
A Novel Antiviral Protein Derived from Oenanthe javanica: Type I Interferon-Dependent Antiviral Signaling and Its Pharmacological Potential

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Expression of Full-Length OPRP
2.2. Synthesis of Small Peptides
2.3. Construction of Recombinant Plasmid pcDNA3.1(+)/OPRP
2.4. Screening of Stable Expression of Raw264.7 Cells Transfected with pcDNA3.1(+)/OPRP
2.5. Cell Culture and Reagents
2.6. Preparation of Splenocytes
2.7. CD4+ Cells
2.8. RNA Isolation and qPCR
2.9. Western Blot Analysis and Co-Immunoprecipitation
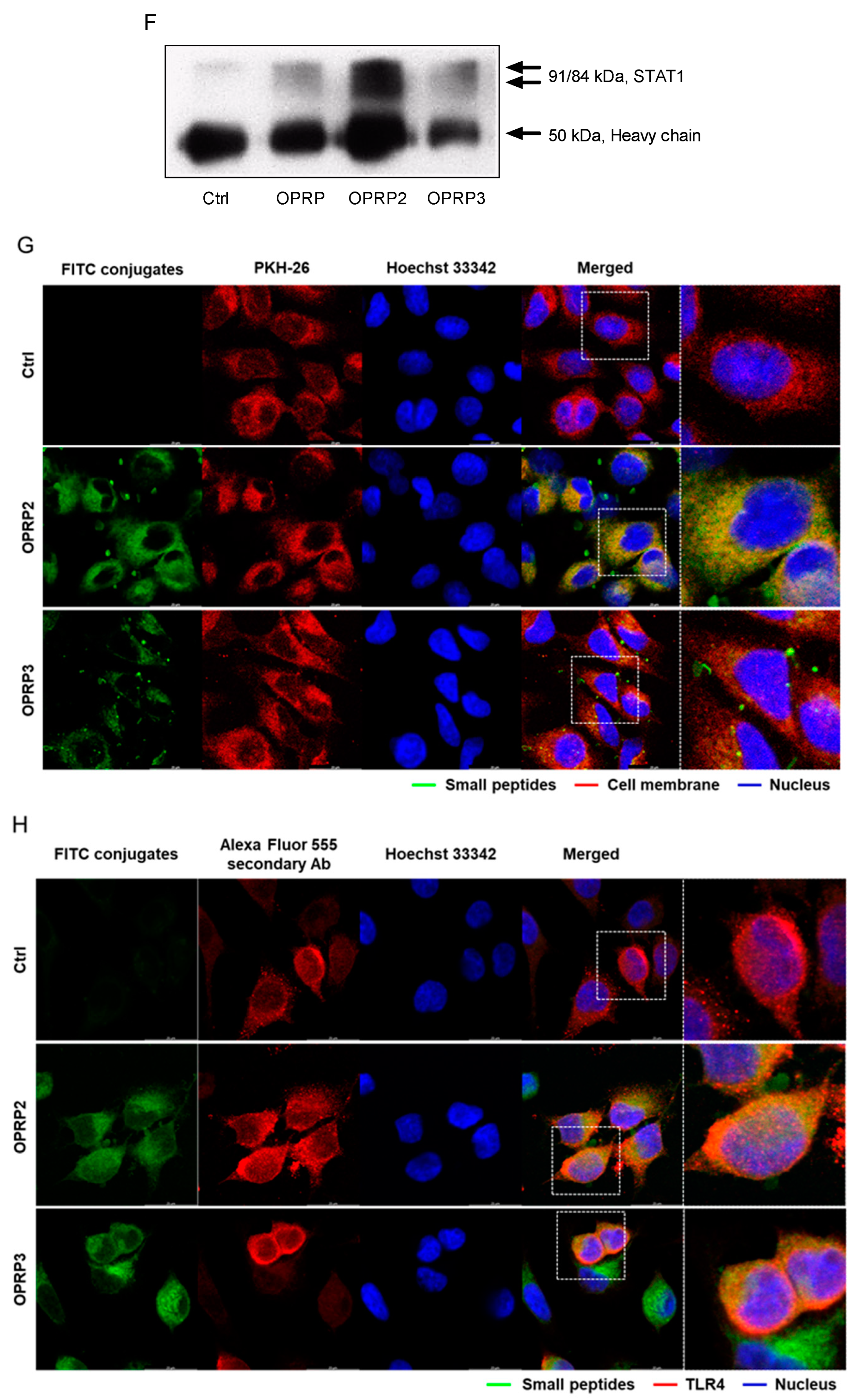
2.10. Immunocytochemistry (ICC)
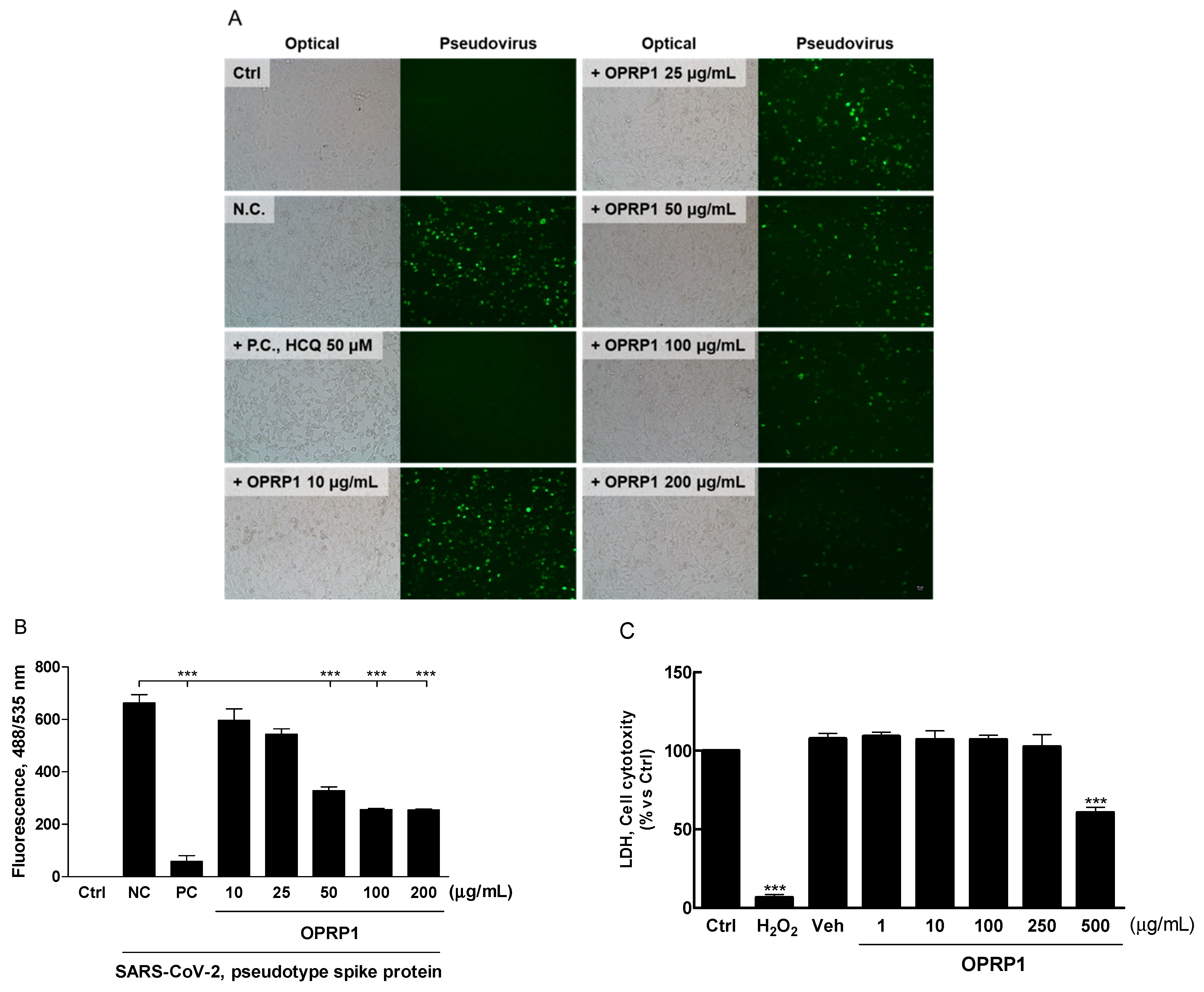
2.11. Inhibition of SARS-CoV-2 Pseudovirus Assay
2.12. Statistical Analysis
3. Results
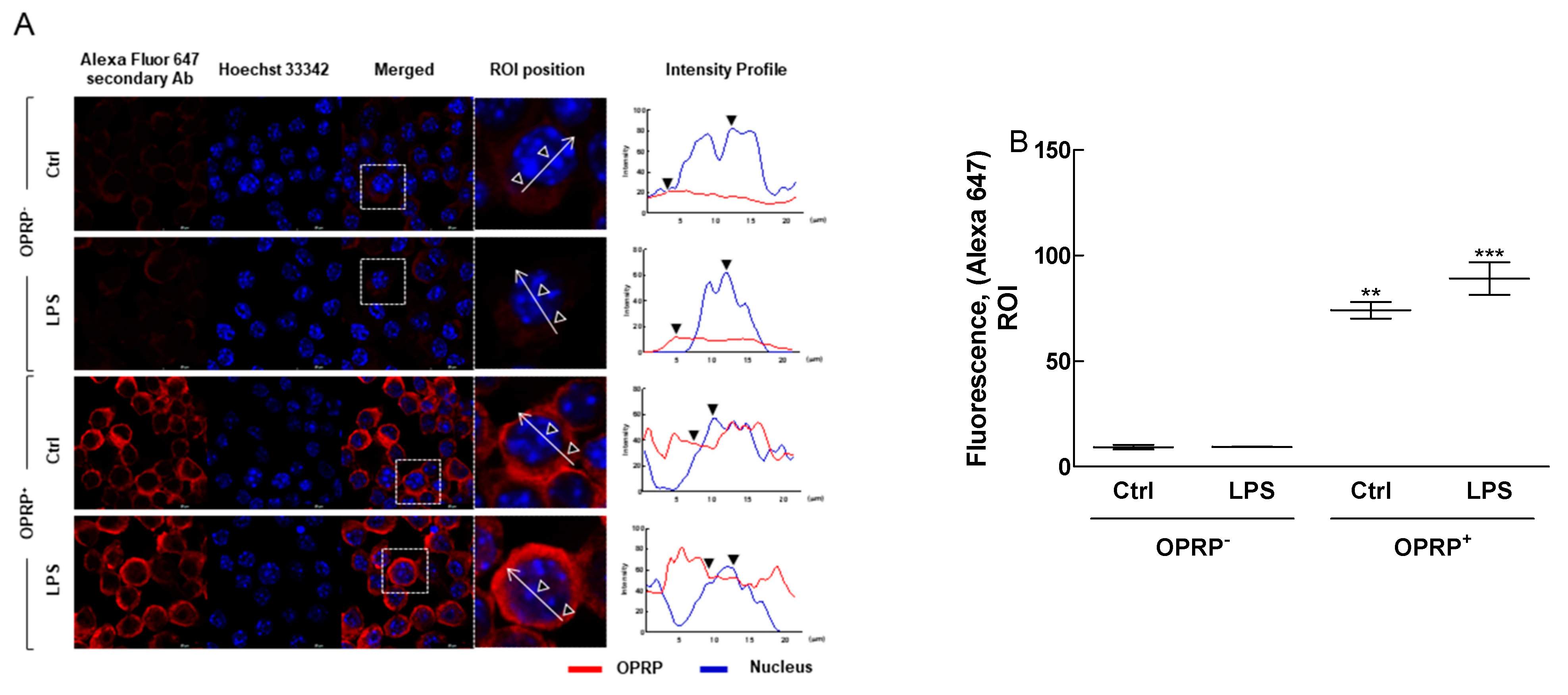
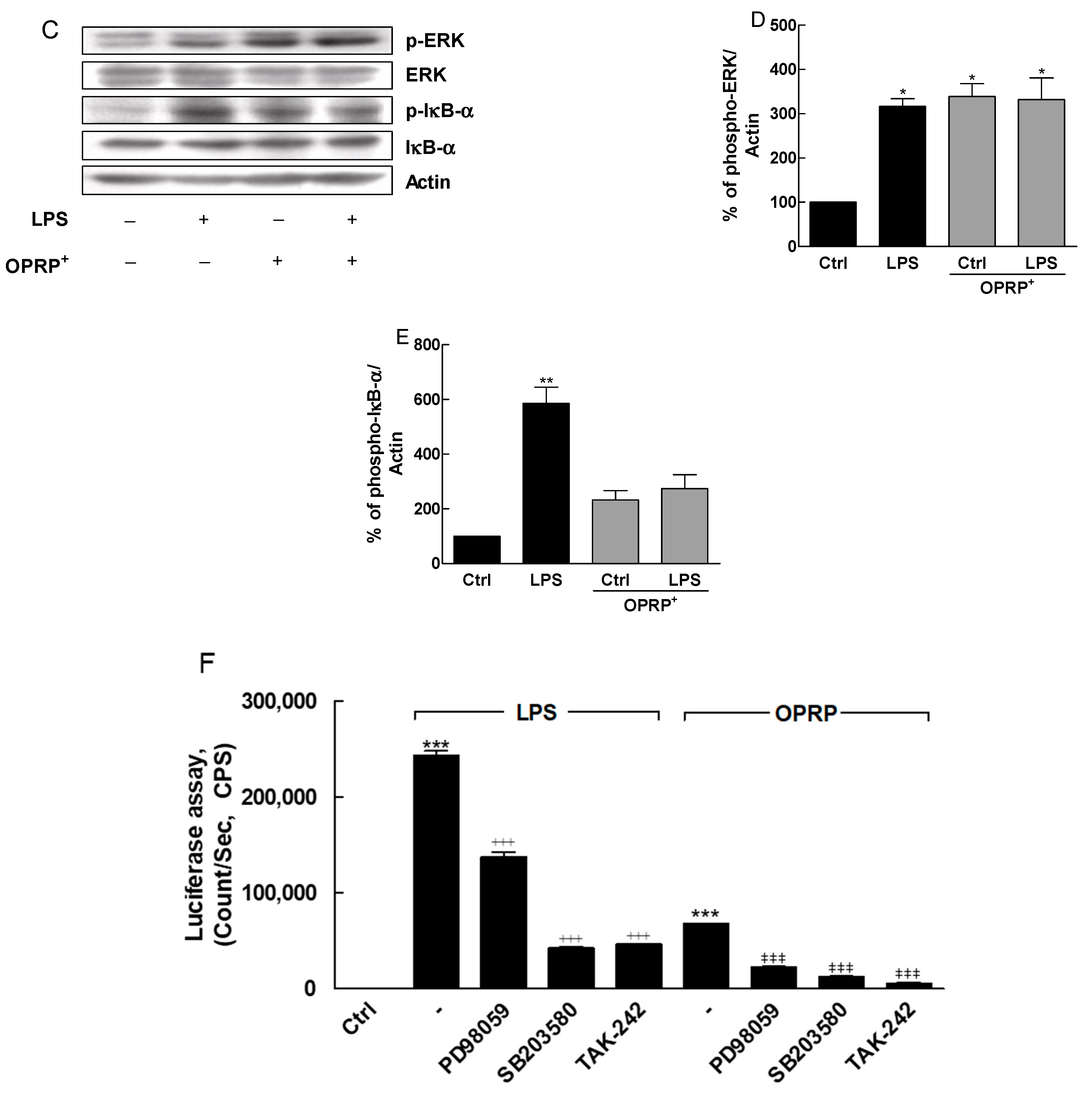
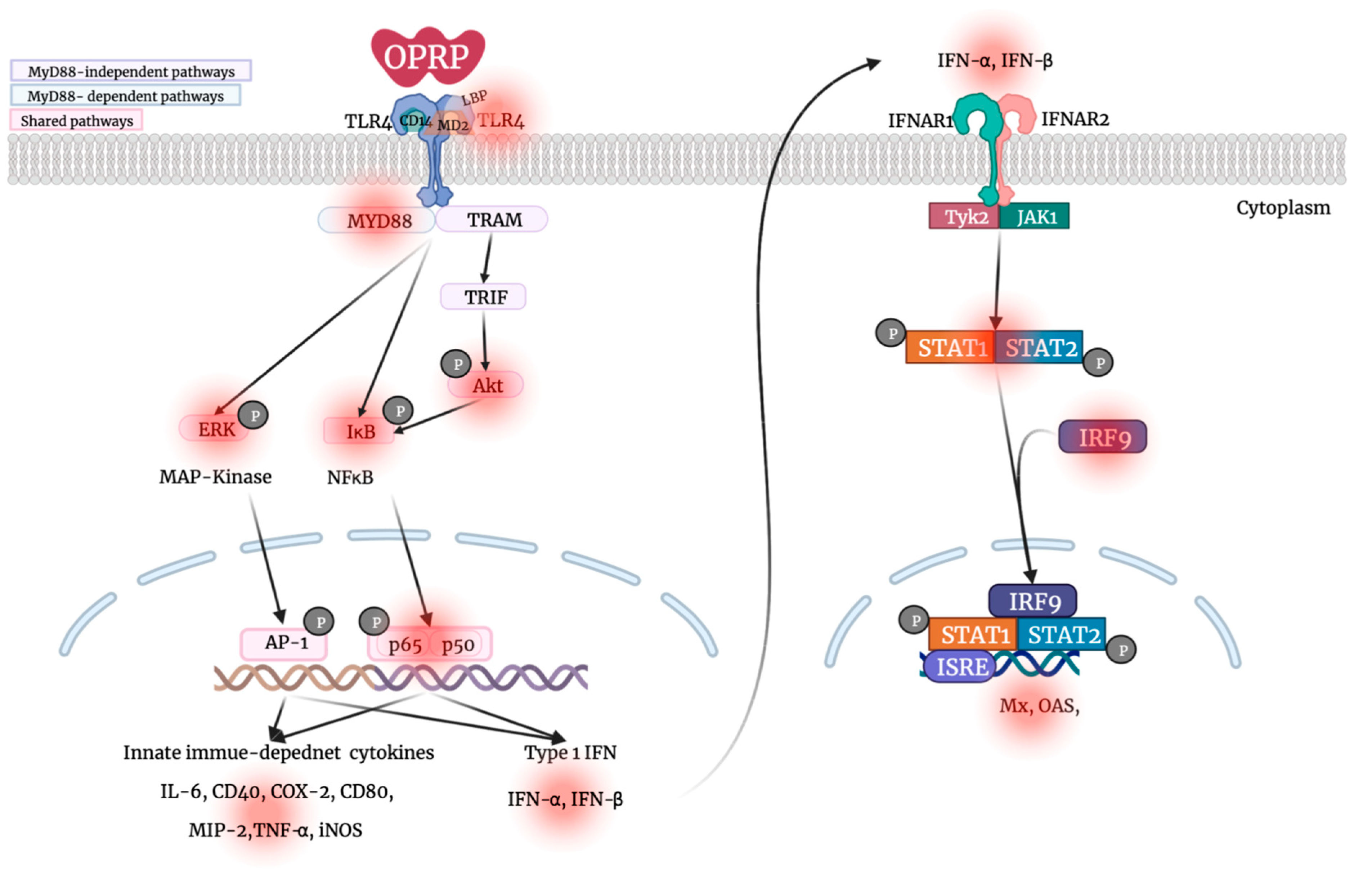
3.1. TLR4-Dependent Signaling Pathway in Macrophages
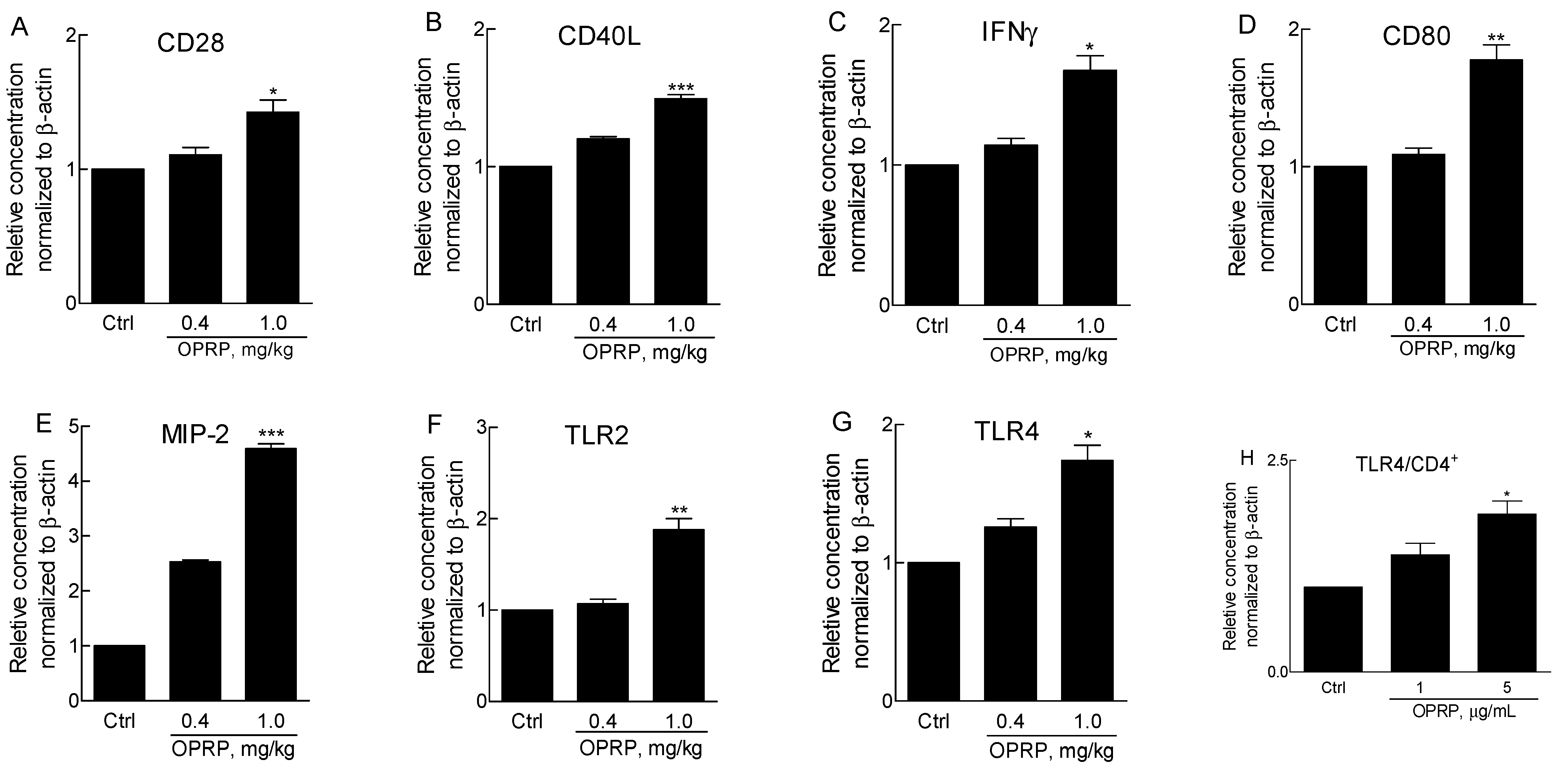
3.2. Activation of Splenocytes and TLR Signaling in CD4+ Cells
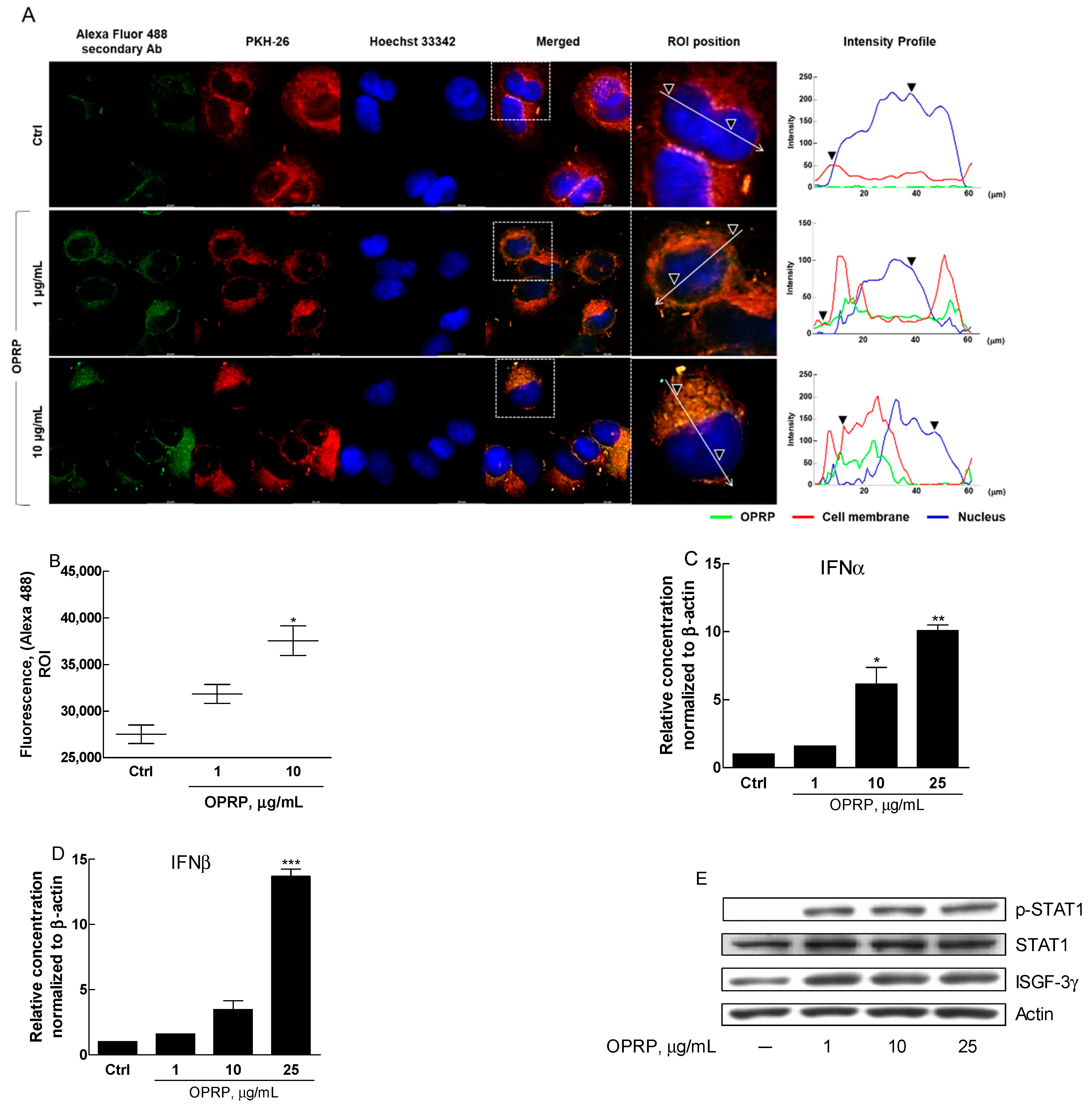
3.3. OPRP-Mediated Cellular Responses
3.4. STAT1/STAT2-Dependent Signaling
3.5. Inhibition of SARS-CoV-2 Pseudo-Virus Entry
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Beutler, B.; Jiang, Z.; Georgel, P.; Crozat, K.; Croker, B.; Rutschmann, S.; Du, X.; Hoebe, K. Genetic analysis of host resistance: Toll-like receptor signaling and immunity at large. Annu. Rev. Immunol. 2006, 24, 353–389. [Google Scholar] [CrossRef] [Green Version]
- Medzhitov, R. Recognition of microorganisms and activation of the immune response. Nature 2007, 449, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. Toll-like receptor and RIG-I-like receptor signaling. Ann. N. Y. Acad. Sci. 2008, 1143, 1–20. [Google Scholar] [CrossRef]
- Vance, R.E.; Isberg, R.R.; Portnoy, D.A. Patterns of pathogenesis: Discrimination of pathogenic and nonpathogenic microbes by the innate immune system. Cell Host Microbe 2009, 6, 10–21. [Google Scholar] [CrossRef] [Green Version]
- Bol, J.F.; Linthorst, H.J.; Cornelissen, B.J. Plant pathogenesis-related proteins induced by virus infection. Annu. Rev. Phytopathol. 1990, 28, 113–138. [Google Scholar] [CrossRef]
- Chintamanani, S.; Hulbert, S.H.; Johal, G.S.; Balint-Kurti, P.J. Identification of a maize locus that modulates the hypersensitive defense response, using mutant-assisted gene identification and characterization. Genetics 2010, 184, 813–825. [Google Scholar] [CrossRef] [Green Version]
- Coll, N.S.; Epple, P.; Dangl, J.L. Programmed cell death in the plant immune system. Cell Death Differ. 2011, 18, 1247–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitham, S.A.; Qi, M.; Innes, R.W.; Ma, W.; Lopes-Caitar, V.; Hewezi, T. Molecular Soybean-Pathogen Interactions. Annu. Rev. Phytopathol. 2016, 54, 443–468. [Google Scholar] [CrossRef] [PubMed]
- Midoro-Horiuti, T.; Brooks, E.G.; Goldblum, R.M. Pathogenesis-related proteins of plants as allergens. Ann. Allergy Asthma Immunol. 2001, 87, 261–271. [Google Scholar] [CrossRef]
- Christensen, A.B.; Cho, B.H.; Naesby, M.; Gregersen, P.L.; Brandt, J.; Madriz-Ordenana, K.; Collinge, D.B.; Thordal-Christensen, H. The molecular characterization of two barley proteins establishes the novel PR-17 family of pathogenesis-related proteins. Mol. Plant Pathol. 2002, 3, 135–144. [Google Scholar] [CrossRef]
- Colditz, F.; Nyamusuren, O.; Niehaus, K.; Eubel, H.; Braun, H.P.; Krajinski, F. Proteomic approach: Identification of Medicago truncatula, proteins included in roots after infection with the pathogenic oomycete Aphanomyces euteiches. Plant Mol. Biol. 2004, 55, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Sels, J.; Mathys, J.; De Coninck, B.M.; Cammue, B.P.; De Bolle, M.F. Plant pathogenesis-related (PR) proteins: A focus on PR peptides. Plant Physiol. Biochem. 2008, 46, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Gajera, H.; Domadiya, R.; Patel, S.; Kapopara, M.; Golakiva, B. Molecular mechanism of trichoderma as bio-control agents against phytopathogen system-a revies. Curr. Res. Microbiol. Biotechnol. 2013, 1, 133–142. [Google Scholar]
- Moosa, A.; Farzand, A.; Sahi, S.T.; Khan, S.A. Transgenic expression of antifungal pathogenesis-related proteins against phytopathogenic fungi-15 years of success. Isr. J. Plant Sci. 2018, 65, 38–54. [Google Scholar] [CrossRef]
- Van Loon, L.C.; Van Strien, E.A. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar] [CrossRef]
- Jo, B.R.; Yu, J.M.; Jang, S.; Ahn, J.W.; Kim, H.S.; Seoung, E.A.; Park, H.Y.; Jin, D.H.; Joo, S.S. Cloning, Expression, and Purification of a Pathogenesis-Related Protein from Oenanthe javanica and Its Biological Properties. Biol. Pharm. Bull. 2020, 43, 158–168. [Google Scholar] [CrossRef] [Green Version]
- Schwandner, R.; Dziarski, R.; Wesche, H.; Rothe, M.; Kirschning, C.J. Peptidoglycan- and lipoteichoic acid-induced cell activation is mediated by toll-like receptor 2. J. Biol. Chem. 1999, 274, 17406–17409. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, M.; Asai, Y.; Ogawa, T. Separation and structural analysis of lipoprotein in a lipopolysaccharide preparation from Porphyromonas gingivalis. Int. Immunol. 2004, 16, 1431–1437. [Google Scholar] [CrossRef]
- Hardiman, G.; Rock, F.L.; Balasubramanian, S.; Kastelein, R.A.; Bazan, J.F. Molecular characterization and modular analysis of human MyD88. Oncogene 1996, 13, 2467–2475. [Google Scholar]
- Burns, K.; Martinon, F.; Esslinger, C.; Pahl, H.; Schneider, P.; Bodmer, J.L.; Di Marco, F.; French, L.; Tschopp, J. MyD88, an adapter protein involved in interleukin-1 signaling. J. Biol. Chem. 1998, 273, 12203–12209. [Google Scholar] [CrossRef] [Green Version]
- Medzhitov, R.; Preston-Hurlburt, P.; Kopp, E.; Stadlen, A.; Chen, C.; Ghosh, S.; Janeway, C.A., Jr. MyD88 is an adaptor protein in the hToll/IL-1 receptor family signaling pathways. Mol. Cell 1998, 2, 253–258. [Google Scholar] [CrossRef]
- O’Neill, L.A.; Bowie, A.G. The family of five: TIR-domain-containing adaptors in Toll-like receptor signalling. Nat. Rev. Immunol. 2007, 7, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Marie, I.; Durbin, J.E.; Levy, D.E. Differential viral induction of distinct interferon-alpha genes by positive feedback through interferon regulatory factor-7. EMBO J. 1998, 17, 6660–6669. [Google Scholar] [CrossRef] [PubMed]
- Tailor, P.; Tamura, T.; Kong, H.J.; Kubota, T.; Kubota, M.; Borghi, P.; Gabriele, L.; Ozato, K. The feedback phase of type I interferon induction in dendritic cells requires interferon regulatory factor 8. Immunity 2007, 27, 228–239. [Google Scholar] [CrossRef] [Green Version]
- Mlcochova, P.; Winstone, H.; Zuliani-Alvarez, L.; Gupta, R.K. TLR4-Mediated Pathway Triggers Interferon-Independent G0 Arrest and Antiviral SAMHD1 Activity in Macrophages. Cell Rep. 2020, 30, 3972–3980.e5. [Google Scholar] [CrossRef]
- Muller, U.; Steinhoff, U.; Reis, L.F.; Hemmi, S.; Pavlovic, J.; Zinkernagel, R.M.; Aguet, M. Functional role of type I and type II interferons in antiviral defense. Science 1994, 264, 1918–1921. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.Y.; Hertzog, P.J.; Holland, K.A.; Sumarsono, S.H.; Tymms, M.J.; Hamilton, J.A.; Whitty, G.; Bertoncello, I.; Kola, I. A null mutation in the gene encoding a type I interferon receptor component eliminates antiproliferative and antiviral responses to interferons alpha and beta and alters macrophage responses. Proc. Natl. Acad. Sci. USA 1995, 92, 11284–11288. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.A.; Jung, Y.S.; Lee, S.J.; Park, S.C.; Kim, M.J.; Lee, E.J.; Byun, H.J.; Jhee, K.H.; Lee, S.P. Hepatoprotective effects of fermented field water-dropwort (Oenanthe javanica) extract and its major constituents. Food Chem. Toxicol. 2014, 67, 154–160. [Google Scholar] [CrossRef]
- Sim, H.J.; Kim, S.M.; Jeon, Y.J.; Lee, Y.E. Antioxidant activity of dropwort (Oenanthe javanica DC) fermented extract and its hepatoprotective effect against alcohol in rats. J. Korean Soc. Food Cult. 2015, 30, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Ai, G.; Huang, Z.M.; Liu, Q.C.; Han, Y.Q.; Chen, X. The protective effect of total phenolics from Oenanthe Javanica on acute liver failure induced by D-galactosamine. J. Ethnopharmacol. 2016, 186, 53–60. [Google Scholar] [CrossRef]
- Park, J.C.; Yu, Y.B.; Lee, J.H.; Hattori, M.; Lee, C.K.; Choi, J.W. Protective effect of Oenanthe javanica on the hepatic lipid peroxidation in bromobenzene-treated rats and its bioactive component. Planta Med. 1996, 62, 488–490. [Google Scholar] [CrossRef] [PubMed]
- Park, J.C.; Choi, J.W. Effects of methanol extract of Oenanthe javanica on the hepatic alcohol-metabolizing enzyme system and its bioactive component. Phytother. Res. 1998, 11, 260–262. [Google Scholar] [CrossRef]
- Yang, J.H.; Kim, S.C.; Shin, B.Y.; Jin, S.H.; Jo, M.J.; Jegal, K.H.; Kim, Y.W.; Lee, J.R.; Ku, S.K.; Cho, I.J.; et al. O-Methylated flavonol isorhamnetin prevents acute inflammation through blocking of NF-kappaB activation. Food Chem. Toxicol. 2013, 59, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.H.; Cho, H.W.; Lee, B.Y.; Yoon, J.Y. Anti-inflammatory effects of Oenanthe javanica ethanol extract and its fraction on LPS-induced inflammation response. J. Korean Soc. Food Sci. Nutr. 2016, 45, 1595–1603. [Google Scholar] [CrossRef] [Green Version]
- Ahn, H.; Lee, G.S. Isorhamnetin and hyperoside derived from water dropwort inhibits inflammasome activation. Phytomedicine 2017, 24, 77–86. [Google Scholar] [CrossRef]
- Lu, C.L.; Li, X.F. A Review of Oenanthe javanica (Blume) DC. as Traditional Medicinal Plant and Its Therapeutic Potential. Evid. Based Complement. Altern. Med. 2019, 42, 482–485. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Kim, K.H.; Lee, Y.J.; Lee, S.H.; Park, J.C.; Nam, D.H. Oenanthe javanica extract accelerates ethanol metabolism in ethanol-treated animals. BMB Rep. 2009, 42, 482–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kongkachuichai, R.; Charoensiri, R.; Yakoh, K.; Kringkasemsee, A.; Insung, P. Nutrients value and antioxidant content of indigenous vegetables from Southern Thailand. Food Chem. 2015, 173, 838–846. [Google Scholar] [CrossRef]
- Han, Y.Q.; Huang, Z.M.; Yang, X.B.; Liu, H.Z.; Wu, G.X. In vivo and in vitro anti-hepatitis B virus activity of total phenolics from Oenanthe javanica. J. Ethnopharmacol. 2008, 118, 148–153. [Google Scholar] [CrossRef]
- Michalska, A.; Blaszczyk, K.; Wesoly, J.; Bluyssen, H.A.R. A Positive Feedback Amplifier Circuit That Regulates Interferon (IFN)-Stimulated Gene Expression and Controls Type I and Type II IFN Responses. Front. Immunol. 2018, 9, 1135. [Google Scholar] [CrossRef] [Green Version]
- Teoh, E.S. (Ed.) Secondary metabolites of plants. In Medicinal Orchids of Asia; Springer: Cham, Switzerland, 2016; pp. 59–73. [Google Scholar]
- Ali, S.; Ganai, B.A.; Kamili, A.N.; Bhat, A.A.; Mir, Z.A.; Bhat, J.A.; Tyagi, A.; Islam, S.T.; Mushtaq, M.; Yadav, P.; et al. Pathogenesis-related proteins and peptides as promising tools for engineering plants with multiple stress tolerance. Microbiol. Res. 2018, 212, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Finkina, E.I.; Melnikova, D.N.; Bogdanov, I.V.; Ovchinnikova, T.V. Plant Pathogenesis-Related Proteins PR-10 and PR-14 as Components of Innate Immunity System and Ubiquitous Allergens. Curr. Med. Chem. 2017, 24, 1772–1787. [Google Scholar] [CrossRef]
- Swain, S.L.; McKinstry, K.K.; Strutt, T.M. Expanding roles for CD4(+) T cells in immunity to viruses. Nat. Rev. Immunol. 2012, 12, 136–148. [Google Scholar] [CrossRef] [PubMed]
- Selby, L.I.; Aurelio, L.; Yuen, D.; Graham, B.; Johnston, A.P.R. Quantifying Cellular Internalization with a Fluorescent Click Sensor. ACS Sens. 2018, 3, 1182–1189. [Google Scholar] [CrossRef] [PubMed]
- Hung, W.S.; Ling, P.; Cheng, J.C.; Chang, S.S.; Tseng, C.P. Disabled-2 is a negative immune regulator of lipopolysaccharide-stimulated Toll-like receptor 4 internalization and signaling. Sci. Rep. 2016, 6, 35343. [Google Scholar] [CrossRef] [Green Version]
- Nitin, P.; Nandhakumar, R.; Vidhya, B.; Rajesh, S.; Sakunthala, A. COVID-19: Invasion, pathogenesis and possible cure—A review. J. Virol. Methods 2022, 300, 114434. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Gene | Direction | Sequence (5′ to 3′) | Accession |
|---|---|---|---|---|
| Mouse | CD28 | Forward | 5′-TACCTAGACAACGAGAGGAG | NM_007642.4 |
| Reverse | 5′-GTCACTTTGAAGGAGTCTGT | |||
| CD40L | Forward | 5′-CTAATCGGGAGCCTTCGAGT | NM_011616.2 | |
| Reverse | 5′-TGGATCACTTGGCTTGCTTC | |||
| CD80 | Forward | 5′-TATTGCTGCCTTGCCGTTAC | NM_001359898.1 | |
| Reverse | 5′-ACCAGGCCCAGGATGATAAG | |||
| IFN-γ | Forward | 5′-GTGACATGAAAATCCTGCAG | NM_008337.4 | |
| Reverse | 5′-GTTGTTGACCTCAAACTTGG | |||
| MIP2 | Forward | 5′-TTCCATTGCCCAGATGTTGT | NM_009140.2 | |
| Reverse | 5′-CTGTGTGGGTGGGATGTAGC | |||
| TLR2 | Forward | 5′-TCAGTGGCCAGAAAAGATGC | NM_011905.3 | |
| Reverse | 5′-ACCAGCAACACAGGGAACAA | |||
| TLR4 | Forward | 5′-CGCTCTGGCATCATCTTCAT | NM_021297.3 | |
| Reverse | 5′-TGTTTGCTCAGGATTCGAGG | |||
| IFN-α | Forward | 5′-ACCTGCAAGGCTGTCTGATG | NM_010502.2 | |
| Reverse | 5′-CAGTCTTCCCAGCACATTGG | |||
| IFN-β | Forward | 5′-GTTCCTGCTGTGCTTCTCCA | NM_010510.1 | |
| Reverse | 5′-CTTTCCATTCAGCTGCTCCA | |||
| Mx1 | Forward | 5′-GAGAGGCAAAGTCTCCTATG | NM_010846.1 | |
| Reverse | 5′-GTCAATGAGAGTCAGGTCTG | |||
| β-actin | Forward | 5′-TCCTGACCCTGAAGTACCCC | NM_007393.5 | |
| Reverse | 5′-ATGCCAGTGGTACGACCAGA | |||
| Human | IFN-α | Forward | 5′-GCACAGATGAGGAGAATCTC | NM_000605.4 |
| Reverse | 5′-TTGTCTAGGAGGGTCTCATC | |||
| IFN-β | Forward | 5′-GTCTCCTCCAAATTGCTCTC | NM_002176.4 | |
| Reverse | 5′-CCTCAGGGATGTCAAAGTTC | |||
| β-actin | Forward | 5′-GTCTGGTGCCTGGTCTGATG | NM_011198 | |
| Reverse | 5′-GGTTGAAAAGGAGCTCTGGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jo, B.-R.; Kim, H.-S.; Ahn, J.-W.; Jeoung, E.-Y.; Jang, S.-K.; Yoo, Y.-M.; Joo, S.-S. A Novel Antiviral Protein Derived from Oenanthe javanica: Type I Interferon-Dependent Antiviral Signaling and Its Pharmacological Potential. Biomolecules 2022, 12, 835. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12060835
Jo B-R, Kim H-S, Ahn J-W, Jeoung E-Y, Jang S-K, Yoo Y-M, Joo S-S. A Novel Antiviral Protein Derived from Oenanthe javanica: Type I Interferon-Dependent Antiviral Signaling and Its Pharmacological Potential. Biomolecules. 2022; 12(6):835. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12060835
Chicago/Turabian StyleJo, Bo-Ram, Hyun-Soo Kim, Jeong-Won Ahn, Eui-Young Jeoung, Su-Kil Jang, Yeong-Min Yoo, and Seong-Soo Joo. 2022. "A Novel Antiviral Protein Derived from Oenanthe javanica: Type I Interferon-Dependent Antiviral Signaling and Its Pharmacological Potential" Biomolecules 12, no. 6: 835. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12060835