KDR (VEGFR2) Genetic Variants and Serum Levels in Patients with Rheumatoid Arthritis

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Characteristics of the Study Population
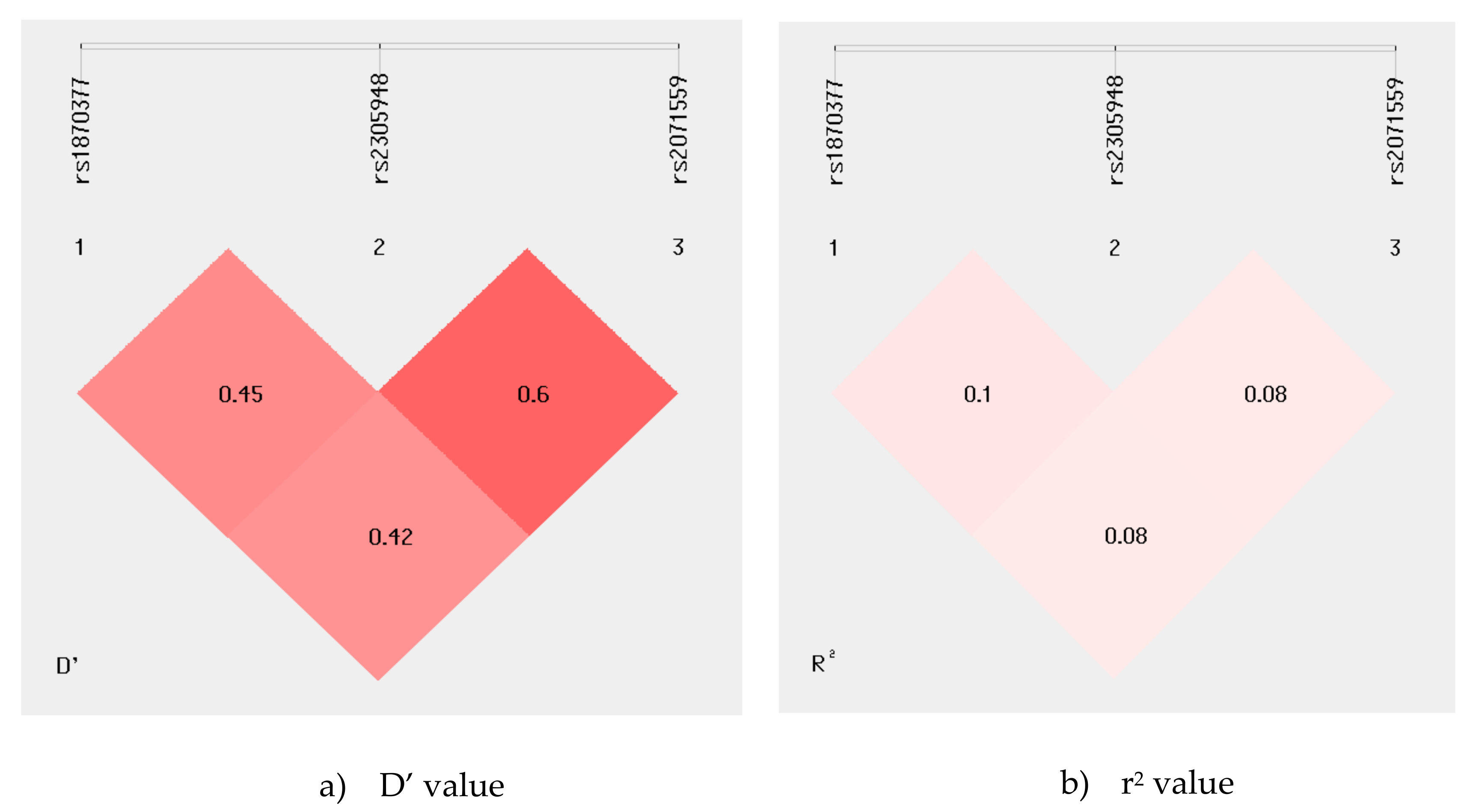
2.2. Information about KDR Gene SNPs as Well as Distribution of the KDR Genetic Variants in RA Patients and Controls
2.3. Genetic Effects of the KDR Haplotypes on the RA risk
2.4. Association between KDR rs2305948 Polymorphism and Clinical Phenotype of Rheumatoid Arthritis
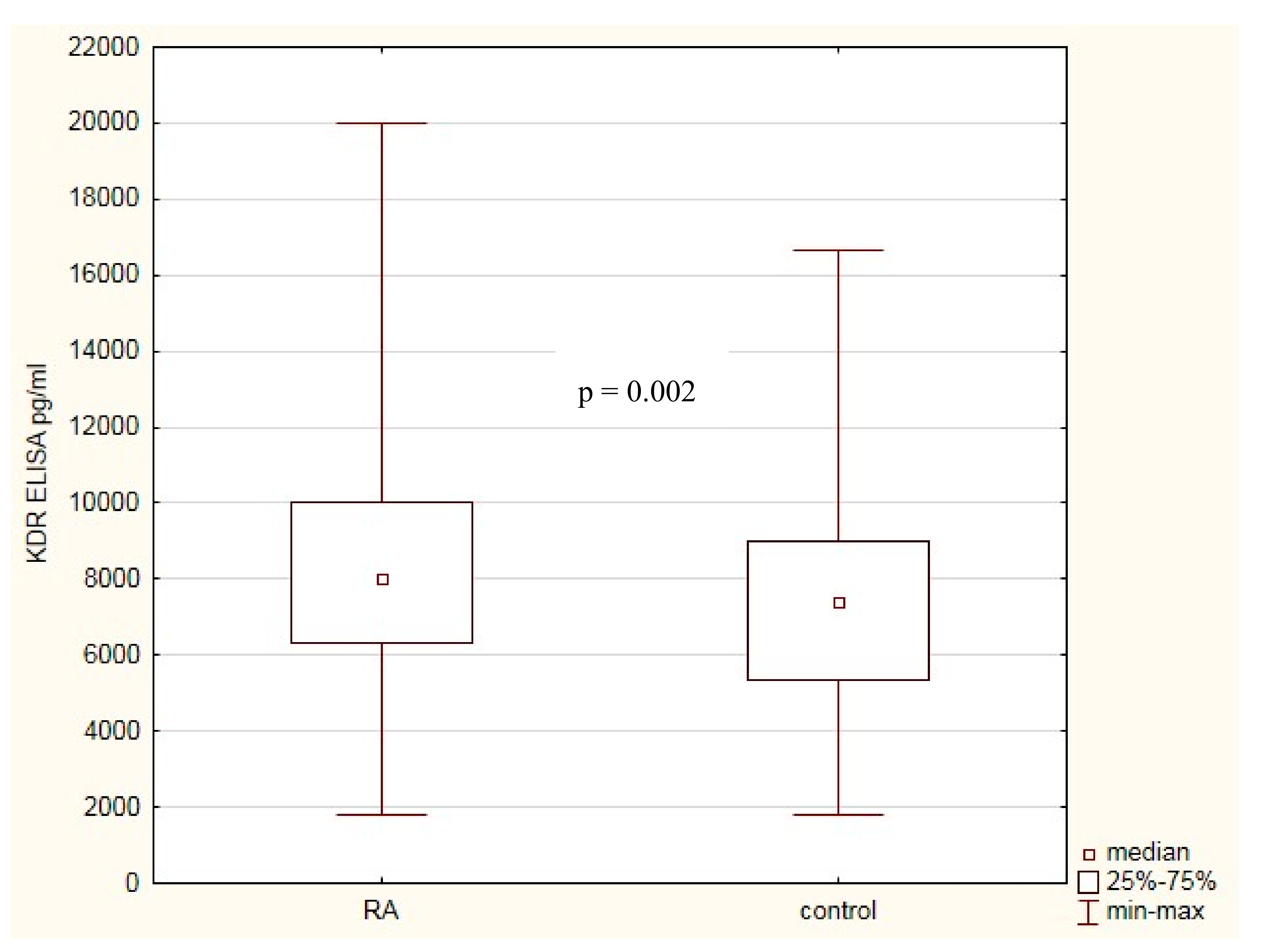
2.5. KDR Protein Level in Healthy Subjects and Patients with RA. Association with RA Clinical Phenotype
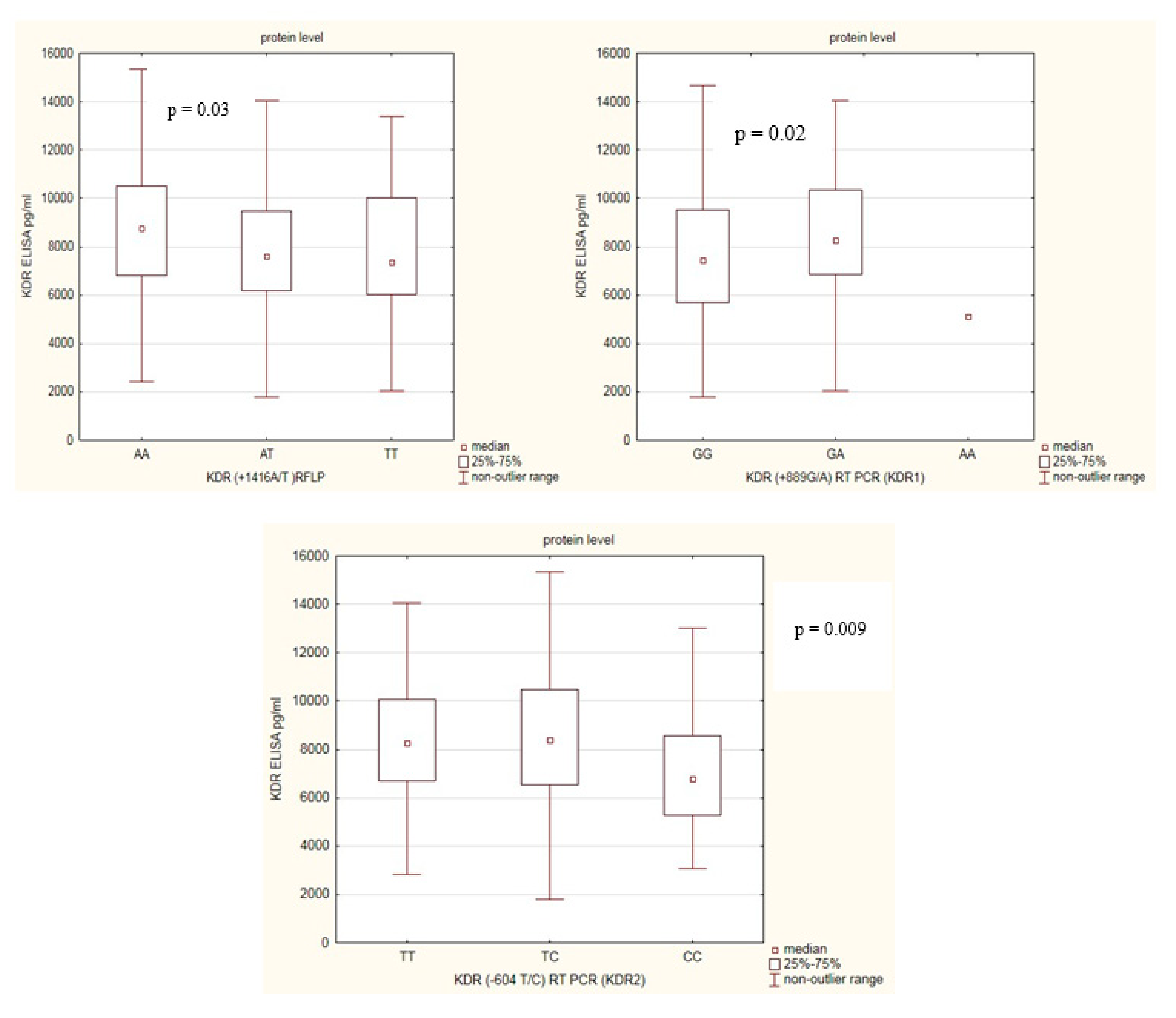
2.6. KDR Genetic Variants with Respect to KDR Protein Levels
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. SNP Selection, DNA Extraction and Genetic Analysis
4.3. Assay for Serum Levels of KDR
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Qiu, H.; Zhang, H.; Wang, L.; Zhuang, C.; Liu, R. Vascular endothelial growth factor A (VEGFA) polymorphisms in Chinese patients with rheumatoid arthritis. Scand. J. Rheumatol. 2013, 42, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Szekanecz, Z.; Koch, A.E. Angiogenesis and its targeting in rheumatoid arthritis. Vasc. Pharmacol. 2009, 90, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-R.; Kim, K.-W.; Kim, B.-M.; Cho, M.-L.; Lee, S.-H. The Effect of Vascular Endothelial Growth Factor on Osteoclastogenesis in Rheumatoid Arthritis. PLoS ONE 2015, 10, e0124909. [Google Scholar] [CrossRef] [PubMed]
- Paleolog, E.M. Angiogenesis in rheumatoid arthritis. Arthritis Res. 2002, 4, S81–S90. [Google Scholar] [CrossRef] [PubMed]
- Azizi, G.; Boghozian, R.; Mirshafiey, A. The potential role of angiogenic factors in rheumatoid arthritis. Int. J. Rheum. Dis. 2014, 17, 369–383. [Google Scholar] [CrossRef] [PubMed]
- Szekanecz, Z.; Besenyei, T.; Paragh, G.; Koch, A.E. Angiogenesis in rheumatoid arthritis. Autoimmunity 2009, 42, 563–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Yang, Y.; Yuan, H.; You, J.; Burkatovskaya, M.; Amar, S. Novel transcriptional regulation of VEGF in inflammatory processes. J. Cell. Mol. Med. 2013, 17, 386–397. [Google Scholar] [CrossRef] [PubMed]
- Rothenberg, M.E. VEGF obstructs the lungs. Nat. Med. 2004, 10, 1041. [Google Scholar] [CrossRef]
- Autiero, M.; Luttun, A.; Tjwa, M.; Carmeliet, P. Placental growth factor and its receptor, vascular endothelial growth factor receptor-1: Novel targets for stimulation of ischemic tissue revascularization and inhibition of angiogenic and inflammatory disorders. J. Thromb. Haemost. 2003, 1, 1356–1370. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Tanaka, K.; Hirata, G.; Hanada, M.; Matsuda, S.; Shuto, T.; Iwamoto, Y. Possible Involvement of the Vascular Endothelial Growth Factor-Flt-1-Focal Adhesion Kinase Pathway in Chemotaxis and the Cell Proliferation of Osteoclast Precursor Cells in Arthritic Joints. J. Immunol. 2002, 168, 5824–5831. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, M.; Hosoda, Y.; Hirose, S.; Okada, Y.; Ikeda, E. Expression of vascular endothelial growth factor isoforms and their receptors Flt-1, KDR, and neuropilin-1 in synovial tissues of rheumatoid arthritis. J. Pathol. 2000, 191, 426–433. [Google Scholar] [CrossRef]
- Shibuya, M.; Claesson-Welsh, L. Signal transduction by VEGF receptors in regulation of angiogenesis and lymphangiogenesis. Exp. Cell Res. 2006, 312, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y. Positive and Negative Modulation of Angiogenesis by VEGFR1 Ligands. Sci. Signal. 2009, 2. [Google Scholar] [CrossRef] [PubMed]
- Sundrani, D.P.; Reddy, U.S.; Joshi, A.A.; Mehendale, S.S.; Chavan-Gautam, P.M.; Hardikar, A.A.; Chandak, G.R.; Joshi, S.R. Differential placental methylation and expression of VEGF, FLT-1 and KDR genes in human term and preterm preeclampsia. Clin. Epigenetics 2013, 5, 6. [Google Scholar] [CrossRef] [PubMed]
- Sisto, M.; Lisi, S.; Lofrumento, D.D.; D’Amore, M.; Frassanito, M.A.; Ribatti, D. Sjögren’s syndrome pathological neovascularization is regulated by VEGF-A-stimulated TACE-dependent crosstalk between VEGFR2 and NF-κB. Genes Immun. 2012, 13, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Hao, Q.; Wang, L.; Tang, H. Vascular endothelial growth factor induces protein kinase D-dependent production of proinflammatory cytokines in endothelial cells. Am. J. Physiol. Physiol. 2009, 296, C821–C827. [Google Scholar] [CrossRef]
- Fava, R.A.; Olsan, N.J.; Spancer-Green, G.; Yeo, K.T.; Yeo, T.K.; Berse, B.; Jackman, R.W.; Senger, D.R.; Dvorak, H.F.; Brown, L.F. Vascular permeability factor/endothelial growth factor (VPF/VEGF): Accumulation and expression in human synovial fluids and rheumatoid synovial tissue. J. Exp. Med. 1994, 180, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Ballara, S.; Taylor, P.C.; Reusch, P.; Feldmann, M.; Maini, R.N.; Paleolog, E.M.; Marmé, D. Raised serum vascular endothelial growth factor levels are associated with destructive change in inflammatory arthritis. Arthritis Rheum. 2001, 44, 2055–2064. [Google Scholar] [CrossRef]
- Clavel, G.; Bessis, N.; Lemeiter, D.; Fardellone, P.; Mejjad, O.; Ménard, J.-F.; Pouplin, S.; Boumier, P.; Vittecoq, O.; Le Loët, X.; et al. Angiogenesis markers (VEGF, soluble receptor of VEGF and angiopoietin-1) in very early arthritis and their association with inflammation and joint destruction. Clin. Immunol. 2007, 124, 158–164. [Google Scholar] [CrossRef]
- Lee, S.S.; Joo, Y.S.; Kim, W.U.; Min, D.J.; Min, J.K.; Park, S.H.; Cho, C.S.; Kim, H.Y. Vascular endothelial growth factor levels in the serum and synovial fluid of patients with rheumatoid arthritis. Clin. Exp. Rheumatol. 2001, 19, 321–324. [Google Scholar]
- Sone, H.; Sakauchi, M.; Takahashi, A.; Suzuki, H.; Inoue, N.; Iida, K.; Shimano, H.; Toyoshima, H.; Kawakami, Y.; Okuda, Y.; et al. Elevated levels of vascular endothelial growth factor in the sero of patients with rheumatoid arthritis correlation with disease activity. Life Sci. 2001, 69, 1861–1869. [Google Scholar] [CrossRef]
- Carvalho, J.F.; Blank, M.; Shoenfeld, Y. Vascular Endothelial Growth Factor (VEGF) in Autoimmune Diseases. J. Clin. Immunol. 2007, 27, 246–256. [Google Scholar] [CrossRef]
- Paradowska-Gorycka, A.; Pawlik, A.; Romanowska-Prochnicka, K.; Haladyj, E.; Malinowski, D.; Stypinska, B.; Manczak, M.; Olesinska, M. Relationship between VEGF Gene Polymorphisms and Serum VEGF Protein Levels in Patients with Rheumatoid Arthritis. PLoS ONE 2016, 11, e0160769. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, M.; Durães, C.; Oliveira, A.; Martins, M.J.; Lucas, R.; Costa, L.; Pereira, J.G.; Ramos, I.; Machado, J.C.; Simões-Ventura, F. LRP5 gene polymorphisms and radiographic joint damage in rheumatoid arthritis patients. Osteoporos. Int. 2018, 29, 2355–2368. [Google Scholar] [CrossRef] [PubMed]
- Zhi, L.; Yao, S.; Ma, W.; Zhang, W.; Chen, H.; Li, M.; Ma, J. Polymorphisms of RAD51B are associated with rheumatoid arthritis and erosion in rheumatoid arthritis patients. Sci. Rep. 2017, 7, 45876. [Google Scholar] [CrossRef]
- Torices, S.; Julia, A.; Muñoz, P.; Varela, I.; Balsa, A.; Marsal, S.; Fernández-Nebro, A.; Blanco, F.; López-Hoyos, M.; Martinez-Taboada, V.; et al. A functional variant of TLR10 modifies the activity of NFkB and may help predict a worse prognosis in patients with rheumatoid arthritis. Arthritis Res. Ther. 2016, 18, 221. [Google Scholar] [CrossRef] [PubMed]
- López-Lasanta, M.; Julia, A.; Maymó, J.; Fernández-Gutiérrez, B.; Ureña-Garnica, I.; Blanco, F.J.; Cañete, J.D.; Alperi-López, M.; Olivé, A.; Corominas, H.; et al. Variation at interleukin-6 receptor gene is associated to joint damage in rheumatoid arthritis. Arthritis Res. Ther. 2015, 17, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addobbati, C.; da Cruz, H.L.A.; Adelino, J.E.; Melo Tavares Ramos, A.L.; Fragoso, T.S.; Domingues, A.; Branco Pinto Duarte, Â.L.; Oliveira, R.D.R.; Louzada-Júnior, P.; Donadi, E.A.; et al. Polymorphisms and expression of inflammasome genes are associated with the development and severity of rheumatoid arthritis in Brazilian patients. Inflamm. Res. 2018, 67, 255–264. [Google Scholar] [CrossRef]
- Paradowska-Gorycka, A.; Sowinska, A.; Pawlik, A.; Malinowski, D.; Stypinska, B.; Haladyj, E.; Romanowska-Prochnicka, K.; Olesinska, M. FLT-1 gene polymorphisms and protein expression profile in rheumatoid arthritis. PLoS ONE 2017, 12, e0172018. [Google Scholar] [CrossRef]
- Vernerova, L.; Špoutil, F.; Vlček, M.; Krskova, K.; Penesova, A.; Meskova, M.; Marko, A.; Raslova, K.; Vohnout, B.; Rovenský, J.; et al. A Combination of CD28 (rs1980422) and IRF5 (rs10488631) Polymorphisms Is Associated with Seropositivity in Rheumatoid Arthritis: A Case Control Study. PLoS ONE 2016, 11, e0153316. [Google Scholar] [CrossRef]
- Wang, Y.; Zheng, Y.; Zhang, W.; Yu, H.; Lou, K.; Zhang, Y.; Qin, Q.; Zhao, B.; Yang, Y.; Hui, R. Polymorphisms of KDR gene are associated with coronary heart disease. J. Am. Coll. Cardiol. 2007, 50, 760–767. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-W.; Shin, E.-S.; Lee, J.; Bahn, J.-W.; Oh, H.-B.; Oh, S.-Y.; Cho, S.-H.; Moon, H.-B.; Min, K.-U. Association between genetic variations of vascular endothelial growth factor receptor 2 and atopy in the Korean population. J. Allergy Clin. Immunol. 2006, 117, 774–779. [Google Scholar] [CrossRef] [PubMed]
- Gałecki, P.; Orzechowska, A.; Berent, D.; Talarowska, M.; Bobińska, K.; Gałecka, E.; Lewiński, A.; Maes, M.; Szemraj, J. Vascular endothelial growth factor receptor 2 gene (KDR) polymorphisms and expression levels in depressive disorder. J. Affect. Disord. 2013, 147, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, M.; Viezens, L.; Fuhrhop, I.; Ruther, W.; Schaefer, C.; Schwarzloh, B.; Algenstaedt, P.; Fink, B.; Hansen-Algenstaedt, N. Angiogenic growth factors in rheumatoid arthritis. Rheumatol. Int. 2013, 33, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Sekimoto, T.; Hamada, K.; Oike, Y.; Matsuoka, T.; Matsumoto, M.; Chosa, E.; Suda, T.; Tajima, N.; Yamamura, K. Effect of direcy angiogenesis inhibition in rheumatoid arthritis using a soluble vascular endothelial growth factor receptor 1 chimeric protein. J. Rheumatol. 2002, 29, 240–245. [Google Scholar] [PubMed]
- Ulyatt, C.; Walker, J.; Ponnambalam, S. Hypoxia differentially regulates VEGFR1 and VEGFR2 levels and alters intracellular signaling and cell migration in endothelial cells. Biochem. Biophys. Res. Commun. 2011, 404, 774–779. [Google Scholar] [CrossRef]
- Clarke, J.M.; I Hurwitz, H. Targeted inhibition of VEGF Receptor-2: An update on Ramucirumab. Expert Opin. Biol. Ther. 2013, 13, 1187–1196. [Google Scholar] [CrossRef] [PubMed]
- Fontanella, C.; Ongaro, E.; Bolzonello, S.; Guardascione, M.; Fasola, G.; Aprile, G. Clinical advances in the development of novel VEGFR2 inhibitors. Ann. Transl. Med. 2014, 2, 123. [Google Scholar]
- Liu, D.; Song, J.; Ji, X.; Liu, Z.; Cong, M.; Hu, B. Association of Genetic Polymorphisms on VEGFA and VEGFR2 With Risk of Coronary Heart Disease. Medicine (Baltimore) 2016, 95, e3413. [Google Scholar] [CrossRef]
- Zhang, W.; Sun, K.; Zhen, Y.; Wang, D.; Wang, Y.; Chen, J.; Xu, J.; Hu, F.B.; Hui, R. VEGF Receptor-2 Variants Are Associated With Susceptibility to Stroke and Recurrence. Stroke 2009, 40, 2720–2726. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Xu, W.; Kamel-Reid, S.; Liu, X.; Jung, C.W.; Kim, S.; Lipton, J.H. Clinical relevance of vascular endothelial growth factor (VEGFA) and VEGF receptor (VEGFR2) gene polymorphism on the treatment outcome following imatinib therapy. Ann. Oncol. 2010, 21, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Sjostrom, S.; Wibom, C.; Andersson, U.; Brännström, T.; Broholm, H.; Johansen, C.; Collatz-Laier, H.; Liu, Y.; Bondy, M.; Henriksson, R.; et al. Genetic variations in VEGF and VEGFR2 and glioblastoma outcome. J. Neurooncol. 2010, 104, 523–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beeghly-Fadiel, A.; Shu, X.O.; Lu, W.; Long, J.; Cai, Q.; Xiang, Y.B.; Zheng, Y.; Zhao, Z.; Gu, K.; Gao, Y.T.; et al. Genetic variation in VEGF family genes and breast cancer risk: A report from the Shanghai Breast Cancer Genetics Study. Cancer Epidemiol. Biomarkers Prev. 2011, 20, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Schneider, B.P.; Wang, M.; Radovich, M.; Sledge, G.W.; Badve, S.; Thor, A.; Flockhart, D.A.; Hancock, B.; Davidson, N.; Gralow, J.; et al. Association of vascular endothelial growth factor and vascular endothe lial growth factor receptor-2 genetic polymorphisms with outcome in a trial of paclitaxel compared with paclitaxel plus bevacizumab in advanced breast cancer: ECOG 2100. J. Clin. Oncol. 2008, 26, 4672–4678. [Google Scholar] [CrossRef] [PubMed]
- Steeghs, N.; Gelderblom, H.; Wessels, J.; Eskens, F.A.; de Bont, N.; Nortier, J.W.; Guchelaar, H.J. Pharmacogenetics of telatinib, a VEGFR-2 and VEGFR-3 tyrosine kinase inhibitor, used in patients with solid tumors. Invest. New Drugs 2011, 29, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Kariž, S.; Petrovič, D. Minor association of kinase insert domain-containing receptor gene polymorphism (rs2071559) with myocardial infarction in Caucasians with type 2 diabetes mellitus: Case-control cross-sectional study. Clin. Biochem. 2014, 47, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.P.; Wu, Y.Z.; Yu, N.; Yu, Z.W.; Xie, F.Y.; Yuan, Q. VEGF Gene Polymorphisms Affect Serum Protein Levels and Alter Disease Activity and Synovial Lesions in Rheumatoid Arthritis. Med. Sci. Monit. 2016, 22, 316–324. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, I.; Riera, P.; Andrés, M.; Altés, A.; Majem, M.; Blanco, R.; Capdevila, L.; Barba, A.; Barnadas, A.; Salazar, J. Prognostic effect of VEGF gene variants in metastatic non-small-cell lung cancer patients. Angiogenesis 2019, 22, 433–440. [Google Scholar] [CrossRef]
- Song, Y.; Yang, Y.; Liu, L.; Liu, X. Association between five polymorphisms in vascular endothelial growth factor gene and urinary bladder cancer risk: A systematic review and meta-analysis involving 6671 subjects. Gene 2019, 698, 186–197. [Google Scholar] [CrossRef]
- Zhang, L.F.; Ren, K.W.; Zuo, L.; Zou, J.G.; Song, N.H.; Mi, Y.Y.; Wang, Z.J.; Zhang, W. VEGF gene rs3025039C/T and rs833052C/A variants are associated with bladder cancer risk in Asian descendants. J. Cell Biochem. 2019, 120, 10402–10412. [Google Scholar] [CrossRef]
- Innocenti, F.; Jiang, C.; Sibley, A.B.; Etheridge, A.S.; Hatch, A.J.; Denning, S.; Niedzwiecki, D.; Shterev, I.D.; Lin, J.; Furukawa, Y.; et al. Genetic variation determines VEGF-A plasma levels in cancer patients. Sci. Rep. 2018, 8, 16332. [Google Scholar] [CrossRef] [PubMed]
- Gong, M.; Dong, W.; Shi, Z.; Qiu, S.; Yuan, R. Vascular endothelial growth factor gene polymorphisms and the risk of renal cell carcinoma: Evidence from eight case-control studies. Oncotarget 2017, 8, 8447–8458. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xu, A.; Xu, H. The roles of vascular endothelial growth factor gene polymorphisms in congenital heart diseases: A meta-analysis. Growth Factors 2018, 36, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Baz-Dávila, R.; Espinoza-Jiménez, A.; Rodríguez-Pérez, M.d.C.; Zulueta, J.; Varo, N.; Montejo, Á.; Almeida-González, D.; Aguirre-Jaime, A.; Córdoba-Lanús, E.; Casanova, C. Role of HIF1A, VEGFA and VEGFR2 SNPs in the Susceptibility and Progression of COPD in a Spanish Population. PLoS ONE 2016, 11, e0154998. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Niu, H.; Li, Y.; He, P.; Li, Q.; Ouyang, Y.; Li, M.; Hu, Z.; Zhong, Y.; Sun, P.; et al. Polymorphisms in VEGF-A are associated with COPD risk in the Chinese population from Hainan province. J. Genet. 2016, 95, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Sellami, N.; Lamine, L.B.; Turki, A.; Sarray, S.; Jailani, M.; Al-Ansari, A.K.; Ghorbel, M.; Mahjoub, T.; Almawi, W.Y. Association of VEGFA variants with altered VEGF secretion and type 2 diabetes: A case-control study. Cytokine 2018, 106, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Tung, M.-C.; Hsieh, M.-J.; Wang, S.-S.; Yang, S.-F.; Chen, S.-S.; Wang, S.-W.; Lee, L.-M.; Lee, W.-J.; Chien, M.-H. Associations of VEGF-C Genetic Polymorphisms with Urothelial Cell Carcinoma Susceptibility Differ between Smokers and Non-Smokers in Taiwan. PLoS ONE 2014, 9, e91147. [Google Scholar] [CrossRef]
- Chien, M.H.; Liu, Y.F.; Hsin, C.H.; Lin, C.H.; Shih, C.H.; Yang, S.F.; Cheng, C.W.; Lin, C.W. Impact of VEGF-C gene polymorphisms and environmental factors on oral cancer susceptibility in Taiwan. PLoS ONE 2013, 8, e60283. [Google Scholar] [CrossRef]
- Hsieh, M.C.; Hsu, H.T.; Hsiao, P.C.; Yang, S.F.; Yeh, C.B.; Bien, M.Y.; Lin, C.H.; Chien, M.H. Role of VEGF-C gene polymorphisms in susceptibility to hepatocellular carcinoma and its pathological development. J. Clin. Lab. Anal. 2014, 28, 237–244. [Google Scholar] [CrossRef]
- Hsueh, K.C.; Lin, Y.J.; Chang, J.S.; Wan, L.; Tsai, Y.H.; Tsai, C.H.; Chen, C.P.; Tsai, F.J. Association of vascular endothelial growth factor C-634 g polymorphism in taiwanese children with Kawasaki disease. Pediatr. Cardiol. 2008, 29, 292–296. [Google Scholar] [CrossRef]
- Yong, Y.; He, L. SHEsis, a powerful software platform for analyses of linkage disequilibrium, haplotype construction, and genetic association at polymorphism loci. Cell Res. 2005, 15, 97–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Zhang, Z.; He, Z.; Tang, W.; Li, T.; Zeng, Z.; He, L.; Shi, Y. A partition-ligation-combination-subdivision EM algorithm for haplotype inference with multiallelic markers: Update of the SHEsis (http://analysis.bio-x.cn). Cell Res. 2009, 19, 519–523. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Characteristics | RA Patients | |
|---|---|---|
| N | Mediana (IQR) | |
| Age (years) | 616 | 56.2 ± 12.5 (22–89) |
| Disease duration (years) | 524 | 11.2 ± 8.7 (0–48) |
| Larsen | 567 | 3.0 ± 1.0 (0–5) |
| Number of swollen joints | 340 | 4.6 ± 4.9 (0–26) |
| Number of tender joints | 338 | 8.0 ± 6.1 (0–28) |
| ESR (mm/h) | 565 | 35.0 ± 24.5 (0–164) |
| CRP (mg/L) | 345 | 22.5 ± 24.2 (0–144) |
| Hemoglobin (g/dL) | 345 | 12.5 ± 1.5 (8.1–16.5) |
| VAS (mm) | 331 | 52.3 ± 23.9 (0–100) |
| DAS 28-CRP | 334 | 4.89 ± 1.40 (0.97–7.95) |
| HAQ | 288 | 1.47 ± 0.72 (0–3.125) |
| PLT (× 103/mm3) | 345 | 321.17 ± 106.10 (0–948) |
| Creatinine | 344 | 0.73 ± 0.24 (0–2.60) |
| N | n (%) | |
| RF presence | 561 | 386 (69%) |
| anti-CCP presence | 348 | 280 (80%) |
| Coronary artery disease CAD | 343 | 46 (13%) |
| Hypertension (HNT) | 344 | 123 (36%) |
| Myocarditis (MI) | 341 | 12 (3.5%) |
| Parameter Continuous Variables | Patients with Cardiovascular Diseases | Patients without Cardiovascular Diseases | p | ||
|---|---|---|---|---|---|
| N | Median (IQR) | N | Median (IQR) | ||
| age (years) | 145 | 62 (56–68) | 199 | 53 (44–60) | 0.000 |
| disease duration (years) | 132 | 10 (6–17) | 167 | 10 (5–17) | 0.932 |
| Larsen | 144 | 3 (2–4) | 199 | 3 (3–4) | 0.913 |
| number of tender joints | 140 | 8 (4–13.5) | 193 | 7 (2–11) | 0.247 |
| number of swollen joints | 141 | 3 (0–7) | 194 | 3 (1–7) | 0.280 |
| ESR (mm/h) | 144 | 30 (15.5–45) | 198 | 24 (12–40) | 0.046 |
| CRP (mg/L) | 141 | 15.7 (8.0–36.7) | 198 | 12.2 (5.1–32.0) | 0.053 |
| Hb | 142 | 12.8 (11.7–13.6) | 197 | 12.6 (11.5–13.5) | 0.431 |
| VAS (mm) | 139 | 58 (32–75) | 188 | 52 (34–69) | 0.539 |
| DAS-28 | 139 | 5.10 (3.98–6.1) | 191 | 5.05 (3.77–5.89) | 0.278 |
| HAQ | 123 | 1.63 (1.00–2.13) | 158 | 1.38 (0.75–1.88) | 0.018 |
| PLT | 142 | 298.5 (243–362) | 197 | 317 (259–386) | 0.277 |
| Creatinine | 142 | 0.70 (0.60–0.87) | 196 | 0.70 (0.60–0.80) | 0.006 |
| parameter discrete variables | patients with cardiovascular diseases | patients without cardiovascular diseases | p | ||
| N | n (%) | N | n (%) | ||
| women | 145 | 133 (92%) | 198 | 182 (92%) | 0.948 |
| RF + | 144 | 102 (71%) | 193 | 117 (61%) | 0.052 |
| anti-CCP + | 143 | 121 (85%) | 197 | 153 (78%) | 0.110 |
| KDR SNP | Genotype | RA n (%) | Controls n (%) | Adjusted OR (95% CI) | p-Value |
|---|---|---|---|---|---|
| rs1870377A/T (+1416A/T) | |||||
| Codominant | AA | 291 (48%) | 167 (53%) | 1 | - |
| AT | 239 (40%) | 120 (38%) | 0.78 (0.55–1.09) | 0.139 | |
| TT | 72 (12%) | 26 (8%) | 1.76 (1.09–2.85) | 0.020 | |
| Dominant | AA | 291 (48%) | 167 (53%) | 1 | |
| AT + TT | 311 (52%) | 146 (47%) | 1.12 (0.91–1.39) | 0.286 | |
| Recessive | AA + AT | 530 (88%) | 287 (92%) | 1 | |
| TT | 72 (12%) | 26 (8%) | 1.53 (1.07–2.20) | 0.019 | |
| Allele | A | 821 (68%) | 454 (73%) | 1 | - |
| T | 383 (32%) | 172 (27%) | 1.38 (1.00–1.92) | 0.053 | |
| rs2305948 G/A (+889G/A) | |||||
| Codominant | GG | 409 (64%) | 245 (72%) | 1 | - |
| GA | 224 (35%) | 94 (28%) | 0.72 (0.21–2.55) | 0.614 | |
| AA | 8 (1%) | 1 (0%) | 3.56 (0.30–42.35) | 0.314 | |
| Dominant | GG | 409 (64%) | 245 (72%) | 1 | |
| GA + AA | 232 (36%) | 95 (28%) | 1.38 (1.10–1.73) | 0.005 | |
| Recessive | GG + GA | 633 (99%) | 339 (100%) | 1 | |
| AA | 8 (1%) | 1 (0%) | 2.73 (0.43–17.37) | 0.287 | |
| Alleles | G | 1042 (81%) | 584 (86%) | 1 | - |
| A | 240 (19%) | 96 (14%) | 1.71 (1.15–2.54) | 0.008 | |
| rs2071559 T/C (−604 T/C) | |||||
| Codominant | TT | 163 (26%) | 17 (5%) | 1 | - |
| TC | 335 (52%) | 239 (70%) | 0.51 (0.37–0.69) | 0.000 | |
| CC | 141 (22%) | 84 (25%) | 0.57 (0.39–0.81) | 0.002 | |
| Dominant | TT | 163 (26%) | 17 (5%) | 1 | |
| TC + CC | 476 (74%) | 323 (95%) | 0.39 (0.27–0.55) | 0.000 | |
| Recessive | TT + TC | 498 (78%) | 256 (75%) | 1 | |
| CC | 141 (22%) | 84 (25%) | 0.90 (0.71–1.14) | 0.360 | |
| Alleles | T | 661 (52%) | 273 (40%) | 1 | - |
| C | 617 (48%) | 407 (60%) | 0.60 (0.45–0.81) | 0.001 | |
| Haplotype 1416A/T 889G/A 604T/C | RA 2n = 1200 (freq) | Control 2n = 622 (freq) | p-Value | Odds Ratio (95%CI) |
|---|---|---|---|---|
| AGC | 449 (0.374) | 310 (0.498) | <0.001 | 0.601 (0.494–0.731) |
| TAT | 116 (0.096) | 47 (0.075) | 0.141 | 1.309 (0.919–1.864) |
| TGT | 154 (0.128) | 67 (0.107) | 0.225 | 1.219 (0.899–1.654) |
| AGT | 286 (0.238) | 107 (0.172) | 0.001 | 1.506 (1.176–1.927) |
| TGC | 85 (0.07) | 47 (0.075) | 0.704 | 0.932 (0.644–1.35) |
| AAT | 66 (0.055) | 23 (0.036) | 0.108 | 1.515 (0.933–2.461) |
| Parameter | GG | GA + AA | p * | ||
|---|---|---|---|---|---|
| N | Median (IQR) | N | Median (IQR) | ||
| Age (years) | 631 | 50 (32–59) | 23 | 52 (33–62) | 0.737 |
| Disease duration (years) | 322 | 10 (4–16) | 202 | 9 (5–15) | 0.588 |
| Larsen | 355 | 3 (3–3) | 211 | 3 (3–4) | 0.030 |
| Number of tender joints | 175 | 8 (4–12) | 163 | 6 (2–11) | 0.062 |
| Number of swollen joints | 176 | 4 (2–9) | 164 | 2 (0–6) | 0.000 |
| ESR (mm/h) | 355 | 31 (19–51) | 209 | 28 (15–40) | 0.006 |
| CRP (mg/L) | 177 | 17 (8–40) | 167 | 11 (5–25) | 0.000 |
| Hemoglobin (g/dL) | 177 | 12.6 (11.6–13.3) | 167 | 12.8 (11.7–13.8) | 0.201 |
| VAS (mm) | 170 | 60 (45–75) | 161 | 47 (30–67) | 0.000 |
| DAS 28-CRP | 172 | 5.3 (4.1–6) | >162 | 4.7 (3.5–5.6) | 0.000 |
| HAQ | >152 | 1.62 (1.06–2) | 135 | 1.37 (0.75–2) | 0.027 |
| PLT (× 103/mm3) | 177 | 312 (255–383) | 166 | 304 (245–370) | 0.567 |
| Creatinine | 174 | 0.7 (0.6–0.8) | 167 | 0.7 (0.6–0.8) | 0.027 |
| p ** | |||||
| N | n (%) | N | n (%) | ||
| Women | 635 | 481 (76%) | 322 | 277 (86%) | 0.000 |
| RF presence | 352 | 241 (68%) | 208 | 144 (69%) | 0.850 |
| anti-CCP presence | 179 | 141 (77%) | 168 | 138 (72%) | 0.429 |
| Parameter | KDR Protein Level | KDR Protein Level | p | ||||
|---|---|---|---|---|---|---|---|
| Parameter Group I | N | Median (IQR) | Parameter Group II | N | Median (IQR) | ||
| age | age ≥ 56 | 146 | 8042 (6575–9935) | age < 56 | 126 | 7893 (5836–10246) | 0.337 |
| sex | women | 253 | 7955 (6226–9848) | men | 19 | 9867 (6575–10910) | 0.093 |
| RF | RF + | 169 | 7964 (6042–10032) | RF - | 97 | 8457 (6595–10246) | 0.413 |
| anti-CCP | a-CCP + | 207 | 8006 (6294–10032) | a-CCP - | 58 | 8254 (6438–10323) | 0.586 |
| disease duration | ≥10 | 151 | 8065 (6429–10032) | <10 | 113 | 7825 (6101–10140) | 0.894 |
| ESR | ≥30 | 124 | 8269 (6434–9966) | <30 | 141 | 7825 (6042–10128) | 0.420 |
| number of tender joints | ≥7 | 155 | 7415 (5852–9997) | <7 | 107 | 8370 (6892–10360) | 0.081 |
| number of swollen joints | ≥3 | 141 | 7375 (5836–9997) | <3 | 121 | 8449 (7008–10323) | 0.081 |
| CRP | ≥13 | 121 | 8006 (6006–9518) | <13 | 143 | 8091 (6556–10450) | 0.191 |
| DAS-28 | ≥5.0 | 127 | 7791 (5894–9997) | <5.0 | 133 | 8032 (6556–10246) | 0.508 |
| HAQ | ≥1.5 | 126 | 7722 (5836–10141) | <1.5 | 113 | 7887 (6556–9642) | 0.789 |
| cardiovascular diseases | CAD + (CAD, HNT, MI) | 117 | 7791 (6423–10360) | CAD - | 145 | 8032 (6303–9848) | 0.889 |
| Genotype | RA Group | Control Group | p | ||
|---|---|---|---|---|---|
| N | Median (IQR) | N | Median (IQR) | ||
| KDR (+1416A/T) | |||||
| AA | 127 | 8766 (6802–10495) | 146 | 7479 (5297–9181) | 0.001 |
| AT | 96 | 7611 (6199–9457) | 97 | 7527 (5341–8963) | 0.512 |
| TT | 39 | 7365 (6042–9997) | 20 | 6997 (5322–9204) | 0.325 |
| KDR (+889G/A) | |||||
| GG | 124 | 7425 (5681–9539) | 205 | 7384 (5348–8973) | 0.398 |
| GA | 147 | 8264 (6875–10360) | 83 | 7371 (5469–8919) | 0.002 |
| AA | 1 | - | 1 | - | - |
| KDR (−604 T/C) | |||||
| TT | 68 | 8269 (6683–10043) | 17 | 8917 (7416–9312) | 0.934 |
| TC | 152 | 8410 (6511–10455) | 200 | 7336 (5341–8826) | 0.001 |
| CC | 51 | 6786 (5282–8568) | 72 | 6979 (5139–9288) | 0.900 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paradowska-Gorycka, A.; Stypinska, B.; Pawlik, A.; Malinowski, D.; Romanowska-Prochnicka, K.; Manczak, M.; Olesinska, M. KDR (VEGFR2) Genetic Variants and Serum Levels in Patients with Rheumatoid Arthritis. Biomolecules 2019, 9, 355. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9080355
Paradowska-Gorycka A, Stypinska B, Pawlik A, Malinowski D, Romanowska-Prochnicka K, Manczak M, Olesinska M. KDR (VEGFR2) Genetic Variants and Serum Levels in Patients with Rheumatoid Arthritis. Biomolecules. 2019; 9(8):355. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9080355
Chicago/Turabian StyleParadowska-Gorycka, Agnieszka, Barbara Stypinska, Andrzej Pawlik, Damian Malinowski, Katarzyna Romanowska-Prochnicka, Malgorzata Manczak, and Marzena Olesinska. 2019. "KDR (VEGFR2) Genetic Variants and Serum Levels in Patients with Rheumatoid Arthritis" Biomolecules 9, no. 8: 355. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9080355