
Kir4.2 Potassium Channels in Retinal Pigment Epithelial Cells In Vitro: Contribution to Cell Viability and Proliferation, and Down-Regulation by Vascular Endothelial Growth Factor

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. RNA Extraction and cDNA Synthesis
2.4. RT-PCR Analysis
2.5. Real-Time RT-PCR Analysis (qRT-PCR)
2.6. siRNA Transfection
2.7. Immunocytochemistry
2.8. Western Blot Analysis
2.9. ELISA, Cell Proliferation and Viability Assays
2.10. Statistical Analysis
3. Results
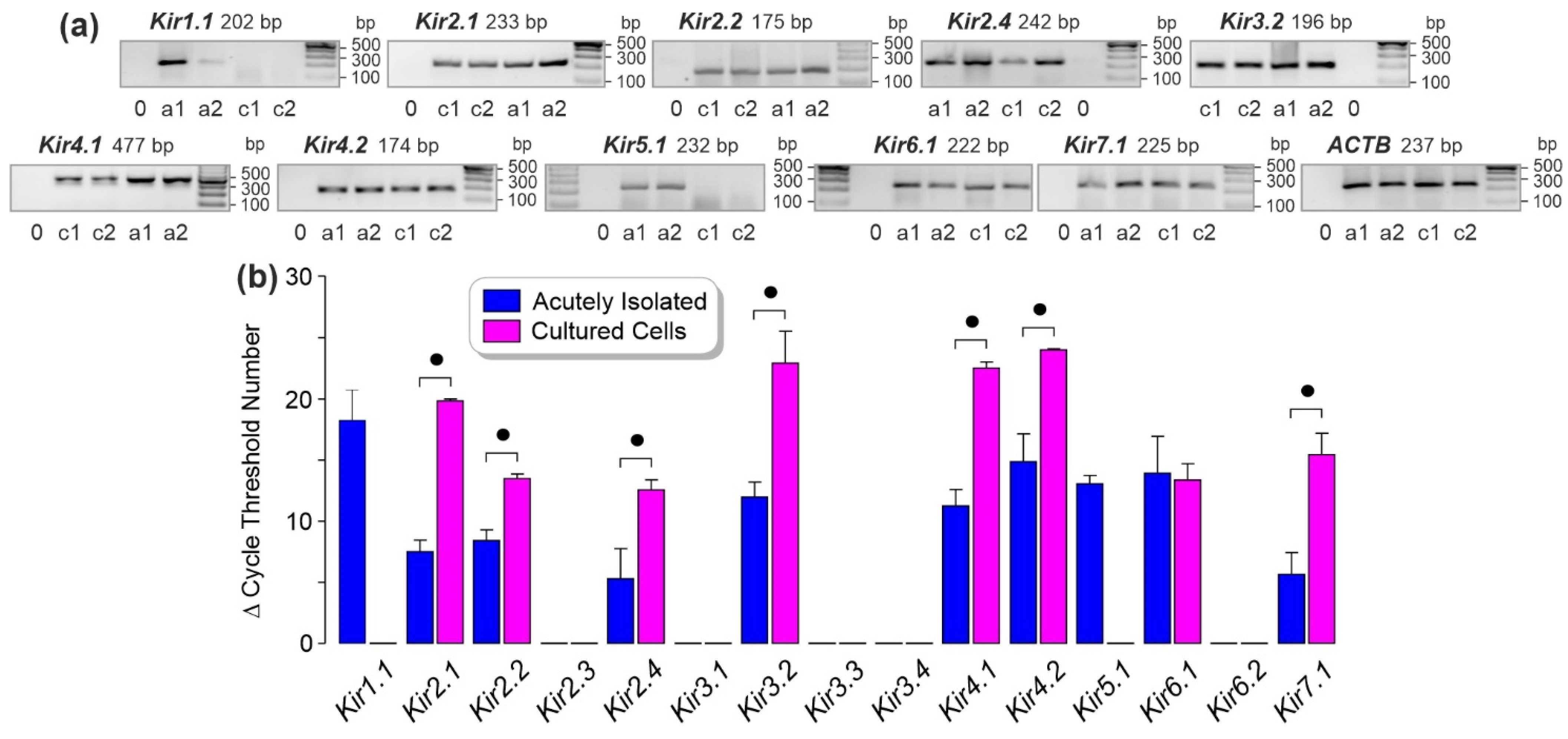
3.1. Kir Channel Gene Expression in Human RPE Cells
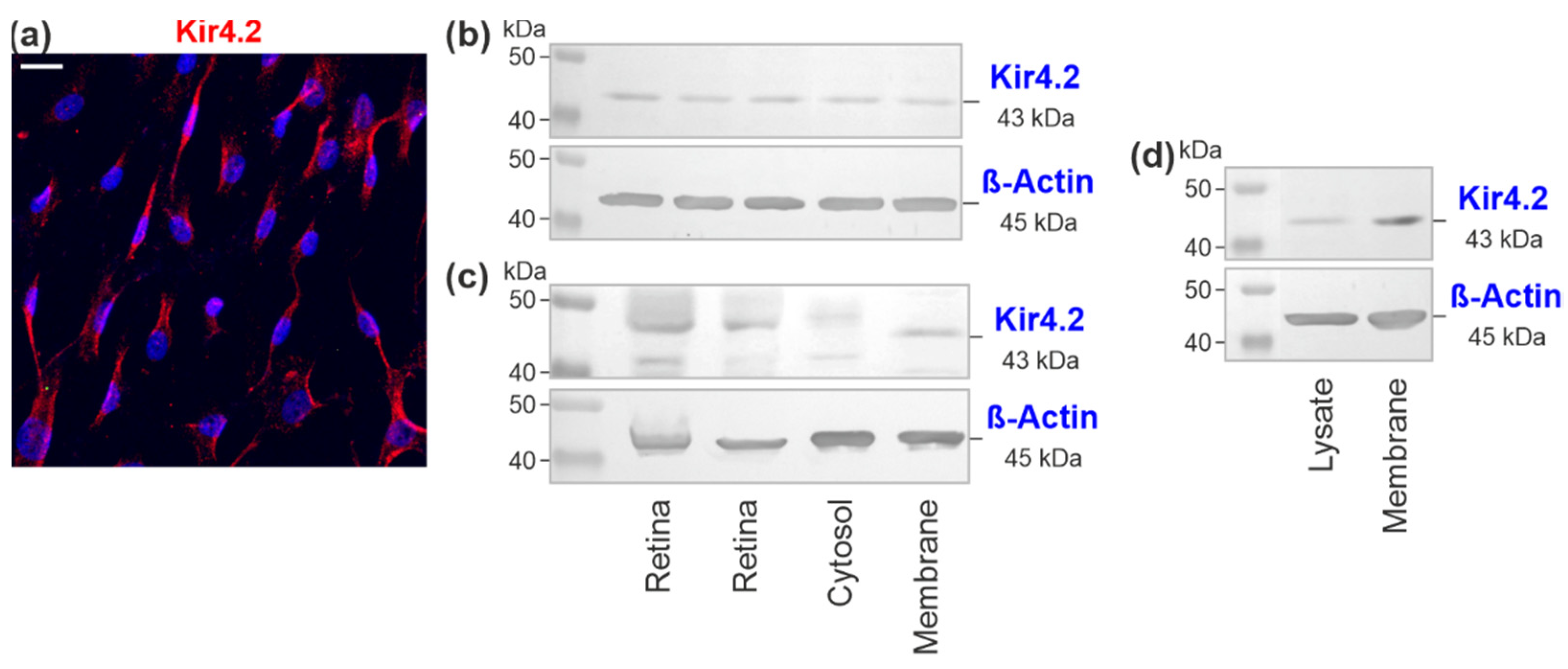
3.2. Kir4.2 Protein in Cultured RPE Cells
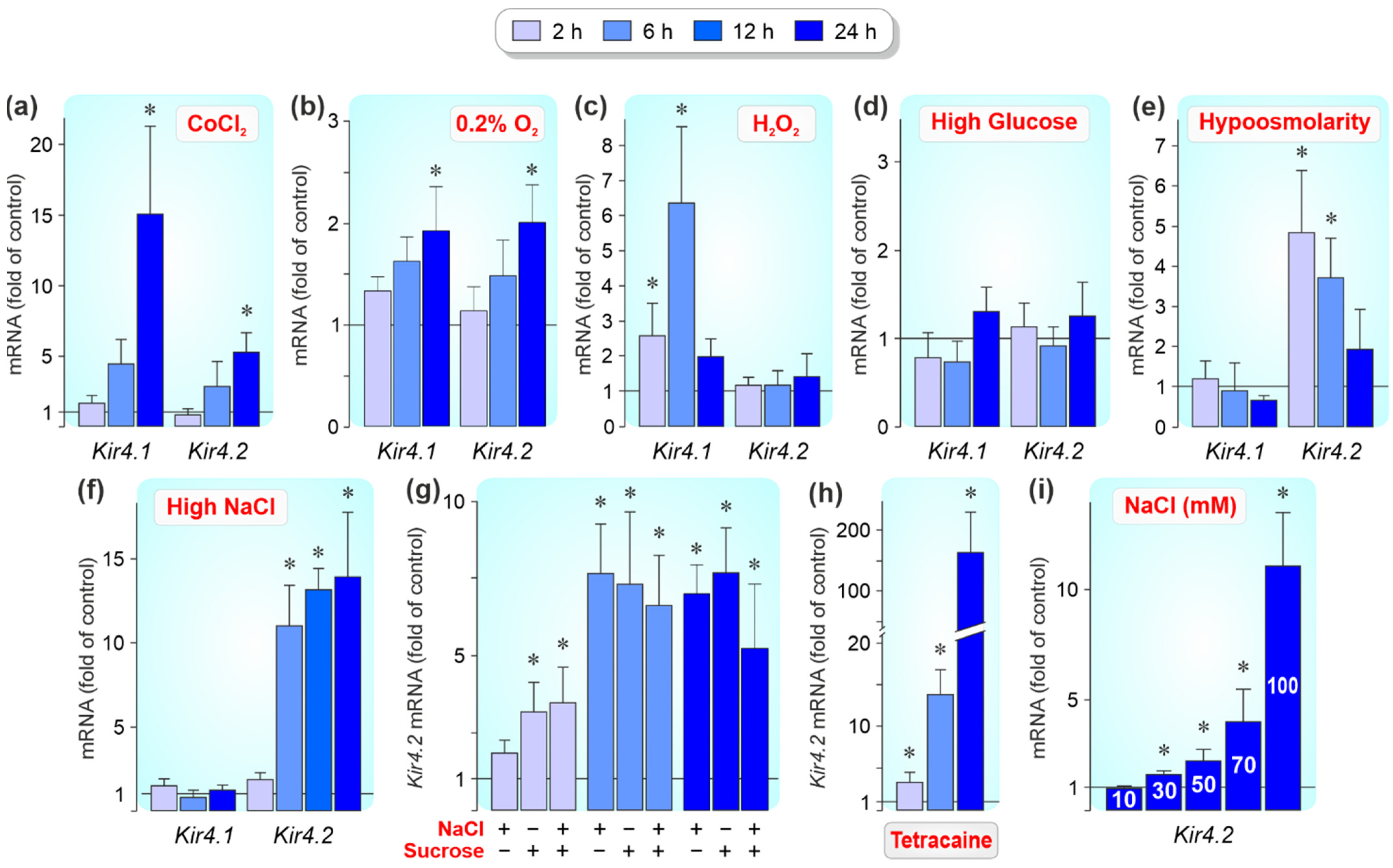
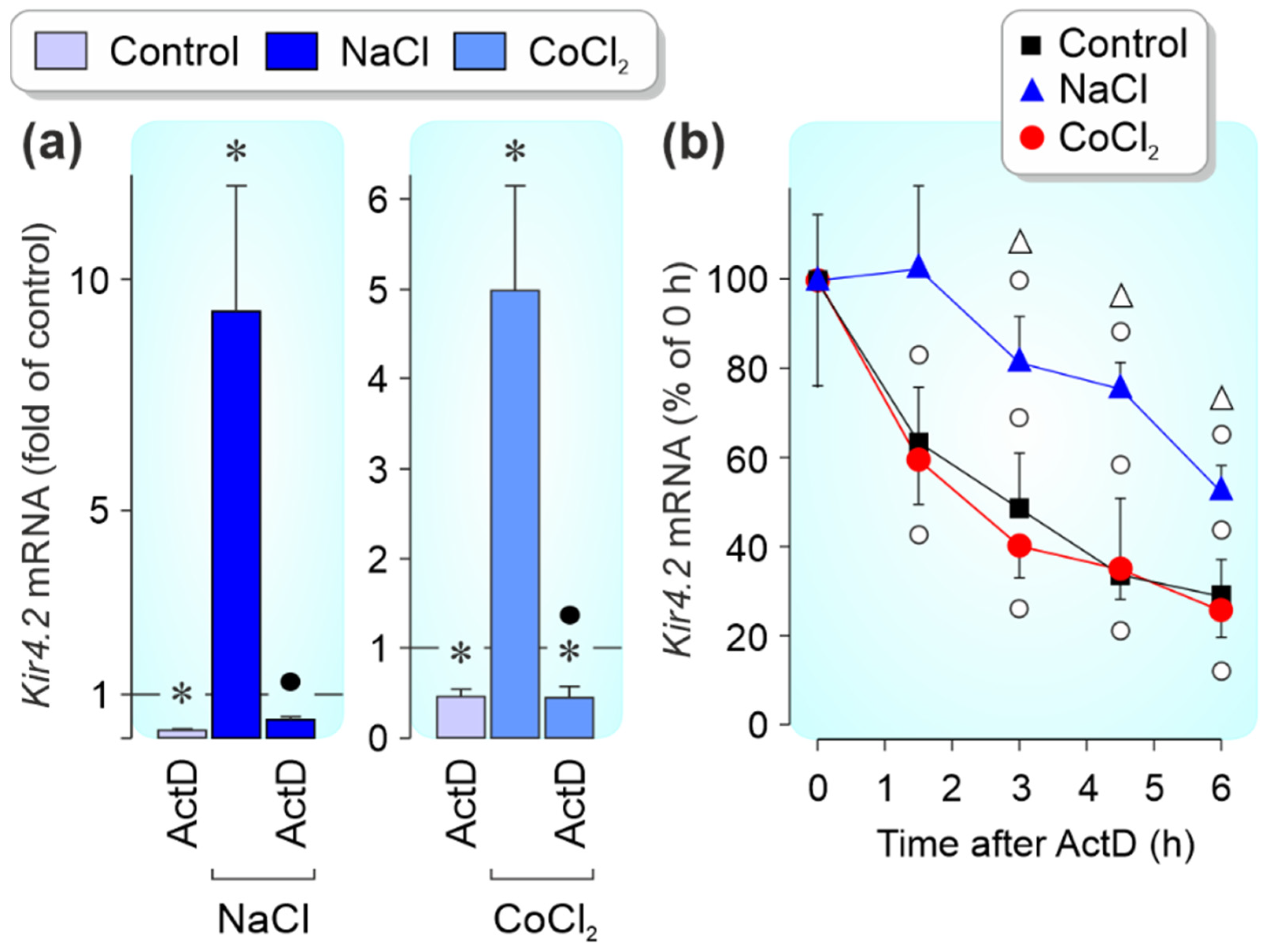
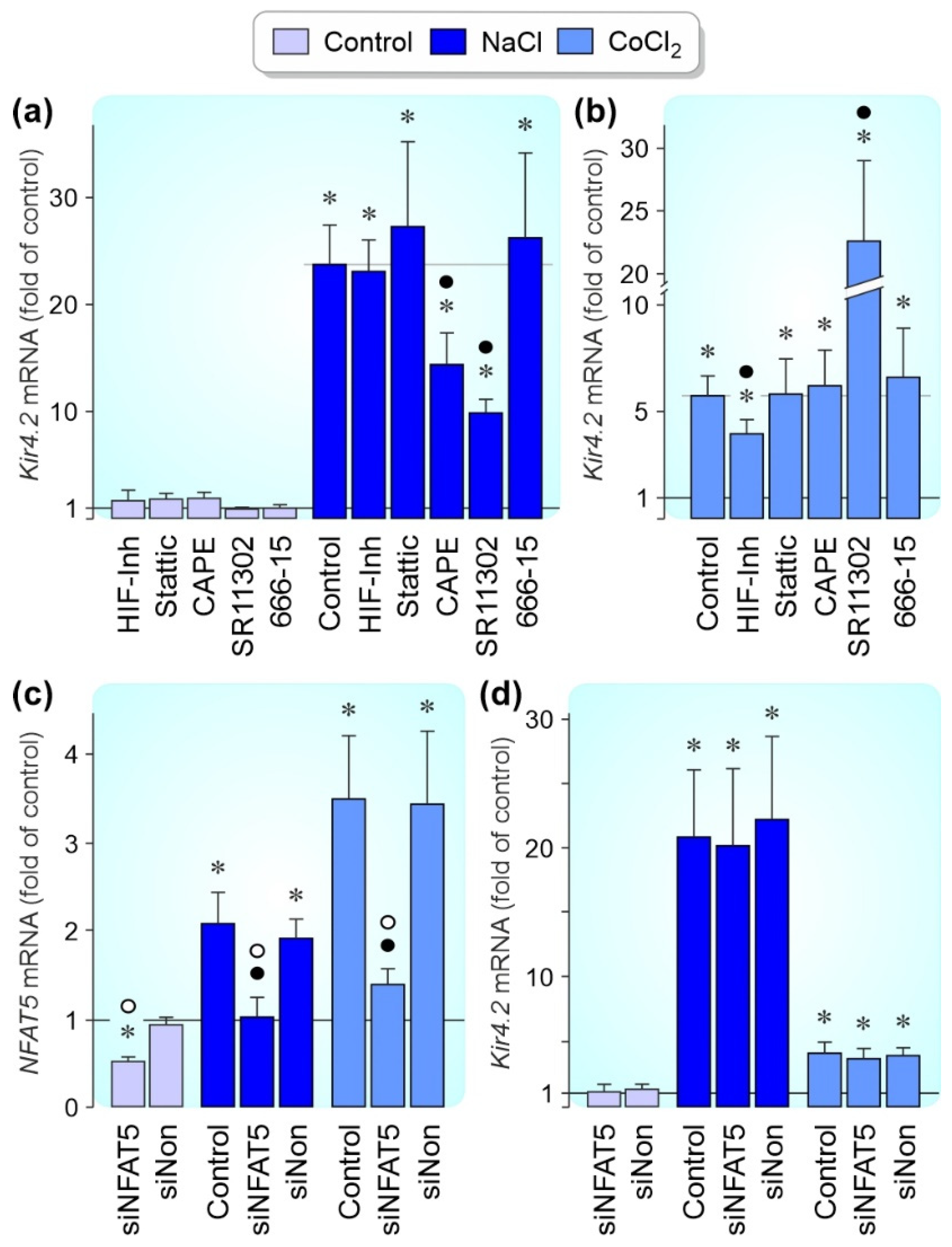
3.3. Regulation of Kir4.1 and Kir4.2 Gene Expression by Pathogenic Conditions
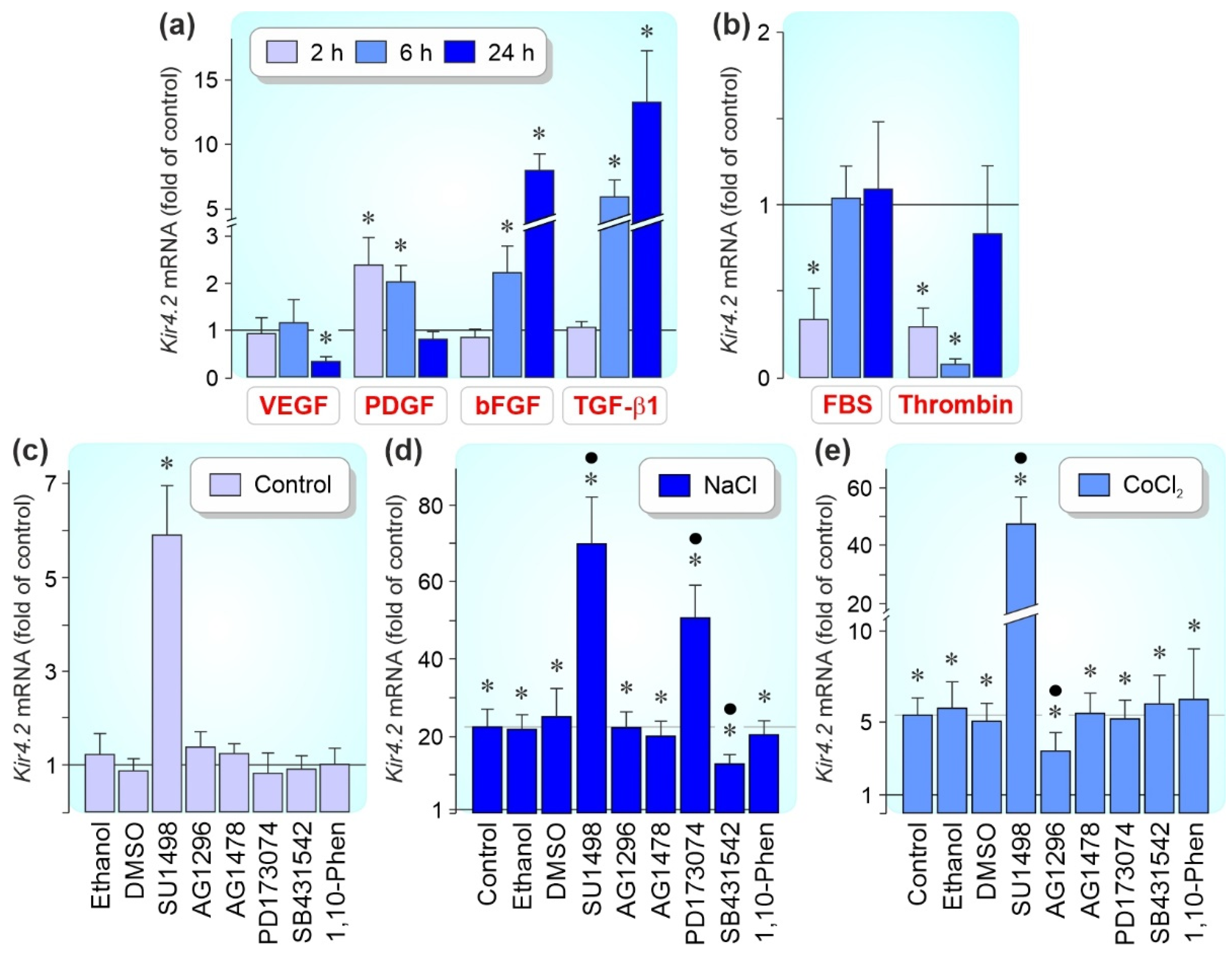
3.4. Regulation of Kir4.2 Gene Expression by Growth Factors
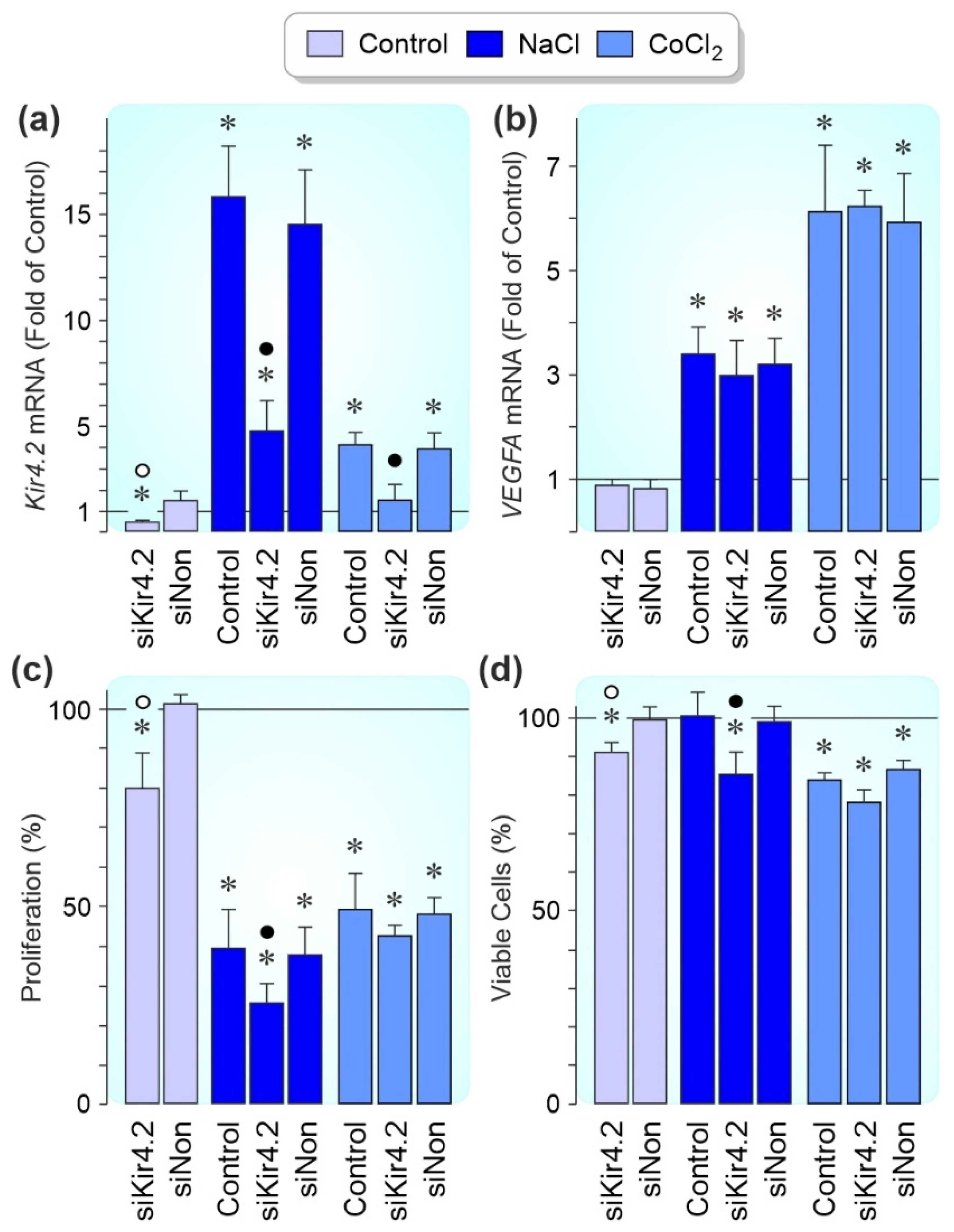
3.5. Effects of Kir4.2 Knockdown on Cell Proliferation and Survival
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Strauss, O. The Retinal Pigment Epithelium in Visual Function. Physiol. Rev. 2005, 85, 845–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bialek, S.; Miller, S.S. K+ and Cl-transport mechanisms in bovine pigment epithelium that could modulate subretinal space volume and composition. J. Physiol. 1994, 475, 401–417. [Google Scholar] [CrossRef] [PubMed]
- Pederson, J.E. Fluid physiology of the subretinal space. In Retina; Wilkinson, C.P., Ed.; Mosby: St Louis, MI, USA, 1994; pp. 1955–1968. [Google Scholar]
- Hughes, B.A.; Takahira, M. Inwardly rectifying K+ currents in isolated human retinal pigment epithelial cells. Investig. Ophthalmol. Vis. Sci. 1996, 37, 1125–1139. [Google Scholar]
- Yang, D.; Zhang, X.; Hughes, B.A. Expression of inwardly rectifying potassium channel subunits in native human retinal pig-ment epithelium. Exp. Eye Res. 2008, 87, 176–183. [Google Scholar] [CrossRef] [Green Version]
- Pearson, W.L.; Dourado, M.; Schreiber, M.; Salkoff, L.; Nichols, C.G. Expression of a functional Kir4 family inward rectifier K + channel from a gene cloned from mouse liver. J. Physiol. 1999, 514, 639–653. [Google Scholar] [CrossRef]
- Kusaka, S.; Inanobe, A.; Fujita, A.; Makino, Y.; Tanemoto, M.; Matsushita, K.; Tano, Y.; Kurachi, Y. Functional Kir7.1 channels localized at the root of apical processes in rat retinal pigment epithelium. J. Physiol. 2001, 531, 27–36. [Google Scholar] [CrossRef]
- Lopez, P.F.; Sippy, B.D.; Lambert, H.M.; Thach, A.B.; Hinton, D.R. Transdifferentiated retinal pigment epithelial cells are immuno-reactive for vascular endothelial growth factor in surgically excised age-related macular degeneration-related choroidal ne-ovascular membranes. Investig. Ophthalmol. Vis. Sci. 1996, 37, 855–868. [Google Scholar]
- Stern, J.; Temple, S. Retinal pigment epithelial cell proliferation. Exp. Biol. Med. 2015, 240, 1079–1086. [Google Scholar] [CrossRef] [Green Version]
- Al-Hussaini, H.; Kilarkaje, N. Effects of diabetes on retinal pigment epithelial cell proliferation and mitogen-activated protein kinase signaling in dark Agouti rats. Exp. Toxicol. Pathol. 2015, 67, 117–124. [Google Scholar] [CrossRef]
- Zhou, M.; Geathers, J.S.; Grillo, S.L.; Weber, S.R.; Wang, W.; Zhao, Y.; Sundstrom, J.M. Role of Epithelial-Mesenchymal Transition in Retinal Pigment Epithelium Dysfunction. Front. Cell Dev. Biol. 2020, 8, 501. [Google Scholar] [CrossRef]
- Ho, J.; Witkin, A.J.; Liu, J.; Chen, Y.; Fujimoto, J.G.; Schuman, J.; Duker, J.S. Documentation of Intraretinal Retinal Pigment Epithelium Migration via High-Speed Ultrahigh-Resolution Optical Coherence Tomography. Ophthalmology 2011, 118, 687–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christenbury, J.G.; Folgar, F.A.; O’Connell, R.V.; Chiu, S.J.; Farsiu, S.; Toth, C.A. Progression of Intermediate Age-related Macular Degeneration with Proliferation and Inner Retinal Migration of Hyperreflective Foci. Ophthalmology 2013, 120, 1038–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miura, M.; Makita, S.; Sugiyama, S.; Hong, Y.-J.; Yasuno, Y.; Elsner, A.E.; Tamiya, S.; Tsukahara, R.; Iwasaki, T.; Goto, H. Evaluation of intraretinal migration of retinal pigment epithelial cells in age-related macular degeneration using polarimetric imaging. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curcio, C.A.; Zanzottera, E.C.; Ach, T.; Balaratnasingam, C.; Freund, K.B. Activated Retinal Pigment Epithelium, an Optical Coherence Tomography Biomarker for Progression in Age-Related Macular Degeneration. Investig. Opthalmol. Vis. Sci. 2017, 58, BIO211–BIO226. [Google Scholar] [CrossRef]
- Roy, R.; Saurabh, K.; Shah, D.; Chowdhury, M.; Goel, S. Choroidal Hyperreflective Foci: A Novel Spectral Domain Optical Coherence Tomography Biomarker in Eyes with Diabetic Macular Edema. Asia-Pac. J. Ophthalmol. 2019, 8, 314–318. [Google Scholar] [CrossRef]
- Lei, J.; Balasubramanian, S.; Abdelfattah, N.S.; Nittala, M.G.; Sadda, S.R. Proposal of a simple optical coherence tomography-based scoring system for progression of age-related macular degeneration. Graefes Arch. Clin. Exp. Ophthalmol. 2017, 255, 1551–1558. [Google Scholar] [CrossRef]
- Nassisi, M.; Fan, W.; Shi, Y.; Lei, J.; Borrelli, E.; Ip, M.; Sadda, S.R. Quantity of Intraretinal Hyperreflective Foci in Patients with Intermediate Age-Related Macular Degeneration Correlates With 1-Year Progression. Investig. Opthalmol. Vis. Sci. 2018, 59, 3431–3439. [Google Scholar] [CrossRef] [Green Version]
- Grossniklaus, H.E.; Ling, J.X.; Wallace, T.M.; Dithmar, S.; Lawson, D.H.; Cohen, C.; Elner, V.; Elner, S.G.; Sternberg, P. Macrophage and retinal pigment epithelium expression of angiogenic cytokines in choroidal neovascularization. Mol. Vis. 2002, 8, 119–126. [Google Scholar]
- Miller, J.W.; Le Couter, J.; Strauss, E.C.; Ferrara, N. Vascular Endothelial Growth Factor A in Intraocular Vascular Disease. Ophthalmology 2013, 120, 106–114. [Google Scholar] [CrossRef]
- Saika, S.; Kono-Saika, S.; Tanaka, T.; Yamanaka, O.; Ohnishi, Y.; Sato, M.; Muragaki, Y.; Ooshima, A.; Yoo, J.; Flanders, K.C.; et al. Smad3 is required for dedifferentiation of retinal pigment epithelium following retinal detachment in mice. Lab. Investig. 2004, 84, 1245–1258. [Google Scholar] [CrossRef]
- Rehak, M.; Hollborn, M.; Iandiev, I.; Pannicke, T.; Karl, A.; Wurm, A.; Kohen, L.; Reichenbach, A.; Wiedemann, P.; Bringmann, A. Retinal gene expression and Müller cell responses after branch retinal vein occlusion in the rat. Investig. Ophthalmol. Vis. Sci. 2009, 50, 2359–2367. [Google Scholar] [CrossRef] [PubMed]
- Radeke, M.J.; Radeke, C.M.; Shih, Y.-H.; Hu, J.; Bok, D.; Johnson, L.V.; Coffey, P.J. Restoration of mesenchymal retinal pigmented epithelial cells by TGFβ pathway inhibitors: Implications for age-related macular degeneration. Genome Med. 2015, 7, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, R.; Klein, B.E.; Moss, S.E.; Cruickshanks, K.J. The Wisconsin epidemiologic study of diabetic retinopathy: XVII: The 14-year incidence and progression of diabetic retinopathy and associated risk factors in type 1 diabetes. Ophthalmology 1998, 105, 1801–1815. [Google Scholar] [CrossRef]
- Klein, R.; Klein, B.E.; Tomany, S.C.; Cruickshanks, K.J. The association of cardiovascular disease with the long-term incidence of age-related maculopathy: The beaver dam eye study. Ophthalmology 2003, 110, 1273–1280. [Google Scholar] [CrossRef]
- Van Leeuwen, R.; Ikram, M.K.; Vingerling, J.R.; Witteman, J.C.; Hofman, A.; de Jong, P.T. Blood pressure, atherosclerosis, and the incidence of age-related maculopathy: The Rotterdam Study. Investig. Ophthalmol. Vis. Sci. 2003, 44, 3771–3777. [Google Scholar] [CrossRef] [Green Version]
- Kamoi, K.; Takeda, K.; Hashimoto, K.; Tanaka, R.; Okuyama, S. Identifying risk factors for clinically significant diabetic macula edema in patients with type 2 diabetes mellitus. Curr. Diabetes Rev. 2013, 9, 209–217. [Google Scholar] [CrossRef]
- Lifton, R.P.; Gharavi, A.G.; Geller, D.S. Molecular Mechanisms of Human Hypertension. Cell 2001, 104, 545–556. [Google Scholar] [CrossRef] [Green Version]
- He, F.J.; Markandu, N.D.; Sagnella, G.A.; de Wardener, H.E.; MacGregor, G.A. Plasma sodium: Ignored and underestimated. Hypertension 2005, 45, 98–102. [Google Scholar] [CrossRef]
- Neuhofer, W. Role of NFAT5 in Inflammatory Disorders Associated with Osmotic Stress. Curr. Genom. 2010, 11, 584–590. [Google Scholar] [CrossRef] [Green Version]
- Obika, L.O.; Amabebe, E.; Ozoene, J.O.; Inneh, C.A. Thirst perception, plasma osmolality and estimated plasma arginine vaso-pressin concentration in dehydrated and oral saline loaded subjects. Niger. J. Physiol. Sci. 2013, 28, 83–89. [Google Scholar]
- Bringmann, A.; Hollborn, M.; Kohen, L.; Wiedemann, P. Intake of dietary salt and drinking water: Implications for the devel-opment of age-related macular degeneration. Mol. Vis. 2016, 22, 1437–1454. [Google Scholar] [PubMed]
- Chen, R.; Hollborn, M.; Grosche, A.; Reichenbach, A.; Wiedemann, P.; Bringmann, A.; Kohen, L. Effects of the vegetable polyphenols epigallocatechin-3-gallate, luteolin, apigenin, myricetin, quercetin, and cyanidin in primary cultures of human retinal pigment epithelial cells. Mol. Vis. 2014, 20, 242–258. [Google Scholar] [PubMed]
- An, W.G.; Kanekal, M.; Simon, M.C.; Maltepe, E.; Blagosklonny, M.V.; Neckers, L.M. Stabilization of wild-type p53 by hypox-ia-inducible factor 1α. Nature 1998, 392, 405–408. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Sánchez, J.; Chánez-Cárdenas, M.E. The use of cobalt chloride as a chemical hypoxia model. J. Appl. Toxicol. 2019, 39, 556–570. [Google Scholar] [CrossRef]
- Cicinnati, V.R.; Shen, Q.; Sotiropoulos, G.C.; Radtke, A.; Gerken, G.; Beckebaum, S. Validation of putative reference genes for gene expression studies in human hepatocellular carcinoma using real-time quantitative RT-PCR. BMC Cancer 2008, 8, 350. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Cahalan, M. Local anesthetic block of sodium channels in normal and pronase-treated squid giant axons. Biophys. J. 1978, 23, 285–311. [Google Scholar] [CrossRef] [Green Version]
- Natarajan, K.; Singh, S.; Burke, T.R., Jr.; Grunberger, D.; Aggarwal, B.B. Caffeic acid phenethyl ester is a potent and specific inhibitor of activation of nuclear transcription factor NF-κB. Proc. Natl. Acad. Sci. USA 1996, 93, 9090–9095. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Lee, J.H.; Boovanahalli, S.K.; Jin, Y.; Lee, M.; Jin, X.; Kim, J.H.; Hong, A.Y.-S.; Lee, J.J. (Aryloxyacetylamino)benzoic Acid Analogues: A New Class of Hypoxia-Inducible Factor-1 Inhibitors. J. Med. Chem. 2007, 50, 1675–1684. [Google Scholar] [CrossRef]
- Schust, J.; Sperl, B.; Hollis, A.; Mayer, T.U.; Berg, T. Stattic: A Small-Molecule Inhibitor of STAT3 Activation and Dimerization. Chem. Biol. 2006, 13, 1235–1242. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Kwon, M.H.; Woo, S. TonEBP transcriptional activator in the cellular response to increased osmolality. Pflügers Arch. 2002, 444, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Hollborn, M.; Vogler, S.; Reichenbach, A.; Wiedemann, P.; Bringmann, A.; Kohen, L. Regulation of the hyperosmotic induction of aquaporin 5 and VEGF in retinal pigment epithelial cells: Involvement of NFAT5. Mol. Vis. 2015, 21, 360–377. [Google Scholar] [PubMed]
- Veltmann, M.; Hollborn, M.; Reichenbach, A.; Wiedemann, P.; Kohen, L.; Bringmann, A. Osmotic Induction of Angiogenic Growth Factor Expression in Human Retinal Pigment Epithelial Cells. PLoS ONE 2016, 11, e0147312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doktor, F.; Prager, P.; Wiedemann, P.; Kohen, L.; Bringmann, A.; Hollborn, M. Hypoxic expression of NLRP3 and VEGF in cultured retinal pigment epithelial cells: Contribution of P2Y2 receptor signaling. Purinergic Signal. 2018, 14, 471–484. [Google Scholar] [CrossRef]
- Burg, M.B.; Ferraris, J.D.; Dmitrieva, N. Cellular Response to Hyperosmotic Stresses. Physiol. Rev. 2007, 87, 1441–1474. [Google Scholar] [CrossRef]
- Arsenijevic, T.; Vujovic, A.; Libert, F.; De Beeck, A.O.; Hébrant, A.; Janssens, S.; Gregoire, F.; Lefort, A.; Bolaky, N.; Perret, J.; et al. Hyperosmotic stress induces cell cycle arrest in retinal pigmented epithelial cells. Cell Death Dis. 2013, 4, e662. [Google Scholar] [CrossRef] [Green Version]
- Winges, A.; Garcia, T.B.; Prager, P.; Wiedemann, P.; Kohen, L.; Bringmann, A.; Hollborn, M. Osmotic expression of aldose reductase in retinal pigment epithelial cells: Involvement of NFAT5. Graefe’s Arch. Clin. Exp. Ophthalmol. 2016, 254, 2387–2400. [Google Scholar] [CrossRef]
- Urrego, D.; Tomczak, A.P.; Zahed, F.; Stühmer, W.; Pardo, L.A. Potassium channels in cell cycle and cell proliferation. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130094. [Google Scholar] [CrossRef] [Green Version]
- Kondratskyi, A.; Kondratska, K.; Skryma, R.; Prevarskaya, N. Ion channels in the regulation of apoptosis. Biochim. Biophys. Acta Biomembr. 2015, 1848, 2532–2546. [Google Scholar] [CrossRef] [Green Version]
- Capiod, T. Cell proliferation, calcium influx and calcium channels. Biochimie 2011, 93, 2075–2079. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Accession Number | Gene ID OMIM | Primer Sequences (5′ → 3′) | Product (bp) |
|---|---|---|---|
| ACTBa,b NM_001101 | 60 102630 | sATGGCCACGGCTGCTTCCAGC asCATGGTGGTGCCGCCAGACAG | 237 |
| β2-microglobulin (B2M)b NM_004048.2 | 567 109700 | sTTGAAAAAGTGGAGCATTCAGA asTCGATCCCACTTAACTATCTTGG | 155 |
| KCNJ1 (Kir1.1)a,b NM_153766.2 | 3758 600359 | sGTGGAGGCACAGTCAAGGTT asCCACACAGGGAGTGTGATTG | 202 |
| KCNJ2 (Kir2.1)a,b NM_000891.2 | 3759 600681 | sCGGTGGATGCTGGTTATCTT asAAAACAGCAATTGGGCATTC | 233 |
| KCNJ12 (Kir2.2)a,b NM_021012.4 | 3768 602323 | sAGAAGAATGGCCAGTGCAAC asGCGATGACCCAGAAGATGAT | 175 |
| KCNJ4 (Kir2.3)a,b NM_004981.1 | 3761 600504 | sCCATCATCATTGTCCACGAG asGAAGACCACAGGCTCAAAGC | 197 |
| KCNJ14 (Kir2.4)a,b NM_013348.3 | 3770 603953 | sGATCGACTCTGCCAGTCCTC asTCCCTGGGACCTCATAAGTG | 242 |
| KCNJ3 (Kir3.1)a,b NM_002239.3 | 3760 601534 | sTGTGGAAACAACTGGGATGA asGGGACGACATGAGAAGCATT | 203 |
| KCNJ6 (Kir3.2)a,b NM_002240.4 | 3763 600877 | sGCTACCGGGTCATCACAGAT asATCAGGCACAGTTTCCCATC | 196 |
| KCNJ9 (Kir3.3)a,b NM_004983.2 | 3765 600932 | sGCTACCTGACGGACCTGTTC asAAGCCGTTGAGGTTGTTGAC | 193 |
| KCNJ5 (Kir3.4)a,b NM_000890.3 | 3762 600734 | sACCGATTCACACCAGTCCTC asATTCTGCTCAGCCTCTGCAT | 212 |
| KCNJ10 (Kir4.1)a,b NM_002241.4 | 3766 602208 | sCCAGGGATACGACGGCGGAGA asGAAACGAATGGTCTCAGCCCG | 477 |
| KCNJ15 (Kir4.2)a NM_002243.4 | 3772 602106 | sCCGTTCCATCACAGAGGAAT asTTCTGCTTGGTGATGACTGC | 174 |
| KCNJ15 (Kir4.2)b NM_002243.4 | sAGGTAGCCAATATGAGGAAGAGC asACAAGCTCAAACTCCTTCTCCTT | 247 | |
| KCNJ16 (Kir5.1)a,b NM_018658.2 | 3773 605722 | sTCCACTGGAACATCTCACCA asACGTGCAGGATTCTCGAACT | 232 |
| KCNJ8 (Kir6.1)a,b NM_004982.3 | 3764 600935 | sGGAGGGAGGATGATGACAGA asTTTCCTCAGGTCACCCACTC | 222 |
| KCNJ11 (Kir6.2)a,b NM_000525.3 | 3767 600937 | sATCATCGTCATCCTGGAAGG asGGTGTTGCCAAACTTGGAG | 162 |
| KCNJ13 (Kir7.1)a,b NM_002242.4 | 3769 603208 | sTCACATGGATGGCAAACCTA asGCCAGAGGACTTGATGGTGT | 225 |
| NFAT5b NM_006599.3 | 10725 604708 | sTCACCATCATCTTCCCACCT asCTGCAATAGTGCATCGCTGT | 174 |
| VEGFA188, 164, 120b NM_003376.5 NM_001287044.1 NM_001025370.2 | 7422 192240 | sCCTGGTGGACATCTTCCAGGAGTA asCTCACCGCCTCGGCTTGTCACA | 479; 407; 275 |
| Method | Antibody | Source | Catalog | Dilution Concentration |
|---|---|---|---|---|
| ICC | rabbit anti-human Kir4.2 | Sigma-Aldrich, Taufkirchen, Germany | HPA016702 | 1:100 2 µg/mL |
| Alexa568-coupled goat anti-rabbit IgG | Invitrogen, Paisley, UK | A11036 | 1:500 4 µg/mL | |
| WB | rabbit anti-human Kir4.2 | Sigma-Aldrich | HPA016702 | 1:500 0.4 µg/mL |
| mouse anti-human Kir4.2 | Santa Cruz Biotechnology, Heidelberg, Germany | sc-376322 | 1:600 0.33 µg/mL | |
| rabbit anti-β-actin | Cell Signaling Technology, Frankfurt/M., Germany | 8457 | 1:1000 37 ng/mL | |
| alkaline phosphatase-coupled goat anti-rabbit IgG | Cell Signaling Technology | 7054 | 1:2000 35.5 ng/mL | |
| alkaline phosphatase-coupled goat anti-mouse IgG | Cell Signaling Technology | 7056 | 1:2000 55.5 ng/mL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beer, M.-C.; Kuhrt, H.; Kohen, L.; Wiedemann, P.; Bringmann, A.; Hollborn, M. Kir4.2 Potassium Channels in Retinal Pigment Epithelial Cells In Vitro: Contribution to Cell Viability and Proliferation, and Down-Regulation by Vascular Endothelial Growth Factor. Biomolecules 2022, 12, 848. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12060848
Beer M-C, Kuhrt H, Kohen L, Wiedemann P, Bringmann A, Hollborn M. Kir4.2 Potassium Channels in Retinal Pigment Epithelial Cells In Vitro: Contribution to Cell Viability and Proliferation, and Down-Regulation by Vascular Endothelial Growth Factor. Biomolecules. 2022; 12(6):848. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12060848
Chicago/Turabian StyleBeer, Marie-Christin, Heidrun Kuhrt, Leon Kohen, Peter Wiedemann, Andreas Bringmann, and Margrit Hollborn. 2022. "Kir4.2 Potassium Channels in Retinal Pigment Epithelial Cells In Vitro: Contribution to Cell Viability and Proliferation, and Down-Regulation by Vascular Endothelial Growth Factor" Biomolecules 12, no. 6: 848. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12060848