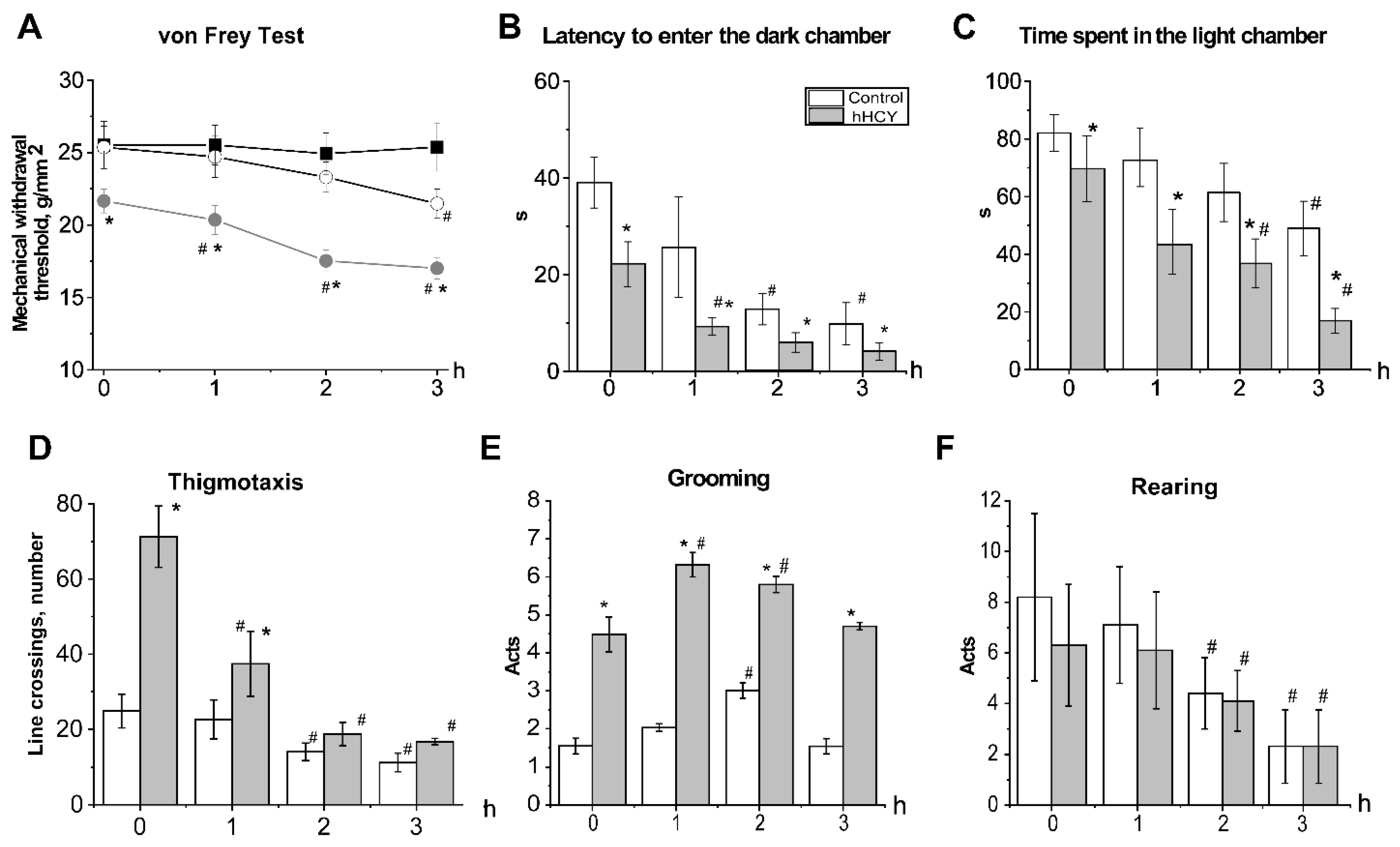
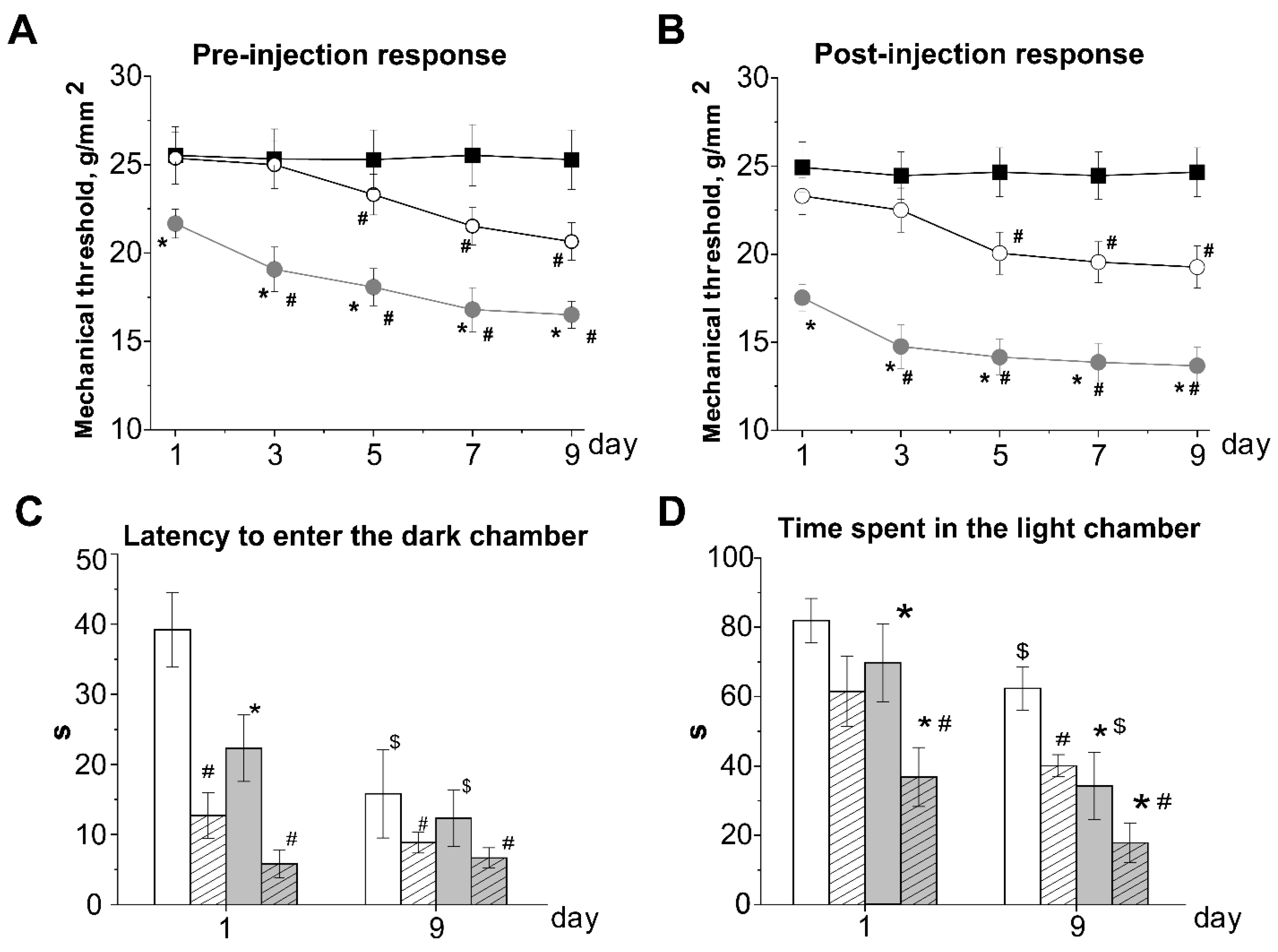
3.1. Effects of Acute and Chronic Nitroglycerine (NTG) Administration on Mechanical Sensitivity of Rats with hHCY
3.2. Anxiety and Photophobia in Rats with hHCY after Administration of Nitroglycerine in the Light-Dark Transition Test
3.3. Effects of Acute Nitroglycerine Administration on the Behavior of Rats with hHCY in the Open Field
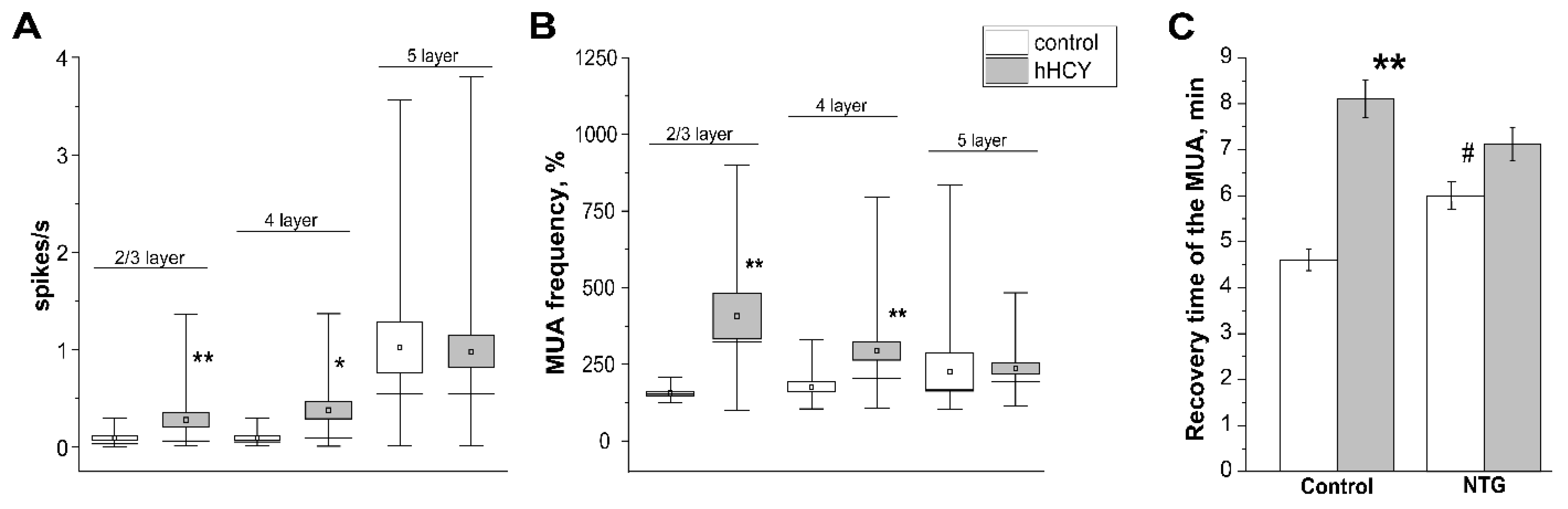
3.4. Effects of Nitroglycerine Administration on CSD and MUA in Rats with hHCY
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| hHCY | hyperhomocysteinemia |
| CSD | cortical spreading depression |
| MUA | multiple unit activity |
| MTHFR | methylenetetrahyrdofolate reductase |
| LFP | local field potential |
| NTG | nitroglycerine |
References
- Petras, M.; Tatarkova, Z.; Kovalska, M.; Mokra, D.; Dobrota, D.; Lehotsky, J.; Drgova, A. Hyperhomocysteinemia as a risk factor for the neuronal system disorders. J. Physiol. Pharmacol. 2014, 65, 15–23. [Google Scholar] [PubMed]
- Sitdikova, G.F.; Hermann, A.; Yakovlev, A.V. Neurotoxic and neuroprotective effects of homocysteine and hydrogen sulfide. Uchenye Zap. Kazan. Univ. Seriya Estestv. Nauk. 2018, 160, 686–704. [Google Scholar]
- Sharma, M.; Tiwari, M.; Tiwari, R.K. Hyperhomocysteinemia: Impact on neurodegenerative diseases. Basic Clin. Pharmacol. Toxicol. 2015, 117, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Vollset, S.E.; Refsum, H.; Irgens, L.M.; Emblem, B.M.; Tverdal, A.; Gjessing, H.K.; Monsen, A.L.; Ueland, P.M. Plasma total homocysteine, pregnancy complications, and adverse pregnancy outcomes: The Hordaland Homocysteine study. Am. J. Clin. Nutr. 2000, 71, 962–968. [Google Scholar] [CrossRef]
- Yakovleva, O.V.; Ziganshina, A.R.; Dmitrieva, S.A.; Arslanova, A.N.; Yakovlev, A.V.; Minibayeva, F.V.; Khaertdinov, N.N.; Ziyatdinova, G.K.; Giniatullin, R.A.; Sitdikova, G.F. Hydrogen sulfide ameliorates developmental impairments of rat offspring with prenatal hyperhomocysteinemia. Oxid. Med. Cell. Longev. 2018, 2018, 2746873. [Google Scholar] [CrossRef] [Green Version]
- Yakovleva, O.; Bogatova, K.; Mukhtarova, R.; Yakovlev, A.; Shakhmatova, V.; Gerasimova, E.; Ziyatdinova, G.; Hermann, A.; Sitdikova, G. Hydrogen sulfide alleviates anxiety, motor, and cognitive dysfunctions in rats with maternal hyperhomocysteinemia via mitigation of oxidative stress. Biomolecules 2020, 10, 995. [Google Scholar] [CrossRef]
- Arutjunyan, A.; Kozina, L.; Stvolinskiy, S.; Bulygina, Y.; Mashkina, A.; Khavinson, V. Pineal on? protects the rat offspring from prenatal hyperhomocysteinemia. Int. J. Clin. Exp. Med. 2012, 5, 179–185. [Google Scholar]
- Škovierová, H.; Vidomanová, E.; Mahmood, S.; Sopková, J.; Drgová, A.; Červeňová, T.; Erika, H.; Lehotský, J. The Molecular and Cellular Effect of Homocysteine Metabolism Imbalance on Human Health. Int. J. Mol. Sci. 2016, 17, 1733. [Google Scholar] [CrossRef]
- Lippi, G.; Mattiuzzi, C.; Meschi, T.; Cervellin, G.; Borghi, L. Homocysteine and migraine. A narrative review. Clin. Chim. Acta 2014, 433, 5–11. [Google Scholar] [CrossRef]
- Moschiano, F.; D’Amico, D.; Usai, S.; Grazzi, L.; Di Stefano, M.; Ciusani, E.; Erba, N.; Bussone, G. Homocysteine plasma levels in patients with migraine with aura. Neurol. Sci. 2008, 29, 173–175. [Google Scholar] [CrossRef]
- Liampas, I.; Siokas, V.; Mentis, A.A.; Aloizou, A.M.; Dastamani, M.; Tsouris, Z.; Aslanidou, P.; Brotis, A.; Dardiotis, E. Serum homocysteine, pyridoxine, folate, and vitamin B12 levels in migraine: Systematic review and meta-analysis. Headache 2020, 620, 1508–1534. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yu, Y.; He, J.; Guo, L.; Li, H.; Teng, J. Effects of MTHFR C677T and A1298C Polymorphisms on Migraine Susceptibility: A Meta-Analysis of 26 Studies. Headache 2019, 59, 891–905. [Google Scholar] [CrossRef]
- Tietjen, G.E.; Collins, S.A. Hypercoagulability and migraine. Headache 2018, 58, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Litvinov, R.I.; Peshkova, A.D.; Le Minh, G.; Khaertdinov, N.N.; Evtugina, N.G.; Sitdikova, G.F.; Weisel, J.W. Effects of Hyperhomocysteinemia on the Platelet-Driven Contraction of Blood Clots. Metabolites 2021, 11, 354. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Dong, Z.; Zhao, Y.; Sai, N.; Liu, H.; Huang, G.; Zhang, X. Homocysteine induces mitochondrial dysfunction involving the crosstalk between oxidative stress and mitochondrial pSTAT3 in rat ischemic brain. Sci. Rep. 2017, 7, 6932. [Google Scholar] [CrossRef]
- Olesen, I.; Tfelt-Hansen, P.; Olesen, J. Animal migraine models for drug development: Status and future perspectives. CNS Drugs 2013, 27, 1049–1068. [Google Scholar] [CrossRef] [PubMed]
- Ayata, C. Cortical spreading depression triggers migraine attack: Pro. Headache 2010, 50, 725–730. [Google Scholar] [CrossRef]
- Harriott, A.M.; Takizawa, T.; Chung, D.Y.; Chen, S.P. Spreading depression as a preclinical model of migraine. J. Headache Pain 2019, 20, 45. [Google Scholar] [CrossRef]
- Colonna, D.M.; Meng, W.; Deal, D.D.; Busija, D.W. Nitric oxide promotes arteriolar dilation during cortical spreading depression in rabbits. Stroke 1994, 25, 2463–2470. [Google Scholar] [CrossRef] [Green Version]
- Strecker, T.; Dux, M.; Messlinger, K. Nitric oxide releases calcitonin-gene-related peptide from rat dura mater encephali promoting increases in meningeal blood flow. J. Vasc. Res. 2002, 39, 489–496. [Google Scholar] [CrossRef]
- Christiansen, I.; Thomsen, L.L.; Daugaard, D.; Ulrich, V.; Olesen, J. Glyceryl trinitrate induces attacks of migraine without aura in sufferers of migraine with aura. Cephalalgia 1999, 19, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Afridi, S.K.; Matharu, M.S.; Lee, L.; Kaube, H.; Friston, K.J.; Frackowiak, R.S.; Goadsby, P.J. A PET study exploring the laterality of brainstem activation in migraine using glyceryl trinitrate. Brain 2005, 128 Pt 4, 932–939. [Google Scholar] [CrossRef]
- Olesen, J. The role of nitric oxide (NO) in migraine, tension-type headache and cluster headache. Pharmacol. Ther. 2008, 120, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Demartini, C.; Greco, R.; Zanaboni, A.M.; Sances, G.; De Icco, R.; Borsook, D.; Tassorelli, C. Nitroglycerin as a comparative experimental model of migraine pain: From animal to human and back. Prog. Neurobiol. 2019, 177, 15–32. [Google Scholar] [CrossRef] [PubMed]
- Greco, R.; Demartini, C.; Zanaboni, A.M.; Tassorelli, C. Chronic and intermittent administration of systemic nitroglycerin in the rat induces an increase in the gene expression of CGRP in central areas: Potential contribution to pain processing. J. Headache Pain 2018, 19, 51. [Google Scholar] [CrossRef] [Green Version]
- Greco, R.; Demartini, C.; Zanaboni, A.; Casini, I.; De Icco, R.; Reggiani, A.; Misto, A.; Piomelli, D.; Tassorelli, C. Characterization of the peripheral FAAH inhibitor, URB937, in animal models of acute and chronic migraine. Neurobiol. Dis. 2021, 147, 105157. [Google Scholar] [CrossRef]
- Bates, E.A.; Nikai, T.; Brennan, K.C.; Fu, Y.H.; Charles, A.C.; Basbaum, A.I.; Ptácek, L.J.; Ahn, A.H. Sumatriptan alleviates nitroglycerin-induced mechanical and thermal allodynia in mice. Cephalalgia 2010, 30, 170–178. [Google Scholar] [CrossRef] [Green Version]
- Markovics, A.; Kormos, V.; Gaszner, B.; Lashgarara, A.; Szoke, E.; Sandor, K.; Szabadfi, K.; Tuka, B.; Tajti, J.; Szolcsanyi, B.J.; et al. Pituitary adenylate cyclase-activating polypeptide plays a key role in nitroglycerol-induced trigeminovascular activation in mice. Neurobiol. Dis. 2012, 45, 633–644. [Google Scholar] [CrossRef]
- Gerasimova, E.; Burkhanova, G.; Chernova, K.; Zakharov, A.; Enikeev, D.; Khaertdinov, N.; Giniatullin, R.; Sitdikova, G. Hyperhomocysteinemia increases susceptibility to cortical spreading depression associated with photophobia, mechanical allodynia, and anxiety in rats. Behav. Brain Res. 2021, 409, 113324. [Google Scholar] [CrossRef]
- Bełtowski, J. Leptin and the regulation of endothelial function in physiological and pathological conditions. Clin. Exp. Pharmacol. Physiol. 2012, 39, 168–178. [Google Scholar] [CrossRef]
- Gerasimova, E.; Yakovleva, O.; Burkhanova, G.; Ziyatdinova, G.; Khaertdinov, N.; Sitdikova, G. Effects of maternal hyperhomocysteinemia, on the early physical development and neurobehavioral maturation of rat offspring. BioNanoScience 2017, 7, 155–158. [Google Scholar] [CrossRef]
- Kumar, S.; Ivanov, S.; Lagunin, A.; Goela, R.K. Attenuation of hyperhomocysteinemia induced vascular dementia by sodium orthovanadate perhaps via PTP1B: Pertinent downstream outcomes. Behav. Brain Res. 2019, 364, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Harrington, M.G.; Chekmenev, E.Y.; Schepkin, V.; Fonteh, A.N.; Arakaki, X. Sodium MRI in a rat migraine model and a NEURON simulation study support a role for sodium in migraine. Cephalalgia 2011, 31, 1254–1265. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Yeo, J.H.; Yoon, S.Y.; Kwon, S.G.; Lee, J.H.; Beitz, A.J.; Roh, D.H. Differential Development of Facial and Hind Paw Allodynia in a Nitroglycerin-Induced Mouse Model of Chronic Migraine: Role of Capsaicin Sensitive Primary Afferents. Biol. Pharm. Bull. 2018, 41, 172–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogdanov, V.B.; Bogdanova, O.V.; Koulchitskya, S.V.; Chauvela, M.V.; Makarchuk, M.Y.; Brennan, K.C.; Renshaw, P.F.; Schoenen, J. Behaviour in the open field predicts the number of KCl induced cortical spreading depressions in rats. Behav. Brain Res. 2013, 236, 90–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuralli, D.; Wattiez, A.; Andrew, F.R.; Bolay, H. Behavioral and cognitive animal models in headache research. J. Headache Pain 2019, 20, 11. [Google Scholar] [CrossRef]
- Crawley, J.N. Behavioral phenotyping of transgenic and knockout mice: Experimental design and evaluation of general health, sensory functions, motor abilities, and specific behavioral tests. Brain Res. 1999, 835, 18–26. [Google Scholar] [CrossRef]
- Moye, L.S.; Pradhan, A.A. Animal Model of Chronic Migraine-Associated Pain. Curr. Protoc. Neurosci. 2017, 80, 9–60. [Google Scholar] [CrossRef]
- Pradhan, A.A.; Smith, M.L.; McGuire, B.; Tarash, I.; Evans, C.J.; Charles, A. Characterization of a novel model of chronic migraine. Pain 2014, 155, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Bourin, M.; Hascoët, M. The mouse light/dark box test. Eur. J. Pharmacol. 2003, 463, 55–65. [Google Scholar] [CrossRef]
- Nasretdinov, A.; Lotfullina, N.; Vinokurova, D.; Lebedeva, J.; Burkhanova, G.; Chernova, K.; Zakharov, A.; Khazipov, R. Direct current coupled recordings of cortical spreading depression using silicone probes. Front. Cell. Neurosci. 2017, 11, 408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toriumi, H.; Shimizu, T.; Ebine, T.; Takizawa, T.; Kayama, Y.; Koh, A.; Shibata, M.; Suzuki, N. Repetitive trigeminal nociceptive stimulation in rats increases their susceptibility to cortical spreading depression. Neurosci. Res. 2016, 106, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Leão, A. Spreading depression of activity in the cerebral cortex. J. Neurophysiol. 1944, 7, 359–390. [Google Scholar] [CrossRef]
- Larrosa, B.; Pastor, J.; López-Aguado, L.; Herreras, O. A role for glutamate and glia in the fast network oscillations preceding spreading depression. Neuroscience 2006, 141, 1057–1068. [Google Scholar] [CrossRef] [PubMed]
- Burstein, R.; Yarnitsky, D.; Goor-Aryeh, I.; Ransil, B.J.; Bajwa, Z.H. An association between migraine and cutaneous allodynia. Ann. Neurol. 2000, 47, 614–662. [Google Scholar] [CrossRef]
- Baldacci, F.; Lucchesi, C.; Cafalli, M.; Poletti, M.; Ulivi, M.; Vedovello, M.; Giuntini, M.; Mazzucchi, S.; Prete, E.D.; Vergallo, A.; et al. Migraine features in migraineurs with and without anxiety–depression symptoms: A hospital-based study. Clin. Neurol. Neurosurg. 2015, 132, 74–78. [Google Scholar] [CrossRef]
- Peres, M.F.P.; Mercante, J.P.P.; Tobo, P.R.; Kamei, H.; Bigal, M.E. Anxiety and depression symptoms and migraine: A symptom-based approach research. J. Headache Pain 2017, 18, 37. [Google Scholar] [CrossRef] [Green Version]
- Goadsby, P.J.; Holland, P.R.; Martins-Oliveira, M.; Hoffmann, J.; Christoph, S.; Akerman, S. Pathophysiology of migraine: A disorder of sensory processing. Physiol. Rev. 2017, 97, 553–622. [Google Scholar] [CrossRef]
- Lea, R.A.; Ovcaric, M.; Sundholm, J.; MacMillan, J.; Griffiths, L.R. The methylenetetrahydrofolate reductase gene variant C677T influences susceptibility to migraine with aura. BMC Med. 2004, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Koutmos, M.; Datta, S.; Pattridge, K.A.; Smith, J.L.; Matthews, R.G. Insights into the reactivation of cobalamin-dependent methionine synthase. Proc. Natl. Acad. Sci. USA 2009, 106, 18527–18532. [Google Scholar] [CrossRef] [Green Version]
- Neeb, L.; Reuter, U. Nitric oxide in migraine. CNS Neurol. Disord. -Drug Targets 2007, 6, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Olesen, J. Nitric oxide-related drug targets in headache. Neurotherapeutics 2010, 7, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Pardutz, A.; Krizbai, I.; Multon, S.; Vecsei, L.; Schoenen, J. Systemic nitroglycerin increases nNOS levels in rat trigeminal nucleus caudalis. Neuroreport 2000, 11, 3071–3075. [Google Scholar] [CrossRef] [PubMed]
- Bellamy, J.; Bowen, E.J.; Russo, A.F.; Durham, P.L. Nitric oxide regulation of calcitonin gene-related peptide gene expression in rat trigeminal ganglia neurons. Eur. J. Neurosci. 2006, 23, 2057–2066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paolucci, M.; Altamura, C.; Vernieri, F. The Role of Endothelial Dysfunction in the Pathophysiology and Cerebrovascular Effects of Migraine: A Narrative Review. J. Clin. Neurol. 2021, 17, 164–175. [Google Scholar] [CrossRef]
- Lassen, L.H.; Ashina, M.; Christiansen, I.; Ulrich, V.; Olesen, J. Nitric oxide synthase inhibition in migraine. Lancet 1997, 8, 349, 401–402. [Google Scholar] [CrossRef]
- Gaifullina, A.S.; Lazniewska, J.; Gerasimova, E.V.; Burkhanova, G.F.; Rzhepetskyy, Y.; Tomin, A.; Rivas-Ramirez, P.; Huang, J.; Cmarko, L.; Zamponi, G.W.; et al. A potential role for T-type calcium channels in homocysteinemia-induced peripheral neuropathy. Pain 2019, 160, 2798–2810. [Google Scholar] [CrossRef]
- Harris, H.M.; Carpenter, J.M.; Black, J.R.; Smitherman, T.A.; Sufka, K.J. The effects of repeated nitroglycerin administrations in rats; modeling migraine-related endpoints and chronification. J. Neurosci. Methods. 2017, 284, 63–70. [Google Scholar] [CrossRef]
- Farajdokht, F.; Mohaddes, G.; Shanehbandi, D.; Karimi, P.; Babri, S. Ghrelin attenuated hyperalgesia induced by chronic nitroglycerin: CGRP and TRPV1 as targets for migraine management. Cephalalgia 2018, 38, 1716–1730. [Google Scholar] [CrossRef]
- Chen, H.; Tang, X.; Li, J.; Hu, B.; Yang, W.; Zhan, M.; Ma, T.; Xu, S. IL-17 crosses the blood-brain barrier to trigger neuroinflammation: A novel mechanism in nitroglycerin-induced chronic migraine. J. Headache Pain 2022, 23, 1. [Google Scholar] [CrossRef]
- Burstein, R.; Jakubowski, M. Managing migraine associated with sensitization. Handb. Clin. Neurol. 2010, 97, 207–215. [Google Scholar] [CrossRef]
- Tardiolo, G.; Bramanti, P.; Mazzon, E. Migraine: Experimental Models and Novel Therapeutic Approaches. Int. J. Mol. Sci. 2019, 20, 2932. [Google Scholar] [CrossRef] [Green Version]
- Sufka, K.J.; Staszko, S.M.; Johnson, A.P.; Davis, M.E.; Davis, R.E.; Smitherman, T.A. Clinically relevant behavioral endpoints in a recurrent nitroglycerin migraine model in rats. J. Headache Pain 2016, 17, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prut, L.; Belzung, C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: A review. Eur. J. Pharmacol. 2003, 463, 3–33. [Google Scholar] [CrossRef]
- Lalonde, R.; Barraud, H.; Ravey, J.; Guéant, J.L.; Bronowicki, J.P.; Strazielle, C. Effects of a B-vitamin-deficient diet on exploratory activity, motor coordination, and spatial learning in young adult Balb/c mice. Brain Res. 2008, 1188, 122–131. [Google Scholar] [CrossRef]
- Wang, L.; Alachkar, A.; Sanathara, N.; Belluzzi, J.D.; Wang, Z.; Civelli, O. A Methionine-Induced Animal Model of Schizophrenia: Face and Predictive Validity. Int. J. Neuropsychopharmacol. 2015, 18, pyv054. [Google Scholar] [CrossRef] [Green Version]
- Alachkar, A.; Wang, L.; Yoshimura, R.; Hamzeh, A.R.; Wang, Z.; Sanathara, N.; Lee, S.M.; Xu, X.; Abbott, G.W.; Civelli, O. Prenatal one-carbon metabolism dysregulation programs schizophrenia-like deficits. Mol. Psychiatry 2018, 23, 282–294. [Google Scholar] [CrossRef] [Green Version]
- Tucker, L.B.; McCabe, J.T. Measuring Anxiety-Like Behaviors in Rodent Models of Traumatic Brain Injury. Front. Behav. Neurosci. 2021, 15, 682935. [Google Scholar] [CrossRef]
- Ferguson, S.A.; Berry, K.J.; Hansen, D.K.; Wall, K.S.; White, G.; Antony, A.C. Behavioral Effects of Prenatal Folate Deficiency in Mice. Birth Defects Res. Part A Clin. Mol. Teratol. 2005, 73, 249–252. [Google Scholar] [CrossRef]
- Blaise, S.A.; Nédélec, E.; Schroeder, H.; Alberto, J.M.; Bossenmeyer-Pourié, C.; Guéant, J.L.; Daval, J.L. Gestational vitamin B deficiency lds to homocysteine-associated brain apoptosis and alters neurobehavioral development in rats. Am. J. Pathol. 2007, 170, 667–679. [Google Scholar] [CrossRef] [Green Version]
- Ishii, I.; Kamata, S.; Ito, S.; Shimonaga, A.; Koizumi, M.; Tsushima, M.; Miura, A.; Nagata, T.; Tosaka, Y.; Ohtani, H.; et al. A High-Methionine Diet for One-Week Induces a High Accumulation of Methionine in the Cerebrospinal Fluid and Confers Bipolar Disorder-like Behavior in Mice. Int. J. Mol. Sci. 2022, 23, 928. [Google Scholar] [CrossRef] [PubMed]
- Lauritzen, M. Pathophysiology of the migraine aura. The spreading depression theory. Brain 1994, 117, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Cacciapuoti, F. Migraine homocysteine-related: Old and new mechanisms. Neurol. Clin. Neurosci. 2017, 5, 137–140. [Google Scholar] [CrossRef]
- Knapp, L.; Szita, B.; Kocsis, K.; Vécsei, L.; Toldi, J. Nitroglycerin enhances the propagation of cortical spreading depression: Comparative studies with sumatriptan and novel kynurenic acid analogues. Drug Des Dev. Ther. 2016, 11, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petzold, G.C.; Haack, S.; von Bohlen Und Halbach, O.; Priller, J.; Lehmann, T.N.; Heinemann, U.; Dirnagl, U.; Dreier, J.P. Nitric oxide modulates spreading depolarization threshold in the human and rodent cortex. Stroke 2008, 39, 1292–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Chang, C.F.; Morales, M.; Chiang, Y.H.; Harvey, B.K.; Su, T.P.; Tsao, L.I.; Chen, S.; Thiemermann, C. Diadenosine tetraphosphate protects against injuries induced by ischemia and 6-hydroxydopamine in rat brain. J. Neurosci. 2003, 23, 7958–7965. [Google Scholar] [CrossRef] [Green Version]
- Džoljić, E.; Grbatinić, I.; Kostić, V. Why is nitric oxide important for our brain? Funct. Neurol. 2015, 30, 159–163. [Google Scholar] [CrossRef]
- Wang, H.G.; Lu, F.M.; Jin, I.; Udo, H.; Kandel, E.R.; de Vente, J.; Walter, U.; Lohmann, S.M.; Hawkins, R.D.; Antonova, I. Presynaptic and postsynaptic roles of NO, cGK, and RhoA in long-lasting potentiation and aggregation of synaptic proteins. Neuron 2005, 45, 389–403. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Takei, H.; Koyanagi, Y.; Koshikawa, N.; Kobayashi, M. Presynaptic cell type-dependent regulation of GABAergic synaptic transmission by nitric oxide in rat insular cortex. Neuroscience 2015, 284, 65–77. [Google Scholar] [CrossRef]
- Elsherbiny, N.M.; Sharma, I.; Kira, D.; Alhusban, S.; Samra, Y.A.; Jadeja, R.; Martin, P.; Al-Shabrawey, M.; Tawfik, A. Homocysteine Induces Inflammation in Retina and Brain. Biomolecules 2020, 10, 393. [Google Scholar] [CrossRef] [Green Version]
- Lai, W.K.; Kan, M.Y. Homocysteine-Induced Endothelial Dysfunction. Ann. Nutr. Metab. 2015, 67, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yakovlev, A.V.; Kurmashova, E.; Zakharov, A.; Sitdikova, G.F. Network-driven activity and neuronal excitability in hippocampus of neonatal rats with prenatal hyperhomocysteinemia. BioNanoScience 2018, 8, 304–309. [Google Scholar] [CrossRef]
- Lipton, S.A.; Kim, W.K.; Choi, Y.B.; Kumar, S.; d’Emilia, D.M.; Rayudu, P.V.; Arnelle, D.R.; Stamler, J.S. Neurotoxicity associated with dual actions of homocysteine at the N-methyl-D-aspartate receptor. Proc. Natl. Acad. Sci. USA 1997, 94, 5923–5928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lea, R.; Colson, N.; Quinlan, S.; McMillan, J.; Griffiths, L. The effects of vitamin supplementation and MTHFR (C677T) genotype on homocysteine-lowering and migraine disability. Pharmacogenet. Genom. 2009, 19, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Shaik, M.M.; Gan, S.H. Vitamin supplementation as possible prophylactic treatment against migraine with aura and menstrual migraine. Biomed Res. Int. 2015, 2015, 469529. [Google Scholar] [CrossRef] [Green Version]
- Liampas, I.N.; Siokas, V.; Aloizou, A.M.; Tsouris, Z.; Dastamani, M.; Aslanidou, P.; Brotis, A.; Dardiotis, E. Pyridoxine, folate and cobalamin for migraine: A systematic review. Acta Neurol. Scand. 2020, 142, 108–120. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (Mean ± SEM) | Hhcy (Mean ± SEM) | |
| Total number of line crossings | ||
| Before NTG | 33.7 ± 5.2 | 80.1 ± 6.8 * |
| +NTG 1h | 26.1 ± 5.6 | 40.7 ± 7.8 #* |
| +NTG 2h | 21.1 ± 4.9 # | 23.5 ± 2.6 # |
| +NTG 3h | 12.3 ± 2.3 # | 19.7 ± 0.4 # |
| Central zone activity | ||
| Before NTG | 6.5 ± 1.2 | 3.1 ± 0.7 * |
| +NTG 1h | 3.2 ± 0.6 # | 2.2 ± 0.6 #* |
| +NTG 2h | 1.4 ± 0.2 #* | 0.8 ± 0.3 # |
| +NTG 3h | 0.8 ± 0.2 # | 0.4 ± 0.2 # |
| Thigmotaxis | ||
| Before NTG | 24.8 ± 4.4 | 71.3 ± 8.1 * |
| +NTG 1h | 22.6 ± 5.2 | 37.4 ± 8.6 #* |
| +NTG 2h | 14.1 ± 2.3 # | 18.7 ± 3.1 # |
| +NTG 3h | 11.2 ± 2.4 # | 16.7 ± 0.8 # |
| KCl concentration | Number of Animals with CSD/all Animals | Number of CSD (Mean ± SEM) | Duration of CSD Occurrence (min) (Mean ± SEM) |
| 0.05 M | |||
| Control | 1/7 | 1 ± 0 | -- |
| hHCY | 3/6 | 1 ± 0 | -- |
| Control + NTG | 1/5 | 1 ± 0 | -- |
| hHCY + NTG | 2/7 | 1 ± 0 | -- |
| 0.1 M | |||
| Control | 3/7 | 2.1 ± 0.4 | 5.4 ± 1.9 |
| hHCY | 5/6 | 2.4 ± 0.3 | 10.4 ± 1.6 |
| Control + NTG | 2/5 | 2.0 ± 1.0 | 6.4 ± 3.1 |
| hHCY + NTG | 5/7 | 2.2 ± 0.7 | 9.1 ± 3.4 |
| 0.2 M | |||
| Control | 4/7 | 4.1 ± 0.6 | 13.3 ± 1.5 |
| hHCY | 6/6 | 4.7 ± 1.0 | 17.6 ± 3.0 |
| Control + NTG | 3/5 | 2.6 ± 0.8 | 7.4 ± 4.2 |
| hHCY + NTG | 5/7 | 3.4 ± 1.0 | 9.0 ± 3.4 |
| 0.3 M | |||
| Control | 4/7 | 3.9 ± 0.7 | 10.2 ± 3.8 |
| hHCY | 6/6 | 3.6 ± 0.7 | 12.3 ± 3.0 |
| Control + NTG | 4/5 | 2.7 ± 0.8 | 10.6 ± 4.8 |
| hHCY + NTG | 7/7 | 1.8 ± 0.4 | 6.4 ± 1.8 |
| 0.6 M | |||
| Control | 6/7 | 5.9 ± 1.2 | 19.3 ± 2.3 |
| hHCY | 6/6 | 6.4 ± 1.5 | 23.2 ± 8.2 |
| Control + NTG | 4/5 | 5.5 ± 1.5 | 20.4 ± 5.2 |
| hHCY + NTG | 7/7 | 3.5 ± 0.9 | 14.6 ± 2.6 |
| 1 M | |||
| Control | 7/7 | 7.2 ± 1.3 | 25.6 ± 4.5 |
| hHCY | 6/6 | 6.8 ± 1.7 | 20.3 ± 6.3 |
| Control + NTG | 5/5 | 5.4 ± 2.5 | 20.1 ± 6.0 |
| hHCY + NTG | 7/7 | 4.1 ± 0.8 | 14.2 ± 2.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gerasimova, E.; Yakovleva, O.; Enikeev, D.; Bogatova, K.; Hermann, A.; Giniatullin, R.; Sitdikova, G. Hyperhomocysteinemia Increases Cortical Excitability and Aggravates Mechanical Hyperalgesia and Anxiety in a Nitroglycerine-Induced Migraine Model in Rats. Biomolecules 2022, 12, 735. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12050735
Gerasimova E, Yakovleva O, Enikeev D, Bogatova K, Hermann A, Giniatullin R, Sitdikova G. Hyperhomocysteinemia Increases Cortical Excitability and Aggravates Mechanical Hyperalgesia and Anxiety in a Nitroglycerine-Induced Migraine Model in Rats. Biomolecules. 2022; 12(5):735. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12050735
Chicago/Turabian StyleGerasimova, Elena, Olga Yakovleva, Daniel Enikeev, Ksenia Bogatova, Anton Hermann, Rashid Giniatullin, and Guzel Sitdikova. 2022. "Hyperhomocysteinemia Increases Cortical Excitability and Aggravates Mechanical Hyperalgesia and Anxiety in a Nitroglycerine-Induced Migraine Model in Rats" Biomolecules 12, no. 5: 735. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12050735