
Myristic Acid Supplementation Aggravates High Fat Diet-Induced Adipose Inflammation and Systemic Insulin Resistance in Mice

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
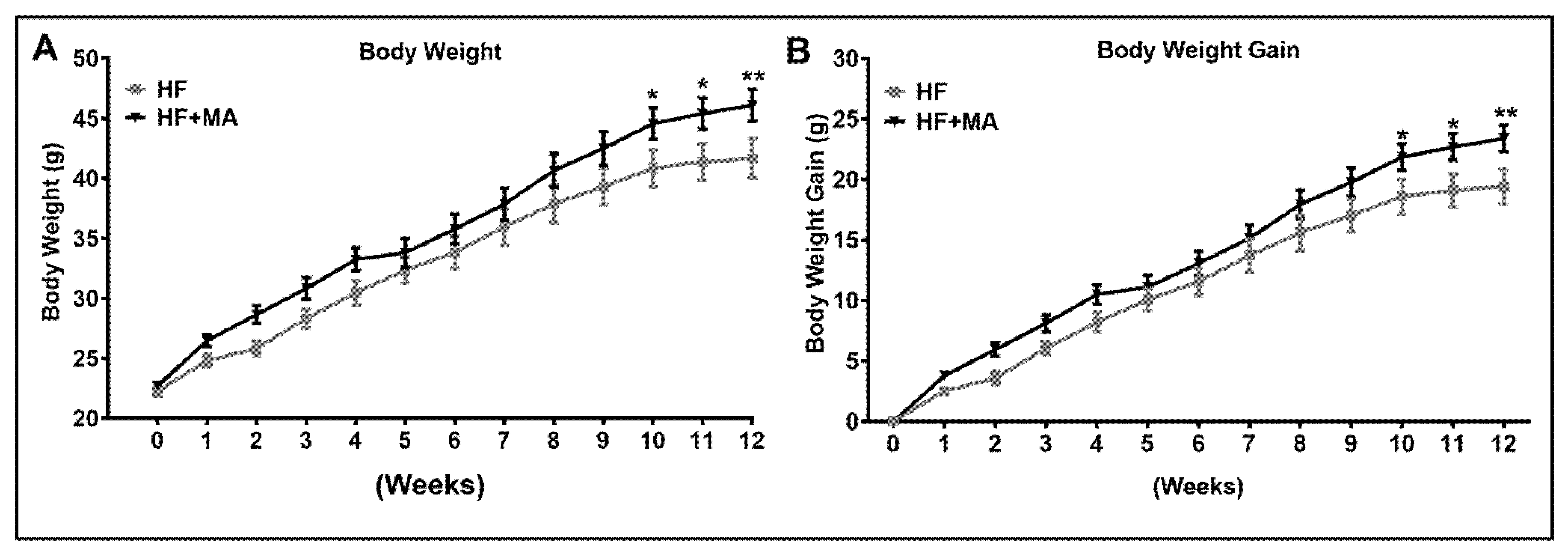
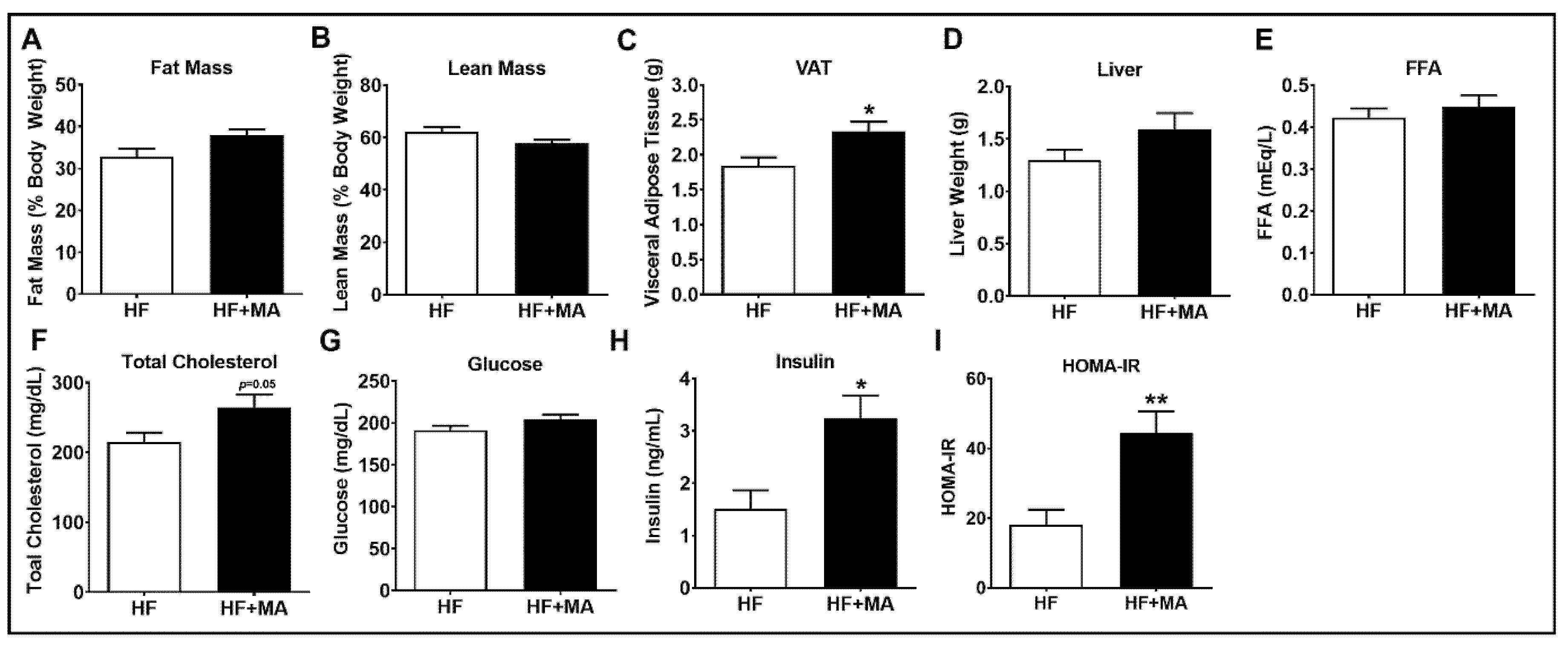
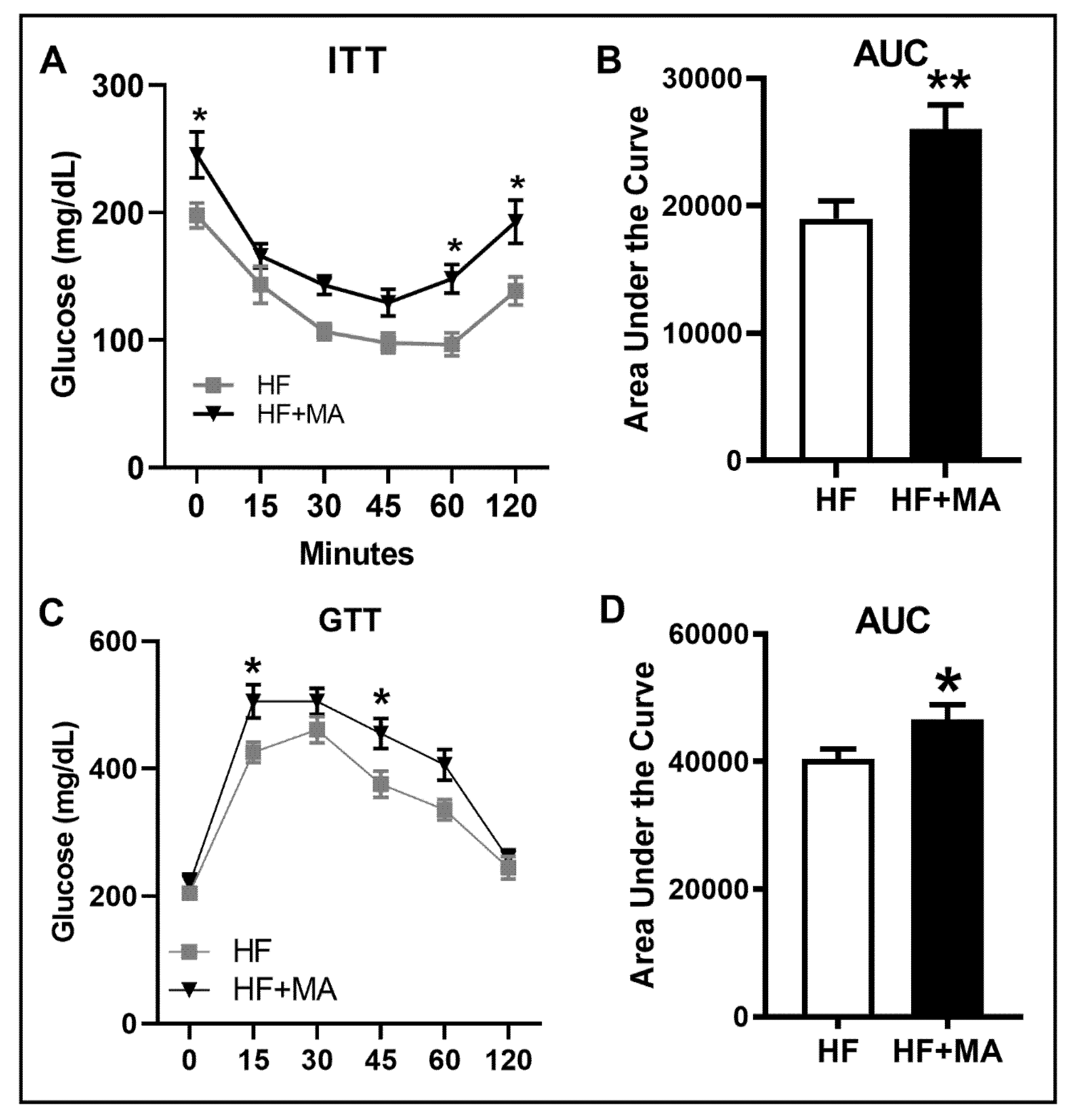
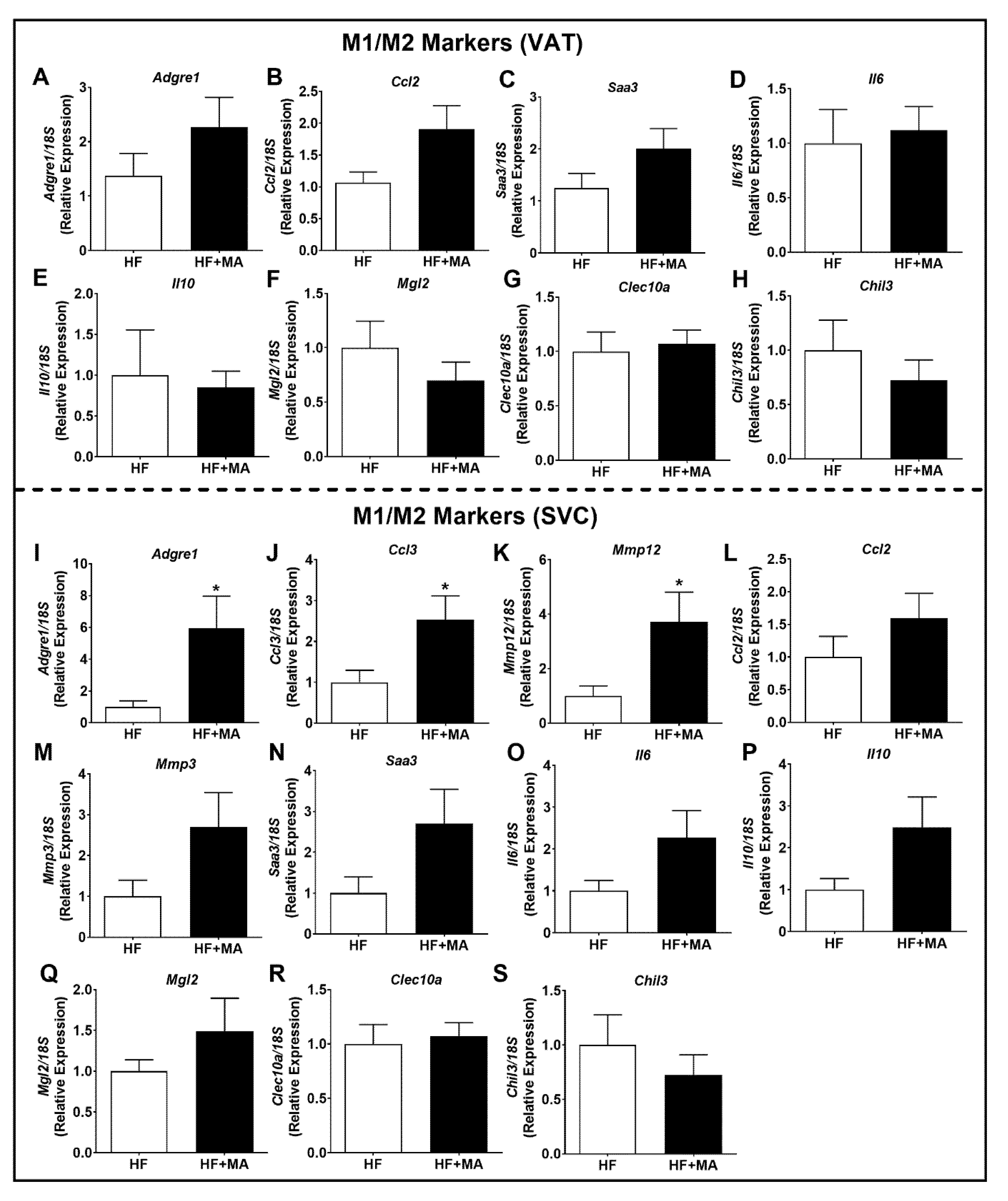
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fatima, S.; Hu, X.; Gong, R.H.; Huang, C.; Chen, M.; Wong, H.L.X.; Bian, Z.; Kwan, H.Y. Palmitic acid is an intracellular signaling molecule involved in disease development. Cell Mol. Life Sci. 2019, 76, 2547–2557. [Google Scholar] [CrossRef] [PubMed]
- Kien, C.L. Dietary interventions for metabolic syndrome: Role of modifying dietary fats. Curr. Diab. Rep. 2009, 9, 43–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houston, M. The relationship of saturated fats and coronary heart disease: Fa(c)t or fiction? A commentary. Ther. Adv. Cardiovasc. Dis. 2018, 12, 33–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekar, S.; Shafie, S.R.; Prasadam, I.; Crawford, R.; Panchal, S.K.; Brown, L.; Xiao, Y. Saturated fatty acids induce development of both metabolic syndrome and osteoarthritis in rats. Sci. Rep. 2017, 7, 46457. [Google Scholar] [CrossRef] [Green Version]
- Unger, A.L.; Torres-Gonzalez, M.; Kraft, J. Dairy Fat Consumption and the Risk of Metabolic Syndrome: An Examination of the Saturated Fatty Acids in Dairy. Nutrients 2019, 11, 2200. [Google Scholar] [CrossRef] [Green Version]
- Praagman, J.; Vissers, L.E.T.; Mulligan, A.A.; Laursen, A.S.D.; Beulens, J.W.J.; van der Schouw, Y.T.; Wareham, N.J.; Hansen, C.P.; Khaw, K.T.; Jakobsen, M.U.; et al. Consumption of individual saturated fatty acids and the risk of myocardial infarction in a UK and a Danish cohort. Int. J. Cardiol. 2019, 279, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Korbecki, J.; Bajdak-Rusinek, K. The effect of palmitic acid on inflammatory response in macrophages: An overview of molecular mechanisms. Inflamm. Res. 2019, 68, 915–932. [Google Scholar] [CrossRef] [Green Version]
- Carta, G.; Murru, E.; Banni, S.; Manca, C. Palmitic Acid: Physiological Role, Metabolism and Nutritional Implications. Front. Physiol. 2017, 8, 902. [Google Scholar] [CrossRef] [Green Version]
- Puri, P.; Wiest, M.M.; Cheung, O.; Mirshahi, F.; Sargeant, C.; Min, H.K.; Contos, M.J.; Sterling, R.K.; Fuchs, M.; Zhou, H.; et al. The plasma lipidomic signature of nonalcoholic steatohepatitis. Hepatology 2009, 50, 1827–1838. [Google Scholar] [CrossRef] [Green Version]
- Chiappini, F.; Coilly, A.; Kadar, H.; Gual, P.; Tran, A.; Desterke, C.; Samuel, D.; Duclos-Vallee, J.C.; Touboul, D.; Bertrand-Michel, J.; et al. Metabolism dysregulation induces a specific lipid signature of nonalcoholic steatohepatitis in patients. Sci. Rep. 2017, 7, 46658. [Google Scholar] [CrossRef] [Green Version]
- Chiappini, F.; Desterke, C.; Bertrand-Michel, J.; Guettier, C.; Le Naour, F. Hepatic and serum lipid signatures specific to nonalcoholic steatohepatitis in murine models. Sci. Rep. 2016, 6, 31587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rioux, V.; Pedrono, F.; Legrand, P. Regulation of mammalian desaturases by myristic acid: N-terminal myristoylation and oTher. modulations. Biochim. Biophys. Acta 2011, 1811, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Udenwobele, D.I.; Su, R.C.; Good, S.V.; Ball, T.B.; Varma Shrivastav, S.; Shrivastav, A. Myristoylation: An Important Protein Modification in the Immune Response. Front. Immunol. 2017, 8, 751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perreault, M.; Zulyniak, M.A.; Badoud, F.; Stephenson, S.; Badawi, A.; Buchholz, A.; Mutch, D.M. A distinct fatty acid profile underlies the reduced inflammatory state of metabolically healthy obese individuals. PLoS ONE 2014, 9, e88539. [Google Scholar] [CrossRef] [Green Version]
- Raatz, S.K.; Conrad, Z.; Johnson, L.K.; Picklo, M.J.; Jahns, L. Relationship of the Reported Intakes of Fat and Fatty Acids to Body Weight in US Adults. Nutrients 2017, 9, 438. [Google Scholar] [CrossRef] [Green Version]
- Aristizabal, J.C.; Barona, J.; Gonzalez-Zapata, L.I.; Deossa, G.C.; Estrada, A. Fatty Acid Content of Plasma Triglycerides May Contribute to the Heterogeneity in the Relationship Between Abdominal Obesity and the Metabolic Syndrome. Metab Syndr. Relat. Disord. 2016, 14, 311–317. [Google Scholar] [CrossRef]
- Masquio, D.C.; de Piano-Ganen, A.; Oyama, L.M.; Campos, R.M.; Santamarina, A.B.; de Souza, G.I.; Gomes, A.D.; Moreira, R.G.; Corgosinho, F.C.; do Nascimento, C.M.; et al. The role of free fatty acids in the inflammatory and cardiometabolic profile in adolescents with metabolic syndrome engaged in interdisciplinary therapy. J. Nutr. Biochem. 2016, 33, 136–144. [Google Scholar] [CrossRef]
- Mukwevho, E.; Joseph, J.S. Calmodulin dependent protein kinase II activation by exercise regulates saturated & unsaturated fatty acids and improves some metabolic syndrome markers. Life Sci. 2014, 111, 53–61. [Google Scholar] [CrossRef]
- Temme, E.H.; Mensink, R.P.; Hornstra, G. Effects of medium chain fatty acids (MCFA), myristic acid, and oleic acid on serum lipoproteins in healthy subjects. J. Lipid Res. 1997, 38, 1746–1754. [Google Scholar] [CrossRef]
- Snook, J.T.; Park, S.; Williams, G.; Tsai, Y.H.; Lee, N. Effect of synthetic triglycerides of myristic, palmitic, and stearic acid on serum lipoprotein metabolism. Eur. J. Clin. Nutr. 1999, 53, 597–605. [Google Scholar] [CrossRef] [Green Version]
- Zock, P.L.; de Vries, J.H.; Katan, M.B. Impact of myristic acid versus palmitic acid on serum lipid and lipoprotein levels in healthy women and men. Arterioscler. Thromb. 1994, 14, 567–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajri, T.; Khosla, P.; Pronczuk, A.; Hayes, K.C. Myristic acid-rich fat raises plasma LDL by stimulating LDL production without affecting fractional clearance in gerbils fed a cholesterol-free diet. J. Nutr. 1998, 128, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Khosla, P.; Hajri, T.; Pronczuk, A.; Hayes, K.C. Decreasing dietary lauric and myristic acids improves plasma lipids more favorably than decreasing dietary palmitic acid in rhesus monkeys fed AHA step 1 type diets. J. Nutr. 1997, 127, 525S–530S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tholstrup, T.; Marckmann, P.; Jespersen, J.; Vessby, B.; Jart, A.; Sandstrom, B. Effect on blood lipids, coagulation, and fibrinolysis of a fat high in myristic acid and a fat high in palmitic acid. Am. J. Clin. Nutr. 1994, 60, 919–925. [Google Scholar] [CrossRef] [Green Version]
- Takato, T.; Iwata, K.; Murakami, C.; Wada, Y.; Sakane, F. Chronic administration of myristic acid improves hyperglycaemia in the Nagoya-Shibata-Yasuda mouse model of congenital type 2 diabetes. Diabetologia 2017, 60, 2076–2083. [Google Scholar] [CrossRef] [Green Version]
- Stakos, D.A.; Boudoulas, K.D.; Gaillard, T.R.; Schuster, D.P.; Osei, K.; Boudoulas, H. Regional and overall aortic function in nondiabetic individuals with insulin resistance and normal glucose tolerance. J. Clin. Endocrinol. Metab 2013, 98, 4457–4463. [Google Scholar] [CrossRef] [Green Version]
- Gopal, T.; Kumar, N.; Perriotte-Olson, C.; Casey, C.A.; Donohue, T.M., Jr.; Harris, E.N.; Talmon, G.; Kabanov, A.V.; Saraswathi, V. Nanoformulated SOD1 ameliorates the combined NASH and alcohol-associated liver disease partly via regulating CYP2E1 expression in adipose tissue and liver. Am. J. Physiol. Gastrointest Liver Physiol. 2020, 318, G428–G438. [Google Scholar] [CrossRef]
- Perriotte-Olson, C.; Adi, N.; Manickam, D.S.; Westwood, R.A.; Desouza, C.V.; Natarajan, G.; Crook, A.; Kabanov, A.V.; Saraswathi, V. Nanoformulated copper/zinc superoxide dismutase reduces adipose inflammation in obesity. Obesity 2016, 24, 148–156. [Google Scholar] [CrossRef] [Green Version]
- Natarajan, G.; Perriotte-Olson, C.; Bhinderwala, F.; Powers, R.; Desouza, C.V.; Talmon, G.A.; Yuhang, J.; Zimmerman, M.C.; Kabanov, A.V.; Saraswathi, V. Nanoformulated copper/zinc superoxide dismutase exerts differential effects on glucose vs lipid homeostasis depending on the diet composition possibly via altered AMPK signaling. Transl. Res. 2017, 188, 10–26. [Google Scholar] [CrossRef]
- Saraswathi, V.; Hasty, A.H. The role of lipolysis in mediating the proinflammatory effects of very low density lipoproteins in mouse peritoneal macrophages. J. Lipid Res. 2006, 47, 1406–1415. [Google Scholar] [CrossRef] [Green Version]
- Saraswathi, V.; Kumar, N.; Gopal, T.; Bhatt, S.; Ai, W.; Ma, C.; Talmon, G.A.; Desouza, C. Lauric Acid versus Palmitic Acid: Effects on Adipose Tissue Inflammation, Insulin Resistance, and Non-Alcoholic Fatty Liver Disease in Obesity. Biology 2020, 9, 346. [Google Scholar] [CrossRef] [PubMed]
- Oosterveer, M.H.; van Dijk, T.H.; Tietge, U.J.; Boer, T.; Havinga, R.; Stellaard, F.; Groen, A.K.; Kuipers, F.; Reijngoud, D.J. High fat feeding induces hepatic fatty acid elongation in mice. PLoS ONE 2009, 4, e6066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, S.B.; Baicu, C.F.; Van Laer, A.; Geng, T.; Kasiganesan, H.; Zile, M.R.; Cowart, L.A. Ceramide synthase 5 mediates lipid-induced autophagy and hypertrophy in cardiomyocytes. J. Clin. Invest. 2012, 122, 3919–3930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubbard, N.E.; Socolich, R.J.; Erickson, K.L. Dietary myristic acid alters acylated proteins in activated murine macrophages. J. Nutr. 1996, 126, 1563–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, L.; Torres, S.; Baulies, A.; Alarcon-Vila, C.; Elena, M.; Fabrias, G.; Casas, J.; Caballeria, J.; Fernandez-Checa, J.C.; Garcia-Ruiz, C. Myristic acid potentiates palmitic acid-induced lipotoxicity and steatohepatitis associated with lipodystrophy by sustaning de novo ceramide synthesis. Oncotarget 2015, 6, 41479–41496. [Google Scholar] [CrossRef] [Green Version]
- Sampey, B.P.; Freemerman, A.J.; Zhang, J.; Kuan, P.F.; Galanko, J.A.; O’Connell, T.M.; Ilkayeva, O.R.; Muehlbauer, M.J.; Stevens, R.D.; Newgard, C.B.; et al. Metabolomic profiling reveals mitochondrial-derived lipid biomarkers that drive obesity-associated inflammation. PLoS ONE 2012, 7, e38812. [Google Scholar] [CrossRef]
- Ebbesson, S.O.; Tejero, M.E.; Lopez-Alvarenga, J.C.; Harris, W.S.; Ebbesson, L.O.; Devereux, R.B.; MacCluer, J.W.; Wenger, C.; Laston, S.; Fabsitz, R.R.; et al. Individual saturated fatty acids are associated with different components of insulin resistance and glucose metabolism: The GOCADAN study. Int. J. Circumpolar. Health 2010, 69, 344–351. [Google Scholar] [CrossRef]
- Kusunoki, M.; Tsutsumi, K.; Nakayama, M.; Kurokawa, T.; Nakamura, T.; Ogawa, H.; Fukuzawa, Y.; Morishita, M.; Koide, T.; Miyata, T. Relationship between serum concentrations of saturated fatty acids and unsaturated fatty acids and the homeostasis model insulin resistance index in Japanese patients with type 2 diabetes mellitus. J. Med. Invest. 2007, 54, 243–247. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Barnes, G.T.; Yang, Q.; Tan, G.; Yang, D.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Invest. 2003, 112, 1821–1830. [Google Scholar] [CrossRef]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Invest. 2003, 112, 1796–1808. [Google Scholar] [CrossRef]
- Funaki, M. Saturated fatty acids and insulin resistance. J. Med. Invest. 2009, 56, 88–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steppan, C.M.; Bailey, S.T.; Bhat, S.; Brown, E.J.; Banerjee, R.R.; Wright, C.M.; Patel, H.R.; Ahima, R.S.; Lazar, M.A. The hormone resistin links obesity to diabetes. Nature 2001, 409, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Muse, E.D.; Obici, S.; Bhanot, S.; Monia, B.P.; McKay, R.A.; Rajala, M.W.; Scherer, P.E.; Rossetti, L. Role of resistin in diet-induced hepatic insulin resistance. J. Clin. Invest. 2004, 114, 232–239. [Google Scholar] [CrossRef]
- Hayes, K.C.; Khosla, P.; Hajri, T.; Pronczuk, A. Saturated fatty acids and LDL receptor modulation in humans and monkeys. Prostaglandins Leukot Essent Fat. Acids 1997, 57, 411–418. [Google Scholar] [CrossRef]
- Noto, D.; Fayer, F.; Cefalu, A.B.; Altieri, I.; Palesano, O.; Spina, R.; Valenti, V.; Pitrone, M.; Pizzolanti, G.; Barbagallo, C.M.; et al. Myristic acid is associated to low plasma HDL cholesterol levels in a Mediterranean population and increases HDL catabolism by enhancing HDL particles trapping to cell surface proteoglycans in a liver hepatoma cell model. Atherosclerosis 2016, 246, 50–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, E.A.; Kahl, S.; Seelig, A.; Begovatz, P.; Irmler, M.; Kupriyanova, Y.; Nowotny, B.; Nowotny, P.; Herder, C.; Barosa, C.; et al. Acute dietary fat intake initiates alterations in energy metabolism and insulin resistance. J. Clin. Invest. 2017, 127, 695–708. [Google Scholar] [CrossRef]
- Ogawa, Y.; Imajo, K.; Honda, Y.; Kessoku, T.; Tomeno, W.; Kato, S.; Fujita, K.; Yoneda, M.; Saito, S.; Saigusa, Y.; et al. Palmitate-induced lipotoxicity is crucial for the pathogenesis of nonalcoholic fatty liver disease in cooperation with gut-derived endotoxin. Sci. Rep. 2018, 8, 11365. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Li, H.; Xu, H.; Halim, V.; Zhang, W.; Wang, H.; Ong, K.T.; Woo, S.L.; Walzem, R.L.; Mashek, D.G.; et al. Palmitoleate induces hepatic steatosis but suppresses liver inflammatory response in mice. PLoS ONE 2012, 7, e39286. [Google Scholar] [CrossRef]
- Benatar, J.R.; Stewart, R.A. The effects of changing dairy intake on trans and saturated fatty acid levels- results from a randomized controlled study. Nutr. J. 2014, 13, 32. [Google Scholar] [CrossRef] [Green Version]
- Khaw, K.T.; Sharp, S.J.; Finikarides, L.; Afzal, I.; Lentjes, M.; Luben, R.; Forouhi, N.G. Randomised trial of coconut oil, olive oil or butter on blood lipids and oTher. cardiovascular risk factors in healthy men and women. BMJ Open 2018, 8, e020167. [Google Scholar] [CrossRef] [Green Version]
- McKenney, J.M.; Proctor, J.D.; Wright, J.T., Jr.; Kolinski, R.J.; Elswick, R.K., Jr.; Coaker, J.S. The effect of supplemental dietary fat on plasma cholesterol levels in lovastatin-treated hypercholesterolemic patients. Pharmacotherapy 1995, 15, 565–572. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | High Fat | High Fat + MA |
| g | ||
| Protein | ||
| Casein | 200 | 200 |
| L-Cystine | 3 | 3 |
| Carbohydrate | ||
| Corn Starch | 72.8 | 72.8 |
| Maltodextrin 10 | 100 | 100 |
| Sucrose | 172.8 | 172.8 |
| Cellulose, BW200 | 50 | 50 |
| Fat | ||
| Soybean Oil | 25 | 25 |
| Lard | 177.5 | 151.7555 |
| Myristic Acid | 0 | 25.7445 |
| Minerals and Vitamins | ||
| Mineral Mix S10026 | 10 | 10 |
| DiCalcium Phosphate | 13 | 13 |
| Calcium Carbonate | 5.5 | 5.5 |
| Potassium Citrate, 1 H2O | 16.5 | 16.5 |
| Vitamin Mix V10001 | 10 | 10 |
| Choline Bitartrate | 2 | 2 |
| Total | 858.15 | 858.15 |
| Gene (Abbr) | Description | Catalog Number |
|---|---|---|
| 18S | 18S ribosomal RNA | 4352930E |
| Adgre1 (EMR-1; F4/80) | EGF-like module containing, mucin-like, hormone receptor-like 1 | Mm00802529_m1 |
| Adipoq | Adiponectin, C1Q, and collagen domain | Mm00456425_m1 |
| Ccl2 | Chemokine ligand 2/monocyte chemotactic protein 1 | Mm00441242_m1 |
| Ccl3 | Chemokine ligand 3/macrophage inflammatory protein 1alpha | Mm00441258_m1 |
| Ccr2 | Chemokine receptor/monocyte chemotactic protein 1 receptor | Mm99999051_gH |
| Chil3 | Chitinase-like 3 | Mm00657889_mH |
| Clec10a | C-type lectin domain family 10, member A | Mm00546125_g1 |
| Elovl6 | ELOVL fatty acid elongase 6 | Mm00851223_s1 |
| Fads1 | Fatty acid desaturase 1 | Mm00507605_m1 |
| Fads2 | Fatty acid desaturase 2 | Mm00517221_m1 |
| Fasn | Fatty acid synthase | Mm01253292_m1 |
| Il6 | Interleukin 6 | Mm00446190_m1 |
| Il10 | Interleukin 10 | Mm99999062_m1 |
| Lep | Leptin | Mm00434759_m1 |
| Mgl2 | Macrophage galactose N-acetyl-galactosamine–specific lectins 2 | Mm00460844_m1 |
| Mmp3 | Matrix metallopeptidase 3 | Mm00440295_m1 |
| Mmp12 | Matrix metallopeptidase 12 | Mm00500554_m1 |
| Retn | Resistin | Mm00445641_m1 |
| Saa3 | Serum amyloid A3 | Mm00441203_m1 |
| Scd1 | Stearoyl-CoA desaturase 1 | Mm00772290_m1 |
| Tnf | Tumor necrosis factor, alpha | Mm00443258_m1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saraswathi, V.; Kumar, N.; Ai, W.; Gopal, T.; Bhatt, S.; Harris, E.N.; Talmon, G.A.; Desouza, C.V. Myristic Acid Supplementation Aggravates High Fat Diet-Induced Adipose Inflammation and Systemic Insulin Resistance in Mice. Biomolecules 2022, 12, 739. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12060739
Saraswathi V, Kumar N, Ai W, Gopal T, Bhatt S, Harris EN, Talmon GA, Desouza CV. Myristic Acid Supplementation Aggravates High Fat Diet-Induced Adipose Inflammation and Systemic Insulin Resistance in Mice. Biomolecules. 2022; 12(6):739. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12060739
Chicago/Turabian StyleSaraswathi, Viswanathan, Narendra Kumar, Weilun Ai, Thiyagarajan Gopal, Saumya Bhatt, Edward N. Harris, Geoffrey A. Talmon, and Cyrus V. Desouza. 2022. "Myristic Acid Supplementation Aggravates High Fat Diet-Induced Adipose Inflammation and Systemic Insulin Resistance in Mice" Biomolecules 12, no. 6: 739. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12060739