Proinflammatory Action of a New Electronegative Low-Density Lipoprotein Epitope

 , and
, and 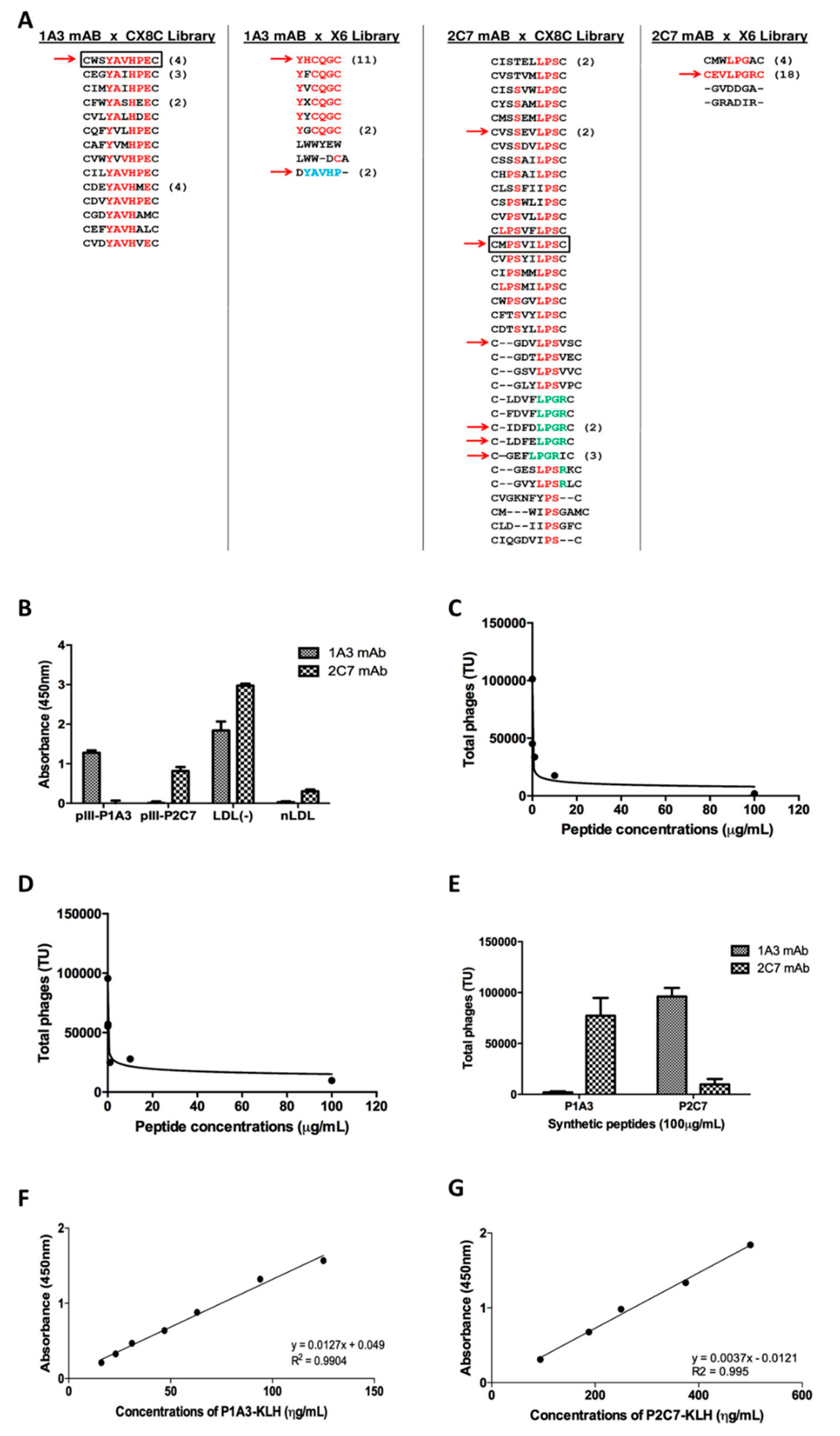{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Phage Display Random Peptide Libraries
2.3. Characterization of the Peptides by Small-Angle X-ray Scattering (SAXS)
2.4. Isolation of Human LDL (−)
2.5. Obtention of Bone Marrow-Derived Macrophages
2.6. Peptide Endocytosis by Macrophages
2.7. Macrophage Activation
2.8. Evaluation of BMDM Polarization by Flow Cytometry
2.9. Cytokine Quantification by Cytometric Bead Array (CBA) Kit
2.10. Nitric Oxide Quantification (NOA)
2.11. Statistical Analysis
2.12. Data Availability
2.13. Human and Animal Rights
3. Results
3.1. Mapping of LDL(-) Mimetic Epitopes
3.2. Structural Characterization of P1A3 and P2C7
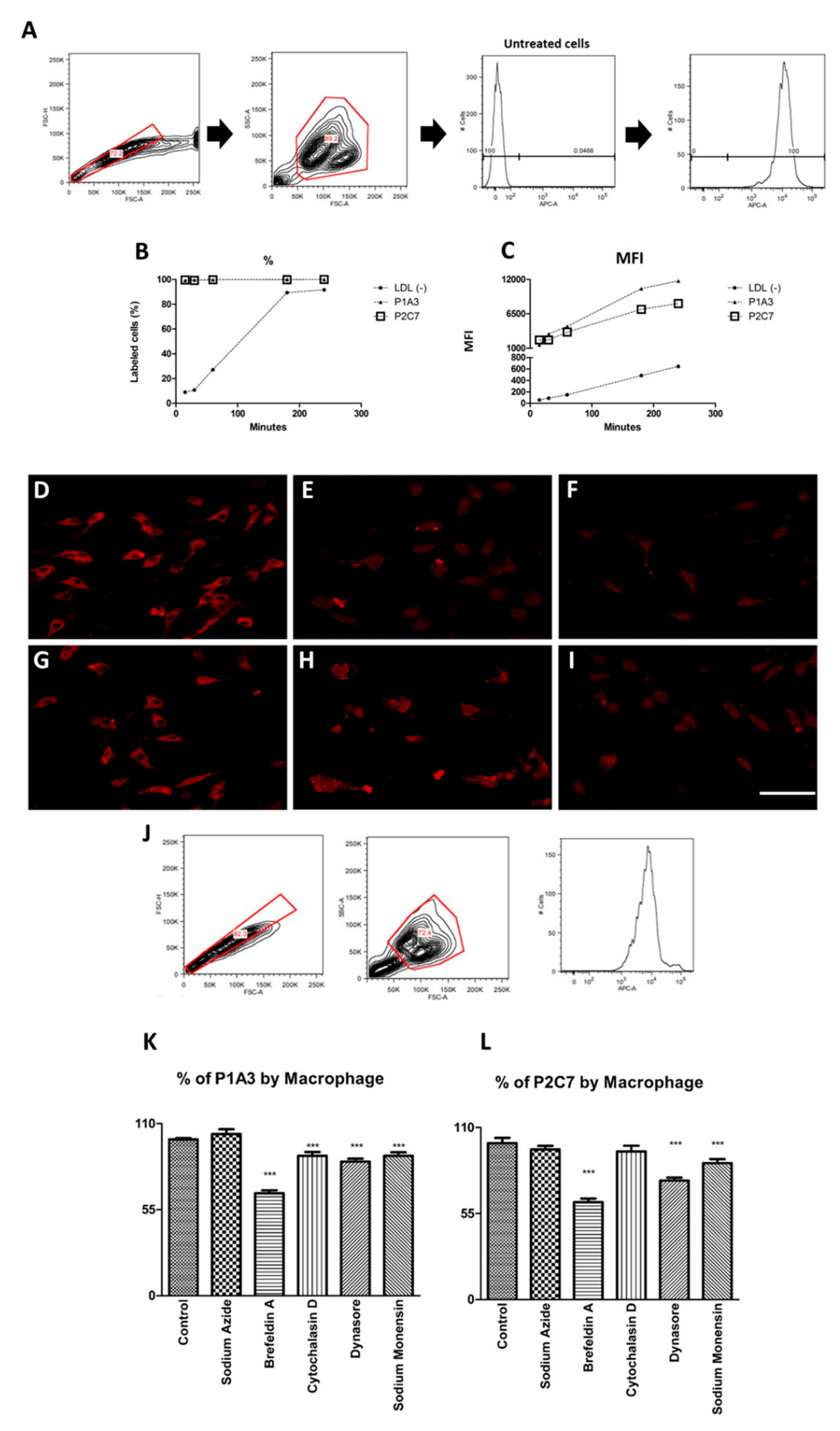
3.3. P1A3 and P2C7 Are Internalized by Macrophages
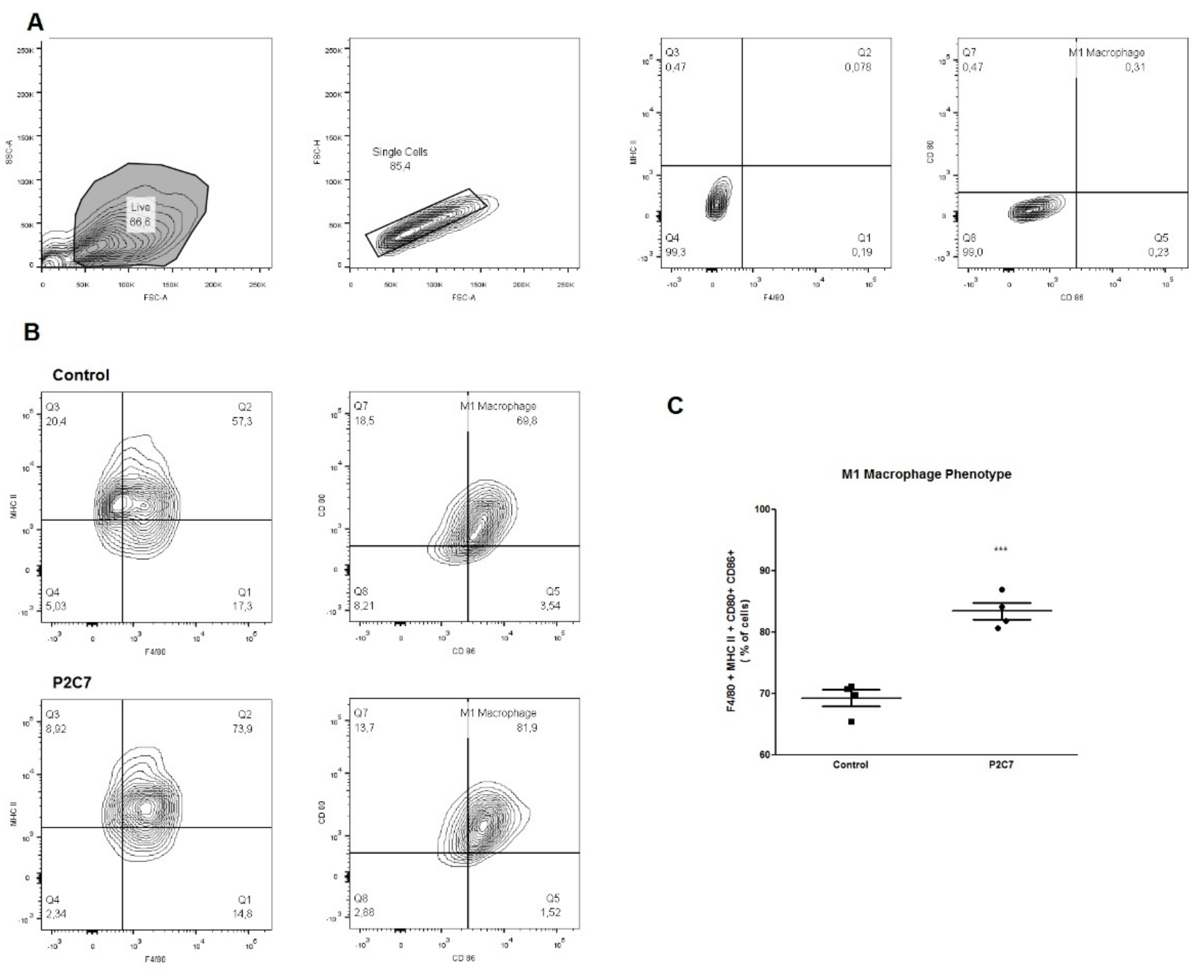
3.4. P2C7 Is a Proinflammatory Stimulus to Macrophages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weber, C.; Noels, H. Atherosclerosis: Current pathogenesis and therapeutic options. Nat. Med. 2011, 17, 1410–1422. [Google Scholar] [CrossRef] [PubMed]
- Eldika, N. Atherosclerosis as an inflammatory disease: Implications for therapy. Front. Biosci. 2004, 9, 2764. [Google Scholar] [CrossRef] [PubMed]
- Avogaro, P.; Bon, G.B.; Cazzolato, G. Presence of a modified low density lipoprotein in humans. Arteriosclerosis 1988, 8, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Mello, A.P.Q.; Da Silva, I.T.; Abdalla, D.S.P.; Damasceno, N.R.T. Electronegative low-density lipoprotein: Origin and impact on health and disease. Atherosclerosis 2011, 215, 257–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estruch, M.; Sánchez-Quesada, J.L.; Ordóñez Llanos, J.; Benítez, S. Electronegative LDL: A Circulating Modified LDL with a Role in Inflammation. Mediat. Inflamm. 2013, 2013, 181324. [Google Scholar] [CrossRef]
- De Castellarnau, C.; Sánchez-Quesada, J.L.; Benítez, S.; Rosa, R.; Caveda, L.; Vila, L.; Ordóñez-Llanos, J. Electronegative LDL from normolipemic subjects induces IL-8 and monocyte chemotactic protein secretion by human endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 2281–2287. [Google Scholar] [CrossRef]
- Abe, Y.; Fornage, M.; Yang, C.-Y.; Bui-Thanh, N.-A.; Wise, V.; Chen, H.-H.; Rangaraj, G.; Ballantyne, C.M. L5, the most electronegative subfraction of plasma LDL, induces endothelial vascular cell adhesion molecule 1 and CXC chemokines, which mediate mononuclear leukocyte adhesion. Atherosclerosis 2007, 192, 56–66. [Google Scholar] [CrossRef]
- Faulin, T.D.E.S.; Cavalcante, M.F.; Abdalla, D.S.P. Role of electronegative LDL and its associated antibodies in the pathogenesis of atherosclerosis. Clin. Lipidol. 2010, 5, 719–729. [Google Scholar] [CrossRef]
- Pedrosa, A.M.C.; Faine, L.A.; Grosso, D.M.; De Las Heras, B.; Boscá, L.; Abdalla, D.S.P. Electronegative LDL induction of apoptosis in macrophages: Involvement of Nrf2. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2010, 1801, 430–437. [Google Scholar] [CrossRef]
- Estruch, M.; Bancells, C.; Beloki, L.; Sanchez-Quesada, J.L.; Ordóñez-Llanos, J.; Benitez, S. CD14 and TLR4 mediate cytokine release promoted by electronegative LDL in monocytes. Atherosclerosis 2013, 229, 356–362. [Google Scholar] [CrossRef]
- Damasceno, N.R.T.; Sevanian, A.; Apolinário, E.; Oliveira, J.M.; Fernandes, I.; Abdalla, D.S.P. Detection of electronegative low-density lipoprotein (LDL−) in plasma and atherosclerotic lesions by monoclonal antibody-based immunoassays. Clin. Biochem. 2006, 39, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Damasceno, N.R.T.; Apolinário, E.; Flauzino, F.D.; Fernandes, I.; Abdalla, D.S.P. Soy isoflavones reduce electronegative low-density lipoprotein (LDL−) and anti-LDL− autoantibodies in experimental atherosclerosis. Eur. J. Nutr. 2007, 46, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Fredrikson, G.N.; Hedblad, B.; Berglund, G.; Alm, R.; Ares, M.; Cercek, B.; Chyu, K.-Y.; Shah, P.K.; Nilsson, J. Identification of Immune Responses Against Aldehyde-Modified Peptide Sequences in ApoB Associated with Cardiovascular Disease. Arter. Thromb. Vasc. Boil. 2003, 23, 872–878. [Google Scholar] [CrossRef] [PubMed]
- Ketelhuth, D.F.J.; Rios, F.J.O.; Wang, Y.; Liu, H.; Johansson, M.E.; Fredrikson, G.N.; Hedin, U.; Gidlund, M.; Nilsson, J.; Hansson, G.K.; et al. Identification of a Danger-Associated Peptide from Apolipoprotein B100 (ApoBDS-1) That Triggers Innate Proatherogenic Responses. Circulation 2011, 124, 2433–2443. [Google Scholar] [CrossRef] [PubMed]
- Hermansson, A.; Johansson, D.K.; Ketelhuth, D.F.; Andersson, J.; Zhou, X.; Hansson, G.K. Immunotherapy with Tolerogenic Apolipoprotein B-100–Loaded Dendritic Cells Attenuates Atherosclerosis in Hypercholesterolemic Mice. Circulation 2011, 123, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Kobiyama, K.; Winkels, H.; Tse, K.; Miller, J.; Vassallo, M.; Wolf, D.; Ryden, C.; Orecchioni, M.; Dileepan, T.; et al. Regulatory CD4+ T Cells Recognize Major Histocompatibility Complex Class II Molecule—Restricted Peptide Epitopes of Apolipoprotein, B. Circulation 2018, 138, 1130–1143. [Google Scholar] [CrossRef] [PubMed]
- Assinger, A.; Wang, Y.; Butler, L.M.; Hansson, G.K.; Yan, Z.Q.; Söderberg-Nauclér, C.; Ketelhuth, D.F.J. Apolipoprotein B100 danger-associated signal 1 (ApoBDS-1) triggers platelet activation and boosts platelet-leukocyte proinflammatory responses. Thromb. Haemost. 2014, 112, 332–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faulin, T.D.E.S.; De Sena, K.C.M.; Telles, A.E.R.; De Mattos Grosso, D.; Faulin, E.J.B.; Abdalla, D.S.P. Validation of a novel ELISA for measurement of electronegative low-density lipoprotein. Clin. Chem. Lab. Med. 2008, 46, 1769–1775. [Google Scholar] [CrossRef]
- Oliveira, J.A.; Sevanian, A.; Rodrigues, R.J.; Apolinário, E.; Abdalla, D.S.P. Minimally modified electronegative LDL and its autoantibodies in acute and chronic coronary syndromes. Clin. Biochem. 2006, 39, 708–714. [Google Scholar] [CrossRef]
- Grosso, D.M.; Ferderbar, S.; Wanschel, A.C.B.A.; Krieger, M.H.; Higushi, M.L.; Abdalla, D.S.P. Antibodies against electronegative LDL inhibit atherosclerosis in LDLr-/-mice. Braz. J. Med Biol. Res. 2008, 41, 1086–1092. [Google Scholar] [CrossRef]
- Kazuma, S.M.; Cavalcante, M.F.; Telles, A.E.; Maranhão, A.Q.; Abdalla, D.S. Cloning and expression of an anti-LDL(-) single-chain variable fragment, and its inhibitory effect on experimental atherosclerosis. mAbs 2013, 5, 763–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faulin, T.D.E.S.; Guilherme, D.F.; Silva, A.S.; Abdalla, D.S.P.; Hering, V.R.; Politi, M.J.; Maranhão, A.Q. GFP-SCFV: Expression and possible applications as a tool for experimental investigations of atherosclerosis. Biotechnol. Prog. 2014, 30, 1206–1213. [Google Scholar] [CrossRef] [PubMed]
- Cavalcante, M.F.; Kazuma, S.M.; Bender, E.A.; Adorne, M.D.; Ullian, M.; Veras, M.M.; Saldiva, P.H.N.; Maranhão, A.Q.; Guterres, S.S.; Pohlmann, A.R.; et al. A nanoformulation containing a scFv reactive to electronegative LDL inhibits atherosclerosis in LDL receptor knockout mice. Eur. J. Pharm. Biopharm. 2016, 107, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Michaloski, J.S.; Redondo, A.R.; Magalhães, L.S.; Cambui, C.C.; Giordano, R.J. Discovery of pan-VEGF inhibitory peptides directed to the extracellular ligand-binding domains of the VEGF receptors. Sci. Adv. 2016, 2, e1600611. [Google Scholar] [CrossRef] [PubMed]
- Beppler, J.; Ben Mkaddem, S.; Michaloski, J.; Honorato, R.V.; Velasco, I.T.; De Oliveira, P.S.L.; Giordano, R.J.; Monteiro, R.C.; Da Silva, F.P.; Oliveira, P.S.L. Negative regulation of bacterial killing and inflammation by two novel CD16 ligands. Eur. J. Immunol. 2016, 46, 1926–1935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, C.L.P. Investigating Macromolecular Complexes in Solution by Small Angle X-Ray Scattering. In Current Trends in X-Ray Crystallography; InTech: Luxembourg, 2011. [Google Scholar] [CrossRef] [Green Version]
- Weischenfeldt, J.; Porse, B. Bone Marrow-Derived Macrophages (BMM): Isolation and Applications; CSH Protocols: Cold Spring Harbor, NY, USA, 2008. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/21356739 (accessed on 1 June 2016).
- Bancells, C.; Canals, F.; Benítez, S.; Colomé, N.; Julve, J.; Ordóñez-Llanos, J.; Sánchez-Quesada, J.L. Proteomic analysis of electronegative low-density lipoprotein. J. Lipid Res. 2010, 51, 3508–3515. [Google Scholar] [CrossRef] [Green Version]
- Klausner, R.D.; Donaldson, J.G.; Lippincott-Schwartz, J. Brefeldin A: Insights into the control of membrane traffic and organelle structure. J. Cell Biol. 1992, 116, 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef]
- Medbury, H.J.; James, V.; Ngo, J.; Hitos, K.; Wang, Y.; Harris, D.C.; Fletcher, J.P. Differing association of macrophage subsets with atherosclerotic plaque stability. Int. Angiol. 2013, 32, 74–84. [Google Scholar]
- Mc Neill, E.; Crabtree, M.J.; Sahgal, N.; Patel, J.; Chuaiphichai, S.; Iqbal, A.J.; Hale, A.B.; Greaves, D.R.; Channon, K.M. Regulation of iNOS function and cellular redox state by macrophage Gch1 reveals specific requirements for tetrahydrobiopterin in NRF2 activation. Free Radic. Boil. Med. 2015, 79, 206–216. [Google Scholar] [CrossRef]
- Jablonski, K.A.; Amici, S.A.; Webb, L.M.; Ruiz-Rosado, J.D.D.; Popovich, P.G.; Partida-Sánchez, S.; Guerau-De-Arellano, M. Novel Markers to Delineate Murine M1 and M2 Macrophages. PLoS ONE 2015, 10, e0145342. [Google Scholar] [CrossRef] [PubMed]
- Nathan, C. Nitric oxide as a secretory product of mammalian cells. FASEB J. 1992, 6, 3051–3064. [Google Scholar] [CrossRef] [PubMed]
- Moncada, S.; Higgs, E.A. Molecular mechanisms and therapeutic strategies related to nitric oxide. FASEB J. 1995, 9, 1319–1330. [Google Scholar] [CrossRef] [PubMed]
- Singh, U.; Jialal, I. Oxidative stress and atherosclerosis. Pathophysiology 2006, 13, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Luoma, J.S.; Strålin, P.; Marklund, S.L.; Hiltunen, T.P.; Särkioja, T.; Ylä-Herttuala, S. Expression of extracellular SOD and iNOS in macrophages and smooth muscle cells in human and rabbit atherosclerotic lesions: Colocalization with epitopes characteristic of oxidized LDL and peroxynitrite-modified proteins. Arter. Thromb. Vasc. Boil. 1998, 18, 157–167. [Google Scholar] [CrossRef]
- Garlanda, C.; Dinarello, C.A.; Mantovani, A. The interleukin-1 family: Back to the future. Immunity 2013, 39, 1003–1018. [Google Scholar] [CrossRef]
- McDowell, T.L.; Symons, J.A.; Duff, G.W. Human interleukin-1α gene expression is regulated by Sp1 and a transcriptional repressor. Cytokine 2005, 30, 141–153. [Google Scholar] [CrossRef]
- McCarthy, D.A.; Ranganathan, A.; Subbaram, S.; Flaherty, N.L.; Patel, N.; Trebak, M.; Hempel, N.; Melendez, J.A. Redox-control of the alarmin, Interleukin-1α. Redox Biol. 2013, 1, 218–225. [Google Scholar] [CrossRef]
- Niemann-Jönsson, A.; Söderberg, I.; Lindholm, M.W.; Jovinge, S.; Nilsson, J.; Fredrikson, G.N. Medial Expression of TNF-α and TNF Receptors Precedes the Development of Atherosclerotic Lesions in Apolipoprotein E/LDL Receptor Double Knockout Mice. Int. J. Biomed. Sci. IJBS 2007, 3, 116–122. [Google Scholar]
- Hashizume, M.; Mihara, M. Blockade of IL-6 and TNF-α inhibited oxLDL-induced production of MCP-1 via scavenger receptor induction. Eur. J. Pharmacol. 2012, 689, 249–254. [Google Scholar] [CrossRef]
- Zhang, S.; Kim, C.C.; Batra, S.; McKerrow, J.H.; Loke, P. Delineation of Diverse Macrophage Activation Programs in Response to Intracellular Parasites and Cytokines. PLoS Negl. Trop. Dis. 2010, 4, e648. [Google Scholar] [CrossRef] [PubMed]
- Witztum, J.L.; Lichtman, A.H. The Influence of Innate and Adaptive Immune Responses on Atherosclerosis. Annu. Rev. Pathol. Mech. Dis. 2014, 9, 73–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, Y.I.; Tsimikas, S. Oxidation-specific epitopes as targets for biotheranostic applications in humans. Curr. Opin. Lipidol. 2013, 24, 426–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, Y.I.; Choi, S.-H.; Wiesner, P.; Fang, L.; Harkewicz, R.; Hartvigsen, K.; Boullier, A.; Gonen, A.; Diehl, C.J.; Que, X.; et al. Oxidation-Specific Epitopes are Danger Associated Molecular Patterns Recognized by Pattern Recognition Receptors of Innate Immunity. Circ. Res. 2011, 108, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Amir, S.; Hartvigsen, K.; Gonen, A.; Leibundgut, G.; Que, X.; Jensen-Jarolim, E.; Wagner, O.; Tsimikas, S.; Witztum, J.L.; Binder, C.J. Peptide mimotopes of malondialdehyde epitopes for clinical applications in cardiovascular disease. J. Lipid Res. 2012, 53, 1316–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honold, L.; Nahrendorf, M. Resident and monocyte-derived macrophages in cardiovascular disease. Circ. Res. 2018, 122, 113–127. [Google Scholar] [CrossRef]
- Bobryshev, Y.V.; Ivanova, E.A.; Chistiakov, D.A.; Nikiforov, N.G.; Orekhov, A.N. Macrophages and Their Role in Atherosclerosis: Pathophysiology and Transcriptome Analysis. BioMed Res. Int. 2016, 2016, 9582430. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faulin, T.d.E.S.; Kazuma, S.M.; Tripodi, G.L.; Cavalcante, M.F.; Wakasuqui, F.; Oliveira, C.L.P.; Degenhardt, M.F.d.S.; Michaloski, J.; Giordano, R.J.; Ketelhuth, D.F.J.; et al. Proinflammatory Action of a New Electronegative Low-Density Lipoprotein Epitope. Biomolecules 2019, 9, 386. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9080386
Faulin TdES, Kazuma SM, Tripodi GL, Cavalcante MF, Wakasuqui F, Oliveira CLP, Degenhardt MFdS, Michaloski J, Giordano RJ, Ketelhuth DFJ, et al. Proinflammatory Action of a New Electronegative Low-Density Lipoprotein Epitope. Biomolecules. 2019; 9(8):386. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9080386
Chicago/Turabian StyleFaulin, Tanize do Espirito Santo, Soraya Megumi Kazuma, Gustavo Luis Tripodi, Marcela Frota Cavalcante, Felipe Wakasuqui, Cristiano Luis Pinto Oliveira, Maximilia Frazão de Souza Degenhardt, Jussara Michaloski, Ricardo José Giordano, Daniel Francisco Jacon Ketelhuth, and et al. 2019. "Proinflammatory Action of a New Electronegative Low-Density Lipoprotein Epitope" Biomolecules 9, no. 8: 386. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9080386