
Macrophages Are a Double-Edged Sword: Molecular Crosstalk between Tumor-Associated Macrophages and Cancer Stem Cells

 , and
, and
Abstract
:1. Introduction
2. Secretory Molecules of CSCs Induce Macrophage Recruitment and Tumor-Promoting Characteristics of the TME
2.1. CSCs Recruit Macrophages into the TME

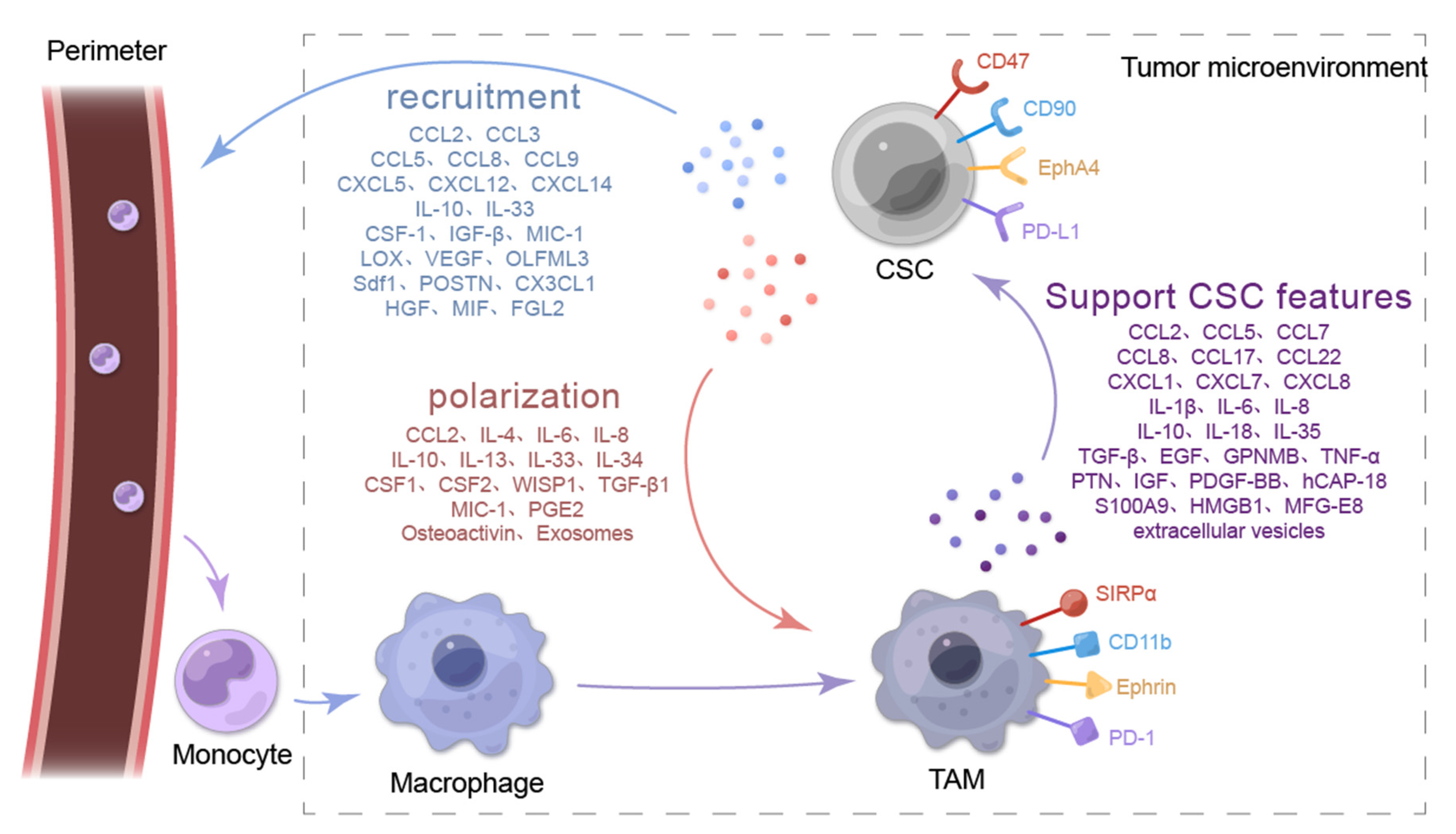{kind=link}
{kind=link}
| Chemokines Produced by CSCs | Chemokine Receptor | Type of Cancer | Associated Signaling Pathway/Mechanism of Chemokine Secretion | Reference |
|---|---|---|---|---|
| CCL2 1 | CCR2 | Glioblastoma | IFNγ | [47] |
| Breast cancer | β-Catenin | [48] | ||
| Bladder cancer | Not shown | [39] | ||
| Hepatocellular carcinoma | Hippo | [43] | ||
| CCL3 | CCR1/CCR5 | Bladder cancer | Not shown | [49,50] |
| Leukemia | Not shown | [51] | ||
| CCL5 | CCR5, CCR1, CCR2 | Glioblastoma | PI3K 2/AKT | [52,53] |
| Optic glioma | Not shown | [54] | ||
| Hepatocellular carcinoma | Not shown | [55] | ||
| CCL8 | CCR2 | Bladder cancer | Not shown | [56] |
| Cutaneous squamous cell carcinoma | Not shown | [57] | ||
| CCL9 | CCR1 | Non-small cell lung cancer | Not shown | [58] |
| CXCL5 3 | CXCR2 | Hepatocellular carcinoma | Sox9 | [59] |
| CXCL12 | CXCR4 | Leukemia | Not shown | [42] |
| Colon cancer | NF-κB 4 | [60] | ||
| CXCL14 | Not shown | Lung cancer | Not shown | [61] |
| IL-10 | IL-10R | Ovarian cancer | WNT | [62] |
| IL-33 | ST2 | Squamous cell carcinoma | NRF2 | [44] |
| CSF1 5 | CSF1R | Hepatocellular carcinoma | Hippo | [43] |
| Non-small cell lung cancer | IRF5 | [63] | ||
| Glioblastoma | STAT3 6 | [64] | ||
| TGF-β 7 | Not shown | Glioblastoma | STAT3 | [64] |
| MIC-1 8 | Not shown | Glioblastoma | Not shown | [64] |
| Pancreatic cancer | Not shown | [65] | ||
| LOX 9 | Not shown | Pancreatic cancer | Not shown | [66] |
| VEGF 10 | VEGFR | Colorectal cancer | Six1/MAPK 11 & PI3K/AKT/mTOR | [67,68] |
| OLFML3 12 | Not shown | Glioblastoma | CLOCK-BMAL1 | [40] |
| Sdf1 13 | Cxcr4b | Brain tumor | AKT1 | [41] |
| POSTN 14 | Integrinαvβ₃ | Glioblastoma | Not shown | [69] |
| CX3CL1 | CX3CR1 | Testicular germ cell tumors | Not shown | [70] |
| HGF 15 | Not shown | Glioblastoma | HGF-MET axis | [71] |
| MIF | CXCR4 | Glioblastoma | Not shown | [72] |
| FGL2 | Not shown | Glioblastoma | Not shown | [73] |
2.1.1. CSCs Recruit Macrophages through the CC Chemokine Family and the CXC Chemokine Subfamily
2.1.2. CSCs Recruit Macrophages by Secreting IL-33
2.1.3. CSCs Recruit Macrophages through Other Types of Secreted Proteins
2.2. CSCs Promote Macrophage Polarization in the TME
| Chemokines Produced by CSCs | Chemokine Receptor | Type of Cancer | Associated Signaling Pathway/Mechanism of Cytokine Secretion | Reference |
|---|---|---|---|---|
| CCL2 | CCR2 | Breast cancer | β-catenin/CCL2 axis | [48] |
| IL-4 | IL-4R | Breast cancer | ERK1/2 1 | [88] |
| IL-6 | IL-6R | Glioblastoma | TLR4 | [89] |
| Triple negative breast cancer | HLF/MCT-1 | [90,91] | ||
| IL-8 | IL-8R | Ovarian cancer | STAT3 | [92] |
| IL-10 | IL-10R | Ovarian cancer | PPARγ 2/NF-κB | [93] |
| IL-13 | IL-13R | Cholangiocarcinoma | Not shown | [94] |
| IL-34 | IL-34R | Cholangiocarcinoma | Not shown | [94] |
| IL-33 | ST-2 | Squamous cell carcinoma | NRF2 | [44] |
| CSF1 | CSF1R | Glioblastoma | Not shown | [64] |
| Non-small cell lung cancer | Oct4 | [95] | ||
| Breast cancer | Not shown | [96] | ||
| CSF2 | CSFR2 | Triple negative breast cancer | NF-κB | [97] |
| Pancreatic cancer | Not shown | [98] | ||
| WISP1 | Integrinα6β1 | Glioblastoma | WNT/β-catenin | [99] |
| TGF-β | TGFBR | Glioblastoma | Not shown | [64] |
| Pancreatic cancer | Not shown | [98] | ||
| Hepatocellular carcinoma | TLR4 | [100] | ||
| MIC-1 | Not shown | Glioblastoma | Not shown | [64] |
| PGE2 3 | EP2 | Ovarian cancer | COX-2 4/PGE2 | [101] |
| Glioblastoma | ARS2 5/MAGL | [102] | ||
| Osteoactivin | Not shown | Cholangiocarcinoma | Not shown | [94] |
| Exosomes | Not shown | Pancreatic cancer | Not shown | [103,104] |
| Glioblastoma | Not shown | [105] | ||
| Esophageal squamous cell carcinoma | Not shown | [106] |
2.2.1. CSCs Induce Macrophage Polarization by Secreting CC Chemokines
2.2.2. CSCs Promote Macrophage Polarization by Secreting ILs
2.2.3. CSCs Promote Macrophage Polarization through Other Cytokines and Soluble Protein Molecules
2.2.4. CSCs Polarize Macrophages by Secreting Exosomes
3. TAMs Enhance the Stemness Characteristics of Tumor Cells
| TAM Type | Factor of TAM Production | Type of Cancer | Associated Signaling Pathway/Mechanism of TAMs on CSCs | Reference |
|---|---|---|---|---|
| M2 | CCL2 | Breast cancer | β-catenin | [48] |
| CCL5 | Prostate cancer | β-catenin/STAT3 | [110] | |
| Glioblastoma | Not shown | [52] | ||
| CCL7 | Ovarian cancer | CCR3/MM9 1 | [111] | |
| CCL8 | Glioblastoma | CCR1/CCR5/ERK1/2 | [112] | |
| Gastric cancer | JAK1/STAT3 | [113] | ||
| CCL17 | Hepatocellular carcinoma | WNT/β-catenin | [114] | |
| CCL22 | Hepatocellular carcinoma | Not shown | [115] | |
| CXCL1 | Breast cancer | Not shown | [116] | |
| CXCL7 | Glioblastoma | Not shown | [73] | |
| CXCL8 | Breast cancer | CXCR2 | [117] | |
| IL-1β | Glioblastoma | p38 MAPK | [118] | |
| IL-6 | Glioblastoma | Not shown | [89] | |
| Ovarian cancer | WNT | [62] | ||
| Hepatocellular carcinoma | STAT3 | [119] | ||
| Breast cancer | STAT3 | [120] | ||
| IL-8 | Ovarian cancer | JAK2/STAT3 | [92] | |
| IL-10 | Ovarian cancer | WNT | [62] | |
| Non-small cell lung cancer | JAK1/STAT1/NF-κB/Notch1 | [121] | ||
| IL-18 | Gastric cancer | Not shown | [122] | |
| IL-35 | Breast cancer | Not shown | [123] | |
| TGF-β | Hepatocellular carcinoma | EMT | [124] | |
| Pancreatic cancer | TGF-β1/smad2/3 axis | [125] | ||
| Squamous cell carcinoma | NRF2 | [44] | ||
| Lung cancer | Not shown | [126] | ||
| Breast cancer | EMT | [127] | ||
| TNF-α | Clear cell renal cell carcinoma | NF-κB | [128] | |
| Chordoma | PI3K/AKT | [129] | ||
| EGF 1 | Breast cancer | EGFR/STAT3/Sox2 | [130] | |
| GPNMB 2 | Breast cancer | PI3K/AKT/mTOR & β-catenin/MAPKs/AMPK/Src | [131] | |
| PTN 3 | Lymphoma | PTPRZ1/β-catenin | [132] | |
| Glioblastoma | PTN-PTPRZ1 | [133] | ||
| IGF 4 | Thyroid cancer | PI3K/AKT/mTOR | [134] | |
| PDGF-BB 5 | Breast cancer | Not shown | [135] | |
| hCAP-18 | Pancreatic ductal adenocarcinoma | Not shown | [136] | |
| S100A9 6 | Hepatocellular carcinoma | NF-κB | [137] | |
| HMGB1 7 | Non-small cell lung cancer | NF-κB | [138] | |
| MFG-E8 8 | Colorectal cancer | STAT3/Sonic Hedgehog | [139] | |
| extracellular vesicles | Pancreatic cancer | KLF3 | [140] | |
| M1 | IL-6 | Oral squamous cell carcinoma | JAK/STAT3 | [141] |
| IL-12 | Glioblastoma | Not shown | [142] | |
| TNF-α | Lung cancer | Not shown | [143] | |
| Oral cancer | Not shown | [144] |
3.1. CC and CXC Chemokines Are Secreted by TAMs to Maintain Tumor Cell Stemness Characteristics
3.2. TAMs Support the Stemness of Tumor Cells by Secreting ILs
3.3. Other Cytokines and Soluble Protein Molecules Secreted by TAMs to Promote the Stemness of Tumor Cells
3.4. TAMs Promote Tumor Cell Stemness through Surface Protein Molecules
3.5. TAMs Promote Tumor Cell Stemness by Secreting Extracellular Vesicles
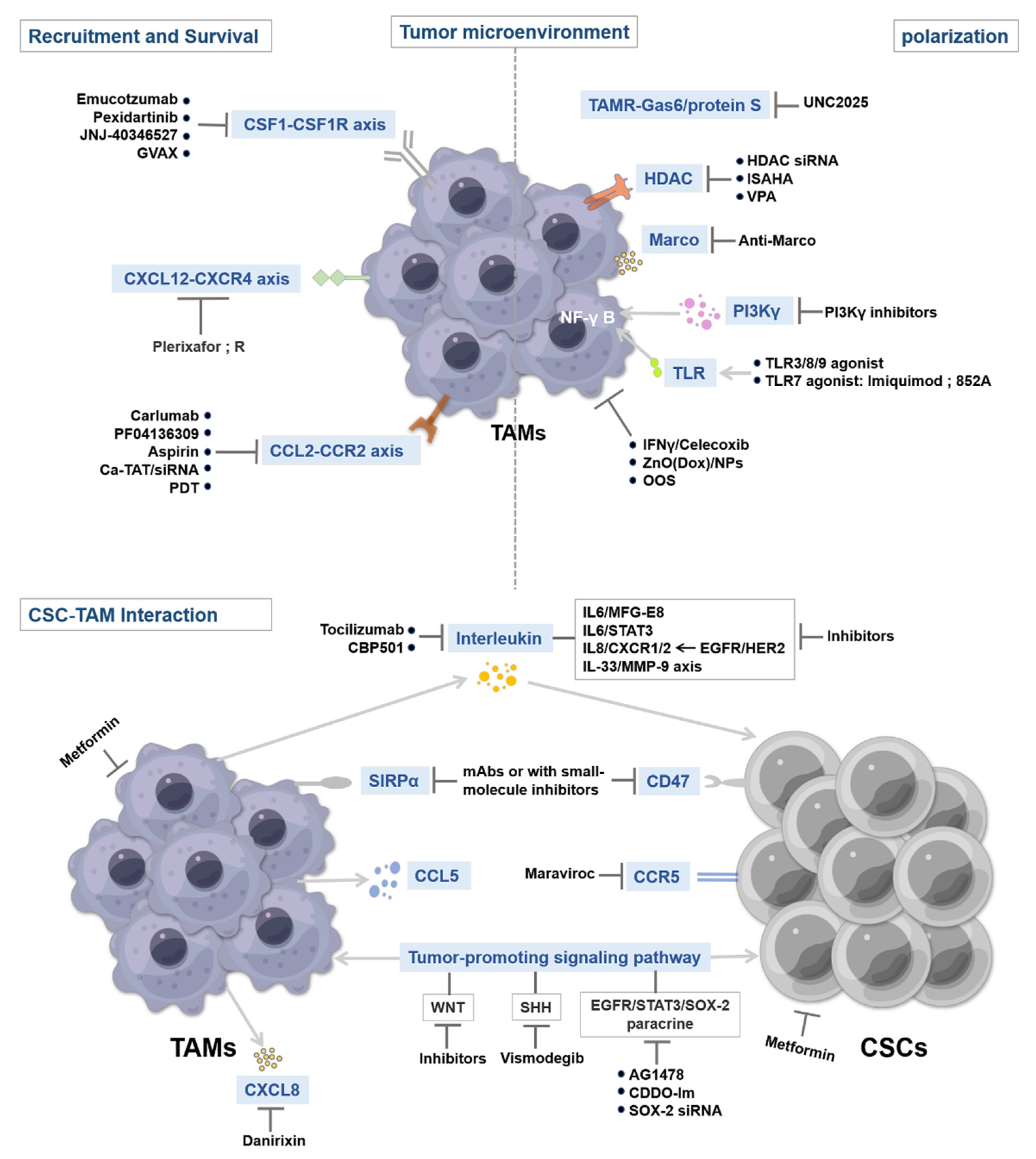
4. Therapeutic Strategies Targeting CSCs and TAMs
4.1. Targeting Tumor-Associated Macrophage Recruitment and Survival
4.1.1. CSF1–CSF1R Inhibitors
4.1.2. CCL2–CCR2 Inhibitors
4.1.3. CXCL12–CXCR4 Inhibitors
4.2. Targeting TAM Activation
4.2.1. TAMR Inhibitors
4.2.2. Anti-MARCO Antibody
4.2.3. Toll-Like Receptors and Stimulator of Interferon Gene Agonists
4.2.4. Others
4.3. Targeting CSC–TAM Interaction
4.3.1. Blockade of the SIRP-α–CD47 Pathway
4.3.2. Targeting IL-Mediated Signaling Pathways
4.3.3. Others
5. Summary and Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, H.M.; Zhang, J.G.; Zhang, X.; Li, Q. Targeting cancer stem cells for reversing therapy resistance: Mechanism, signaling, and prospective agents. Signal Transduct. Target. Ther. 2021, 6, 62. [Google Scholar] [CrossRef] [PubMed]
- Lapidot, T.; Sirard, C.; Vormoor, J.; Murdoch, B.; Hoang, T.; Caceres-Cortes, J.; Minden, M.; Paterson, B.; Caligiuri, M.A.; Dick, J.E. A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature 1994, 367, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Abdoli Shadbad, M.; Hosseinkhani, N.; Asadzadeh, Z.; Derakhshani, A.; Karim Ahangar, N.; Hemmat, N.; Lotfinejad, P.; Brunetti, O.; Silvestris, N.; Baradaran, B. A Systematic Review to Clarify the Prognostic Values of CD44 and CD44(+)CD24(-) Phenotype in Triple-Negative Breast Cancer Patients: Lessons Learned and the Road Ahead. Front. Oncol. 2021, 11, 689839. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Liu, C.; Yao, J.; Wan, H.; Wan, G.; Li, Y.; Chen, N. Breast cancer stem cells, heterogeneity, targeting therapies and therapeutic implications. Pharmacol. Res. 2021, 163, 105320. [Google Scholar] [CrossRef]
- Kemper, K.; Prasetyanti, P.R.; De Lau, W.; Rodermond, H.; Clevers, H.; Medema, J.P. Monoclonal antibodies against Lgr5 identify human colorectal cancer stem cells. Stem Cells 2012, 30, 2378–2386. [Google Scholar] [CrossRef]
- Wang, Z.; Tang, Y.; Xie, L.; Huang, A.; Xue, C.; Gu, Z.; Wang, K.; Zong, S. The Prognostic and Clinical Value of CD44 in Colorectal Cancer: A Meta-Analysis. Front. Oncol. 2019, 9, 309. [Google Scholar] [CrossRef]
- Huang, J.L.; Oshi, M.; Endo, I.; Takabe, K. Clinical relevance of stem cell surface markers CD133, CD24, and CD44 in colorectal cancer. Am. J. Cancer Res. 2021, 11, 5141–5154. [Google Scholar]
- Chong, C.; Müller, M.; Pak, H.; Harnett, D.; Huber, F.; Grun, D.; Leleu, M.; Auger, A.; Arnaud, M.; Stevenson, B.J.; et al. Integrated proteogenomic deep sequencing and analytics accurately identify non-canonical peptides in tumor immunopeptidomes. Nat. Commun. 2020, 11, 1293. [Google Scholar] [CrossRef] [Green Version]
- Rambow, F.; Marine, J.C.; Goding, C.R. Melanoma plasticity and phenotypic diversity: Therapeutic barriers and opportunities. Genes Dev. 2019, 33, 1295–1318. [Google Scholar] [CrossRef] [Green Version]
- Prieto-Vila, M.; Takahashi, R.U.; Usuba, W.; Kohama, I.; Ochiya, T. Drug Resistance Driven by Cancer Stem Cells and Their Niche. Int. J. Mol. Sci. 2017, 18, 2574. [Google Scholar] [CrossRef] [Green Version]
- Plaks, V.; Kong, N.; Werb, Z. The cancer stem cell niche: How essential is the niche in regulating stemness of tumor cells? Cell Stem Cell 2015, 16, 225–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Ye, P.; Chen, Y.; Liu, T.; Cha, J.H.; Yan, X.; Yang, W.H. Involvement of the Estrogen and Progesterone Axis in Cancer Stemness: Elucidating Molecular Mechanisms and Clinical Significance. Front. Oncol. 2020, 10, 1657. [Google Scholar] [CrossRef] [PubMed]
- Al Haq, A.T.; Tseng, H.Y.; Chen, L.M.; Wang, C.C.; Hsu, H.L. Targeting prooxidant MnSOD effect inhibits triple-negative breast cancer (TNBC) progression and M2 macrophage functions under the oncogenic stress. Cell Death Dis. 2022, 13, 49. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Yu, Y.; Wang, X.; Zhang, T. Tumor-Associated Macrophages in Tumor Immunity. Front. Immunol. 2020, 11, 583084. [Google Scholar] [CrossRef]
- Song, W.; Mazzieri, R.; Yang, T.; Gobe, G.C. Translational Significance for Tumor Metastasis of Tumor-Associated Macrophages and Epithelial-Mesenchymal Transition. Front. Immunol. 2017, 8, 1106. [Google Scholar] [CrossRef] [Green Version]
- Zeng, J.; Liu, Z.; Sun, S.; Xie, J.; Cao, L.; Lv, P.; Nie, S.; Zhang, B.; Xie, B.; Peng, S.; et al. Tumor-associated macrophages recruited by periostin in intrahepatic cholangiocarcinoma stem cells. Oncol. Lett. 2018, 15, 8681–8686. [Google Scholar] [CrossRef]
- Zhang, H.; Lu, H.; Xiang, L.; Bullen, J.W.; Zhang, C.; Samanta, D.; Gilkes, D.M.; He, J.; Semenza, G.L. HIF-1 regulates CD47 expression in breast cancer cells to promote evasion of phagocytosis and maintenance of cancer stem cells. Proc. Natl. Acad. Sci. USA 2015, 112, E6215–E6223. [Google Scholar] [CrossRef] [Green Version]
- Castelli, G.; Pelosi, E.; Testa, U. Liver Cancer: Molecular Characterization, Clonal Evolution and Cancer Stem Cells. Cancers 2017, 9, 127. [Google Scholar] [CrossRef] [Green Version]
- Cioffi, M.; Trabulo, S.; Hidalgo, M.; Costello, E.; Greenhalf, W.; Erkan, M.; Kleeff, J.; Sainz, B., Jr.; Heeschen, C. Inhibition of CD47 Effectively Targets Pancreatic Cancer Stem Cells via Dual Mechanisms. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2015, 21, 2325–2337. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Zhang, L.; Yang, L.; Li, H.; Li, R.; Yu, J.; Yang, L.; Wei, F.; Yan, C.; Sun, Q.; et al. Anti-CD47 Antibody as a Targeted Therapeutic Agent for Human Lung Cancer and Cancer Stem Cells. Front. Immunol. 2017, 8, 404. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Clauser, K.R.; Tam, W.L.; Fröse, J.; Ye, X.; Eaton, E.N.; Reinhardt, F.; Donnenberg, V.S.; Bhargava, R.; Carr, S.A.; et al. A breast cancer stem cell niche supported by juxtacrine signalling from monocytes and macrophages. Nat. Cell Biol. 2014, 16, 1105–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage Polarization: Different Gene Signatures in M1(LPS+) vs. Classically and M2(LPS-) vs. Alternatively Activated Macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef] [PubMed]
- Guerra, A.D.; Yeung, O.W.H.; Qi, X.; Kao, W.J.; Man, K. The Anti-Tumor Effects of M1 Macrophage-Loaded Poly (ethylene glycol) and Gelatin-Based Hydrogels on Hepatocellular Carcinoma. Theranostics 2017, 7, 3732–3744. [Google Scholar] [CrossRef] [PubMed]
- Nusblat, L.M.; Carroll, M.J.; Roth, C.M. Crosstalk between M2 macrophages and glioma stem cells. Cell. Oncol. 2017, 40, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.J.; Xiang, S.F.; Chen, Y.Q.; Zhang, N.; Cao, J.; Zhu, H.; Yang, B.; Zhou, Q.; Ying, M.D.; He, Q.J. Inhibition of M2-like macrophages by all-trans retinoic acid prevents cancer initiation and stemness in osteosarcoma cells. Acta Pharmacol. Sin. 2019, 40, 1343–1350. [Google Scholar] [CrossRef]
- Ito, A.; Kagawa, S.; Sakamoto, S.; Kuwada, K.; Kajioka, H.; Yoshimoto, M.; Kikuchi, S.; Kuroda, S.; Yoshida, R.; Tazawa, H.; et al. Extracellular vesicles shed from gastric cancer mediate protumor macrophage differentiation. BMC Cancer 2021, 21, 102. [Google Scholar] [CrossRef]
- Pedraza-Brindis, E.J.; Sánchez-Reyes, K.; Hernández-Flores, G.; Bravo-Cuellar, A.; Jave-Suárez, L.F.; Aguilar-Lemarroy, A.; Gómez-Lomelí, P.; López-López, B.A.; Ortiz-Lazareno, P.C. Culture supernatants of cervical cancer cells induce an M2 phenotypic profile in THP-1 macrophages. Cell. Immunol. 2016, 310, 42–52. [Google Scholar] [CrossRef]
- Sánchez-Reyes, K.; Pedraza-Brindis, E.J.; Hernández-Flores, G.; Bravo-Cuellar, A.; López-López, B.A.; Rosas-González, V.C.; Ortiz-Lazareno, P.C. The supernatant of cervical carcinoma cells lines induces a decrease in phosphorylation of STAT-1 and NF-κB transcription factors associated with changes in profiles of cytokines and growth factors in macrophages derived from U937 cells. Innate Immun. 2019, 25, 344–355. [Google Scholar] [CrossRef]
- Wang, D.; Wang, X.; Si, M.; Yang, J.; Sun, S.; Wu, H.; Cui, S.; Qu, X.; Yu, X. Exosome-encapsulated miRNAs contribute to CXCL12/CXCR4-induced liver metastasis of colorectal cancer by enhancing M2 polarization of macrophages. Cancer Lett. 2020, 474, 36–52. [Google Scholar] [CrossRef]
- Gok Yavuz, B.; Gunaydin, G.; Gedik, M.E.; Kosemehmetoglu, K.; Karakoc, D.; Ozgur, F.; Guc, D. Cancer associated fibroblasts sculpt tumour microenvironment by recruiting monocytes and inducing immunosuppressive PD-1(+) TAMs. Sci. Rep. 2019, 9, 3172. [Google Scholar] [CrossRef]
- Cavalcante, R.S.; Ishikawa, U.; Silva, E.S.; Silva-Júnior, A.A.; Araújo, A.A.; Cruz, L.J.; Chan, A.B.; de Araújo Júnior, R.F. STAT3/NF-κB signalling disruption in M2 tumour-associated macrophages is a major target of PLGA nanocarriers/PD-L1 antibody immunomodulatory therapy in breast cancer. Br. J. Pharmacol. 2021, 178, 2284–2304. [Google Scholar] [CrossRef] [PubMed]
- Ouzounova, M.; Lee, E.; Piranlioglu, R.; El Andaloussi, A.; Kolhe, R.; Demirci, M.F.; Marasco, D.; Asm, I.; Chadli, A.; Hassan, K.A.; et al. Monocytic and granulocytic myeloid derived suppressor cells differentially regulate spatiotemporal tumour plasticity during metastatic cascade. Nat. Commun. 2017, 8, 14979. [Google Scholar] [CrossRef] [PubMed]
- Clara, J.A.; Monge, C.; Yang, Y.; Takebe, N. Targeting signalling pathways and the immune microenvironment of cancer stem cells—A clinical update. Nat. Rev. Clin. Oncol. 2020, 17, 204–232. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zheng, Y.; Zhao, M. Exosome-Based Cancer Therapy: Implication for Targeting Cancer Stem Cells. Front. Pharmacol. 2016, 7, 533. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.H.; Wang, Y.N.; Yang, W.H.; Xia, W.; Wei, Y.; Chan, L.C.; Wang, Y.H.; Jiang, Z.; Xu, S.; Yao, J.; et al. Human ribonuclease 1 serves as a secretory ligand of ephrin A4 receptor and induces breast tumor initiation. Nat. Commun. 2021, 12, 2788. [Google Scholar] [CrossRef]
- Cheng, H.; Wang, Z.; Fu, L.; Xu, T. Macrophage Polarization in the Development and Progression of Ovarian Cancers: An Overview. Front. Oncol. 2019, 9, 421. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Hsu, W.H.; Han, J.; Xia, Y.; DePinho, R.A. Cancer Stemness Meets Immunity: From Mechanism to Therapy. Cell Rep. 2021, 34, 108597. [Google Scholar] [CrossRef]
- Wei, J.; Chen, P.; Gupta, P.; Ott, M.; Zamler, D.; Kassab, C.; Bhat, K.P.; Curran, M.A.; de Groot, J.F.; Heimberger, A.B. Immune biology of glioma-associated macrophages and microglia: Functional and therapeutic implications. Neuro-Oncol. 2020, 22, 180–194. [Google Scholar] [CrossRef]
- Kobatake, K.; Ikeda, K.I.; Nakata, Y.; Yamasaki, N.; Ueda, T.; Kanai, A.; Sentani, K.; Sera, Y.; Hayashi, T.; Koizumi, M.; et al. Kdm6a Deficiency Activates Inflammatory Pathways, Promotes M2 Macrophage Polarization, and Causes Bladder Cancer in Cooperation with p53 Dysfunction. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2020, 26, 2065–2079. [Google Scholar] [CrossRef]
- Chen, P.; Hsu, W.H.; Chang, A.; Tan, Z.; Lan, Z.; Zhou, A.; Spring, D.J.; Lang, F.F.; Wang, Y.A.; DePinho, R.A. Circadian Regulator CLOCK Recruits Immune-Suppressive Microglia into the GBM Tumor Microenvironment. Cancer Discov. 2020, 10, 371–381. [Google Scholar] [CrossRef] [Green Version]
- Chia, K.; Mazzolini, J.; Mione, M.; Sieger, D. Tumor initiating cells induce Cxcr4-mediated infiltration of pro-tumoral macrophages into the brain. eLife 2018, 7, e31918. [Google Scholar] [CrossRef] [PubMed]
- Roversi, F.M.; Bueno, M.L.P.; Pericole, F.V.; Saad, S.T.O. Hematopoietic Cell Kinase (HCK) Is a Player of the Crosstalk between Hematopoietic Cells and Bone Marrow Niche through CXCL12/CXCR4 Axis. Front. Cell Dev. Biol. 2021, 9, 634044. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhao, Y.; Yan, H.; Yang, Y.; Shen, S.; Dai, X.; Ji, X.; Ji, F.; Gong, X.G.; Li, L.; et al. Single tumor-initiating cells evade immune clearance by recruiting type II macrophages. Genes Dev. 2017, 31, 247–259. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, S.; Elhance, A.; Van Duzer, A.; Kumar, S.; Leitenberger, J.J.; Oshimori, N. Tumor-initiating cells establish an IL-33-TGF-β niche signaling loop to promote cancer progression. Science 2020, 369, eaay1813. [Google Scholar] [CrossRef]
- Pathria, P.; Louis, T.L.; Varner, J.A. Targeting Tumor-Associated Macrophages in Cancer. Trends Immunol. 2019, 40, 310–327. [Google Scholar] [CrossRef]
- Qian, B.Z.; Li, J.; Zhang, H.; Kitamura, T.; Zhang, J.; Campion, L.R.; Kaiser, E.A.; Snyder, L.A.; Pollard, J.W. CCL2 recruits inflammatory monocytes to facilitate breast-tumour metastasis. Nature 2011, 475, 222–225. [Google Scholar] [CrossRef] [Green Version]
- Gangoso, E.; Southgate, B.; Bradley, L.; Rus, S.; Galvez-Cancino, F.; McGivern, N.; Güç, E.; Kapourani, C.A.; Byron, A.; Ferguson, K.M.; et al. Glioblastomas acquire myeloid-affiliated transcriptional programs via epigenetic immunoediting to elicit immune evasion. Cell 2021, 184, 2454–2470.e2426. [Google Scholar] [CrossRef]
- Zhang, F.; Li, P.; Liu, S.; Yang, M.; Zeng, S.; Deng, J.; Chen, D.; Yi, Y.; Liu, H. β-Catenin-CCL2 feedback loop mediates crosstalk between cancer cells and macrophages that regulates breast cancer stem cells. Oncogene 2021, 40, 5854–5865. [Google Scholar] [CrossRef]
- Cheah, M.T.; Chen, J.Y.; Sahoo, D.; Contreras-Trujillo, H.; Volkmer, A.K.; Scheeren, F.A.; Volkmer, J.P.; Weissman, I.L. CD14-expressing cancer cells establish the inflammatory and proliferative tumor microenvironment in bladder cancer. Proc. Natl. Acad. Sci. USA 2015, 112, 4725–4730. [Google Scholar] [CrossRef] [Green Version]
- Korbecki, J.; Grochans, S.; Gutowska, I.; Barczak, K.; Baranowska-Bosiacka, I. CC Chemokines in a Tumor: A Review of Pro-Cancer and Anti-Cancer Properties of Receptors CCR5, CCR6, CCR7, CCR8, CCR9, and CCR10 Ligands. Int. J. Mol. Sci. 2020, 21, 7619. [Google Scholar] [CrossRef]
- Baba, T.; Mukaida, N. Role of macrophage inflammatory protein (MIP)-1α/CCL3 in leukemogenesis. Mol. Cell. Oncol. 2014, 1, e29899. [Google Scholar] [CrossRef] [PubMed]
- Novak, M.; Koprivnikar Krajnc, M.; Hrastar, B.; Breznik, B.; Majc, B.; Mlinar, M.; Rotter, A.; Porčnik, A.; Mlakar, J.; Stare, K.; et al. CCR5-Mediated Signaling Is Involved in Invasion of Glioblastoma Cells in Its Microenvironment. Int. J. Mol. Sci. 2020, 21, 4199. [Google Scholar] [CrossRef] [PubMed]
- Kranjc, M.K.; Novak, M.; Pestell, R.G.; Lah, T.T. Cytokine CCL5 and receptor CCR5 axis in glioblastoma multiforme. Radiol. Oncol. 2019, 53, 397–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Pan, Y.; Gutmann, D.H. Genetic and genomic alterations differentially dictate low-grade glioma growth through cancer stem cell-specific chemokine recruitment of T cells and microglia. Neuro-Oncol. 2019, 21, 1250–1262. [Google Scholar] [CrossRef]
- Zhuang, Y.; Zhao, X.; Yuan, B.; Zeng, Z.; Chen, Y. Blocking the CCL5-CCR5 Axis Using Maraviroc Promotes M1 Polarization of Macrophages Cocultured with Irradiated Hepatoma Cells. J. Hepatocell. Carcinoma 2021, 8, 599–611. [Google Scholar] [CrossRef]
- Li, Y.; Chen, X.; Li, D.; Yang, Z.; Bai, Y.; Hu, S.; Liu, Z.; Gu, J.; Zhang, X. Identification of prognostic and therapeutic value of CC chemokines in Urothelial bladder cancer: Evidence from comprehensive bioinformatic analysis. BMC Urol. 2021, 21, 173. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Wang, P.; Zhou, Q.; Zhu, L.; Zhang, H.; Zhang, Y.; Zheng, Z.; Bhatta, A.K.; Zhang, G.; Wang, X. CCL8 enhances sensitivity of cutaneous squamous cell carcinoma to photodynamic therapy by recruiting M1 macrophages. Photodiagn. Photodyn. Ther. 2019, 26, 235–243. [Google Scholar] [CrossRef]
- Kortlever, R.M.; Sodir, N.M.; Wilson, C.H.; Burkhart, D.L.; Pellegrinet, L.; Brown Swigart, L.; Littlewood, T.D.; Evan, G.I. Myc Cooperates with Ras by Programming Inflammation and Immune Suppression. Cell 2017, 171, 1301–1315.e1314. [Google Scholar] [CrossRef]
- Ren, Z.; Chen, Y.; Shi, L.; Shao, F.; Sun, Y.; Ge, J.; Zhang, J.; Zang, Y. Sox9/CXCL5 axis facilitates tumour cell growth and invasion in hepatocellular carcinoma. FEBS J. 2022. [Google Scholar] [CrossRef]
- Jiang, X.; Cao, G.; Gao, G.; Wang, W.; Zhao, J.; Gao, C. Triptolide decreases tumor-associated macrophages infiltration and M2 polarization to remodel colon cancer immune microenvironment via inhibiting tumor-derived CXCL12. J. Cell. Physiol. 2021, 236, 193–204. [Google Scholar] [CrossRef]
- Takiguchi, S.; Korenaga, N.; Inoue, K.; Sugi, E.; Kataoka, Y.; Matsusue, K.; Futagami, K.; Li, Y.J.; Kukita, T.; Teramoto, N.; et al. Involvement of CXCL14 in osteolytic bone metastasis from lung cancer. Int. J. Oncol. 2014, 44, 1316–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghavan, S.; Mehta, P.; Xie, Y.; Lei, Y.L.; Mehta, G. Ovarian cancer stem cells and macrophages reciprocally interact through the WNT pathway to promote pro-tumoral and malignant phenotypes in 3D engineered microenvironments. J. Immunother. Cancer 2019, 7, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashina, T.; Baghdadi, M.; Yoneda, A.; Kinoshita, I.; Suzu, S.; Dosaka-Akita, H.; Jinushi, M. Cancer stem-like cells derived from chemoresistant tumors have a unique capacity to prime tumorigenic myeloid cells. Cancer Res. 2014, 74, 2698–2709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, A.; Wei, J.; Kong, L.Y.; Wang, Y.; Priebe, W.; Qiao, W.; Sawaya, R.; Heimberger, A.B. Glioma cancer stem cells induce immunosuppressive macrophages/microglia. Neuro-Oncol. 2010, 12, 1113–1125. [Google Scholar] [CrossRef] [PubMed]
- Jelski, W.; Mroczko, B. Biochemical diagnostics of pancreatic cancer—Present and future. Clin. Chim. Acta Int. J. Clin. Chem. 2019, 498, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Janakiram, N.B.; Mohammed, A.; Bryant, T.; Ritchie, R.; Stratton, N.; Jackson, L.; Lightfoot, S.; Benbrook, D.M.; Asch, A.S.; Lang, M.L.; et al. Loss of natural killer T cells promotes pancreatic cancer in LSL-Kras(G12D/+) mice. Immunology 2017, 152, 36–51. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.; Zhang, Z.; Yao, H.; Shen, L. Tim-4 promotes the growth of colorectal cancer by activating angiogenesis and recruiting tumor-associated macrophages via the PI3K/AKT/mTOR signaling pathway. Cancer Lett. 2018, 436, 119–128. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, Y.; Peña, M.M.; Pirisi, L.; Creek, K.E. Six1 promotes colorectal cancer growth and metastasis by stimulating angiogenesis and recruiting tumor-associated macrophages. Carcinogenesis 2017, 38, 281–292. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Ke, S.Q.; Huang, Z.; Flavahan, W.; Fang, X.; Paul, J.; Wu, L.; Sloan, A.E.; McLendon, R.E.; Li, X.; et al. Periostin secreted by glioblastoma stem cells recruits M2 tumour-associated macrophages and promotes malignant growth. Nat. Cell Biol. 2015, 17, 170–182. [Google Scholar] [CrossRef] [Green Version]
- Batool, A.; Wang, Y.Q.; Hao, X.X.; Chen, S.R.; Liu, Y.X. A miR-125b/CSF1-CX3CL1/tumor-associated macrophage recruitment axis controls testicular germ cell tumor growth. Cell Death Dis. 2018, 9, 962. [Google Scholar] [CrossRef]
- Sa, J.K.; Kim, S.H.; Lee, J.K.; Cho, H.J.; Shin, Y.J.; Shin, H.; Koo, H.; Kim, D.; Lee, M.; Kang, W.; et al. Identification of genomic and molecular traits that present therapeutic vulnerability to HGF-targeted therapy in glioblastoma. Neuro-Oncol. 2019, 21, 222–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otvos, B.; Silver, D.J.; Mulkearns-Hubert, E.E.; Alvarado, A.G.; Turaga, S.M.; Sorensen, M.D.; Rayman, P.; Flavahan, W.A.; Hale, J.S.; Stoltz, K.; et al. Cancer Stem Cell-Secreted Macrophage Migration Inhibitory Factor Stimulates Myeloid Derived Suppressor Cell Function and Facilitates Glioblastoma Immune Evasion. Stem Cells 2016, 34, 2026–2039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Zhao, Q.; Wang, J.; Tian, X.; Wang, J.; Xia, X.; Ott, M.; Rao, G.; Heimberger, A.B.; Li, S. FGL2-wired macrophages secrete CXCL7 to regulate the stem-like functionality of glioma cells. Cancer Lett. 2021, 506, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Bauer, D.; Mazzio, E.; Soliman, K.F.A. Whole Transcriptomic Analysis of Apigenin on TNFα Immuno-activated MDA-MB-231 Breast Cancer Cells. Cancer Genom. Proteom. 2019, 16, 421–431. [Google Scholar] [CrossRef] [Green Version]
- Fang, W.B.; Yao, M.; Brummer, G.; Acevedo, D.; Alhakamy, N.; Berkland, C.; Cheng, N. Targeted gene silencing of CCL2 inhibits triple negative breast cancer progression by blocking cancer stem cell renewal and M2 macrophage recruitment. Oncotarget 2016, 7, 49349–49367. [Google Scholar] [CrossRef]
- Zhang, J.; Patel, L.; Pienta, K.J. CC chemokine ligand 2 (CCL2) promotes prostate cancer tumorigenesis and metastasis. Cytokine Growth Factor Rev. 2010, 21, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, X.; Wang, Y.; Li, S.; Wang, X. Krüppel like factor 6 splice variant 1 (KLF6-SV1) overexpression recruits macrophages to participate in lung cancer metastasis by up-regulating TWIST1. Cancer Biol. Ther. 2019, 20, 680–691. [Google Scholar] [CrossRef]
- Aldinucci, D.; Lorenzon, D.; Cattaruzza, L.; Pinto, A.; Gloghini, A.; Carbone, A.; Colombatti, A. Expression of CCR5 receptors on Reed-Sternberg cells and Hodgkin lymphoma cell lines: Involvement of CCL5/Rantes in tumor cell growth and microenvironmental interactions. Int. J. Cancer 2008, 122, 769–776. [Google Scholar] [CrossRef]
- Kodama, T.; Koma, Y.I.; Arai, N.; Kido, A.; Urakawa, N.; Nishio, M.; Shigeoka, M.; Yokozaki, H. CCL3-CCR5 axis contributes to progression of esophageal squamous cell carcinoma by promoting cell migration and invasion via Akt and ERK pathways. Lab. Investig. J. Tech. Methods Pathol. 2020, 100, 1140–1157. [Google Scholar] [CrossRef]
- Gutmann, D.H.; Kettenmann, H. Microglia/Brain Macrophages as Central Drivers of Brain Tumor Pathobiology. Neuron 2019, 104, 442–449. [Google Scholar] [CrossRef]
- Shi, H.; Sun, Y.; Ruan, H.; Ji, C.; Zhang, J.; Wu, P.; Li, L.; Huang, C.; Jia, Y.; Zhang, X.; et al. 3,3’-Diindolylmethane Promotes Gastric Cancer Progression via β-TrCP-Mediated NF-κB Activation in Gastric Cancer-Derived MSCs. Front. Oncol. 2021, 11, 603533. [Google Scholar] [CrossRef] [PubMed]
- Özcan, Y.; Çağlar, F.; Celik, S.; Demir, A.B.; Erçetin, A.P.; Altun, Z.; Aktas, S. The role of cancer stem cells in immunotherapy for bladder cancer: An in vitro study. Urol. Oncol. 2020, 38, 476–487. [Google Scholar] [CrossRef] [PubMed]
- Pavon, L.F.; Sibov, T.T.; de Souza, A.V.; da Cruz, E.F.; Malheiros, S.M.F.; Cabral, F.R.; de Souza, J.G.; Boufleur, P.; de Oliveira, D.M.; de Toledo, S.R.C.; et al. Tropism of mesenchymal stem cell toward CD133(+) stem cell of glioblastoma in vitro and promote tumor proliferation in vivo. Stem Cell Res. Ther. 2018, 9, 310. [Google Scholar] [CrossRef] [PubMed]
- Wani, N.; Nasser, M.W.; Ahirwar, D.K.; Zhao, H.; Miao, Z.; Shilo, K.; Ganju, R.K. C-X-C motif chemokine 12/C-X-C chemokine receptor type 7 signaling regulates breast cancer growth and metastasis by modulating the tumor microenvironment. Breast Cancer Res. BCR 2014, 16, R54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Boeck, A.; Ahn, B.Y.; D’Mello, C.; Lun, X.; Menon, S.V.; Alshehri, M.M.; Szulzewsky, F.; Shen, Y.; Khan, L.; Dang, N.H.; et al. Glioma-derived IL-33 orchestrates an inflammatory brain tumor microenvironment that accelerates glioma progression. Nat. Commun. 2020, 11, 4997. [Google Scholar] [CrossRef]
- Liu, M.; Sun, X.; Shi, S. MORC2 Enhances Tumor Growth by Promoting Angiogenesis and Tumor-Associated Macrophage Recruitment via Wnt/β-Catenin in Lung Cancer. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 51, 1679–1694. [Google Scholar] [CrossRef]
- Liu, Y.; Li, X.; Zhang, Y.; Wang, H.; Rong, X.; Peng, J.; He, L.; Peng, Y. An miR-340-5p-macrophage feedback loop modulates the progression and tumor microenvironment of glioblastoma multiforme. Oncogene 2019, 38, 7399–7415. [Google Scholar] [CrossRef]
- Hu, J.; Jo, M.; Eastman, B.M.; Gilder, A.S.; Bui, J.D.; Gonias, S.L. uPAR induces expression of transforming growth factor β and interleukin-4 in cancer cells to promote tumor-permissive conditioning of macrophages. Am. J. Pathol. 2014, 184, 3384–3393. [Google Scholar] [CrossRef] [Green Version]
- Dzaye, O.; Hu, F.; Derkow, K.; Haage, V.; Euskirchen, P.; Harms, C.; Lehnardt, S.; Synowitz, M.; Wolf, S.A.; Kettenmann, H. Glioma Stem Cells but Not Bulk Glioma Cells Upregulate IL-6 Secretion in Microglia/Brain Macrophages via Toll-like Receptor 4 Signaling. J. Neuropathol. Exp. Neurol. 2016, 75, 429–440. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Yang, P.; Wang, J.; Zhang, J.; Ma, Q.; Jiang, Y.; Wu, Y.; Han, T.; Xiang, D. HLF regulates ferroptosis, development and chemoresistance of triple-negative breast cancer by activating tumor cell-macrophage crosstalk. J. Hematol. Oncol. 2022, 15, 2. [Google Scholar] [CrossRef]
- Weng, Y.S.; Tseng, H.Y.; Chen, Y.A.; Shen, P.C.; Al Haq, A.T.; Chen, L.M.; Tung, Y.C.; Hsu, H.L. MCT-1/miR-34a/IL-6/IL-6R signaling axis promotes EMT progression, cancer stemness and M2 macrophage polarization in triple-negative breast cancer. Mol. Cancer 2019, 18, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, Y.; Cui, Y.; Li, X.; Cao, X.; Chen, A.; Xu, C.; Cao, J.; Luo, X. Co-culture of ovarian cancer stem-like cells with macrophages induced SKOV3 cells stemness via IL-8/STAT3 signaling. Biomed. Pharmacother. 2018, 103, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Zhang, P.; Liang, T.; Deng, S.; Chen, X.; Zhu, L. Ovarian cancer stem cells induce the M2 polarization of macrophages through the PPARγ and NF-κB pathways. Int. J. Mol. Med. 2015, 36, 449–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raggi, C.; Correnti, M.; Sica, A.; Andersen, J.B.; Cardinale, V.; Alvaro, D.; Chiorino, G.; Forti, E.; Glaser, S.; Alpini, G.; et al. Cholangiocarcinoma stem-like subset shapes tumor-initiating niche by educating associated macrophages. J. Hepatol. 2017, 66, 102–115. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.S.; Shiau, A.L.; Su, B.H.; Hsu, T.S.; Wang, C.T.; Su, Y.C.; Tsai, M.S.; Feng, Y.H.; Tseng, Y.L.; Yen, Y.T.; et al. Oct4 promotes M2 macrophage polarization through upregulation of macrophage colony-stimulating factor in lung cancer. J. Hematol. Oncol. 2020, 13, 62. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, Z.Z.; Aoshima, K.; Cai, W.L.; Sun, H.; Xu, T.; Zhang, Y.; An, Y.; Chen, J.F.; Chan, L.H.; et al. CECR2 drives breast cancer metastasis by promoting NF-κB signaling and macrophage-mediated immune suppression. Sci. Transl. Med. 2022, 14, eabf5473. [Google Scholar] [CrossRef]
- Sami, E.; Paul, B.T.; Koziol, J.A.; ElShamy, W.M. The Immunosuppressive Microenvironment in BRCA1-IRIS-Overexpressing TNBC Tumors Is Induced by Bidirectional Interaction with Tumor-Associated Macrophages. Cancer Res. 2020, 80, 1102–1117. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Wu, H.; Li, Y.; Zhang, R.; Kleeff, J.; Zhang, X.; Cui, M.; Liu, J.; Li, T.; Gao, J.; et al. Combined blockade of TGf-β1 and GM-CSF improves chemotherapeutic effects for pancreatic cancer by modulating tumor microenvironment. Cancer Immunol. Immunother. CII 2020, 69, 1477–1492. [Google Scholar] [CrossRef]
- Tao, W.; Chu, C.; Zhou, W.; Huang, Z.; Zhai, K.; Fang, X.; Huang, Q.; Zhang, A.; Wang, X.; Yu, X.; et al. Dual Role of WISP1 in maintaining glioma stem cells and tumor-supportive macrophages in glioblastoma. Nat. Commun. 2020, 11, 3015. [Google Scholar] [CrossRef]
- Tsukamoto, H.; Mishra, L.; Machida, K. Alcohol, TLR4-TGF-β antagonism, and liver cancer. Hepatol. Int. 2014, 8 (Suppl. S2), 408–412. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Cai, D.J.; Li, B. Ovarian cancer stem-like cells elicit the polarization of M2 macrophages. Mol. Med. Rep. 2015, 11, 4685–4693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Kim, S.S.; Choi, E.; Oh, Y.T.; Lin, W.; Kim, T.H.; Sa, J.K.; Hong, J.H.; Park, S.H.; Kwon, H.J.; et al. ARS2/MAGL signaling in glioblastoma stem cells promotes self-renewal and M2-like polarization of tumor-associated macrophages. Nat. Commun. 2020, 11, 2978. [Google Scholar] [CrossRef] [PubMed]
- Heiler, S.; Wang, Z.; Zöller, M. Pancreatic cancer stem cell markers and exosomes—The incentive push. World J. Gastroenterol. 2016, 22, 5971–6007. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Luo, G.; Zhang, K.; Cao, J.; Huang, C.; Jiang, T.; Liu, B.; Su, L.; Qiu, Z. Correction: Hypoxic Tumor-Derived Exosomal miR-301a Mediates M2 Macrophage Polarization via PTEN/PI3Kγ to Promote Pancreatic Cancer Metastasis. Cancer Res. 2020, 80, 922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabrusiewicz, K.; Li, X.; Wei, J.; Hashimoto, Y.; Marisetty, A.L.; Ott, M.; Wang, F.; Hawke, D.; Yu, J.; Healy, L.M.; et al. Glioblastoma stem cell-derived exosomes induce M2 macrophages and PD-L1 expression on human monocytes. Oncoimmunology 2018, 7, e1412909. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Song, T.N.; Wang, F.R.; Yin, C.; Li, Z.; Lin, J.P.; Meng, Y.Q.; Feng, H.M.; Jing, T. Tumor-derived exosomal HMGB1 promotes esophageal squamous cell carcinoma progression through inducing PD1(+) TAM expansion. Oncogenesis 2019, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.; Ma, T.; Lin, Y.; Lu, X.; Zhang, C.; Chen, S.; Jian, Z. IL-6/STAT3 pathway intermediates M1/M2 macrophage polarization during the development of hepatocellular carcinoma. J. Cell. Biochem. 2018, 119, 9419–9432. [Google Scholar] [CrossRef]
- Myers, K.V.; Amend, S.R.; Pienta, K.J. Targeting Tyro3, Axl and MerTK (TAM receptors): Implications for macrophages in the tumor microenvironment. Mol. Cancer 2019, 18, 94. [Google Scholar] [CrossRef] [Green Version]
- Gomez, K.E.; Wu, F.; Keysar, S.B.; Morton, J.J.; Miller, B.; Chimed, T.S.; Le, P.N.; Nieto, C.; Chowdhury, F.N.; Tyagi, A.; et al. Cancer Cell CD44 Mediates Macrophage/Monocyte-Driven Regulation of Head and Neck Cancer Stem Cells. Cancer Res. 2020, 80, 4185–4198. [Google Scholar] [CrossRef]
- Huang, R.; Wang, S.; Wang, N.; Zheng, Y.; Zhou, J.; Yang, B.; Wang, X.; Zhang, J.; Guo, L.; Wang, S.; et al. CCL5 derived from tumor-associated macrophages promotes prostate cancer stem cells and metastasis via activating β-catenin/STAT3 signaling. Cell Death Dis. 2020, 11, 234. [Google Scholar] [CrossRef]
- Jeong, M.; Wang, Y.Y.; Choi, J.Y.; Lim, M.C.; Choi, J.H. CC Chemokine Ligand 7 Derived from Cancer-Stimulated Macrophages Promotes Ovarian Cancer Cell Invasion. Cancers 2021, 13, 2745. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, L.; Dang, W.Q.; Cao, M.F.; Xiao, J.F.; Lv, S.Q.; Jiang, W.J.; Yao, X.H.; Lu, H.M.; Miao, J.Y.; et al. CCL8 secreted by tumor-associated macrophages promotes invasion and stemness of glioblastoma cells via ERK1/2 signaling. Lab. Investig. J. Tech. Methods Pathol. 2020, 100, 619–629. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Chen, D.; Wu, J.; Sui, C.; Deng, X.; Zhang, P.; Chen, Z.; Liu, D.; Yu, J.; Shi, J.; et al. Yes associated protein 1 promotes resistance to 5-fluorouracil in gastric cancer by regulating GLUT3-dependent glycometabolism reprogramming of tumor-associated macrophages. Arch. Biochem. Biophys. 2021, 702, 108838. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Li, X.; Chen, S.; Zeng, Q.; Zhao, Y.; Luo, F. Tumor-associated macrophage or chemokine ligand CCL17 positively regulates the tumorigenesis of hepatocellular carcinoma. Med. Oncol. 2016, 33, 17. [Google Scholar] [CrossRef]
- Yeung, O.W.; Lo, C.M.; Ling, C.C.; Qi, X.; Geng, W.; Li, C.X.; Ng, K.T.; Forbes, S.J.; Guan, X.Y.; Poon, R.T.; et al. Alternatively activated (M2) macrophages promote tumour growth and invasiveness in hepatocellular carcinoma. J. Hepatol. 2015, 62, 607–616. [Google Scholar] [CrossRef]
- Wang, S.; Wang, N.; Huang, X.; Yang, B.; Zheng, Y.; Zhang, J.; Wang, X.; Lin, Y.; Wang, Z. Baohuoside i suppresses breast cancer metastasis by downregulating the tumor-associated macrophages/C-X-C motif chemokine ligand 1 pathway. Phytomed. Int. J. Phytother. Phytopharmacol. 2020, 78, 153331. [Google Scholar] [CrossRef]
- Nie, G.; Cao, X.; Mao, Y.; Lv, Z.; Lv, M.; Wang, Y.; Wang, H.; Liu, C. Tumor-associated macrophages-mediated CXCL8 infiltration enhances breast cancer metastasis: Suppression by Danirixin. Int. Immunopharmacol. 2021, 95, 107153. [Google Scholar] [CrossRef]
- Feng, X.; Szulzewsky, F.; Yerevanian, A.; Chen, Z.; Heinzmann, D.; Rasmussen, R.D.; Alvarez-Garcia, V.; Kim, Y.; Wang, B.; Tamagno, I.; et al. Loss of CX3CR1 increases accumulation of inflammatory monocytes and promotes gliomagenesis. Oncotarget 2015, 6, 15077–15094. [Google Scholar] [CrossRef] [Green Version]
- Wan, S.; Zhao, E.; Kryczek, I.; Vatan, L.; Sadovskaya, A.; Ludema, G.; Simeone, D.M.; Zou, W.; Welling, T.H. Tumor-associated macrophages produce interleukin 6 and signal via STAT3 to promote expansion of human hepatocellular carcinoma stem cells. Gastroenterology 2014, 147, 1393–1404. [Google Scholar] [CrossRef] [Green Version]
- Zhou, N.; Zhang, Y.; Zhang, X.; Lei, Z.; Hu, R.; Li, H.; Mao, Y.; Wang, X.; Irwin, D.M.; Niu, G.; et al. Exposure of tumor-associated macrophages to apoptotic MCF-7 cells promotes breast cancer growth and metastasis. Int. J. Mol. Sci. 2015, 16, 11966–11982. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Dong, Y.; Li, Y.; Wang, D.; Liu, S.; Wang, D.; Gao, Q.; Ji, S.; Chen, X.; Lei, Q.; et al. IL-10 derived from M2 macrophage promotes cancer stemness via JAK1/STAT1/NF-κB/Notch1 pathway in non-small cell lung cancer. Int. J. Cancer 2019, 145, 1099–1110. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Seppänen, H.; Vainionpää, S.; Ye, Y.; Wang, S.; Mustonen, H.; Puolakkainen, P. IL10, IL11, IL18 are differently expressed in CD14+ TAMs and play different role in regulating the invasion of gastric cancer cells under hypoxia. Cytokine 2012, 59, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Torres, S.J.; Bourn, J.R.; Benight, N.M.; Hunt, B.G.; Lester, C.; Waltz, S.E. Macrophage-mediated RON signaling supports breast cancer growth and progression through modulation of IL-35. Oncogene 2022, 41, 321–333. [Google Scholar] [CrossRef]
- Fan, Q.M.; Jing, Y.Y.; Yu, G.F.; Kou, X.R.; Ye, F.; Gao, L.; Li, R.; Zhao, Q.D.; Yang, Y.; Lu, Z.H.; et al. Tumor-associated macrophages promote cancer stem cell-like properties via transforming growth factor-beta1-induced epithelial-mesenchymal transition in hepatocellular carcinoma. Cancer Lett. 2014, 352, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Ye, H.; Ren, X.; Zheng, S.; Zhou, Q.; Chen, C.; Lin, Q.; Li, G.; Wei, L.; Fu, Z.; et al. Macrophage-expressed CD51 promotes cancer stem cell properties via the TGF-β1/smad2/3 axis in pancreatic cancer. Cancer Lett. 2019, 459, 204–215. [Google Scholar] [CrossRef]
- Zhang, X.; Zhu, M.; Hong, Z.; Chen, C. Co-culturing polarized M2 Thp-1-derived macrophages enhance stemness of lung adenocarcinoma A549 cells. Ann. Transl. Med. 2021, 9, 709. [Google Scholar] [CrossRef]
- Liu, Q.; Hodge, J.; Wang, J.; Wang, Y.; Wang, L.; Singh, U.; Li, Y.; Yao, Y.; Wang, D.; Ai, W.; et al. Emodin reduces Breast Cancer Lung Metastasis by suppressing Macrophage-induced Breast Cancer Cell Epithelial-mesenchymal transition and Cancer Stem Cell formation. Theranostics 2020, 10, 8365–8381. [Google Scholar] [CrossRef]
- Ma, C.; Komohara, Y.; Ohnishi, K.; Shimoji, T.; Kuwahara, N.; Sakumura, Y.; Matsuishi, K.; Fujiwara, Y.; Motoshima, T.; Takahashi, W.; et al. Infiltration of tumor-associated macrophages is involved in CD44 expression in clear cell renal cell carcinoma. Cancer Sci. 2016, 107, 700–707. [Google Scholar] [CrossRef] [Green Version]
- Gulluoglu, S.; Tuysuz, E.C.; Sahin, M.; Yaltirik, C.K.; Kuskucu, A.; Ozkan, F.; Dalan, A.B.; Sahin, F.; Ture, U.; Bayrak, O.F. The role of TNF-α in chordoma progression and inflammatory pathways. Cell. Oncol. 2019, 42, 663–677. [Google Scholar] [CrossRef]
- Yang, J.; Liao, D.; Chen, C.; Liu, Y.; Chuang, T.H.; Xiang, R.; Markowitz, D.; Reisfeld, R.A.; Luo, Y. Tumor-associated macrophages regulate murine breast cancer stem cells through a novel paracrine EGFR/Stat3/Sox-2 signaling pathway. Stem Cells 2013, 31, 248–258. [Google Scholar] [CrossRef]
- Liguori, M.; Digifico, E.; Vacchini, A.; Avigni, R.; Colombo, F.S.; Borroni, E.M.; Farina, F.M.; Milanesi, S.; Castagna, A.; Mannarino, L.; et al. The soluble glycoprotein NMB (GPNMB) produced by macrophages induces cancer stemness and metastasis via CD44 and IL-33. Cell. Mol. Immunol. 2021, 18, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Yang, S.; Pu, X.; He, S.; Yang, Z.; Sheng, X.; Meng, X.; Chen, X.; Jin, L.; Chen, W.; et al. Tumor-associated macrophages increase the proportion of cancer stem cells in lymphoma by secreting pleiotrophin. Am. J. Transl. Res. 2019, 11, 6393–6402. [Google Scholar] [PubMed]
- Shi, Y.; Ping, Y.F.; Zhou, W.; He, Z.C.; Chen, C.; Bian, B.S.; Zhang, L.; Chen, L.; Lan, X.; Zhang, X.C.; et al. Tumour-associated macrophages secrete pleiotrophin to promote PTPRZ1 signalling in glioblastoma stem cells for tumour growth. Nat. Commun. 2017, 8, 15080. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Liu, C.; Chen, F.K.; Feng, Z.P.; Jia, L.; Liu, P.J.; Yang, Z.X.; Hou, F.; Deng, Z.Y. M2-like tumour-associated macrophage-secreted IGF promotes thyroid cancer stemness and metastasis by activating the PI3K/AKT/mTOR pathway. Mol. Med. Rep. 2021, 24, 1–10. [Google Scholar] [CrossRef]
- Okuda, H.; Kobayashi, A.; Xia, B.; Watabe, M.; Pai, S.K.; Hirota, S.; Xing, F.; Liu, W.; Pandey, P.R.; Fukuda, K.; et al. Hyaluronan synthase HAS2 promotes tumor progression in bone by stimulating the interaction of breast cancer stem-like cells with macrophages and stromal cells. Cancer Res. 2012, 72, 537–547. [Google Scholar] [CrossRef] [Green Version]
- Sainz, B., Jr.; Alcala, S.; Garcia, E.; Sanchez-Ripoll, Y.; Azevedo, M.M.; Cioffi, M.; Tatari, M.; Miranda-Lorenzo, I.; Hidalgo, M.; Gomez-Lopez, G.; et al. Microenvironmental hCAP-18/LL-37 promotes pancreatic ductal adenocarcinoma by activating its cancer stem cell compartment. Gut 2015, 64, 1921–1935. [Google Scholar] [CrossRef] [Green Version]
- Wei, R.; Zhu, W.W.; Yu, G.Y.; Wang, X.; Gao, C.; Zhou, X.; Lin, Z.F.; Shao, W.Q.; Wang, S.H.; Lu, M.; et al. S100 calcium-binding protein A9 from tumor-associated macrophage enhances cancer stem cell-like properties of hepatocellular carcinoma. Int. J. Cancer 2021, 148, 1233–1244. [Google Scholar] [CrossRef]
- Xu, H.Z.; Li, T.F.; Wang, C.; Ma, Y.; Liu, Y.; Zheng, M.Y.; Liu, Z.J.; Chen, J.B.; Li, K.; Sun, S.K.; et al. Synergy of nanodiamond-doxorubicin conjugates and PD-L1 blockade effectively turns tumor-associated macrophages against tumor cells. J. Nanobiotechnol. 2021, 19, 268. [Google Scholar] [CrossRef]
- Jinushi, M.; Chiba, S.; Yoshiyama, H.; Masutomi, K.; Kinoshita, I.; Dosaka-Akita, H.; Yagita, H.; Takaoka, A.; Tahara, H. Tumor-associated macrophages regulate tumorigenicity and anticancer drug responses of cancer stem/initiating cells. Proc. Natl. Acad. Sci. USA 2011, 108, 12425–12430. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Li, H.; Zhu, Z.; Mei, P.; Hu, W.; Xiong, X.; Tao, J. microRNA-21-5p from M2 macrophage-derived extracellular vesicles promotes the differentiation and activity of pancreatic cancer stem cells by mediating KLF3. Cell Biol. Toxicol. 2021. [Google Scholar] [CrossRef]
- You, Y.; Tian, Z.; Du, Z.; Wu, K.; Xu, G.; Dai, M.; Wang, Y.; Xiao, M. M1-like tumor-associated macrophages cascade a mesenchymal/stem-like phenotype of oral squamous cell carcinoma via the IL6/Stat3/THBS1 feedback loop. J. Exp. Clin. Cancer Res. CR 2022, 41, 10. [Google Scholar] [CrossRef] [PubMed]
- Tabu, K.; Liu, W.; Kosaku, A.; Terashima, K.; Murota, Y.; Aimaitijiang, A.; Nobuhisa, I.; Hide, T.; Taga, T. Glioma stem cell (GSC)-derived autoschizis-like products confer GSC niche properties involving M1-like tumor-associated macrophages. Stem Cells 2020, 38, 921–935. [Google Scholar] [CrossRef] [PubMed]
- Mine, N.; Yamamoto, S.; Saito, N.; Sato, T.; Sakakibara, K.; Kufe, D.W.; VonHoff, D.D.; Kawabe, T. CBP501 suppresses macrophage induced cancer stem cell like features and metastases. Oncotarget 2017, 8, 64015–64031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradhan, R.; Chatterjee, S.; Hembram, K.C.; Sethy, C.; Mandal, M.; Kundu, C.N. Nano formulated Resveratrol inhibits metastasis and angiogenesis by reducing inflammatory cytokines in oral cancer cells by targeting tumor associated macrophages. J. Nutr. Biochem. 2021, 92, 108624. [Google Scholar] [CrossRef] [PubMed]
- Sawa-Wejksza, K.; Kandefer-Szerszeń, M. Tumor-Associated Macrophages as Target for Antitumor Therapy. Arch. Immunol. Ther. Exp. 2018, 66, 97–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, Y.; Feng, W.; Cao, X.; Ren, K.; Quan, M.; Chen, A.; Xu, C.; Qiu, Y.; Cao, J.; Li, X.; et al. Genistein inhibits stemness of SKOV3 cells induced by macrophages co-cultured with ovarian cancer stem-like cells through IL-8/STAT3 axis. J. Exp. Clin. Cancer Res. CR 2019, 38, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hide, T.; Komohara, Y.; Miyasato, Y.; Nakamura, H.; Makino, K.; Takeya, M.; Kuratsu, J.I.; Mukasa, A.; Yano, S. Oligodendrocyte Progenitor Cells and Macrophages/Microglia Produce Glioma Stem Cell Niches at the Tumor Border. EBioMedicine 2018, 30, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Fang, M.; Li, Y.; Huang, K.; Qi, S.; Zhang, J.; Zgodzinski, W.; Majewski, M.; Wallner, G.; Gozdz, S.; Macek, P.; et al. IL33 Promotes Colon Cancer Cell Stemness via JNK Activation and Macrophage Recruitment. Cancer Res. 2017, 77, 2735–2745. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Liu, Y.; Kong, X.; Wu, R.; Peng, Q.; Zhang, Y.; Zhou, L.; Duan, L. Fusobacterium nucleatum Facilitates M2 Macrophage Polarization and Colorectal Carcinoma Progression by Activating TLR4/NF-κB/S100A9 Cascade. Front. Immunol. 2021, 12, 658681. [Google Scholar] [CrossRef]
- Wang, Y.; Gong, X.; Li, J.; Wang, H.; Xu, X.; Wu, Y.; Wang, J.; Wang, S.; Li, Y.; Zhang, Z. M2 macrophage microvesicle-inspired nanovehicles improve accessibility to cancer cells and cancer stem cells in tumors. J. Nanobiotechnol. 2021, 19, 397. [Google Scholar] [CrossRef]
- Kaneda, M.M.; Cappello, P.; Nguyen, A.V.; Ralainirina, N.; Hardamon, C.R.; Foubert, P.; Schmid, M.C.; Sun, P.; Mose, E.; Bouvet, M.; et al. Macrophage PI3Kγ Drives Pancreatic Ductal Adenocarcinoma Progression. Cancer Discov. 2016, 6, 870–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid, M.C.; Khan, S.Q.; Kaneda, M.M.; Pathria, P.; Shepard, R.; Louis, T.L.; Anand, S.; Woo, G.; Leem, C.; Faridi, M.H.; et al. Integrin CD11b activation drives anti-tumor innate immunity. Nat. Commun. 2018, 9, 5379. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, G.; Pötner, C.; Aust, G.; Schierle, K.; Dietel, C.; Benzing, C.; Krenzien, F.; Bartels, M.; Eichfeld, U.; Schmelzle, M.; et al. TIE2-expressing monocytes and M2-polarized macrophages impact survival and correlate with angiogenesis in adenocarcinoma of the pancreas. Oncotarget 2018, 9, 29715–29726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.J.; Wang, X.H.; Gao, S.T.; Chen, C.; Xu, X.Y.; Sun, Q.; Zhou, Z.H.; Wu, G.Z.; Yu, Q.; Xu, G.; et al. Tumor-associated macrophages correlate with phenomenon of epithelial-mesenchymal transition and contribute to poor prognosis in triple-negative breast cancer patients. J. Surg. Res. 2018, 222, 93–101. [Google Scholar] [CrossRef]
- Sørensen, M.D.; Dahlrot, R.H.; Boldt, H.B.; Hansen, S.; Kristensen, B.W. Tumour-associated microglia/macrophages predict poor prognosis in high-grade gliomas and correlate with an aggressive tumour subtype. Neuropathol. Appl. Neurobiol. 2018, 44, 185–206. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Li, P.; Wang, L.; Xia, Z.; Huang, H.; Lu, Y.; Li, Z. High numbers of CD68+ tumor-associated macrophages correlate with poor prognosis in extranodal NK/T-cell lymphoma, nasal type. Ann. Hematol. 2015, 94, 1535–1544. [Google Scholar] [CrossRef]
- Zhang, Y.; Cheng, S.; Zhang, M.; Zhen, L.; Pang, D.; Zhang, Q.; Li, Z. High-infiltration of tumor-associated macrophages predicts unfavorable clinical outcome for node-negative breast cancer. PLoS ONE 2013, 8, e76147. [Google Scholar] [CrossRef]
- Kaneda, M.M.; Messer, K.S.; Ralainirina, N.; Li, H.; Leem, C.J.; Gorjestani, S.; Woo, G.; Nguyen, A.V.; Figueiredo, C.C.; Foubert, P.; et al. PI3Kγ is a molecular switch that controls immune suppression. Nature 2016, 539, 437–442. [Google Scholar] [CrossRef] [Green Version]
- DeNardo, D.G.; Ruffell, B. Macrophages as regulators of tumour immunity and immunotherapy. Nat. Rev. Immunol. 2019, 19, 369–382. [Google Scholar] [CrossRef]
- Mantovani, A.; Marchesi, F.; Malesci, A.; Laghi, L.; Allavena, P. Tumour-associated macrophages as treatment targets in oncology. Nat. Rev. Clin. Oncol. 2017, 14, 399–416. [Google Scholar] [CrossRef]
- Cannarile, M.A.; Weisser, M.; Jacob, W.; Jegg, A.M.; Ries, C.H.; Rüttinger, D. Colony-stimulating factor 1 receptor (CSF1R) inhibitors in cancer therapy. J. Immunother. Cancer 2017, 5, 53. [Google Scholar] [CrossRef] [PubMed]
- Pyonteck, S.M.; Akkari, L.; Schuhmacher, A.J.; Bowman, R.L.; Sevenich, L.; Quail, D.F.; Olson, O.C.; Quick, M.L.; Huse, J.T.; Teijeiro, V.; et al. CSF-1R inhibition alters macrophage polarization and blocks glioma progression. Nat. Med. 2013, 19, 1264–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Knolhoff, B.L.; Meyer, M.A.; Nywening, T.M.; West, B.L.; Luo, J.; Wang-Gillam, A.; Goedegebuure, S.P.; Linehan, D.C.; DeNardo, D.G. CSF1/CSF1R blockade reprograms tumor-infiltrating macrophages and improves response to T-cell checkpoint immunotherapy in pancreatic cancer models. Cancer Res. 2014, 74, 5057–5069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Lu, Y.; Li, R.; Jiang, Y.; Zheng, Y.; Qian, J.; Bi, E.; Zheng, C.; Hou, J.; Wang, S.; et al. Therapeutic effects of CSF1R-blocking antibodies in multiple myeloma. Leukemia 2018, 32, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Ruffell, B.; Chang-Strachan, D.; Chan, V.; Rosenbusch, A.; Ho, C.M.; Pryer, N.; Daniel, D.; Hwang, E.S.; Rugo, H.S.; Coussens, L.M. Macrophage IL-10 blocks CD8+ T cell-dependent responses to chemotherapy by suppressing IL-12 expression in intratumoral dendritic cells. Cancer Cell 2014, 26, 623–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ries, C.H.; Cannarile, M.A.; Hoves, S.; Benz, J.; Wartha, K.; Runza, V.; Rey-Giraud, F.; Pradel, L.P.; Feuerhake, F.; Klaman, I.; et al. Targeting tumor-associated macrophages with anti-CSF-1R antibody reveals a strategy for cancer therapy. Cancer Cell 2014, 25, 846–859. [Google Scholar] [CrossRef] [Green Version]
- Tap, W.D.; Gelderblom, H.; Palmerini, E.; Desai, J.; Bauer, S.; Blay, J.Y.; Alcindor, T.; Ganjoo, K.; Martín-Broto, J.; Ryan, C.W.; et al. Pexidartinib versus placebo for advanced tenosynovial giant cell tumour (ENLIVEN): A randomised phase 3 trial. Lancet 2019, 394, 478–487. [Google Scholar] [CrossRef]
- Yan, D.; Kowal, J.; Akkari, L.; Schuhmacher, A.J.; Huse, J.T.; West, B.L.; Joyce, J.A. Inhibition of colony stimulating factor-1 receptor abrogates microenvironment-mediated therapeutic resistance in gliomas. Oncogene 2017, 36, 6049–6058. [Google Scholar] [CrossRef]
- Coniglio, S.J.; Eugenin, E.; Dobrenis, K.; Stanley, E.R.; West, B.L.; Symons, M.H.; Segall, J.E. Microglial stimulation of glioblastoma invasion involves epidermal growth factor receptor (EGFR) and colony stimulating factor 1 receptor (CSF-1R) signaling. Mol. Med. 2012, 18, 519–527. [Google Scholar] [CrossRef]
- Sakamoto, C.; Kohara, H.; Inoue, H.; Narusawa, M.; Ogawa, Y.; Hirose-Yotsuya, L.; Miyamoto, S.; Matsumura, Y.; Yamada, K.; Takahashi, A.; et al. Therapeutic vaccination based on side population cells transduced by the granulocyte-macrophage colony-stimulating factor gene elicits potent antitumor immunity. Cancer Gene Ther. 2017, 24, 165–174. [Google Scholar] [CrossRef]
- Wang, C.Y.; Hua, R.; Liu, L.; Zhan, X.; Chen, S.; Quan, S.; Chu, Q.J.; Zhu, Y.T. Immunotherapy against metastatic bladder cancer by combined administration of granulocyte macrophage-colony stimulating factor and interleukin-2 surface modified MB49 bladder cancer stem cells vaccine. Cancer Med. 2017, 6, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.Y.; Yuzhalin, A.E.; Gordon-Weeks, A.N.; Muschel, R.J. Targeting the CCL2-CCR2 signaling axis in cancer metastasis. Oncotarget 2016, 7, 28697–28710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sainz, B., Jr.; Carron, E.; Vallespinós, M.; Machado, H.L. Cancer Stem Cells and Macrophages: Implications in Tumor Biology and Therapeutic Strategies. Med. Inflamm. 2016, 2016, 9012369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalbasi, A.; Komar, C.; Tooker, G.M.; Liu, M.; Lee, J.W.; Gladney, W.L.; Ben-Josef, E.; Beatty, G.L. Tumor-Derived CCL2 Mediates Resistance to Radiotherapy in Pancreatic Ductal Adenocarcinoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2017, 23, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Loberg, R.D.; Ying, C.; Craig, M.; Yan, L.; Snyder, L.A.; Pienta, K.J. CCL2 as an important mediator of prostate cancer growth in vivo through the regulation of macrophage infiltration. Neoplasia 2007, 9, 556–562. [Google Scholar] [CrossRef] [Green Version]
- Loberg, R.D.; Ying, C.; Craig, M.; Day, L.L.; Sargent, E.; Neeley, C.; Wojno, K.; Snyder, L.A.; Yan, L.; Pienta, K.J. Targeting CCL2 with systemic delivery of neutralizing antibodies induces prostate cancer tumor regression in vivo. Cancer Res. 2007, 67, 9417–9424. [Google Scholar] [CrossRef] [Green Version]
- Sanford, D.E.; Belt, B.A.; Panni, R.Z.; Mayer, A.; Deshpande, A.D.; Carpenter, D.; Mitchem, J.B.; Plambeck-Suess, S.M.; Worley, L.A.; Goetz, B.D.; et al. Inflammatory monocyte mobilization decreases patient survival in pancreatic cancer: A role for targeting the CCL2/CCR2 axis. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2013, 19, 3404–3415. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Yue, D.L.; Wang, D.; Chen, X.F.; Yin, X.Y.; Wang, Y.P.; Yang, L.; Zhang, Y. [Aspirin inhibits cell stemness of esophageal cancer by downregulation of chemokine CCL2]. Zhonghua Zhong Liu Za Zhi [Chin. J. Oncol.] 2018, 40, 744–749. [Google Scholar] [CrossRef]
- Izumi, K.; Fang, L.Y.; Mizokami, A.; Namiki, M.; Li, L.; Lin, W.J.; Chang, C. Targeting the androgen receptor with siRNA promotes prostate cancer metastasis through enhanced macrophage recruitment via CCL2/CCR2-induced STAT3 activation. EMBO Mol. Med. 2013, 5, 1383–1401. [Google Scholar] [CrossRef]
- Udartseva, O.O.; Zhidkova, O.V.; Ezdakova, M.I.; Ogneva, I.V.; Andreeva, E.R.; Buravkova, L.B.; Gollnick, S.O. Low-dose photodynamic therapy promotes angiogenic potential and increases immunogenicity of human mesenchymal stromal cells. J. Photochem. Photobiol. B Biol. 2019, 199, 111596. [Google Scholar] [CrossRef]
- Eckert, F.; Schilbach, K.; Klumpp, L.; Bardoscia, L.; Sezgin, E.C.; Schwab, M.; Zips, D.; Huber, S.M. Potential Role of CXCR4 Targeting in the Context of Radiotherapy and Immunotherapy of Cancer. Front. Immunol. 2018, 9, 3018. [Google Scholar] [CrossRef] [PubMed]
- Burger, J.A.; Peled, A. CXCR4 antagonists: Targeting the microenvironment in leukemia and other cancers. Leukemia 2009, 23, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Grande, F.; Giancotti, G.; Ioele, G.; Occhiuzzi, M.A.; Garofalo, A. An update on small molecules targeting CXCR4 as starting points for the development of anti-cancer therapeutics. Eur. J. Med. Chem. 2017, 139, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Taromi, S.; Kayser, G.; Catusse, J.; von Elverfeldt, D.; Reichardt, W.; Braun, F.; Weber, W.A.; Zeiser, R.; Burger, M. CXCR4 antagonists suppress small cell lung cancer progression. Oncotarget 2016, 7, 85185–85195. [Google Scholar] [CrossRef] [Green Version]
- Bertolini, G.; Cancila, V.; Milione, M.; Lo Russo, G.; Fortunato, O.; Zaffaroni, N.; Tortoreto, M.; Centonze, G.; Chiodoni, C.; Facchinetti, F.; et al. A novel CXCR4 antagonist counteracts paradoxical generation of cisplatin-induced pro-metastatic niches in lung cancer. Mol. Ther. J. Am. Soc. Gene Ther. 2021, 29, 2963–2978. [Google Scholar] [CrossRef]
- Hao, Q.; Vadgama, J.V.; Wang, P. CCL2/CCR2 signaling in cancer pathogenesis. Cell Commun. Signal. 2020, 18, 82. [Google Scholar] [CrossRef]
- Laparidou, M.; Schlickenrieder, A.; Thoma, T.; Lengyel, K.; Schusser, B. Blocking of the CXCR4-CXCL12 Interaction Inhibits the Migration of Chicken B Cells into the Bursa of Fabricius. Front. Immunol. 2019, 10, 3057. [Google Scholar] [CrossRef]
- Poh, A.R.; Ernst, M. Targeting Macrophages in Cancer: From Bench to Bedside. Front. Oncol. 2018, 8, 49. [Google Scholar] [CrossRef] [Green Version]
- Linger, R.M.; Cohen, R.A.; Cummings, C.T.; Sather, S.; Migdall-Wilson, J.; Middleton, D.H.; Lu, X.; Barón, A.E.; Franklin, W.A.; Merrick, D.T.; et al. Mer or Axl receptor tyrosine kinase inhibition promotes apoptosis, blocks growth and enhances chemosensitivity of human non-small cell lung cancer. Oncogene 2013, 32, 3420–3431. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Wang, M.; Zhao, H.; Cui, W. Function of Axl receptor tyrosine kinase in non-small cell lung cancer. Oncol. Lett. 2018, 15, 2726–2734. [Google Scholar] [CrossRef] [Green Version]
- Xie, S.; Li, Y.; Li, X.; Wang, L.; Yang, N.; Wang, Y.; Wei, H. Mer receptor tyrosine kinase is frequently overexpressed in human non-small cell lung cancer, confirming resistance to erlotinib. Oncotarget 2015, 6, 9206–9219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, T.; Ben-Batalla, I.; Schultze, A.; Loges, S. Macrophage-tumor crosstalk: Role of TAMR tyrosine kinase receptors and of their ligands. Cell. Mol. Life Sci. 2012, 69, 1391–1414. [Google Scholar] [CrossRef] [PubMed]
- Cummings, C.T.; Zhang, W.; Davies, K.D.; Kirkpatrick, G.D.; Zhang, D.; DeRyckere, D.; Wang, X.; Frye, S.V.; Earp, H.S.; Graham, D.K. Small Molecule Inhibition of MERTK Is Efficacious in Non-Small Cell Lung Cancer Models Independent of Driver Oncogene Status. Mol. Cancer Ther. 2015, 14, 2014–2022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgoudaki, A.M.; Prokopec, K.E.; Boura, V.F.; Hellqvist, E.; Sohn, S.; Östling, J.; Dahan, R.; Harris, R.A.; Rantalainen, M.; Klevebring, D.; et al. Reprogramming Tumor-Associated Macrophages by Antibody Targeting Inhibits Cancer Progression and Metastasis. Cell Rep. 2016, 15, 2000–2011. [Google Scholar] [CrossRef] [Green Version]
- La Fleur, L.; Boura, V.F.; Alexeyenko, A.; Berglund, A.; Pontén, V.; Mattsson, J.S.M.; Djureinovic, D.; Persson, J.; Brunnström, H.; Isaksson, J.; et al. Expression of scavenger receptor MARCO defines a targetable tumor-associated macrophage subset in non-small cell lung cancer. Int. J. Cancer 2018, 143, 1741–1752. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.W.; Joyce, J.A. Alternative activation of tumor-associated macrophages by IL-4: Priming for protumoral functions. Cell Cycle 2010, 9, 4824–4835. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like Receptors and the Control of Immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef]
- McWhirter, S.M.; Jefferies, C.A. Nucleic Acid Sensors as Therapeutic Targets for Human Disease. Immunity 2020, 53, 78–97. [Google Scholar] [CrossRef]
- Adams, S.; Kozhaya, L.; Martiniuk, F.; Meng, T.C.; Chiriboga, L.; Liebes, L.; Hochman, T.; Shuman, N.; Axelrod, D.; Speyer, J.; et al. Topical TLR7 agonist imiquimod can induce immune-mediated rejection of skin metastases in patients with breast cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2012, 18, 6748–6757. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, H.; Barber, G.N. STING is an endoplasmic reticulum adaptor that facilitates innate immune signalling. Nature 2008, 455, 674–678. [Google Scholar] [CrossRef]
- Vanpouille-Box, C.; Hoffmann, J.A.; Galluzzi, L. Pharmacological modulation of nucleic acid sensors—Therapeutic potential and persisting obstacles. Nat. Rev. Drug Discov. 2019, 18, 845–867. [Google Scholar] [CrossRef] [PubMed]
- Tallerico, R.; Garofalo, C.; Carbone, E. A New Biological Feature of Natural Killer Cells: The Recognition of Solid Tumor-Derived Cancer Stem Cells. Front. Immunol. 2016, 7, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donini, C.; Rotolo, R.; Proment, A.; Aglietta, M.; Sangiolo, D.; Leuci, V. Cellular Immunotherapy Targeting Cancer Stem Cells: Preclinical Evidence and Clinical Perspective. Cells 2021, 10, 543. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, J.L.; Sotayo, A.; Ponichtera, H.E.; Castrillon, J.A.; Pourzia, A.L.; Schad, S.; Johnson, S.F.; Carrasco, R.D.; Lazo, S.; Bronson, R.T.; et al. Class IIa HDAC inhibition reduces breast tumours and metastases through anti-tumour macrophages. Nature 2017, 543, 428–432. [Google Scholar] [CrossRef]
- Neele, A.E.; de Winther, M.P.J. Repressing the repressor: Ezh2 mediates macrophage activation. J. Exp. Med. 2018, 215, 1269–1271. [Google Scholar] [CrossRef]
- Qi, B.; Yang, C.; Zhu, Z.; Chen, H. EZH2-Inhibited MicroRNA-454-3p Promotes M2 Macrophage Polarization in Glioma. Front. Cell Dev. Biol. 2020, 8, 574940. [Google Scholar] [CrossRef]
- Hamaidia, M.; Gazon, H.; Hoyos, C.; Hoffmann, G.B.; Louis, R.; Duysinx, B.; Willems, L. Inhibition of EZH2 methyltransferase decreases immunoediting of mesothelioma cells by autologous macrophages through a PD-1-dependent mechanism. JCI Insight 2019, 4, e128474. [Google Scholar] [CrossRef]
- Christofides, A.; Karantanos, T.; Bardhan, K.; Boussiotis, V.A. Epigenetic regulation of cancer biology and anti-tumor immunity by EZH2. Oncotarget 2016, 7, 85624–85640. [Google Scholar] [CrossRef] [Green Version]
- Yamagishi, M.; Uchimaru, K. Targeting EZH2 in cancer therapy. Curr. Opin. Oncol. 2017, 29, 375–381. [Google Scholar] [CrossRef]
- Eich, M.L.; Athar, M.; Ferguson, J.E., 3rd; Varambally, S. EZH2-Targeted Therapies in Cancer: Hype or a Reality. Cancer Res. 2020, 80, 5449–5458. [Google Scholar] [CrossRef]
- Straining, R.; Eighmy, W. Tazemetostat: EZH2 Inhibitor. J. Adv. Pract. Oncol. 2022, 13, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Italiano, A.; Soria, J.C.; Toulmonde, M.; Michot, J.M.; Lucchesi, C.; Varga, A.; Coindre, J.M.; Blakemore, S.J.; Clawson, A.; Suttle, B.; et al. Tazemetostat, an EZH2 inhibitor, in relapsed or refractory B-cell non-Hodgkin lymphoma and advanced solid tumours: A first-in-human, open-label, phase 1 study. Lancet. Oncol. 2018, 19, 649–659. [Google Scholar] [CrossRef]
- Ren, F.; Fan, M.; Mei, J.; Wu, Y.; Liu, C.; Pu, Q.; You, Z.; Liu, L. Interferon-γ and celecoxib inhibit lung-tumor growth through modulating M2/M1 macrophage ratio in the tumor microenvironment. Drug Des. Dev. Ther. 2014, 8, 1527–1538. [Google Scholar] [CrossRef] [Green Version]
- Hernández-SanMiguel, E.; Gargini, R.; Cejalvo, T.; Segura-Collar, B.; Núñez-Hervada, P.; Hortigüela, R.; Sepúlveda-Sánchez, J.M.; Hernández-Laín, A.; Pérez-Núñez, A.; Sanz, E.; et al. Ocoxin Modulates Cancer Stem Cells and M2 Macrophage Polarization in Glioblastoma. Oxidative Med. Cell. Longev. 2019, 2019, 9719730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Lee, J.S.; Kim, D.; Zhu, L. Exploration of Zinc Oxide Nanoparticles as a Multitarget and Multifunctional Anticancer Nanomedicine. ACS Appl. Mater. Interfaces 2017, 9, 39971–39984. [Google Scholar] [CrossRef]
- Chan, K.S.; Espinosa, I.; Chao, M.; Wong, D.; Ailles, L.; Diehn, M.; Gill, H.; Presti, J., Jr.; Chang, H.Y.; van de Rijn, M.; et al. Identification, molecular characterization, clinical prognosis, and therapeutic targeting of human bladder tumor-initiating cells. Proc. Natl. Acad. Sci. USA 2009, 106, 14016–14021. [Google Scholar] [CrossRef] [Green Version]
- Willingham, S.B.; Volkmer, J.P.; Gentles, A.J.; Sahoo, D.; Dalerba, P.; Mitra, S.S.; Wang, J.; Contreras-Trujillo, H.; Martin, R.; Cohen, J.D.; et al. The CD47-signal regulatory protein alpha (SIRPa) interaction is a therapeutic target for human solid tumors. Proc. Natl. Acad. Sci. USA 2012, 109, 6662–6667. [Google Scholar] [CrossRef] [Green Version]
- Grégoire, H.; Roncali, L.; Rousseau, A.; Chérel, M.; Delneste, Y.; Jeannin, P.; Hindré, F.; Garcion, E. Targeting Tumor Associated Macrophages to Overcome Conventional Treatment Resistance in Glioblastoma. Front. Pharmacol. 2020, 11, 368. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Shi, X.; Chen, C.; He, H.; Liu, L.; Wu, J.; Yan, H. High expression of CD47 in triple negative breast cancer is associated with epithelial-mesenchymal transition and poor prognosis. Oncol. Lett. 2019, 18, 3249–3255. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Li, B.; Chen, J.; Dang, J.; Chen, S.; Gunes, E.G.; Xu, B.; Tian, L.; Muend, S.; Raoof, M.; et al. Effect of cabazitaxel on macrophages improves CD47-targeted immunotherapy for triple-negative breast cancer. J. Immunother. Cancer 2021, 9, e002022. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Y.; Fan, J.; Chen, W.; Luan, J.; Mei, X.; Wang, S.; Li, Y.; Ye, L.; Li, S.; et al. Blocking CD47 efficiently potentiated therapeutic effects of anti-angiogenic therapy in non-small cell lung cancer. J. Immunother. Cancer 2019, 7, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Fan, J.; Wang, S.; Li, Y.; Wang, Y.; Li, S.; Luan, J.; Wang, Z.; Song, P.; Chen, Q.; et al. Targeting CD47 and Autophagy Elicited Enhanced Antitumor Effects in Non-Small Cell Lung Cancer. Cancer Immunol. Res. 2017, 5, 363–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, P.; Chi, X.; Cha, J.H.; Luo, S.; Yang, G.; Yan, X.; Yang, W.H. Potential of E3 Ubiquitin Ligases in Cancer Immunity: Opportunities and Challenges. Cells 2021, 10, 3309. [Google Scholar] [CrossRef] [PubMed]
- Chin, A.R.; Wang, S.E. Cytokines driving breast cancer stemness. Mol. Cell. Endocrinol. 2014, 382, 598–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agliano, A.; Calvo, A.; Box, C. The challenge of targeting cancer stem cells to halt metastasis. Semin. Cancer Biol. 2017, 44, 25–42. [Google Scholar] [CrossRef]
- Gao, Y.Y.; Tao, T.; Wu, D.; Zhuang, Z.; Lu, Y.; Wu, L.Y.; Liu, G.J.; Zhou, Y.; Zhang, D.D.; Wang, H.; et al. MFG-E8 attenuates inflammation in subarachnoid hemorrhage by driving microglial M2 polarization. Exp. Neurol. 2021, 336, 113532. [Google Scholar] [CrossRef]
- Shimagaki, T.; Yoshio, S.; Kawai, H.; Sakamoto, Y.; Doi, H.; Matsuda, M.; Mori, T.; Osawa, Y.; Fukai, M.; Yoshida, T.; et al. Serum milk fat globule-EGF factor 8 (MFG-E8) as a diagnostic and prognostic biomarker in patients with hepatocellular carcinoma. Sci. Rep. 2019, 9, 15788. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, W.; Sun, X.; Dang, R.; Zhou, R.; Bai, H.; Ben, J.; Zhu, X.; Zhang, Y.; Yang, Q.; et al. Class A1 scavenger receptor modulates glioma progression by regulating M2-like tumor-associated macrophage polarization. Oncotarget 2016, 7, 50099–50116. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Li, F.; Dong, B.; Zhang, J.; Rao, Y.; Cong, Y.; Mao, B.; Chen, X. Inhibition of STAT3 and ErbB2 suppresses tumor growth, enhances radiosensitivity, and induces mitochondria-dependent apoptosis in glioma cells. Int. J. Radiat. Oncol. Biol. Phys. 2010, 77, 1223–1231. [Google Scholar] [CrossRef]
- Singh, J.K.; Simões, B.M.; Clarke, R.B.; Bundred, N.J. Targeting IL-8 signalling to inhibit breast cancer stem cell activity. Expert Opin. Ther. Targets 2013, 17, 1235–1241. [Google Scholar] [CrossRef] [Green Version]
- Singh, J.K.; Farnie, G.; Bundred, N.J.; Simões, B.M.; Shergill, A.; Landberg, G.; Howell, S.J.; Clarke, R.B. Targeting CXCR1/2 significantly reduces breast cancer stem cell activity and increases the efficacy of inhibiting HER2 via HER2-dependent and -independent mechanisms. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2013, 19, 643–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, P.; Yang, Y.; Hosaka, K.; Zhang, Y.; Fischer, C.; Braun, H.; Liu, S.; Yu, G.; Liu, S.; Beyaert, R.; et al. Molecular mechanisms of IL-33-mediated stromal interactions in cancer metastasis. JCI Insight 2018, 3, e122375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, S.; Adhikari, N.; Banerjee, S.; Amin, S.A.; Jha, T. Matrix metalloproteinase-9 (MMP-9) and its inhibitors in cancer: A minireview. Eur. J. Med. Chem. 2020, 194, 112260. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Yang, N.; Pan, G.; Jin, B.; Wang, S.; Ji, W. Elevated IL-33 promotes expression of MMP2 and MMP9 via activating STAT3 in alveolar macrophages during LPS-induced acute lung injury. Cell. Mol. Biol. Lett. 2018, 23, 52. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, V.; Nataraj, R.; Thangaraj, G.S.; Karthikeyan, M.; Gnanasekaran, A.; Kaginelli, S.B.; Kuppanna, G.; Kallappa, C.G.; Basalingappa, K.M. Targeting Notch signalling pathway of cancer stem cells. Stem Cell Investig. 2018, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Sekulic, A.; Migden, M.R.; Oro, A.E.; Dirix, L.; Lewis, K.D.; Hainsworth, J.D.; Solomon, J.A.; Yoo, S.; Arron, S.T.; Friedlander, P.A.; et al. Efficacy and safety of vismodegib in advanced basal-cell carcinoma. N. Engl. J. Med. 2012, 366, 2171–2179. [Google Scholar] [CrossRef] [Green Version]
- Yao, Z.; Zhang, J.; Zhang, B.; Liang, G.; Chen, X.; Yao, F.; Xu, X.; Wu, H.; He, Q.; Ding, L.; et al. Imatinib prevents lung cancer metastasis by inhibiting M2-like polarization of macrophages. Pharmacol. Res. 2018, 133, 121–131. [Google Scholar] [CrossRef]
- Keir, H.R.; Richardson, H.; Fillmore, C.; Shoemark, A.; Lazaar, A.L.; Miller, B.E.; Tal-Singer, R.; Chalmers, J.D.; Mohan, D. CXCL-8-dependent and -independent neutrophil activation in COPD: Experiences from a pilot study of the CXCR2 antagonist danirixin. ERJ Open Res. 2020, 6, 00583-2020. [Google Scholar] [CrossRef]
- Busch-Petersen, J.; Carpenter, D.C.; Burman, M.; Foley, J.; Hunsberger, G.E.; Kilian, D.J.; Salmon, M.; Mayer, R.J.; Yonchuk, J.G.; Tal-Singer, R. Danirixin: A Reversible and Selective Antagonist of the CXC Chemokine Receptor 2. J. Pharmacol. Exp. Ther. 2017, 362, 338–346. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.R.; Chan, D.K.; Shank, J.J.; Griffith, K.A.; Fan, H.; Szulawski, R.; Yang, K.; Reynolds, R.K.; Johnston, C.; McLean, K.; et al. Phase II clinical trial of metformin as a cancer stem cell-targeting agent in ovarian cancer. JCI Insight 2020, 5, e133247. [Google Scholar] [CrossRef]
- Chen, B.; Cha, J.H.; Yan, M.; Cao, N.; Ye, P.; Yan, X.; Yang, W.H. ATXN7L3B promotes hepatocellular carcinoma stemness and is downregulated by metformin. Biochem. Biophys. Res. Commun. 2021, 573, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Lin, Y.; Xiong, X.; Wang, L.; Guo, Y.; Chen, Y.; Chen, S.; Wang, G.; Lin, P.; Chen, H.; et al. Low-Dose Metformin Reprograms the Tumor Immune Microenvironment in Human Esophageal Cancer: Results of a Phase II Clinical Trial. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2020, 26, 4921–4932. [Google Scholar] [CrossRef] [PubMed]
- Ejarque, M.; Ceperuelo-Mallafré, V.; Serena, C.; Pachón, G.; Núñez-Álvarez, Y.; Terrón-Puig, M.; Calvo, E.; Núñez-Roa, C.; Oliva-Olivera, W.; Tinahones, F.J.; et al. Survivin, a key player in cancer progression, increases in obesity and protects adipose tissue stem cells from apoptosis. Cell Death Dis. 2017, 8, e2802. [Google Scholar] [CrossRef] [PubMed]


| Targets | Drugs | Cancer Type | Trial Phase | ClinicalTrials.Gov ID |
|---|---|---|---|---|
| CSF-1/CSF-1R | CM082 | Small cell lung cancer | II | NCT03904719 |
| ABSK021 | Advanced solid tumor | I | NCT04192344 | |
| AMB-051 | Tenosynovial giant cell tumor | II | NCT04731675 | |
| SNDX-6532 | Unresectable intrahepatic cholangiocarcinoma | II | NCT04301778 | |
| Chiauranib | Triple-negative breast cancer | II | NCT05336721 | |
| Small cell lung cancer | III | NCT04830813 | ||
| NMS-03592088 | Acute myeloid leukemia Chronic myelomonocytic leukemia | I/II | NCT03922100 | |
| Pexidartinib | Tenosynovial giant cell tumor | III | NCT04488822 | |
| Q702 | Advanced solid cancer | I | NCT04648254 | |
| Surufatinib | Neuroendocrine tumor Non-hematologic malignancy | I/II | NCT05077384 | |
| Hepatocellular carcinoma | II | NCT05171439 | ||
| Refractory metastatic digestive system carcinoma Primary peritoneal cancer | II | NCT05030246 | ||
| TPX-0022 | Advanced metastatic solid tumor | I/II | NCT03993873 | |
| Vorolanib | Refractory thoracic tumor | I/II | NCT03583086 | |
| Metastatic renal cell cancer | III | NCT03095040 | ||
| CCL2/CCR2 | BMS-813160 | Locally advanced pancreatic ductal Adenocarcinoma Pancreatic ductal adenocarcinoma | I/II | NCT03767582 |
| CCL5/CCR5 | Maraviroc | Colorectal cancer metastatic | I | NCT04721301 |
| Leronlimab | Solid tumor | II | NCT04504942 | |
| POL6326 | Advanced breast cancer | I/II | NCT04826016 | |
| Metastatic breast cancer | III | NCT03786094 | ||
| Motixafortide | Pancreatic cancer | II | NCT04543071 | |
| CD40/CD40L | ABBV-368 | Advanced solid tumors | I | NCT04196283 |
| Triple-negative breast cancer Non small cell lung cancer Metastatic solid tumor | I | NCT03893955 | ||
| ABBV-927 | Advanced solid tumor Triple-negative breast cancer Non-small cell lung cancer Metastatic solid tumor | I | NCT03893955 | |
| Pancreatic cancer | II | NCT04807972 | ||
| CDX-1140 | Pancreatic cancer | II | NCT04536077 | |
| Solid tumor Diffuse large B-cell lymphoma Mantle cell lymphoma | I | NCT03329950 | ||
| Ovarian clear cell adenocarcinoma | II | NCT05231122 | ||
| Melanoma | I/II | NCT04364230 | ||
| Metastatic triple negative breast cancer | I | NCT05029999 | ||
| Malignant epithelial neoplasm | I | NCT04520711 | ||
| GEN-1042 | Malignant solid tumor Non-small cell lung cancer Colorectal cancer Melanoma Head and Neck squamous cell carcinoma Pancreatic ductal adenocarcinoma | I/II | NCT04083599 | |
| Mitazalimab | Metastatic pancreatic ductal adenocarcinoma | I/II | NCT04888312 | |
| NG-350A | Epithelial tumor Metastatic cancer | I | NCT05165433 | |
| Selicrelumab | Colorectal cancer | I/II | NCT03555149 | |
| Triple-negative breast cancer | I/II | NCT03424005 | ||
| Pancreatic adenocarcinoma | I/II | NCT03193190 | ||
| YH-003 | Advanced solid tumor | I/II | NCT04481009 | |
| Melanoma Pancreatic ductal adenocarcinoma | II | NCT05031494 | ||
| SL-172154 | Ovarian cancer Fallopian tube cancer Primary peritoneal carcinoma | I | NCT04406623 | |
| Cutaneous squamous cell carcinoma Head and Neck squamous cell carcinoma | I | NCT04502888 | ||
| Acute myeloid leukemia | I | NCT05275439 | ||
| BDB-001 | Solid tumor | II | NCT04819373 | |
| Pancreatic cancer Virus-associated tumor Non-small cell lung cancer Melanoma Bladder cancer Triple-negative breast cancer | II | NCT03915678 | ||
| BDC-1001 | HER2 positive solid tumor | I/II | NCT04278144 | |
| BNT-411 | Extensive-stage small cell lung cancer | I/II | NCT04101357 | |
| DSP-0509 | Neoplasm | I/II | NCT03416335 | |
| Imiquimod | Metastatic melanoma | I | NCT03276832 | |
| Oral cancer | I | NCT04883645 | ||
| Squamous cell carcinoma | I | NCT03370406 | ||
| Resiquimod | Locally advanced solid tumor Metastatic solid tumor | I/II | NCT04799054 | |
| Anaplastic astrocytoma | II | NCT01204684 | ||
| SBT-6050 | HER2 positive solid tumor | I | NCT04460456 | |
| TLR9 | AST008 | Advanced or Metastatic merkel cell carcinoma | I/II | NCT03684785 |
| CMP-001 | Head and Neck squamous cell carcinoma | II | NCT04633278 | |
| Metastatic melanoma | II | NCT04698187 | ||
| Merkel cell carcinoma | II | NCT04916002 | ||
| Malignant colorectal neoplasm | I | NCT03507699 | ||
| Advanced malignancy Non-small cell lung cancer Ovarian cancer Urothelial cancer Solid tumor | III | NCT05059522 | ||
| Tilsotolimod | Advanced solid tumor | I | NCT04196283 | |
| Malignant melanoma | II | NCT04126876 | ||
| SD-101 | Metastatic uveal melanoma in the Liver | I | NCT04935229 | |
| Hepatocellular carcinoma Intrahepatic cholangiocarcinoma | I/II | NCT05220722 | ||
| B-Cell Non-Hodgkin lymphoma | I | NCT03410901 | ||
| Metastatic pancreatic adenocarcinoma | I | NCT04050085 | ||
| TLR3 | Poly-ICLC | Hepatocellular carcinoma | I | NCT05281926 |
| Glioblastoma multiforme | I/II | NCT03665545 | ||
| Rintatolimod | Hematopoietic and lymphoid cell neoplasm | I/II | NCT04379518 | |
| Prostate adenocarcinoma | II | NCT03899987 | ||
| PI3Kγ signal pathway | Copanlisib | Non-small cell lung cancer | I | NCT04895579 |
| Diffuse large B-cell lymphoma | II | NCT04433182 | ||
| Refractory/Recurrent primary central system lymphoma | I/II | NCT03581942 | ||
| Follicular lymphoma Endometrial cancer | II | NCT04750941 | ||
| Mantle cell lymphoma | I/II | NCT03877055 | ||
| Head and Neck squamous cell carcinoma Hepatocellular carcinoma | I | NCT03735628 | ||
| Duvelisib | Non-Hodgkin lymphoma | I | NCT05065866 | |
| Unresectable melanoma | I/II | NCT04688658 | ||
| Recurrent diffuse large B-Cell lymphoma | I | NCT04890236 | ||
| Eganelisib | Bladder cancer Urothelial carcinoma | II | NCT03980041 | |
| Non-small cell lung cancer Melanoma Head and Neck squamous cell cancer Triple-negative breast cancer Adrenocortical carcinoma Mesothelioma High-circulating myeloid-derived suppressor cells | I | NCT02637531 | ||
| Breast cancer Renal cell carcinoma | II | NCT03961698 | ||
| Gedatolisib | HER2-positive breast cancer Metastatic breast cancer | II | NCT03698383 | |
| Triple-negative breast cancer | I/II | NCT03911973 | ||
| Squamous cell lung cancer Solid tumor Head and Neck cancer Pancreatic cancer | I | NCT03065062 | ||
| Tenalisib | Locally advanced breast cancer Metastatic breast cancer | II | NCT05021900 | |
| CD47/SIRPα pathway | ALX-148 | Microsatellite stable metastatic colorectal cancer | II | NCT05167409 |
| Aggressive B-Cell Non-Hodgkin lymphoma | I/II | NCT05025800 | ||
| Head and Neck squamous cell carcinoma | II | NCT04675294 | ||
| Acute myeloid leukemia | I/II | NCT04755244 | ||
| Gastroesophageal junction adenocarcinoma | II/III | NCT05002127 | ||
| Non-Hodgkin lymphoma | I | NCT03013218 | ||
| AO-176 | Multiple myeloma | I/II | NCT04445701 | |
| Solid tumor | I/II | NCT03834948 | ||
| HX-009 | Advanced solid tumor | II | NCT04886271 | |
| Relapsed/Refractory lymphoma | I/II | NCT05189093 | ||
| IBI-188 | Acute myeloid leukemia | I/II | NCT04485052 | |
| Lung adenocarcinoma Osteosarcoma | I | NCT04861948 | ||
| IBI-322 | Advanced malignant tumor Lymphoma | I | NCT04338659 | |
| Non-small cell lung cancer | II | NCT05296278 | ||
| Hematologic malignancy | I | NCT04795128 | ||
| Advanced malignancy | I | NCT04328831 | ||
| Myeloid tumor | I | NCT05148442 | ||
| IMC-002 | Advanced cancer | I | NCT05276310 | |
| Solid tumor Lymphoma | I | NCT04306224 | ||
| IMM-01 | Acute myeloid leukemia | I/II | NCT05140811 | |
| IMM-0306 | B-cell non-Hodgkin’s lymphoma | I | NCT04746131 | |
| Lemzoparlimab | Acute myeloid leukemia Myelodysplastic syndrome | I | NCT04912063 | |
| Multiple myeloma | I | NCT04895410 | ||
| Magrolimab | Malignant brain tumor | I | NCT05169944 | |
| Recurrent neuroblastoma | I | NCT04751383 | ||
| Solid tumor | II | NCT04827576 | ||
| Recurrent acute myeloid leukemia | I/II | NCT04435691 | ||
| Metastatic colorectal cancer | II | NCT05330429 | ||
| Multiple myeloma | II | NCT04892446 | ||
| Head and neck squamous cell carcinoma | II | NCT04854499 | ||
| Metastatic colorectal cancer | II | NCT05330429 | ||
| Multiple myeloma | II | NCT04892446 | ||
| Urothelial carcinoma Bladder cancer | I/II | NCT03869190 | ||
| RRx-001 | Central nervous system neoplasm | I | NCT04525014 | |
| Small cell lung carcinoma | III | NCT03699956 | ||
| Small cell carcinoma Non-small cell lung carcinoma Neuroendocrine tumor Ovarian epithelial cancer | II | NCT02489903 | ||
| TTI-622 | Multiple myeloma | I | NCT05139225 | |
| Fallopian tube cancer Primary peritoneal carcinoma | I/II | NCT05261490 | ||
| Acute myeloid leukemia Diffuse large B-Cell lymphoma | I | NCT03530683 | ||
| STING pathway | SNX-281 | Advanced solid tumor Advanced lymphoma | I | NCT04609579 |
| BMS-986301 | Advanced solid cancer | I | NCT03956680 | |
| GSK-3745417 | Neoplasm | I | NCT03843359 | |
| E−7766 | Lymphoma Advanced solid tumor | I | NCT04144140 | |
| SYNB-1891 | Metastatic solid neoplasm lymphoma | I | NCT04167137 | |
| TAK-676 | Non-small cell lung carcinoma Triple-negative breast neoplasm Head and Neck squamous cell carcinoma | I | NCT04879849 | |
| Solid neoplasm | I | NCT04420884 | ||
| MK-2118 | Lymphoma | I | NCT03249792 | |
| IMSA101 | Solid tumor | I/II | NCT04020185 | |
| CDK-002 | Advanced solid tumor | I/II | NCT04592484 | |
| MK-1454 | Head and neck squamous cell carcinoma | II | NCT04220866 | |
| NOX66 | Late-stage prostate cancer | I | NCT03307629 | |
| Metastatic soft-tissue sarcoma | I | NCT05100628 | ||
| Metastatic castration-resistant prostate cancer | I/II | NCT04957290 | ||
| SB-11285 | Melanoma Head and neck squamous cell carcinoma | I | NCT04096638 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, S.; Yang, G.; Ye, P.; Cao, N.; Chi, X.; Yang, W.-H.; Yan, X. Macrophages Are a Double-Edged Sword: Molecular Crosstalk between Tumor-Associated Macrophages and Cancer Stem Cells. Biomolecules 2022, 12, 850. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12060850
Luo S, Yang G, Ye P, Cao N, Chi X, Yang W-H, Yan X. Macrophages Are a Double-Edged Sword: Molecular Crosstalk between Tumor-Associated Macrophages and Cancer Stem Cells. Biomolecules. 2022; 12(6):850. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12060850
Chicago/Turabian StyleLuo, Shahang, Guanghui Yang, Peng Ye, Nengqi Cao, Xiaoxia Chi, Wen-Hao Yang, and Xiuwen Yan. 2022. "Macrophages Are a Double-Edged Sword: Molecular Crosstalk between Tumor-Associated Macrophages and Cancer Stem Cells" Biomolecules 12, no. 6: 850. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12060850