Characterization of Terpene Synthase from Tea Green Leafhopper Being Involved in Formation of Geraniol in Tea (Camellia sinensis) Leaves and Potential Effect of Geraniol on Insect-Derived Endobacteria


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Collection and Analysis of Geraniol
2.3. Extraction and Assay Activity of Geraniol Synthase from Tea Leaves
2.4. Extraction and Activity Assay of Geraniol Synthase from Empoasca (Matsumurasca) onukii
2.5. Empoasca (Matsumurasca) onukii Terpene Synthase Genes Cloning
2.6. Recombinant Expression of Empoasca (Matsumurasca) onukii Terpene Synthases in Escherichia Coli
2.7. Recombinant Expression of Empoasca (Matsumurasca) onukii Terpene Synthases in Insect Cell Sf9
2.8. Recombinant Empoasca (Matsumurasca) onukii Terpene Synthase Enzyme Assay
2.9. Extraction and Activity Assay of Geraniol Synthase from Other Insects
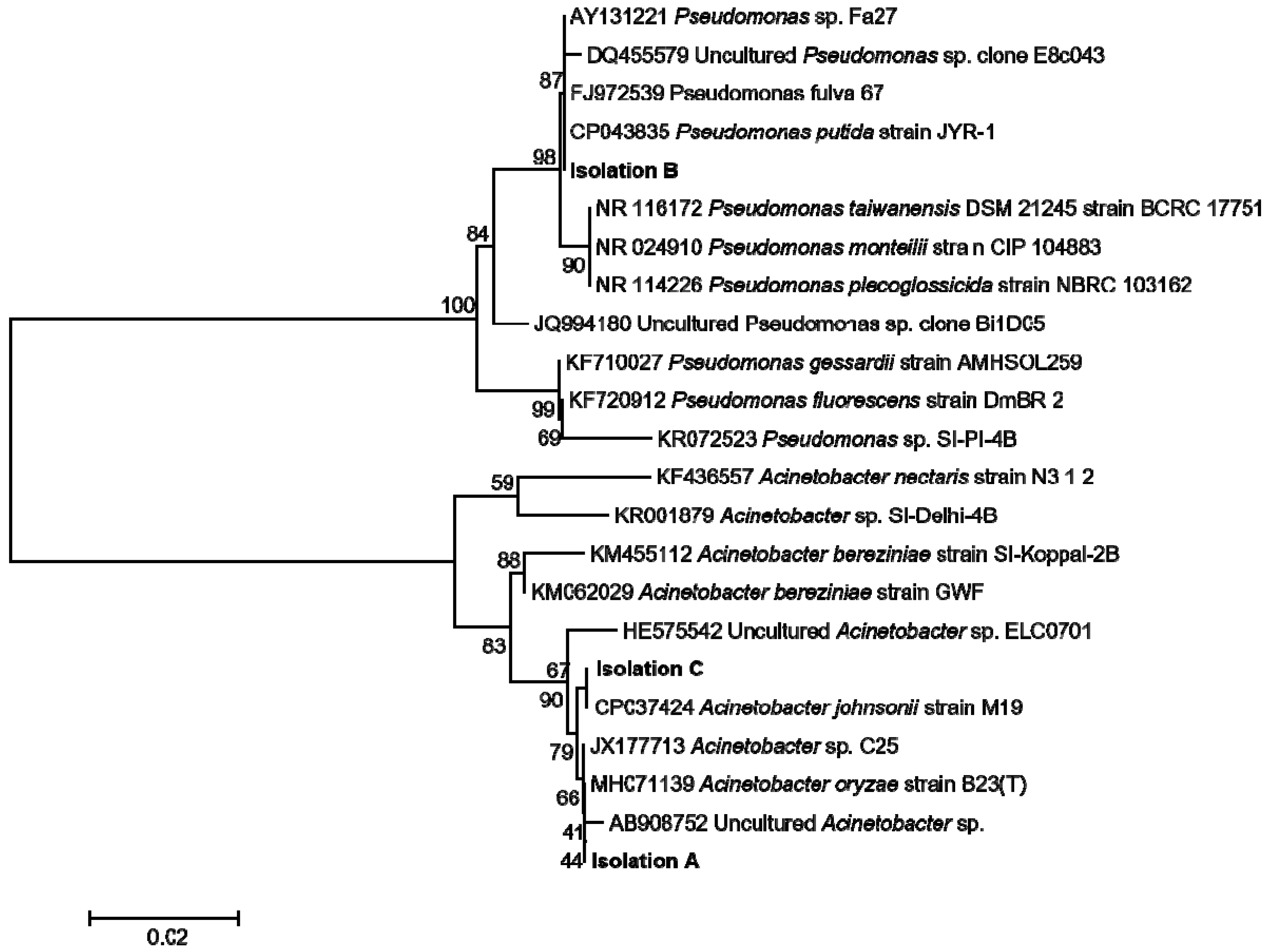
2.10. Empoasca (Matsumurasca) onukii Endobateria Isolation, Identification, and Antibacterial Activity Assay of Geraniol
2.11. Statistical Analyses
3. Results
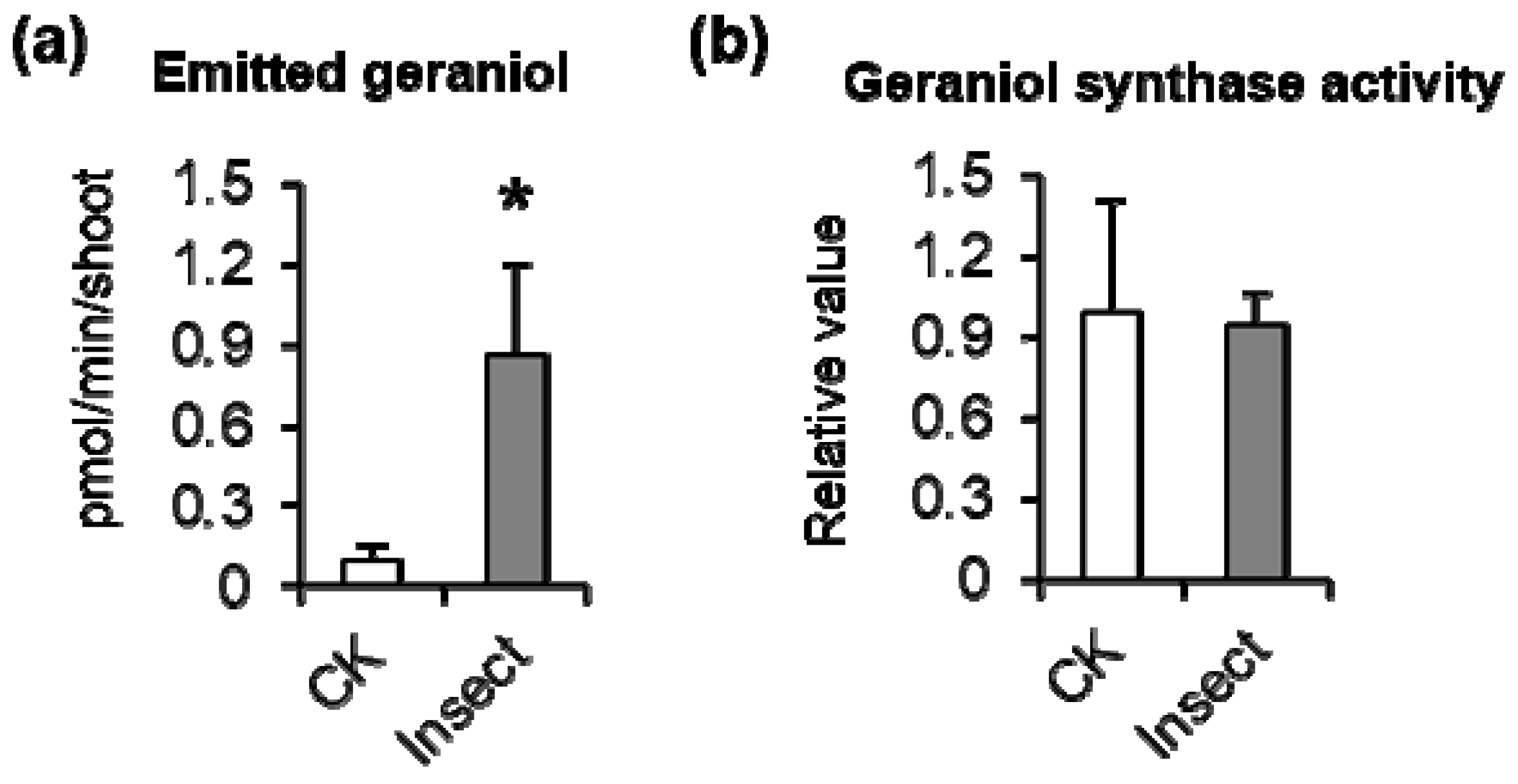
3.1. Empoasca (Matsumurasca) onukii Attack Significantly Increased Geraniol Emission from Tea Leaves
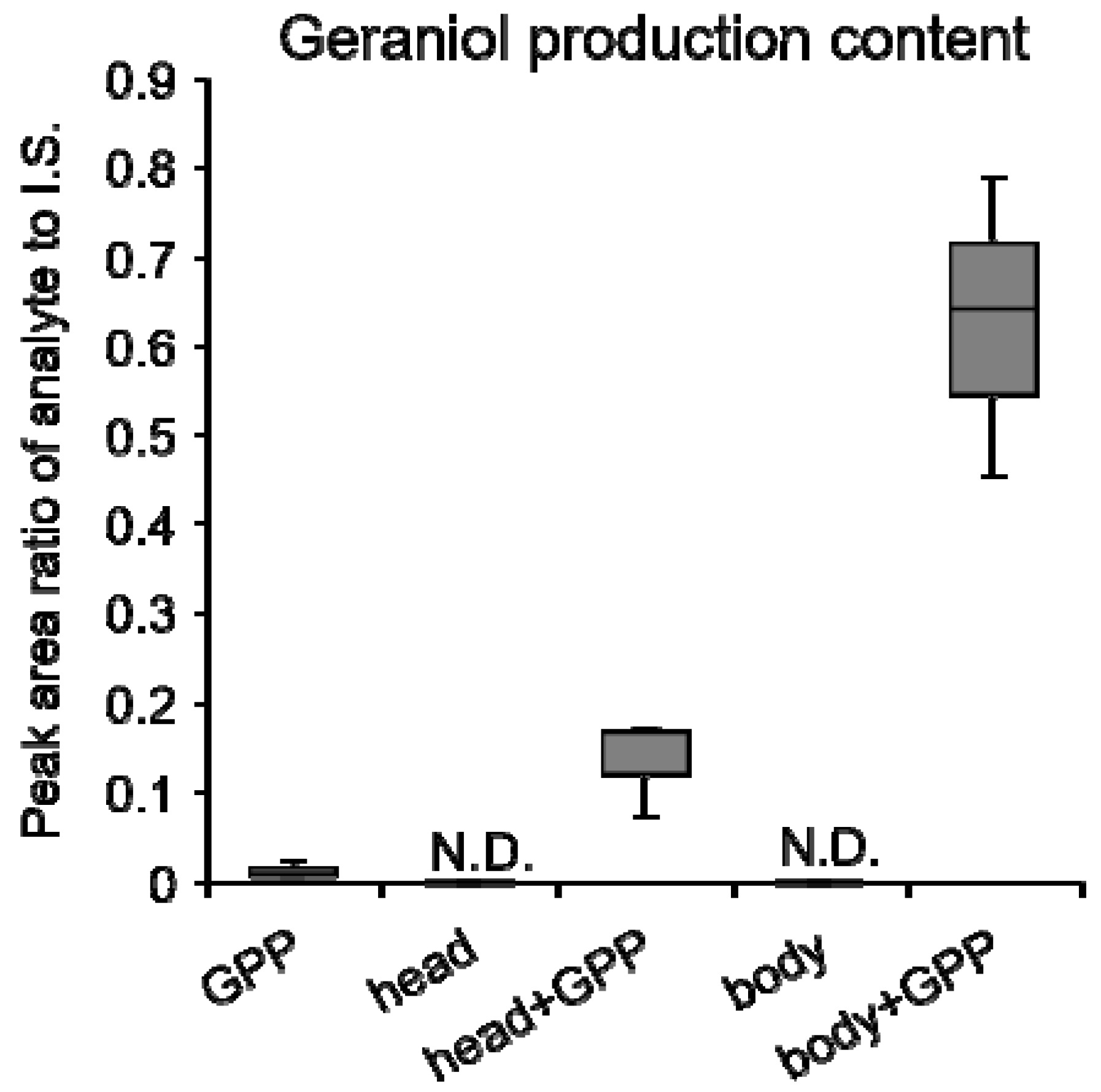
3.2. Crude Enzyme Extract of Empoasca (Matsumurasca) onukii Produced Geraniol from Greanyl Diphosphate
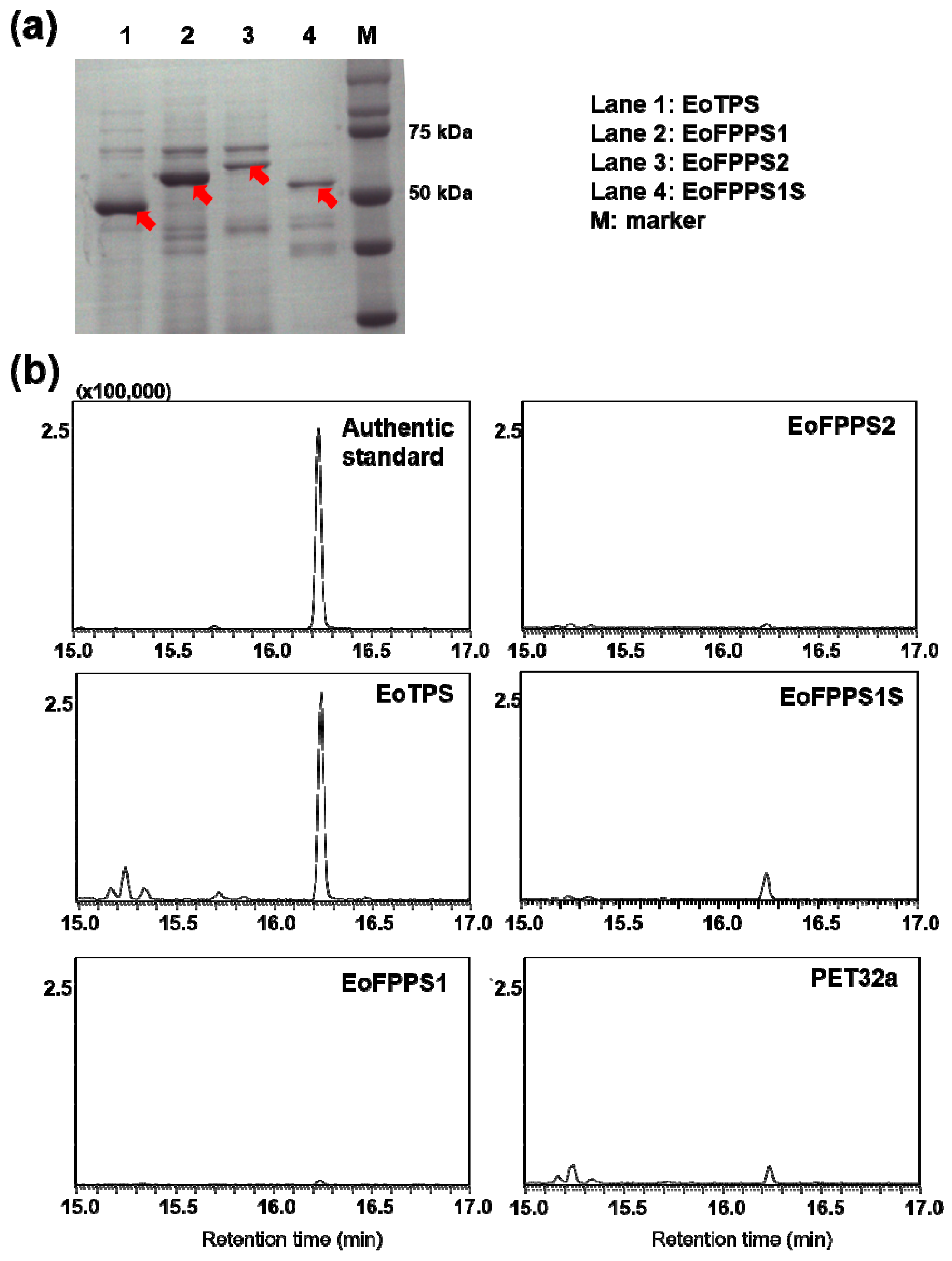
3.3. Terpene synthase from Empoasca (Matsumurasca) onukii Was Able to Convert Greanyl Diphosphate to Geraniol
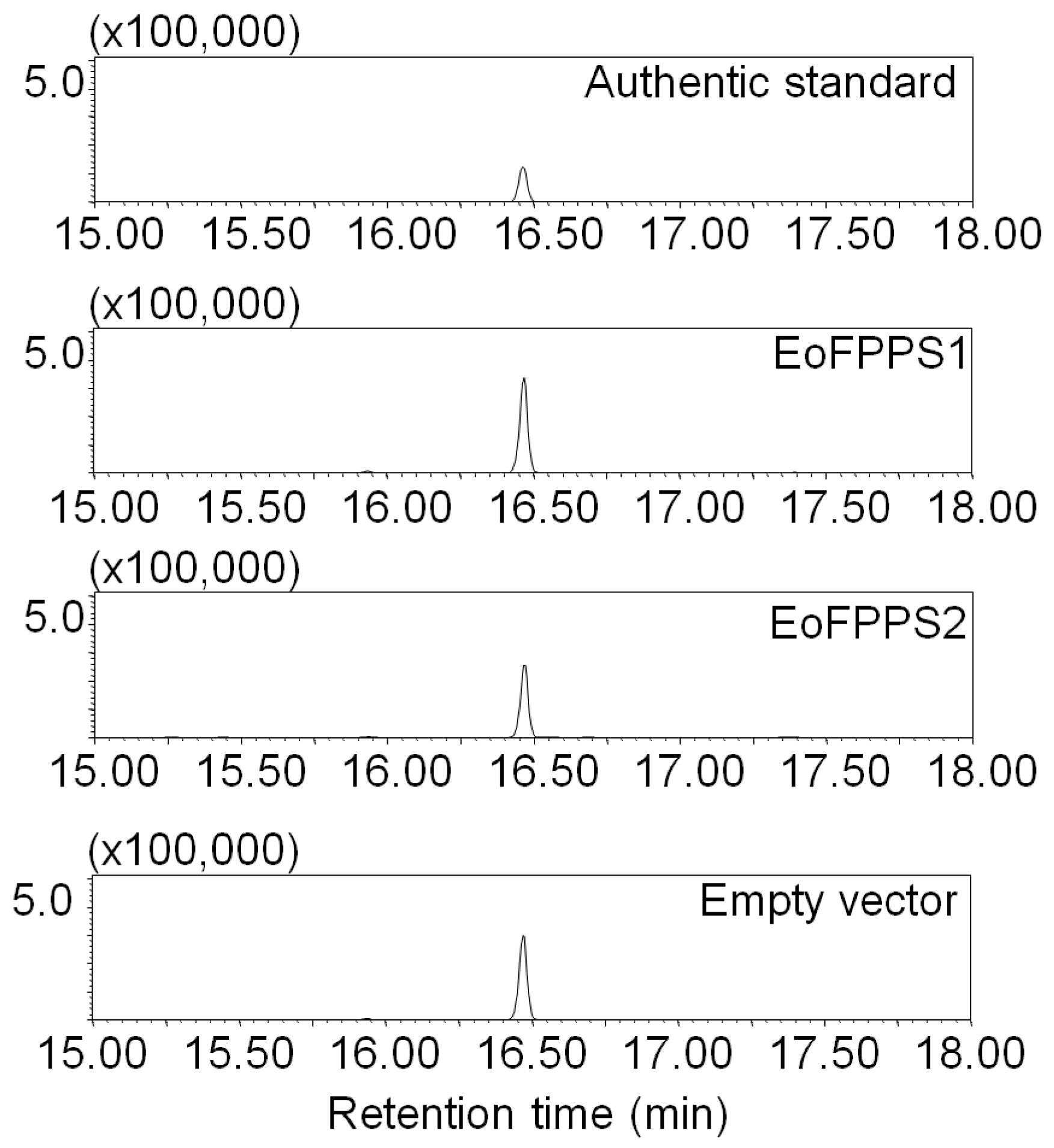
3.4. Geraniol Synthase Also Existed in Other Insects
3.5. Geraniol Had in Vitro Ability to Inhibit the Growth of Endobacterial Isolated from Empoasca (Matsumurasca) onukii
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pichersky, E.; Gershenzon, J. The formation and function of plant volatiles: Perfumes for pollinator attraction and defense. Curr. Opin. Plant Biol. 2002, 5, 237–243. [Google Scholar] [CrossRef]
- Dong, F.; Fu, X.M.; Watanabe, N.; Su, X.G.; Yang, Z.Y. Recent advances in the emission and functions of plant vegetative volatiles. Molecules 2016, 21, 124. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.E.; MithÖfer, A.; Boland, W. Before gene expression: Early events in plant-insect interaction. Trends Plant Sci. 2007, 12, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Mithöfer, A.; Wanner, G.; Boland, W. Effects of feeding Spodoptera littoralis on lima bean leaves. II. Continuous mechanical wounding resembling insect feeding is sufficient to elicit herbivory-related volatile emission. Plant Physiol. 2005, 137, 1160–1168. [Google Scholar] [CrossRef] [PubMed]
- Mei, X.; Liu, X.Y.; Zhou, Y.; Wang, X.Q.; Zeng, L.T.; Fu, X.M.; Li, J.; Tang, J.; Dong, F.; Yang, Z.Y. Formation and emission of linalool in tea (Camellia sinensis) leaves infested by tea green leafhopper (Empoasca (Matsumurasca) onukii Matsuda. Food Chem. 2017, 237, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Alborn, H.T.; Hansen, T.V.; Jones, T.H.; Bennett, D.C.; Tumlinson, J.H.; Schmelz, E.A.; Peter, E.A. Disulfooxy fatty acids from the American bird grasshopper Schistocerca americana, elicitors of plant volatiles. Proc. Natl. Acad. Sci. USA 2007, 104, 12976–12981. [Google Scholar] [CrossRef] [PubMed]
- Bonaventure, G.; VanDoorn, A.; Baldwin, I.T. Herbivore-associated elicitors: FAC signaling and metabolism. Trends Plant Sci. 2011, 16, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Mattiacci, L.; Dicke, M.; Posthumus, M.A. β-Glucosidase: An elicitor of herbivore induced plant odor that attracts host-searching parasitic wasps. Proc. Natl. Acad. Sci. USA 1995, 92, 2039–2040. [Google Scholar] [CrossRef]
- Schmelz, E.A.; Carroll, M.J.; LeClere, S.; Phipps, S.M.; Meredith, J.; Chourey, P.S.; Alborn, H.T.; Teal, P.E.A. Fragments of ATP synthase mediate plant perception of insect attack. Proc. Natl. Acad. Sci. USA 2006, 103, 8894–8899. [Google Scholar] [CrossRef]
- Allmann, S.; Baldwin, I.T. Insects betray themselves in nature to predators by rapid isomerization of green leaf volatiles. Science 2010, 329, 1075–1078. [Google Scholar] [CrossRef]
- Han, B.Y.; Chen, Z.M. Composition of the volatiles from intact and mechanically pierced tea aphid-tea shoot complexes and their attraction to natural enemies of the tea aphid. J. Agric. Food Chem. 2002, 50, 2571–2575. [Google Scholar] [CrossRef] [PubMed]
- Ishiwari, H.; Suzuki, T.; Maeda, T. Essential compounds in herbivore-induced plant volatiles that attract the predatory mite. Neoseiulus womersleyi. J. Chem. Ecol. 2007, 33, 1670–1681. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.; Yang, Z.Y.; Baldermann, S.; Sato, Y.; Asai, T.; Watanabe, N. Herbivore-induced volatiles from tea (Camellia sinensis) plants and their involvement in intraplant communication and changes in endogenous nonvolatile metabolites. J. Agric. Food Chem. 2011, 59, 13131–13135. [Google Scholar] [CrossRef]
- Yang, Z.Y.; Baldermann, S.; Watanabe, N. Recent studies of the volatile compounds in tea. Food Res. Int. 2013, 53, 585–599. [Google Scholar] [CrossRef]
- Cho, J.Y.; Mizutani, M.; Shimizu, B.; Kinoshita, T.; Ogura, M.; Tokoro, K.; Lin, M.; Sakata, K. Chemical profiling and gene expression profiling during the manufacturing process of Taiwan oolong tea “Oriental Beauty”. Biosci. Biotech. Bioch. 2007, 71, 1476–1486. [Google Scholar] [CrossRef] [PubMed]
- Ong, P.K.C.; Acree, T.E. Gas chromatography/olfactory analysis of lychee (Litchi chinesis Sonn.). J. Agric. Food Chem. 1998, 46, 2282–2286. [Google Scholar] [CrossRef]
- Luan, F.; Mosandl, A.; Münch, A.; Wüst, M. Metabolism of geraniol in grape berry mesocarp of Vitis vinifera L. cv. Scheurebe: Demonstration of stereoselective reduction, E/Z-isomerization, oxidation and glycosylation. Phytochemistry 2005, 66, 295–303. [Google Scholar] [CrossRef]
- Lan-Phi, N.T.; Shimamura, T.; Ukeda, H.; Sawamura, M. Chemical and aroma profiles of yuzu (Citrus junos) peel oils of different cultivars. Food Chem. 2009, 115, 1042–1047. [Google Scholar] [CrossRef]
- Abbott, E.; Hall, D.; Hamberger, B.; Bohlmann, J. Laser microdissection of conifer stem tissues: Isolation and analysis of high quality RNA, terpene synthase enzyme activity and terpenoid metabolites from resin ducts and cambial zone tissue of white spruce (Picea glauca). BMC Plant Biol. 2010, 10, 106. [Google Scholar] [CrossRef]
- Zhou, Y.; Zeng, L.; Liu, X.; Gui, J.; Mei, X.; Fu, X.; Dong, F.; Tang, J.; Zhang, L.; Yang, Z. Formation of (E)-nerolidol in tea (Camellia sinensis) leaves exposed to multiple stresses during tea manufacturing. Food Chem. 2017, 231, 78–86. [Google Scholar] [CrossRef]
- Hojo, M.; Matsumoto, T.; Miura, T. Cloning and expression of a geranylgeranyl diphosphate synthase gene: Insights into the synthesis of termite defence secretion. Insect Mol. Biol. 2007, 16, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Beran, F.; Rahfeld, P.; Luck, K.; Nagel, R.; Vogel, H.; Wielsch, N.; Irmisch, S.; Ramasamy, S.; Gershenzon, J.; Heckel, D.G.; et al. Novel family of terpene synthases evolved from trans-isoprenyl diphosphate synthases in a flea beetle. Proc. Natl. Acad. Sci. USA 2016, 113, 2922–2927. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Moleucar evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Qin, D.Z.; Zhang, L.; Xiao, Q.; Dietrich, C.; Matsumura, M. Clarification of the identity of the tea green leafhopper based on morphological comparison between Chinese and Japanese Specimens. PLoS ONE 2015, 10, e0139202. [Google Scholar] [CrossRef] [PubMed]
- Gilg, A.B.; Tittiger, C.; Blomquist, G.J. Unique animal prenyltransferase with monoterpene synthase activity. Naturwissenschaften 2009, 96, 731–735. [Google Scholar] [CrossRef] [PubMed]
- Perlatti, B.; Luiz, A.L.; Prieto, E.L.; Fernandes, J.B.; da Silva, M.F.G.; Ferreira, D.; Costa, E.N.; Júnior, A.L.B.; Forim, M.R. MALDI-TOF MS identification of microbiota associated with pest insect Diabrotica speciosa. Agric. For. Entomol. 2017, 19, 408–417. [Google Scholar] [CrossRef]
- Faulde, M.; Spiesberger, M. Role of the moth fly Clogmia albipunctata (Diptera: Psychodinae) as a mechanical vector of bacterial pathogens in German hospitals. J. Hosp. Infect. 2013, 83, 51–60. [Google Scholar] [CrossRef]
- Pidlyar, V.J.; Jangid, K.; Patole, M.S.; Shouche, Y.S. Studies on cultured and uncultured microbiota of wild Culex Quinquefasciatus mosquito midgut based on 16S ribosomal RNA gene analysis. Am. J. Trop. Med. Hyg. 2004, 70, 597–603. [Google Scholar] [CrossRef]
- Ceja-Navarro, J.A.; Vega, F.E.; Karaoz, U.; Hao, Z.; Jenkins, S.; Lim, H.C.; Kosina, P.; Infante, F.; Northen, T.R.; Brodie, E.L. Gut microbiota mediate caffeine detoxification in the primary insect pest of coffee. Nat. Commun. 2015, 6, 7618. [Google Scholar] [CrossRef]
- Jabs, T.; Dietrich, R.A.; Dangl, J.L. Initiation of runaway cell death in an Arabidopsis mutant by extracellular superoxide. Science 1996, 273, 1853–1856. [Google Scholar] [CrossRef]
- Izumi, S.; Nishio, Y.; Takashima, O.; Hirata, T. Monoterpenoids, potent inducer of apoptosis in cells of Marchantia polymorpha. Chem. Lett. 1997, 26, 837–838. [Google Scholar] [CrossRef]
- Izumi, S.; Takashima, O.; Hirata, T. Geraniol is a potent inducer of apoptosis-like cell death in the cultured shoot primordia of Matricaria chamomilla. Biochem. Biophys. Res. Commun. 1999, 259, 519–522. [Google Scholar] [CrossRef] [PubMed]
- James, D.G. Further field evaluation of synthetic herbivore-induced plant volatiles as attractants for beneficial insect. J. Chem. Ecol. 2005, 31, 481–495. [Google Scholar] [CrossRef] [PubMed]
- Aisaka, H.; Kosuge, M.; Yamanishi, T. Comparison of the flavors of Chinese “Keemun” black tea and Ceylon black tea. Agric. Biol. Chem. 1978, 42, 2157–2159. [Google Scholar] [CrossRef]
- Iijima, Y.; Gang, D.R.; Fridman, E.; Lewinsohn, E.; Pichersky, E. Characterization of geraniol synthase from the peltate glands of sweet basil. Plant Physiol. 2004, 134, 370–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Li, J.; Wang, H.X.; Zeng, Y. A geraniol-synthase gene from Cinnamomum tenuipilum. Phytochemistry 2005, 66, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Honda, G. Geraniol synthases from perilla and their taxonomical significance. Phytochemistry 2007, 68, 446–453. [Google Scholar] [CrossRef]
- Masumoto, N.; Korin, M.; Ito, M. Geraniol and linalool synthases from wild species of perilla. Phytochemistry 2010, 71, 1068–1075. [Google Scholar] [CrossRef]
- Simkin, A.J.; Miettinen, K.; Claudel, P.; Burlat, V.; Guirimand, G.; Courdavault, V.; Papon, N.; Meyer, S.; Godet, S.; St-Pierre, B.; et al. Characterization of the plastidial geraniol synthase from Madagascar periwinkle which initiates the monoterpenoid branch of the alkaloid pathway in internal phloem associated parenchyma. Phytochemistry 2013, 85, 36–43. [Google Scholar] [CrossRef]
- Boch, R.; Shearer, D.A. Identification of geraniol as the active component in the Nassanoff pheromone of the honey bee. Nature 1962, 194, 704–706. [Google Scholar] [CrossRef]
- Lorenz, M.; Boland, W.; Dettner, K. Biosynthesis of iridodials in the defense glands of beetle larvae (Chrysomelinae). Angew. Chem. Int. Ed. Engl. 1993, 32, 912–917. [Google Scholar] [CrossRef]
- Laurent, P.; Braekman, J.C.; Daloze, D.; Pasteels, J. Biosynthesis of defensive compounds from beetles and ants. Eur. J. Org. Chem. 2003, 2003, 2733–2743. [Google Scholar] [CrossRef]
- Burse, A.; Schmidt, A.; Frick, S.; Kuhn, J.; Gershenzon, J.; Boland, W. Iridoid biosynthesis in Chrysomelina larvae: Fat body produces early terpenoid precursors. Insect Biochem. Mol. Biol. 2007, 37, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Jarau, S.; van Veen, J.W.; Twele, R.; Reichle, C.; Herrera, E.; Gonzales, E.H.; Aguilar, I.; Francke, W.; Ayasse, M. Workers make the queens in Melipona bees: Identification of geraniol as a caster determining compound from labial glands of nurse bees. J. Chem. Ecol. 2010, 36, 565–569. [Google Scholar] [CrossRef]
- Keeling, C.; Blomquist, G.J.; Tittiger, C. Coordinated gene expression for pheromone biosynthesis in the pine engraver beetle, Ips pini (Coleoptera: Scolytidae). Naturwissenschaften 2004, 91, 324–328. [Google Scholar] [CrossRef]
- Alborn, H.T.; Turlings, T.C.J.; Jones, T.H.; Stenhagen, G.; Loughrin, J.H.; Tumlinson, J. An elicitor of plant volatiles from beet armyworm oral secretion. Science 1997, 276, 945–949. [Google Scholar] [CrossRef]
- Sorokan, A.V.; Rumyantsev, S.D.; Benkovskaya, G.V.; Maksimov, I.V. The ecological role of microsymbionts in the interaction of plants and herbivorous insect. Biol. Bull. Rev. 2019, 9, 275. [Google Scholar] [CrossRef]
- Frago, E.; Mala, M.; Weldegergis, B.T.; Yang, C.; McLean, A.; Godfray, H.C.J.; Gols, R.; Dicke, M. Symbionts protect aphids from parasitic wasps by attenuating herbivore-induced plant volatiles. Nat. Commun. 2017, 8, 1860. [Google Scholar] [CrossRef] [Green Version]
- Nasiou, E.; Giannakou, I.O. Effect of geraniol, a plant-based alcohol monoterpene oil, against Meloidogyne javanica. Eur. J. Plant Pathol. 2018, 152, 701–710. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, T.; Mi, N.; Wang, Y.; Li, G.; Wang, L.; Xie, Y. Antifungal activity of monoterpenes against wood white-rot fungi. Int. Biodeterior. Biodegrad. 2016, 106, 157–160. [Google Scholar] [CrossRef]
- Nardoni, S.; Ebani, V.V.; D’Ascenzi, C.; Pistelli, L.; Mancianti, F. Sensitivity of entomopathogenic fungi and bacteria to plants secondary metabolites, for an alternative control Rhipicephalus (Boophilus) microplus in cattle. Front. Pharmacol. 2018, 9, 937. [Google Scholar] [CrossRef] [PubMed]
- Müller, G.C.; Junnila, A.; Butler, J.; Kravchenko, V.D.; Revay, E.E.; Weiss, R.W.; Schlein, Y. Efficacy of the botanical repellents geraniol, linalool, and citronella against mosquitoes. J. Vector Ecol. 2009, 34, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Reis, S.L.; Mantello, A.G.; Macedo, J.M.; Gelfuso, E.A.; Silva, C.P.; Fachin, A.L.; Cardoso, A.M.; Beleboni, R.O. Typical monoterpenes as insecticides and repellents against strored grain pests. Molecules 2016, 21, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, A.E. Nutritional interactions in insect-microbial symbioses: Aphids and their symbiotic Bacteria Buchnera. Annu. Rev. Entomol. 1998, 43, 17–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boone, C.K.; Keefover-Ring, K.; Mapes, A.C.; Adams, A.; Bohlmann, J.; Raffa, K.F. Bacteria associated with a tree-killing insect reduce concentrations of plant defense compounds. J. Chem. Ecol. 2013, 39, 1003–1006. [Google Scholar] [CrossRef] [PubMed]
- Johanowicz, D.L.; Hoy, M.N. The manipulation of arthropod reproduction by Wolbachia endosymbionts. Fla. Entomol. 1998, 81, 310–317. [Google Scholar] [CrossRef]
- Achaleke, J.; Martin, T.; Ghogomu, R.T.; Vaissayre, M.; Brévault, T. Esterase-mediated resistance to pyrethroids in field populations of Helicoverpa armigera (Lepidoptera: Noctuidae) from Central Africa. Pest Manag. Sci. 2009, 65, 1147–1154. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Geraniol Concentration (μg/mL) | OD600 |
|---|---|
| 0 | >3 |
| 100 | >3 |
| 250 | 11.980 ± 0.232 |
| 500 | 0.018 ± 0.006 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Liu, X.; Yang, Z. Characterization of Terpene Synthase from Tea Green Leafhopper Being Involved in Formation of Geraniol in Tea (Camellia sinensis) Leaves and Potential Effect of Geraniol on Insect-Derived Endobacteria. Biomolecules 2019, 9, 808. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9120808
Zhou Y, Liu X, Yang Z. Characterization of Terpene Synthase from Tea Green Leafhopper Being Involved in Formation of Geraniol in Tea (Camellia sinensis) Leaves and Potential Effect of Geraniol on Insect-Derived Endobacteria. Biomolecules. 2019; 9(12):808. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9120808
Chicago/Turabian StyleZhou, Ying, Xiaoyu Liu, and Ziyin Yang. 2019. "Characterization of Terpene Synthase from Tea Green Leafhopper Being Involved in Formation of Geraniol in Tea (Camellia sinensis) Leaves and Potential Effect of Geraniol on Insect-Derived Endobacteria" Biomolecules 9, no. 12: 808. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9120808