
Genomic Variation-Mediating Fluconazole Resistance in Yeast
by
Wen-Yao Wang
, 
Hong-Qing Cai
,
Si-Yuan Qu
,
Wei-Hao Lin
,
Cheng-Cheng Liang
,
Hao Liu
,
Ze-Xiong Xie
* and
Ying-Jin Yuan
Frontiers Science Center for Synthetic Biology, Key Laboratory of Systems Bioengineering (Ministry of Education), School of Chemical Engineering and Technology, Tianjin University, Tianjin 300072, China
*
Author to whom correspondence should be addressed.
Biomolecules 2022, 12(6), 845; https://0-doi-org.brum.beds.ac.uk/10.3390/biom12060845
Submission received: 28 April 2022
/
Revised: 7 June 2022
/
Accepted: 9 June 2022
/
Published: 17 June 2022
(This article belongs to the Special Issue Theme Issue Honoring Scientist Louis Pasteur on His 200th Birthday)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Fungal infections pose a serious and growing threat to public health. These infections can be treated with antifungal drugs by killing hazardous fungi in the body. However, the resistance can develop over time when fungi are exposed to antifungal drugs by generating genomic variations, including mutation, aneuploidy, and loss of heterozygosity. The variations could reduce the binding affinity of a drug to its target or block the pathway through which drugs exert their activity. Here, we review genomic variation-mediating fluconazole resistance in the yeast Candida, with the hope of highlighting the functional consequences of genomic variations for the antifungal resistance.
1. Introduction
Nearly 1 billion people suffer from fungal infections, and fungal diseases caused over 1.6 million deaths annually, and more than 10 million people have mucosal candidiasis [1,2]. The yeast Candida is one of the most frequent fungal pathogens, including Candida albicans, Candida tropicalis, Candida parapsilosis, Candida auris, and Candida glabrata. The yeast Candida is an opportunistic pathogen present in the normal microbiome, which can overgrow in immune-compromised individuals and cause disseminated infections [1,3]. Available antifungal drugs are limited, and fluconazole is one of the most widely used antifungal agents [4]. Fluconazole is an efficient inhibitor of lanosterol 14-α-demethylase, which functions to convert lanosterol to ergosterol. Subsequently, the synthesis of normal sterols is blocked, and toxic 14-α-methyl sterols are accumulated in fungi, which is responsible for the fungistatic activity [3,5].
However, with the long-term use of low-concentration antifungal drugs, some types of fungi have become resistant to undergoing abundant genomic variations, including point mutations, aneuploidy, and loss of heterozygosity (LOH). Approximately 0.5–2% of C. albicans isolates are resistant to fluconazole, while the proportion of resistant C. tropicalis, C. parapsilosis, and C. glabrata amounts to 4–9%, 2–6%, and 11–13%, respectively [6]. Genomic variations usually lead to increased drug efflux and decreased affinity between the drug and the target. A better understanding of the antifungal resistance mechanism is critical to developing better therapeutics and to improving strategies that may overcome resistance [7]. Here, we focus on fluconazole resistance in yeast pathogens, highlighting the resistance mediated by genomic variations.
2. Antifungal Resistances Mediated by Genetic Mutations
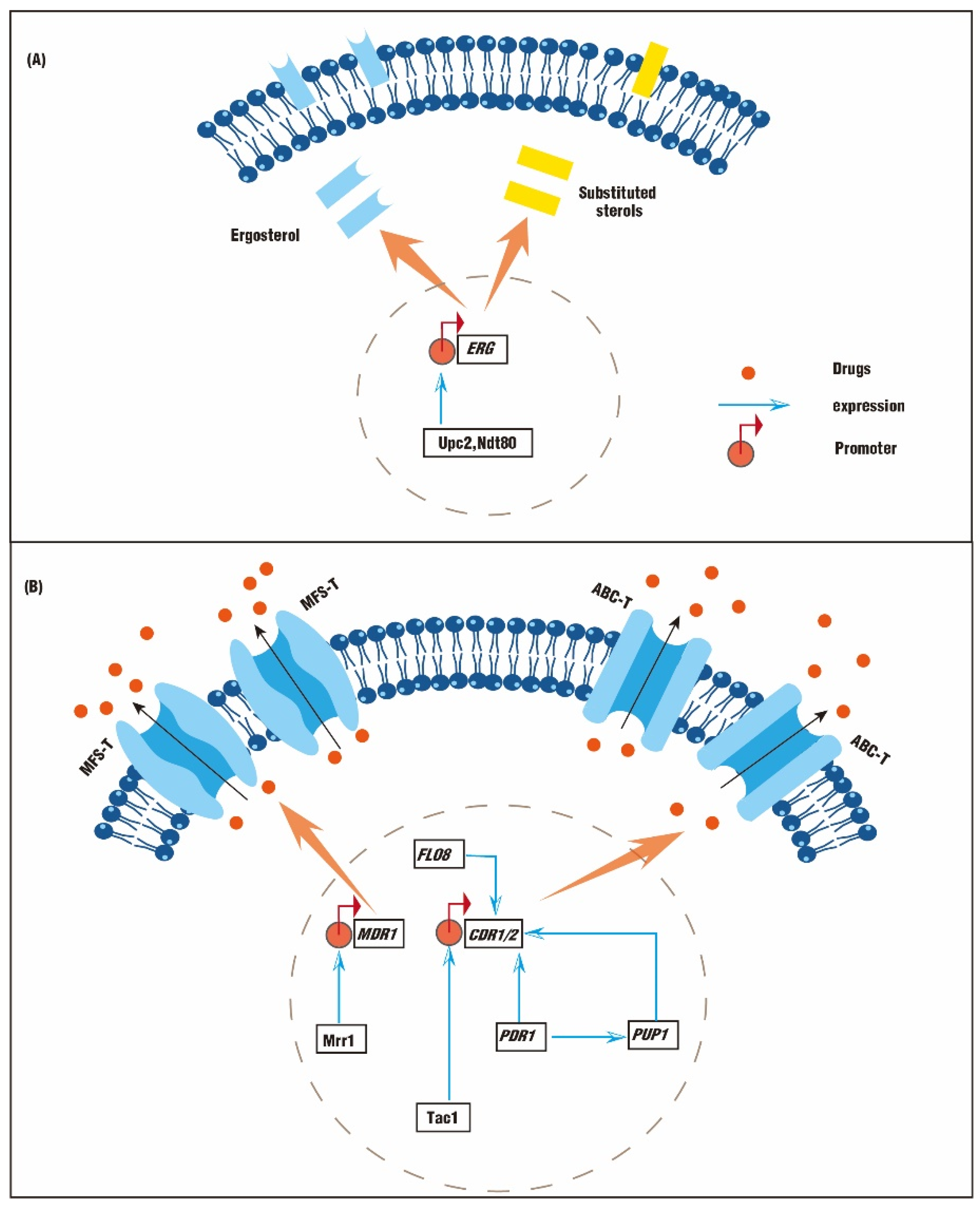
Slight alterations in DNA nucleotides in necessary genes can enable notable antifungal resistance. The ergosterol biosynthesis pathway is a complex route involving about 20 enzymes, and most of these enzymes are encoded by a series of ERG genes. Mutations in ERG genes could affect ergosterol synthesis and lead to fluconazole resistance. Interestingly, functional mutations are mainly represented in ERG3 and ERG11, encoding C-5 sterol desaturase and sterol 14-demethylase, respectively. Additionally, the facilitator superfamily transporter (MFS-T) and the ATP-binding cassette transporter (ABC-T) are also responsible for the fungistatic activity of fluconazole. The MFS-T is encoded by MDR1, and the ABC-T is encoded by CDR1/CDR2. While functional mutations are represented in MDR1 or CDR1/CDR2, the overexpression of efflux pumps can also lead to fluconazole resistance (Figure 1A,B) [3].
2.1. Genetic Mutations Lead to Abnormal Synthesis of Sterols
The synthesis of ergosterol can be affected by ERG mutations. Two mutations, Y132F and K143R, in ERG11 affect the catalytic efficiency of Erg11. Therefore, the binding affinity of fluconazole to its target is reduced. The isolate with these mutations exhibited a 16-fold increase in fluconazole minimum inhibitory concentration (MIC) [8]. Missense mutations and silent point mutations in ERG3 are thought to be responsible for the mis-synthesized sterols and high levels of drug resistance [9]. A C. albicans isolate containing T51C, T434C, and C1052T substitutions in ERG3 exhibits an increased fluconazole resistance, with ergosterol substituted by ergosta-7,22-dienol (Figure 1A) [9]. In addition, clinical Candida isolates with ERG5 and ERG11 mutations exhibit a complete reduction in ergosterol and a cross-resistance to fluconazole and amphotericin B (AmB) (Figure 1A) [10]. UPC2 encodes a zinc cluster transcription factor, Upc2, which regulates the uptake and metabolism of sterol. Upc2 and Ndt80 (also a zinc cluster transcription factor) can bind promoters of ERG genes [11]. Therefore, overexpressed UPC2 and NDT80 will upregulate expressions of ERG2, ERG3, ERG4, ERG6, ERG11, and ERG25, generating fluconazole-resistant Candida isolates. On the contrary, the strain susceptibility to fluconazole can be increased by 500 times when UPC2 is deleted alone or in combination with NDT80 (Figure 1A) [11]. Genetic mutations are also associated with cross-resistance. For example, C. albicans isolates containing the A516P mutation in ERG11 exhibit an upregulated ERG11 expression and a notable cross-resistance to fluconazole, itraconazole, and voriconazole [12].
2.2. Genetic Mutations Lead to An Abnormal Efflux Pump
The overexpression of cell membrane transporters can also be responsible for the fungistatic activity of fluconazole. The efficiency of efflux pumps can be increased by the overexpression of CDR1/CDR2 or MDR. Therefore, drug molecules are pumped out of the cell, exhibiting enhanced antifungal resistance (Figure 1B). The binding and terminal modification of cis-regulatory elements also affect antifungal resistance. PDR genes function during the synthesis of ABC-Ts. The PDR12 promoter contains a cis-regulatory element for Upc2 binding. Isolates without UPC2 exhibit abnormal sterols accumulation and membrane permeability, as well as increased susceptibility to fluconazole [13]. While the C-terminal modification of PDR1 with FLAG tag enables a decreased mRNA concentration by about 50%, the N-terminal modification leads to a 2-fold increase in mRNA concentration. The reduction in PDR1 leads to a significant reduction in fluconazole resistance. The C-terminal modifications also interfere with the action of Gal11A, which subsequently decrease the expression of PDR1, CDR1, and PUP1 [14]. Moreover, the deletion of FLO8 upregulates the expression of CDR1/CDR2 and is responsible for increased resistance to various drugs (Figure 1B) [15].
Gain-of-function (GOF) mutations can enable resistance to fluconazole in some species of Candida. In some fluconazole-resistant C. glabrata isolates, a total of ten synonymous mutations and two missense mutations were identified in PDR1, four of which are considered GOF mutations. The corresponding genes are over-transcribed 20 to 40 times and further upregulate the expression of CDR1 and PUP1 (CDR1 upregulated genes), which are responsible for increased fluconazole resistance (Figure 1B) [14,16]. Tac1 and Mrr1 function as transcriptional activators of CDR1 and MDR1, respectively (Figure 1B). GOF mutations of MRR1 and TAC1 can upregulate the expression of CDR1 and MDR1, and increase the MIC of C. albicans to fluconazole by 60 to 120 times [17,18].
3. Antifungal Resistances Mediated by Aneuploidy
Fungal pathogens have significant genomic plasticity for genomic evolutions and adaptions to environmental perturbations. Aneuploidy is often observed across the Candida species and leads to fluconazole resistance. For example, among forty-two detected fluconazole-resistant isolates, twenty-one isolates are aneuploids [19]. While genetic mutations affect protein function, aneuploidies affect gene expression through gene dosage effects [20]. These alterations make strains less susceptible to antimicrobials by affecting drug targets, efflux pumps, and expressions of other genes that are associated with drug resistance [21]. A reported clinical C. glabrata aneuploidy isolate, which exhibits a two-fold ergosterol concentration, is resistant to fluconazole. The duplicated chromosome contains the ERG11 gene, which encodes the target of fluconazole, suggesting that chromosome replication is associated with fluconazole resistance [22,23].
Both whole chromosomes and chromosome segments can be duplicated or deleted, generating aneuploidy. Seventy fluconazole-resistant and fluconazole-susceptible C. albicans clinical and laboratory isolates were analyzed by using comparative genomic hybridization (CGH) arrays. The result reveals that the chr5L isochromosome [i(5L)] is the aneuploidy most frequently associated with fluconazole resistance (Figure 2A) [19]. Analyses of expression profiles confirm that the expression of ERG11 and TAC1 on chr5L is increased, which contributes to drug resistance and tolerance [19].
Trisomy is also prevalent in fluconazole-resistant Candida species. In fluconazole-resistant C. albicans isolates, chromosome 4 (chr4) trisomy is identified, on which there are no mutations in resistance-associated genes [24]. Trisomy of chromosome 3 (chr3), chromosome 6 (chr6), and chromosome 7 (chr7) is also detected in fluconazole-resistant isolates [25]. Not surprisingly, most of these chromosomes contain genes that affect azole resistance. For example, chr3 contains CDR1, CDR2, and MRR1. MRR1 encodes a transcription factor promoting MDR1 expression (Figure 2B) [26,27,28]. MDR1 is present on chr6, and NCP1, a gene encoding the Erg11 cofactor, is present on chr4 [25].
Aneuploidy can also enable cross-adaptations to unrelated drugs. An isolate of C. albicans with chromosome 2 (chr2) trisomy is adapted to both the anticancer drug hydroxyurea and the antifungal drug caspofungin. The cross-adaptation may occur because alleles of two adaptor genes are located on the same chromosome [20]. In fluconazole-resistant Cryptococcus neoformans isolates, those with disomy of chromosome 1 (chr1) are adapted to flucytosine (5FC), and those with disomy of both chr1 and chr4 exhibit cross-tolerance to AMB and 5FC [29].
Aneuploidy is not only a readily available variation but also reversible. When exposed to conditions without drugs, resistant aneuploidies could gradually regain their susceptibility to drugs by losing aneuploidy chromosomes. Itraconazole- and fluconazole-resistant C. albicans isolates were incubated for more than 150 generations in conditions without drugs. The resistance to itraconazole disappeared, and the resistance to fluconazole decreased to 43% of that of the initial strains [22]. Studies on C. neoformans reveal that the loss of aneuploidy is associated with the apoptosis-inducing factor Aif1. The inactivation of the AIF1 gene enables stable chr1 dimers in the absence of drugs [23].
Polyploids occur during the growth and reproduction of fungi, and different ploidies have different potential to produce genomic changes. Upon exposure to fluconazole, diploid C. albicans forms unstable tetraploid intermediates by altering cell cycle progression and undergoing abnormal mitosis [30]. The genome of tetraploid C. albicans is more unstable than diploids [31,32]. Tetraploids undergo rapid and dramatic genome reductions, abnormal chromosome segregations, and gene rearrangements, leading to aneuploidy while shrinking to diploids [31].
Indeed, the accuracy of chromosome segregation during mitosis is altered under stressful duress, resulting in karyotypic diversity and promoting adaptive cellular evolution. Fluconazole affects ergosterol biosynthesis and nuclear membrane fluidity, which can further affect the proper nucleus separation. Therefore, normal chromosome segregations are disrupted, leading to the frequent occurrence of fluconazole-resistant polyploids or aneuploids [33]. When exposed to fluconazole and AmB, haploid yeast cells exhibit a high probability of chromosome loss. Similarly, in strains lacking the checkpoint gene MAD2, aneuploid cells are frequently detected, and these isolates exhibit fluconazole resistance [34].
4. Antifungal Resistances Mediated by LOH
LOH refers to a cross-chromosomal event that results in the loss of entire alleles and the surrounding chromosomal region in a heterozygous cell. LOH is an efficient strategy that allows species to adapt to new environments. In particular, LOH is frequently detected in fluconazole-resistant C. albicans isolates [35].
4.1. Chromosomal Regions of LOH Associated with Drug Resistance
The diploid genome of C. albicans exhibits a high degree of heterozygosity. However, LOH could be identified at specific sites in fluconazole-resistant C. albicans isolates [36]. Genomic analysis revealed that these LOHs are the results of recombination events or the loss and duplication of a whole chromosome [26]. Therefore, it can be assumed that the presence of fluconazole may induce genomic rearrangements, which further enhance drug resistance. Unlike aneuploidy, an LOH-associated drug resistance increase is persistent and recurrent [37]. The LOH of chr1, the right arm of chromosome 3 (chr3R), and the left arm of chromosome 5 (chr5L) are identified in clinical isolates, and chr3R and chr5L are statistically recurrent [37].
The chr5 of C. albicans is representative of LOH events, on which the genes of ERG11, TAC1, and the mating-type locus (MTL locus) exist [35]. C. albicans usually exhibits as a diploid with three different mating types (MTLa/α, MTLa/a, or MTLα/α). MTLa/α diploid cells exhibit an MTL-heterozygous biofilm; otherwise, an MTL-homozygous biofilm appears. The mating is prevented by MTL-heterozygous biofilms, and multiple drug-resistant C. albicans strains are homozygous [38]. TAC1 is located 14 kb from the MTL locus, and the LOH of TAC1 tends to extend to the MTL locus. Therefore, strains with homozygous TAC1 alleles are usually accompanied by homozygous MTL loci [39]. The poly(A) polymerase responsible for mRNA adenylation, encoded by PAP1, PAP1-a/PAP1-α, is also adjacent to the MTL locus. Homozygous PAP1-α alleles enable hyperadenylation and increase the stability of CDR1 transcripts [40]. Strains with homozygous PAP1 alleles are also usually accompanied by homozygous MTL loci. Further distal to chr5L, LOH of the EEG11 allele is also often accompanied by homozygous MTL loci due to recombination events [38]. Surprisingly, a homozygous mutation in MRR1 on chr3 results in an increased LOH rate of chr5 [35]. It is assumed that the LOH of MRR1 can promote the homozygous MTL through the loss of chr5 [38].
4.2. Different Types of LOH Associated with Drug Resistance
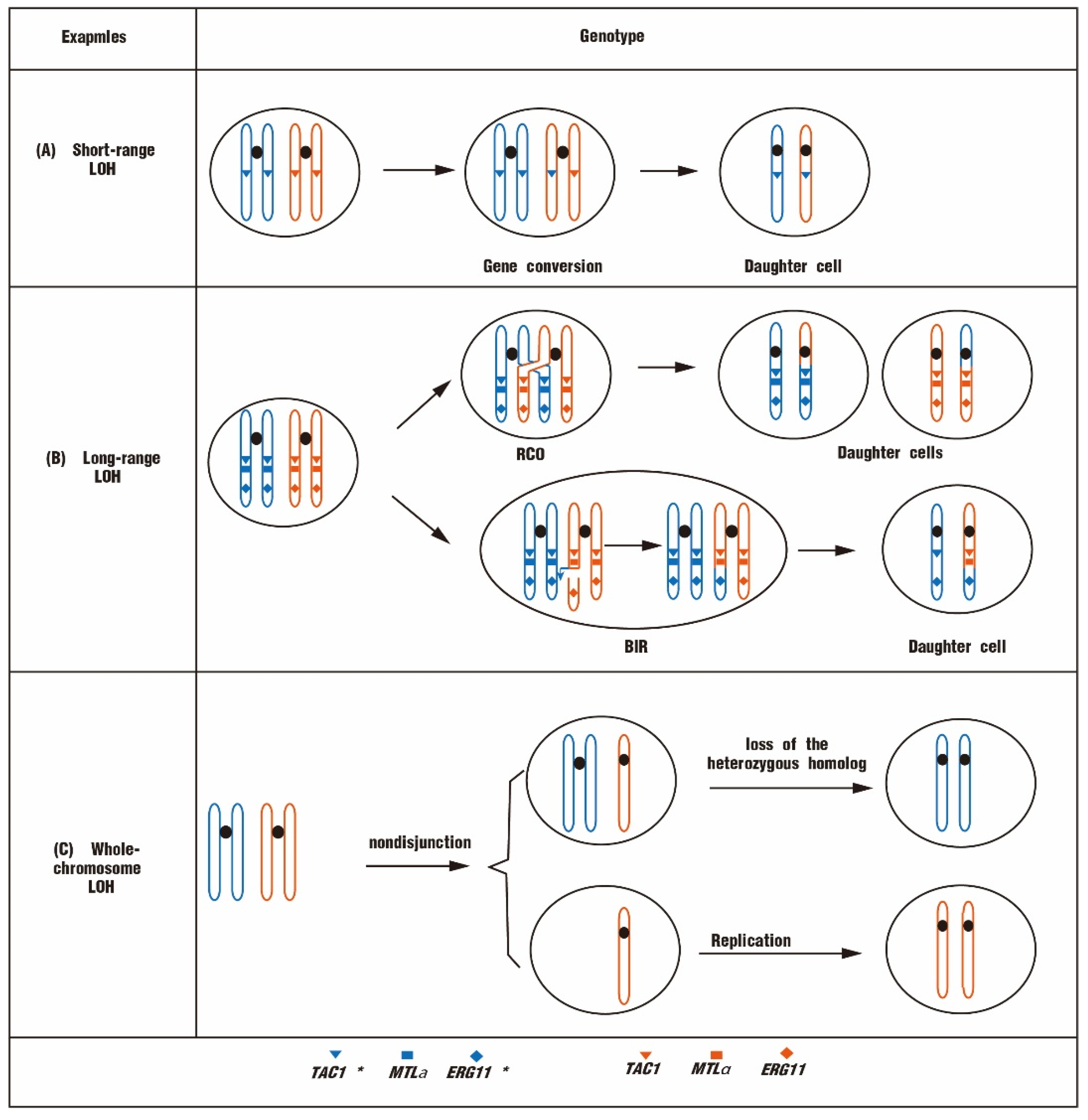
According to the length of DNA fragments, LOHs are classified into short-range LOH, long-range LOH, or whole-chromosome LOH [33,41]. All three types of LOH are associated with antifungal resistance. Short-range LOHs are also called interstitial LOHs (I-LOHs), which undergo small-scale DNA fragment transfers (<50 kb) (Figure 3A). I-LOH events are the most frequent LOHs and are usually formed by gene conversion [42]. For example, of 96 clinical C. albicans isolates analyzed, the majority of drug-resistant strains exhibited homozygous MTL loci [43]. The homozygous MTL could facilitate the mating and chromosome reassemblies to generate resistant progeny. In a fluconazole-resistant C. albicans isolate, a GOF mutation in the TAC1 allele, N977D, is accompanied by an LOH on chr5 [39]. This LOH event may be due to the gene conversion of TAC1 after the GOF mutation, leading to an overall enhancement of Tac1 activity [39].
Long-range LOHs are also called terminal LOHs (T-LOHs) with long chromosomal fragments in homozygous LOHs (>50 kb) (Figure 3B). T-LOHs are usually formed by reciprocal cross-over (RCO) or break-induced replication (BIR) events. LOHs in the yeast genome usually correspond to large-scale chromosomal regions containing multiple adjacent genes [44]. Fluconazole-resistant strains usually contain extremely active TAC1 alleles and homozygous ERG11 alleles. TAC1 hyperactivity is generally due to GOF mutations, whereas the homozygosity of ERG11 is acquired through mitotic recombination events. ERG11 recombination events occur with the LOH of TAC1 and MTL when the chromosomal breakage is adjacent to the centromere, generating a T-LOH [39]. The MDR1 gene is located on chr3, and its overexpression is one of the main causes of fluconazole resistance for C. albicans [45]. Surprisingly, most fluconazole-resistant isolates exhibit a constitutive upregulation of MDR1 and homozygous MRR1 mutations [28]. It reveals that the LOH of MRR1 can be caused by a mitotic recombination event involving a cross-over between the centromere and the MRR1 locus. The recombination event results in a T-LOH from the cross-over locus to the telomere [26].
Whole-chromosome LOH (whole-chr LOH) is a phenomenon of chromosome non-disjunction, causing chromosome imbalance in daughter cells. The zygote with one chromatid undergoes a duplication of the remaining chromosome, generating a homozygous diploid [46,47]. The zygote with three chromatids undergoes a loss of the heterozygous homolog to become homozygous (Figure 3C) [47]. Whole-chr LOHs of chr1, chr3, and chr5 are detected in fluconazole-resistant C. albicans isolates, which contain numerous fluconazole resistance genes [26,37].
4.3. Factors That Could Affect LOHs
When exposed to antifungal drugs, C. albicans undergo more genomic recombination than mutations. For instance, when exposed to fluconazole, rates of total LOHs and whole-chr LOHs increase by approximately 285 and 5 times, respectively [33]. The clinical isolates exhibit a higher rate of LOH than laboratory strains. The host-associated strains suffer approximately 10-fold more LOHs than strains that grow in nature [31]. It is assumed that innate immune components, including antimicrobial peptides (AMPs) and reactive oxygen species (ROS), make host-carried C. albicans generate more large-scale genomic variations. Meanwhile, chromosomal or segmental aneuploidy is frequently detected in LOH strains [48]. Furthermore, tetraploids endure a more severe drug-induced genomic instability than diploids. When exposed to fluconazole, generated LOHs in tetraploids are 60 times greater than those in diploids [32]. Adaptive mistranslations at CUG sites also affect the rate of LOH events and accelerate the generation of fluconazole-resistant C. albicans isolates [49]. In addition, the rate of LOH in strains can be increased by UV and γ-radiation, CRISPR/Cas9, and SCRaMbLE [50,51,52].
5. Conclusions
Antifungal resistance is a severe threat to human health. The yeast Candida is the most common pathogen, and fluconazole is an efficient therapeutic drug for fungal disease. Unfortunately, fungi can become resistant over time by generating genomic variations when exposed to fluconazole. The resistant fungi are capable of being isolated from the clinical environment and analyzed by genomic analysis. Therefore, we know that fluconazole resistance is related to genomic variations at different levels, including mutations, aneuploidies, and LOHs.
Genetic mutations generate amino acid substitutions, which are closely related to drug resistance. In particular, missense mutations and GOF mutations can alter protein structures or promote the expression of resistance genes. Aneuploidies and LOHs are capable of affecting the copy number of genes and their expression, which further affects drug targets, efflux pumps, and the expression of other drug-resistant genes. Aneuploidy and LOHs are unstable, which can be rapidly acquired under drug stimulation and recovered under drug-free conditions.
In general, the generation of fungal resistance is facilitated by chromosome breakages and recombination. Thus, new therapeutic strategies are being developed to reduce the frequency of antifungal resistance. Agents capable of reducing chromosome breakage and recombination can be used as adjuvants to fluconazole [19]. However, the mechanism of drug resistance is complex and is also related to biofilms, the host environment, and the pharmacokinetics of the drug. To fully elucidate the mechanism of drug resistance, unremitting efforts and powerful research technologies are necessary.
The better we understand these variations, the better we can treat the fungi disease precisely. The development of synthetic biology provides new opportunities for fungal resistant analysis from screening to creation. Through genome synthesis and inducible rearrangement, we can create drug-resistant fungi. The SCRaMbLE technology can accelerate the evolution of synthetic genomes and generate large strain libraries with numerous genomic variants, including aneuploidies, LOHs, and structural variations. It provides an efficient platform for the detection of drug-resistant strains, the analysis of drug resistance mechanisms, and the identification of new drug targets. Furthermore, automated biofoundries and machine learning can be employed to facilitate strain construction and analysis, promoting the understanding of drug-resistant impacts of genomic variations and rational clinical drug use.
Author Contributions
Conceptualization, Z.-X.X. and Y.-J.Y.; investigation, W.-Y.W., H.-Q.C., S.-Y.Q., W.-H.L., C.-C.L. and H.L.; writing—original draft preparation, Z.-X.X., W.-Y.W., H.-Q.C., S.-Y.Q., W.-H.L. and C.-C.L.; writing—review and editing, Z.-X.X. and W.-Y.W.; supervision, Z.-X.X.; funding acquisition, Z.-X.X. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Key Research and Development Program of China (2019YFA0906600), the National Natural Science Foundation of China (21907074), and the Natural Science Foundation of Tianjin (20JCQNJC02090).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bongomin, F.; Gago, S.; Oladele, R.O.; Denning, D.W. Global and multi-national prevalence of fungal diseases-estimate precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef]
- Almeida, F.; Rodrigues, M.L.; Coelho, C. The still underestimated problem of fungal diseases worldwide. Front. Microbiol. 2019, 10, 214. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, S.; Sae-Tia, S.; Fries, B.C. Candidiasis and mechanisms of antifungal resistance. Antibiotics 2020, 9, 312. [Google Scholar] [CrossRef]
- Costa-de-Oliveira, S.; Rodrigues, A.G. Candida albicans antifungal resistance and tolerance in bloodstream infections: The triad yeast-host-antifungal. Microorganisms 2020, 8, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, R.S.; Robbins, N.; Cowen, L.E. Regulatory circuitry governing fungal development, drug resistance, and disease. Microbiol. Mol. Biol. Rev. 2011, 75, 213–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berkow, E.L.; Lockhart, S.R. Fluconazole resistance in Candida species: A current perspective. Infect. Drug Resist. 2017, 10, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Cowen, L.E.; Sanglard, D.; Howard, S.J.; Rogers, P.D.; Perlin, D.S. Mechanisms of antifungal drug resistance. Cold Spring Harb. Perspect. Med. 2015, 5, a019752. [Google Scholar] [CrossRef]
- Rybak, J.M.; Sharma, C.; Doorley, L.A.; Barker, K.S.; Palmer, G.E.; Rogers, P.D. Delineation of the direct contribution of Candida auris ERG11 mutations to clinical triazole resistance. Microbiol. Spectr. 2021, 9, e01585-21. [Google Scholar] [CrossRef]
- Vale-Silva, L.A.; Coste, A.T.; Ischer, F.; Parker, J.E.; Kelly, S.L.; Pinto, E.; Sanglard, D. Azole resistance by loss of function of the sterol delta(5,6)-desaturase gene (ERG3) in Candida albicans does not necessarily decrease virulence. Antimicrob. Agents Chemother. 2012, 56, 1960–1968. [Google Scholar] [CrossRef] [Green Version]
- Martel, C.M.; Parker, J.E.; Bader, O.; Weig, M.; Gross, U.; Warrilow, A.G.S.; Kelly, D.E.; Kelly, S.L. A clinical isolate of Candida albicans with mutations in ERG11 (encoding sterol 14 alpha-demethylase) and ERG5 (encoding c22 desaturase) is cross resistant to azoles and amphotericin B. Antimicrob. Agents Chemother. 2010, 54, 3578–3583. [Google Scholar] [CrossRef] [Green Version]
- Branco, J.; Ola, M.; Silva, R.M.; Fonseca, E.; Gomes, N.C.; Martins-Cruz, C.; Silva, A.P.; Silva-Dias, A.; Pina-Vaz, C.; Erraught, C.; et al. Impact of ERG3 mutations and expression of ergosterol genes controlled by UPC2 and NDT80 in Candida parapsilosis azole resistance. Clin. Microbiol. Infect. 2017, 23, 575.e1–575.e8. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.L.; Yang, J.; Ma, Y.; Xi, Z.Q.; Zhao, X.Q.; Zhao, X.X.; Zhao, M. The effects of secreted aspartyl proteinase inhibitor ritonavir on azoles-resistant strains of Candida albicans as well as regulatory role of SAP2 and ERG11. Immun. Inflamm. Dis. 2021, 9, 667–680. [Google Scholar] [CrossRef]
- Hervay, N.T.; Bencova, A.; Valachovic, M.; Morvova, M.; Gbelska, Y. UPC2 gene deletion modifies sterol homeostasis and susceptibility to metabolic inhibitors in Kluyveromyces lactis. Yeast 2020, 37, 647–657. [Google Scholar] [CrossRef]
- Tian, Y.; Gao, N.; Ni, Q.; Mao, Y.; Dong, D.; Huang, X.; Jiang, C.; Li, Z.; Zhang, L.; Wang, X.; et al. Sequence modification of the master regulator Pdr1 interferes with its transcriptional autoregulation and confers altered azole resistance in Candida glabrata. FEMS Yeast Res. 2018, 18, foy038. [Google Scholar] [CrossRef]
- Li, W.J.; Liu, J.Y.; Shi, C.; Zhao, Y.; Meng, L.N.; Wu, F.; Xiang, M.J. FLO8 deletion leads to azole resistance by upregulating CDR1 and CDR2 in Candida albicans. Res. Microbiol. 2019, 170, 272–279. [Google Scholar] [CrossRef]
- Ni, Q.; Wang, C.; Tian, Y.; Dong, D.F.; Jiang, C.; Mao, E.Q.; Peng, Y.B. CgPDR1 gain-of-function mutations lead to azole-resistance and increased adhesion in clinical Candida glabrata strains. Mycoses 2018, 61, 430–440. [Google Scholar] [CrossRef]
- Lohberger, A.; Coste, A.T.; Sanglard, D. Distinct roles of Candida albicans drug resistance transcription factors TAC1, MRR1, and UPC2 in virulence. Eukaryot. Cell 2014, 13, 127–142. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.L.; Rossi, J.M.; Myers, L.C. Candida albicans zn cluster transcription factors Tac1 and Znc1 are activated by farnesol to upregulate a transcriptional program including the multidrug efflux pump CDR1. Antimicrob. Agents Chemother. 2018, 62, e00968-18. [Google Scholar] [CrossRef] [Green Version]
- Selmecki, A.; Forche, A.; Berman, J. Aneuploidy and isochromosome formation in drug-resistant Candida albicans. Science 2006, 313, 367–370. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Teoh, F.; Tan, A.S.M.; Cao, Y.B.; Pavelka, N.; Berman, J. Aneuploidy enables cross-adaptation to unrelated drugs. Mol. Biol. Evol. 2019, 36, 1768–1782. [Google Scholar] [CrossRef]
- Lee, Y.; Puumala, E.; Robbins, N.; Cowen, L.E. Antifungal drug resistance: Molecular mechanisms in Candida albicans and beyond. Chem. Rev. 2021, 121, 3390–3411. [Google Scholar] [CrossRef] [PubMed]
- vanden Bossche, H.; Marichal, P.; Odds, F.C.; Le Jeune, L.; Coene, M.C. Characterization of an azole-resistant Candida glabrata isolate. Antimicrob. Agents Chemother. 1992, 36, 2602–2610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon-Chung, K.J.; Chang, Y.C. Aneuploidy and drug resistance in pathogenic fungi. PLoS Pathog. 2012, 8, e1003022. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.Z.; Saha, A.; Haseeb, A.; Bennett, R.J. A chromosome 4 trisomy contributes to increased fluconazole resistance in a clinical isolate of Candida albicans. Microbiology 2017, 163, 856–865. [Google Scholar] [CrossRef] [PubMed]
- Selmecki, A.M.; Dulmage, K.; Cowen, L.E.; Anderson, J.B.; Berman, J. Acquisition of aneuploidy provides increased fitness during the evolution of antifungal drug resistance. PLoS Genet. 2009, 5, e1000705. [Google Scholar] [CrossRef]
- Dunkel, N.; Blass, J.; Rogers, P.D.; Morschhauser, J. Mutations in the multi-drug resistance regulator MRR1, followed by loss of heterozygosity, are the main cause of MDR1 overexpression in fluconazole-resistant Candida albicans strains. Mol. Microbiol. 2008, 69, 827–840. [Google Scholar] [CrossRef] [Green Version]
- Akins, R.A. An update on antifungal targets and mechanisms of resistance in Candida albicans. Med. Mycol. 2005, 43, 285–318. [Google Scholar] [CrossRef] [Green Version]
- Morschhauser, J.; Barker, K.S.; Liu, T.T.; Blass-Warmuth, J.; Homayouni, R.; Rogers, P.D. The transcription factor Mrr1p controls expression of the MDR1 efflux pump and mediates multidrug resistance in Candida albicans. PLoS Pathog. 2007, 3, 1603–1616. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Gritsenko, V.; Lu, H.; Zhen, C.; Gao, L.; Berman, J.; Jiang, Y.Y. Adaptation to fluconazole via aneuploidy enables cross-adaptation to amphotericin B and flucytosine in Cryptococcus neoformans. Microbiol. Spectr. 2021, 9, e00723-21. [Google Scholar] [CrossRef]
- Harrison, B.D.; Hashemi, J.; Bibi, M.; Pulver, R.; Bavli, D.; Nahmias, Y.; Wellington, M.; Sapiro, G.; Berman, J. A tetraploid intermediate precedes aneuploid formation in yeasts exposed to fluconazole. PLoS Biol. 2014, 12, e1001815. [Google Scholar] [CrossRef]
- Smith, A.C.; Hickman, M.A. Host-Induced genome instability rapidly generates phenotypic variation across Candida albicans strains and ploidy states. mSphere 2020, 5, e00433-20. [Google Scholar] [CrossRef] [PubMed]
- Avramovska, O.; Hickman, M.A. The magnitude of Candida albicans stress-induced genome instability results from an interaction between ploidy and antifungal drugs. Genes Genomes Genet. 2019, 9, 4019–4027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forche, A.; Abbey, D.; Pisithkul, T.; Weinzierl, M.A.; Ringstrom, T.; Bruck, D.; Petersen, K.; Berman, J. Stress alters rates and types of loss of heterozygosity in Candida albicans. mBio 2011, 2, e00129-11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.B.; Bradford, W.D.; Seidel, C.W.; Li, R. Hsp90 stress potentiates rapid cellular adaptation through induction of aneuploidy. Nature 2012, 482, 246–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morschhauser, J. The development of fluconazole resistance in Candida albicans—An example of microevolution of a fungal pathogen. J. Microbiol. 2016, 54, 192–201. [Google Scholar] [CrossRef]
- Sasse, C.; Dunkel, N.; Schafer, T.; Schneider, S.; Dierolf, F.; Ohlsen, K.; Morschhauser, J. The stepwise acquisition of fluconazole resistance mutations causes a gradual loss of fitness in Candida albicans. Mol. Microbiol. 2012, 86, 539–556. [Google Scholar] [CrossRef]
- Ford, C.B.; Funt, J.M.; Abbey, D.; Issi, L.; Guiducci, C.; Martinez, D.A.; Delorey, T.; Li, B.Y.; White, T.C.; Cuomo, C.; et al. The evolution of drug resistance in clinical isolates of Candida albicans. eLife 2015, 4, e00662. [Google Scholar] [CrossRef] [Green Version]
- Popp, C.; Ramirez-Zavala, B.; Schwanfelder, S.; Kruger, I.; Morschhauser, J. Evolution of fluconazole-resistant Candida albicans strains by drug-induced mating competence and parasexual recombination. mBio 2019, 10, e02740-18. [Google Scholar] [CrossRef] [Green Version]
- Coste, A.; Selmecki, A.; Forche, A.; Diogo, D.; Bougnoux, M.E.; d’Enfert, C.; Berman, J.; Sanglard, D. Genotypic evolution of azole resistance mechanisms in sequential Candida albicans isolates. Eukaryot. Cell 2007, 6, 1889–1904. [Google Scholar] [CrossRef] [Green Version]
- Manoharlal, R.; Gorantala, J.; Sharma, M.; Sanglard, D.; Prasad, R. PAP1 poly(A) polymerase 1 homozygosity and hyperadenylation are major determinants of increased mRNA stability of CDR1 in azole-resistant clinical isolates of Candida albicans. Microbiology 2010, 156, 313–326. [Google Scholar] [CrossRef]
- Sui, Y.; Qi, L.; Wu, J.K.; Wen, X.P.; Tang, X.X.; Ma, Z.J.; Wu, X.C.; Zhang, K.; Kokoska, R.J.; Zheng, D.Q.; et al. Genome-wide mapping of spontaneous genetic alterations in diploid yeast cells. Proc. Natl. Acad. Sci. USA 2020, 117, 28191–28200. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.H.; Bennett, R.J. The impact of gene dosage and heterozygosity on the diploid pathobiont Candida albicans. J. Fungi 2020, 6, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rustad, T.R.; Stevens, D.A.; Pfaller, M.A.; White, T.C. Homozygosity at the Candida albicans MTL locus associated with azole resistance. Microbiology 2002, 148, 1061–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsit, S.; Leducq, J.B.; Durand, E.; Marchant, A.; Filteau, M.; Landry, C.R. Evolutionary biology through the lens of budding yeast comparative genomics. Nat. Rev. Genet. 2017, 18, 581–598. [Google Scholar] [CrossRef]
- Hiller, D.; Sanglard, D.; Morschhauser, J. Overexpression of the MDR1 gene is sufficient to confer increased resistance to toxic compounds in Candida albicans. Antimicrob. Agents Chemother. 2006, 50, 1365–1371. [Google Scholar] [CrossRef] [Green Version]
- Hickman, M.A.; Zeng, G.S.; Forche, A.; Hirakawa, M.P.; Abbey, D.; Harrison, B.D.; Wang, Y.M.; Su, C.H.; Bennett, R.J.; Wang, Y.; et al. The ‘obligate diploid’ Candida albicans forms mating-competent haploids. Nature 2016, 530, 242. [Google Scholar] [CrossRef]
- Bennett, R.J.; Forche, A.; Berman, J. Rapid mechanisms for generating genome diversity: Whole ploidy shifts, aneuploidy, and loss of heterozygosity. Cold Spring Harb. Perspect. Med. 2014, 4, a019604. [Google Scholar] [CrossRef]
- Smith, A.C.; Morran, L.T.; Hickman, M.A. Host defense mechanisms induce genome instability leading to rapid evolution in an opportunistic fungal pathogen. Infect. Immun. 2022, 90, 12. [Google Scholar] [CrossRef]
- Weil, T.; Santamaria, R.; Lee, W.; Rung, J.; Tocci, N.; Abbey, D.; Bezerra, A.R.; Carreto, L.; Moura, G.R.; Bayes, M.; et al. Adaptive mistranslation accelerates the evolution of fluconazole resistance and induces major genomic and gene expression alterations in Candida albicans. mSphere 2017, 2, e00167-17. [Google Scholar] [CrossRef] [Green Version]
- St Charles, J.; Hazkani-Covo, E.; Yin, Y.; Andersen, S.L.; Dietrich, F.S.; Greenwell, P.W.; Malc, E.; Mieczkowski, P.; Petes, T.D. High-resolution genome-wide analysis of irradiated (uv and gamma-rays) diploid yeast cells reveals a high frequency of genomic loss of heterozygosity (LOH) events. Genetics 2012, 190, 1267–1284. [Google Scholar] [CrossRef] [Green Version]
- de Vries, A.R.G.; Couwenberg, L.G.F.; van den Broek, M.; Cortes, P.D.; ter Horst, J.; Pronk, J.T.; Daran, J.M.G. Allele-specific genome editing using CRISPR-Cas9 is associated with loss of heterozygosity in diploid yeast. Nucleic Acids Res. 2019, 47, 1362–1372. [Google Scholar]
- Li, Y.X.; Wu, Y.; Ma, L.; Guo, Z.; Xiao, W.H.; Yuan, Y.J. Loss of heterozygosity by SCRaMbLEing. Sci. China Life Sci. 2019, 62, 381–393. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The mechanisms of drug resistance in fungi. (A) The resistance is induced by abnormal sterol substitution or accumulation. (B) The resistance is induced by abnormal efflux pump function.
Figure 1.
The mechanisms of drug resistance in fungi. (A) The resistance is induced by abnormal sterol substitution or accumulation. (B) The resistance is induced by abnormal efflux pump function.

Figure 2.
The aneuploidy-mediated fluconazole resistance. (A) The segmental aneuploidy i(5L) is related to fluconazole resistance in yeast. (B) The trisomy of chr3 is related to fluconazole resistance.
Figure 2.
The aneuploidy-mediated fluconazole resistance. (A) The segmental aneuploidy i(5L) is related to fluconazole resistance in yeast. (B) The trisomy of chr3 is related to fluconazole resistance.

Figure 3.
Three different types of LOH are associated with fluconazole resistance in yeast. (A) Short-range LOH that is usually produced by gene conversion through drug-resistant mutant genes, leading to an increased copy number of drug-resistant genes. (B) Long-range LOH. It results from RCO or BIR events, with intact replication or exchange from the break site to the telomere. (C) Whole-chromosome LOH. Nondisjunction or chromosome missegregation events can yield trisomic or monosomic progeny, respectively. Whole-chromosome LOH arises by either loss of the heterozygous homolog in trisomic individuals or reduplication of the hemizygous homolog in monosomic individuals. TAC1* and ERG11* represent the genes with mutations.
Figure 3.
Three different types of LOH are associated with fluconazole resistance in yeast. (A) Short-range LOH that is usually produced by gene conversion through drug-resistant mutant genes, leading to an increased copy number of drug-resistant genes. (B) Long-range LOH. It results from RCO or BIR events, with intact replication or exchange from the break site to the telomere. (C) Whole-chromosome LOH. Nondisjunction or chromosome missegregation events can yield trisomic or monosomic progeny, respectively. Whole-chromosome LOH arises by either loss of the heterozygous homolog in trisomic individuals or reduplication of the hemizygous homolog in monosomic individuals. TAC1* and ERG11* represent the genes with mutations.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, W.-Y.; Cai, H.-Q.; Qu, S.-Y.; Lin, W.-H.; Liang, C.-C.; Liu, H.; Xie, Z.-X.; Yuan, Y.-J. Genomic Variation-Mediating Fluconazole Resistance in Yeast. Biomolecules 2022, 12, 845. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12060845
AMA Style
Wang W-Y, Cai H-Q, Qu S-Y, Lin W-H, Liang C-C, Liu H, Xie Z-X, Yuan Y-J. Genomic Variation-Mediating Fluconazole Resistance in Yeast. Biomolecules. 2022; 12(6):845. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12060845
Chicago/Turabian StyleWang, Wen-Yao, Hong-Qing Cai, Si-Yuan Qu, Wei-Hao Lin, Cheng-Cheng Liang, Hao Liu, Ze-Xiong Xie, and Ying-Jin Yuan. 2022. "Genomic Variation-Mediating Fluconazole Resistance in Yeast" Biomolecules 12, no. 6: 845. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12060845
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.