
Alternative c-MYC mRNA Transcripts as an Additional Tool for c-Myc2 and c-MycS Production in BL60 Tumors

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
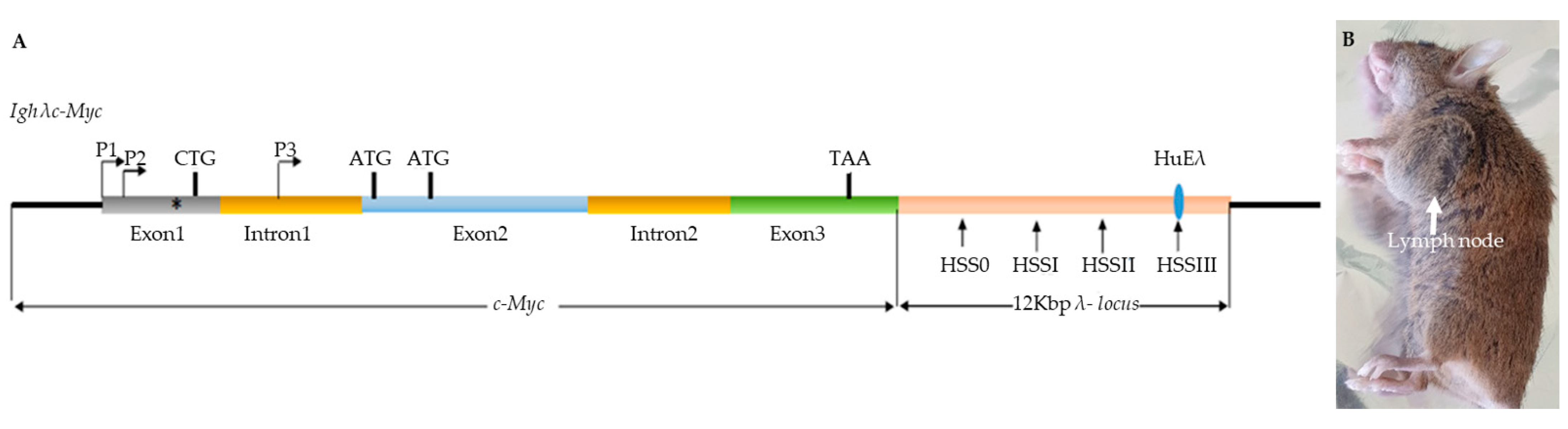
2.1. Mouse Models
2.2. Cell Lines
2.3. RNA Extraction
2.4. Identification of 5′UTR, 3′UTR and Full-Length of cDNA
2.5. In Vitro Expression of c-Myc1, c-Myc2 and c-MycS Reading Frames
2.6. Protein Extraction and Western Blot
3. Results
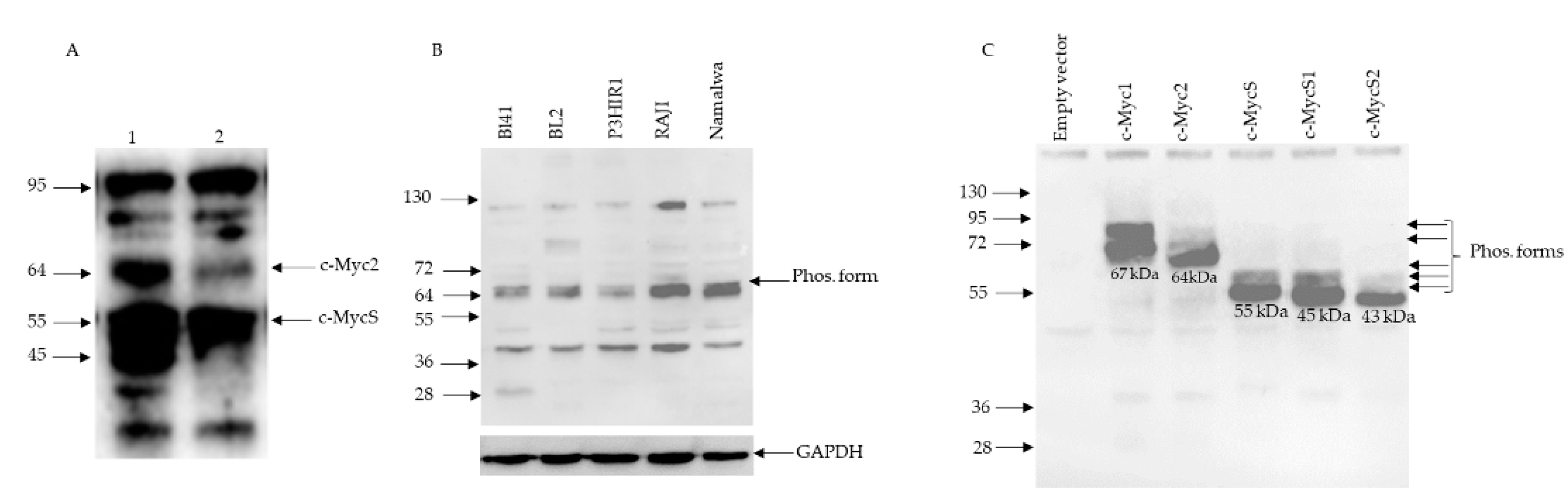
3.1. c-MYC Gene Translocation and Synthesis of c-Myc Proteins in Burkitt Lymphomas
3.2. c-Myc Protein Production by Cell-Free Translation of a Single Cloned Reading Frame
3.3. c-MYC Transcripts in Lymph Nodes from λc-MYC Mice
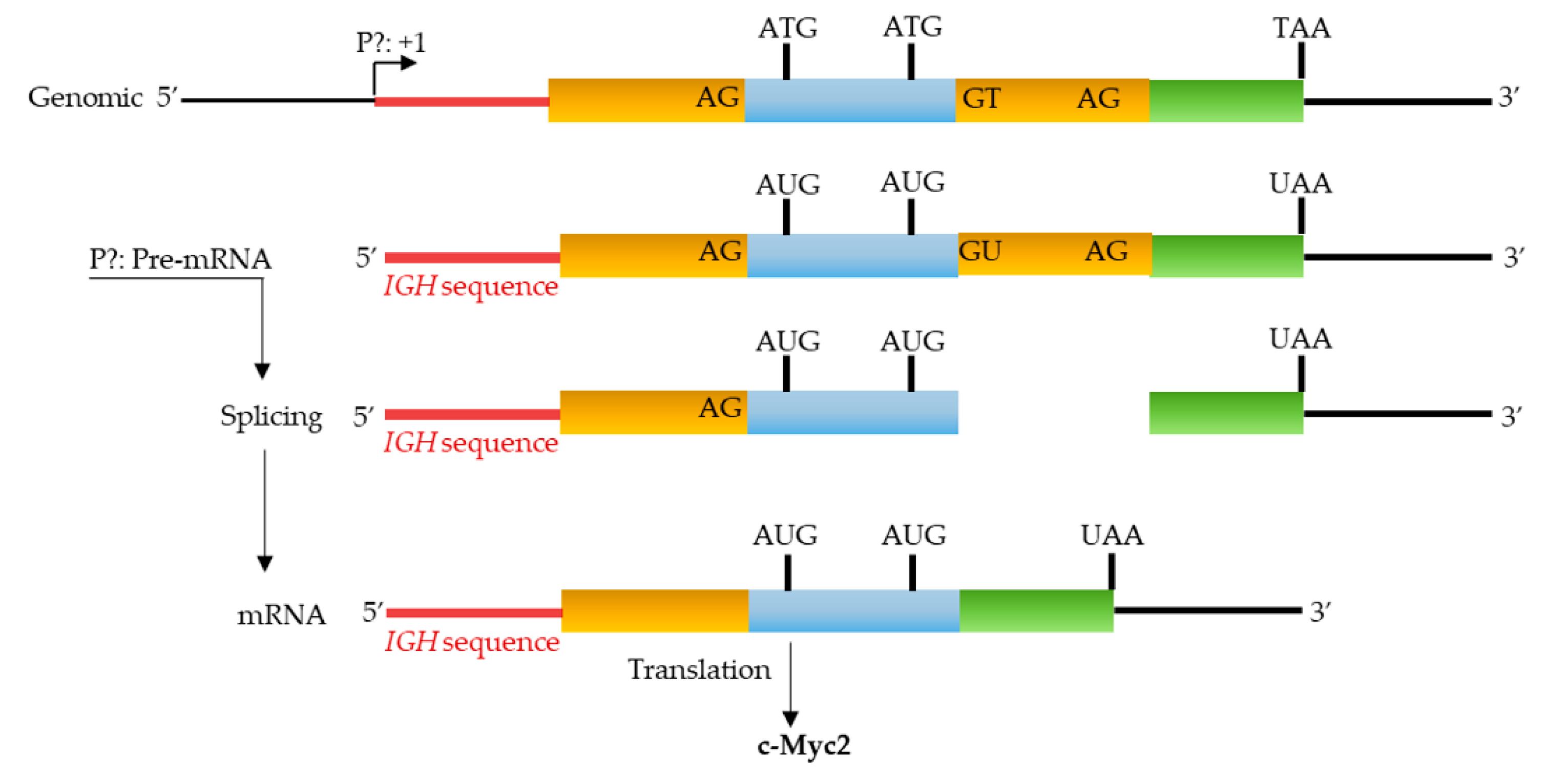
3.4. P1 Promoter Transcripts and c-Myc Proteins
3.5. P3 Promoter Transcripts and c-Myc Proteins
3.6. c-MYC Transcripts and c-Myc Proteins in the BL41 Cell Line
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Thompson, E.B. The many roles of c-Myc in apoptosis. Annu. Rev. Physiol. 1998, 60, 575–600. [Google Scholar] [CrossRef]
- Hann, S.R.; King, M.W.; Bentley, D.L.; Anderson, C.W.; Eisenman, R.N. A non-AUG translational initiation in c-myc exon 1 generates an N-terminally distinct protein whose synthesis is disrupted in Burkitt’s lymphomas. Cell 1988, 52, 185–195. [Google Scholar] [CrossRef]
- Spotts, G.D.; Patel, S.V.; Xiao, Q.; Han, S.R. Identification of downstream-initiated c-Myc proteins which are dominant- negative inhibitors of transactivation by full-length c-Myc proteins. Mol. Cell. Biol. 1997, 17, 1459–1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wierstra, I.; Alves, J. The c-myc promoter: Still MysterY and challenge. Adv. Cancer Res. 2008, 99, 113–333. [Google Scholar]
- Eick, D.; Polack, A.; Kofler, E.; Lenoir, G.M. Expression of P0-and P3-RNA from the normal and translocated c-myc allele in Burkitt’s lymphoma cells. Oncogene 1990, 5, 1397–1402. [Google Scholar]
- Hann, S.R.; Dixit, M.; Sears, R.C.; Sealy, L. The alternatively initiated c-Myc proteins differentially regulate transcription through a noncanonical DNA-binding site. Genes Dev. 1994, 8, 2441–2452. [Google Scholar] [CrossRef] [Green Version]
- Benassayag, C.; Montero, L.; Colombie, N.; Gallant, P.; Cribbs, D.; Morello, D. Human c-Myc isoforms differentially regulate cell growth and apoptosis in Drosophila melanogaster. Mol. Cell. Biol. 2005, 25, 9897–9909. [Google Scholar] [CrossRef] [Green Version]
- Escamilla-Powers, J.R.; Sears, R.C. A conserved pathway that controls c-Myc protein stability through opposing phosphorylation events occurs in yeast. J. Biol. Chem. 2007, 282, 5432–5442. [Google Scholar] [CrossRef] [Green Version]
- Elżbieta Kasprzyk, M.; Sura, W.; Dzikiewicz-Krawczyk, A. Enhancing B-Cell Malignancies-On Repurposing Enhancer Activity towards Cancer. Cancers 2021, 13, 3270. [Google Scholar] [CrossRef]
- Roix, J.J.; McQueen, P.G.; Munson, P.J.; Parada, L.A.; Misteli, T. Spatial proximity of translocation-prone gene loci in human lymphomas. Nat. Genet. 2003, 34, 287–291. [Google Scholar] [CrossRef]
- Gerbitz, A.; Mautner, J.; Geltinger, C.; Hörtnagel, K.; Christoph, B.; Asenbauer, H.; Klobeck, G.; Polack, A.; Bornkamm, G.W. Deregulation of the proto-oncogene c-myc through t(8; 22) translocation in Burkitt’s lymphoma. Oncogene 1999, 18, 1745–1753. [Google Scholar] [CrossRef] [Green Version]
- Dooley, S.; Wundrack, I.; Welter, C.; Blin, N. Constitutive C-myc overexpression and p1/p2 promoter shift in a small-cell lung-cancer cell-line. Int. J. Oncol. 1994, 5, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Ferrad, M.; Ghazzaui, N.; Issaoui, H.; Cook-Moreau, J.; Denizot, Y. Mouse models of c-myc deregulation driven by IgH locus enhancers as models of B-cell lymphomagenesis. Front. Immunol. 2020, 11, 1564. [Google Scholar] [CrossRef] [PubMed]
- Kovalchuk, A.L.; Qi, C.F.; Torrey, T.A.; Taddesse-Heath, L.; Feigenbaum, L.; Park, S.S.; Gerbitz, A.; Klobeck, G.; Hoertnagel, K.; Polack, A.; et al. Burkitt lymphoma in the mouse. J. Exp. Med. 2000, 192, 1183–1190. [Google Scholar] [CrossRef]
- Cesarman, E.; Dalla-Favera, R.; Bentley, D.; Groudine, M. Mutations in the first exon are associated with altered transcription of c-myc in Burkitt lymphoma. Science 1987, 238, 1272–1275. [Google Scholar] [CrossRef]
- Bayda, B.; Tilloy, V.; Chaunavel, A.; Bahri, R.; Halabi, M.A.; Feuillard, J.; Jaccard, A.; Ranger-Rogez, S. Comprehensive Epstein-Barr Virus Transcriptome by RNA-Sequencing in Angioimmunoblastic T Cell Lymphoma (AITL) and Other Lymphomas. Cancers 2021, 13, 610. [Google Scholar] [CrossRef]
- Wasylishen, A.R.; Chan-Seng-Yue, M.; Bros, C.; Dingar, D.; Tu, W.B.; Kalkat, M.; Chan, P.-K.; Mullen, P.J.; Huang, L.; Meyer, N.; et al. MYC phosphorylation at novel regulatory regions suppresses transforming activity. Cancer Res. 2013, 73, 6504–6515. [Google Scholar] [CrossRef] [Green Version]
- Laird-Offringa, I.A.; ElfTerich, P.; Knaken, H.J.; de Ruiter, J.; van der Eb, A.J. Analysis of polyadenylation site usage of the c-myc oncogene. Nucleic Acids Res. 1989, 17, 6499–6514. [Google Scholar] [CrossRef] [Green Version]
- Batsché, E.; Crémisi, C. Opposite transcriptional activity between the wild type c-myc gene coding for c-Myc1 and c-Myc2 proteins and c-Myc1 and c-Myc2 separately. Oncogene 1999, 18, 5662–5671. [Google Scholar] [CrossRef] [Green Version]
- de Arce, A.J.D.; Noderer, W.L.; Wang, C.L. Complete motif analysis of sequence requirements for translation initiation at non-AUG start codons. Nucleic Acids Res. 2018, 46, 985–994. [Google Scholar]
- Paronetto, M.P.; Passacantilli, I.; Sette, C. Alternative splicing and cell survival: From tissue homeostasis to disease. Cell Death Differ. 2016, 23, 1919–1929. [Google Scholar] [CrossRef] [PubMed]
- Zaika, A.I.; Slade, N.; Erster, S.H.; Sansome, C.; Joseph, T.W.; Pearl, M.; Chalas, E.; Moll, U.M. ΔNp73, a dominant-negative inhibitor of wild-type p53 and TAp73, is up- regulated in human tumors. J. Exp. Med. 2002, 196, 765–780. [Google Scholar] [CrossRef] [PubMed]
- Gill, T.; Wang, H.; Bandaru, R.; Lawlor, M.; Lu, C.; Nieman, L.T.; Tao, J.; Zhang, Y.; Anderson, D.G.; Ting, D.T.; et al. Selective targeting of MYC mRNA by stabilized antisense oligonucleotides. Oncogene 2021, 40, 6527–6539. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer (P) | 5′ to 3′ Sequence | Localisation | Use For |
|---|---|---|---|
| P5′UTR1 | CTGGTTTTCCACTACCCGAAA | Exon 1 | 5′UTR Amplification |
| P5′UTR2 | CGTTGAGGGGCATCGTCGCGGG | Exon 2 | 5′UTR Amplification |
| P3′UTR1 | CTACGGAACTCTTGTGCGTAA | Start at the End of the Last Exon | 3′UTR Amplification |
| PFull1 | GACCCCCGAGCTGTGCTGCTC | Start at P1:+1 | Full-Length cDNA |
| PFull2 | GGGAACAGCCGCAGCGGAGGG | Start at P3:+1 | Full-Length cDNA |
| PFull3 | TTACGCACAAGAGTTCCGTAG | Start at the End of the Last Exon | Full-Length cDNA |
| PFull4 | GGAGATAGTGGGGCTCAGAGC | Start at P?:+1 | Full-Length cDNA |
| Orf-c-Myc1 | ATCATATGCTGGATTTTTTTCGGG | CTG Initiation Codon | Full-Length Orf-c-Myc1 |
| Orf-c-Myc2 | ATCATATGCCCCTCAACGTTAGCT | ATG Initiation Codon | Full-Length Orf-c-Myc2 |
| Orf-c-MycS | ATCATATGGTGACCGAGCTGCTG | ATG Initiation Codon | Full-Length Orf-c-MycS |
| Orfs | ATCTCGAGTTACGCACAAGAGTTC | TTA Stop Codon | Reverse for all Orfs |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, D.; Prévaud, L.; Faumont, N.; Troutaud, D.; Feuillard, J.; Diab-Assaf, M.; Oulmouden, A. Alternative c-MYC mRNA Transcripts as an Additional Tool for c-Myc2 and c-MycS Production in BL60 Tumors. Biomolecules 2022, 12, 836. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12060836
Ibrahim D, Prévaud L, Faumont N, Troutaud D, Feuillard J, Diab-Assaf M, Oulmouden A. Alternative c-MYC mRNA Transcripts as an Additional Tool for c-Myc2 and c-MycS Production in BL60 Tumors. Biomolecules. 2022; 12(6):836. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12060836
Chicago/Turabian StyleIbrahim, Dina, Léa Prévaud, Nathalie Faumont, Danielle Troutaud, Jean Feuillard, Mona Diab-Assaf, and Ahmad Oulmouden. 2022. "Alternative c-MYC mRNA Transcripts as an Additional Tool for c-Myc2 and c-MycS Production in BL60 Tumors" Biomolecules 12, no. 6: 836. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12060836