
Improved Biogeography-Based Optimization Based on Affinity Propagation

Abstract
:1. Introduction
- This work proposed an improved BBO algorithm using the AP strategy to modify the migration operation to promote exploration and
- proposed a MBBO algorithm using the Memetic framework and SA as the local search strategy to promote exploitation.
2. Improved BBO with Affinity Propagation based on Memetic Framework
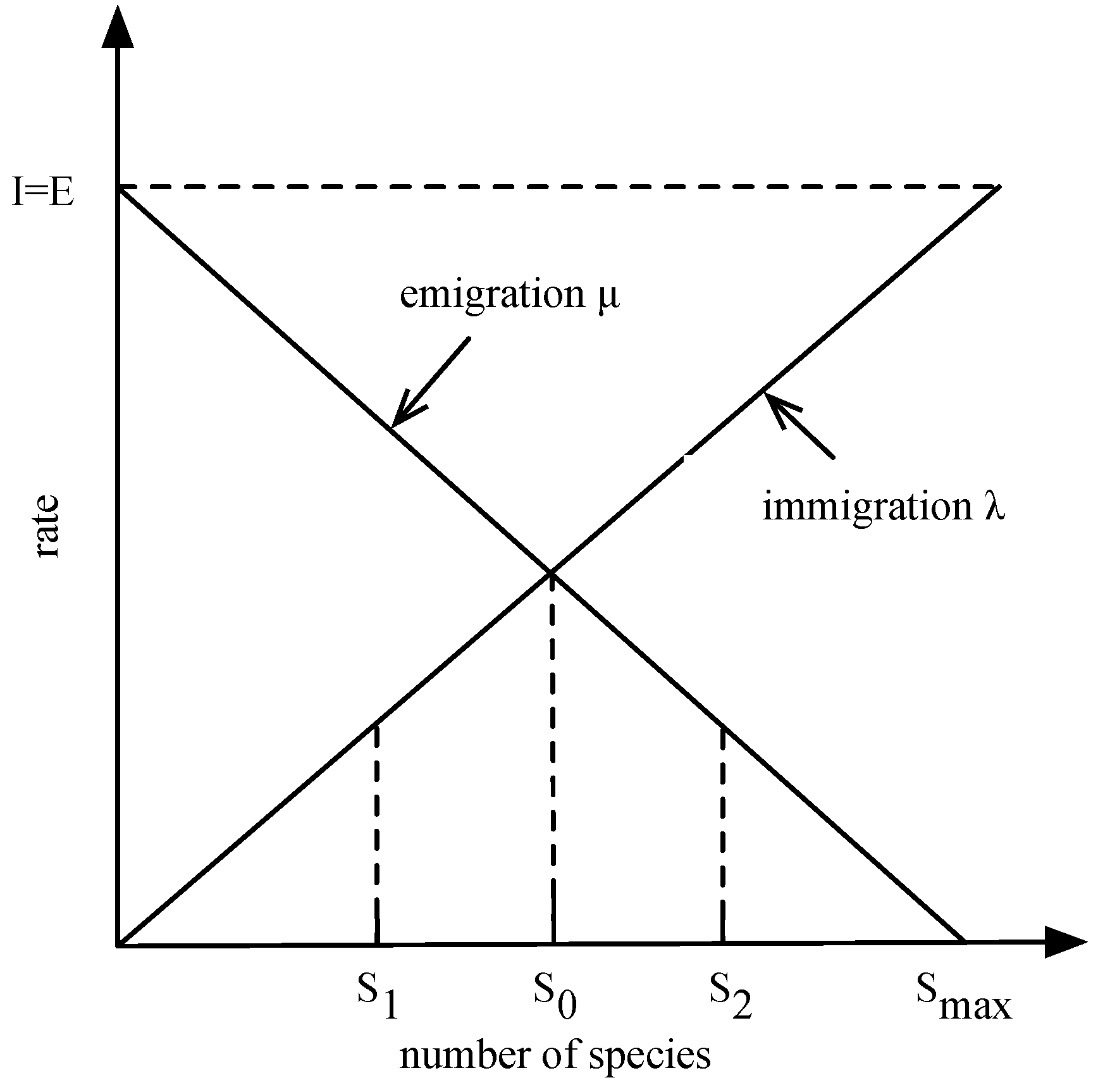
2.1. BBO
- At t time, there were S species in the habitat and no migration behavior in period Δt;
- At t time, there were S + 1 species in the habitat, with one species immigrating in period λs;
- At t time, there were S − 1 species in the habitat, with one species emigrating in period λs.
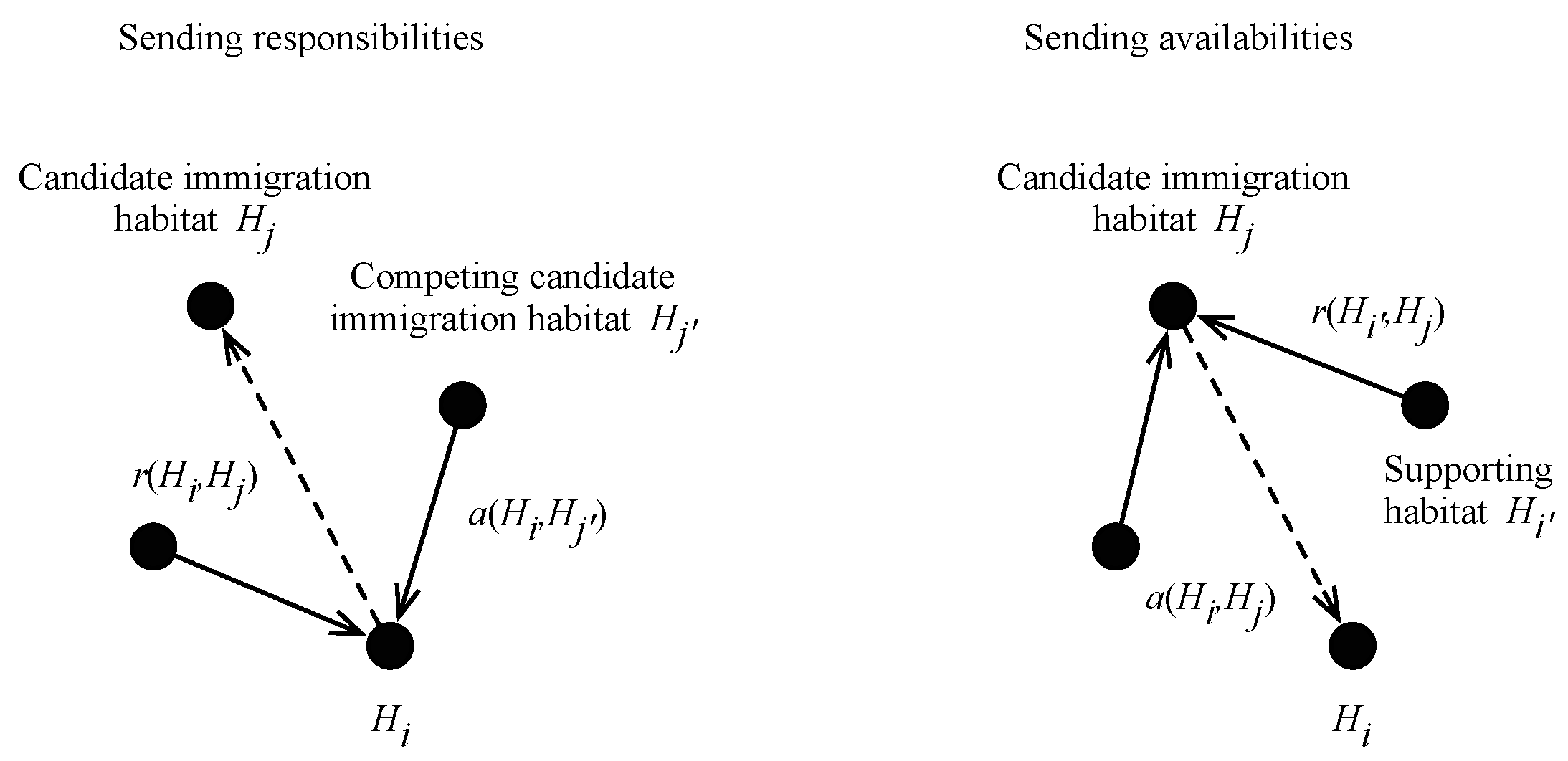
2.2. BBO with Affinity Propagation
2.3. Local Search Strategies
2.4. MBBO Algorithm
| Algorithm 1: MBBO |
|
3. Experimental Settings and Results
3.1. CEC 2015 Benchmarks
3.2. CEC 2015 Benchmarks Results and Analysis
3.2.1. Experiment Parameter Setting
3.2.2. Experiment Result and Analysis
3.2.3. Statistical Analysis
- MBBO algorithm and SPS-L-SHADE-EIG algorithm: A Wilcoxon signed-rank test showed that the MBBO algorithm did not elicit a statistically significant change in , F2 (z = −0.920, , and . In these four functions, the median change from MBBO algorithm to SPS-L-SHADE-EIG algorithm is not significantly different from zero. This suggests that with confidence, the difference between the algorithms is statistically significant for the remaining 11 functions.
- MBBO algorithm and DEsPA algorithm: A Wilcoxon signed-rank test showed that the MBBO algorithm did not elicit a statistically significant change in , F2 (z = −0.178, and with a 95 % confidence level. Therefore, the test has not provided statistically significant evidence that the two algorithms are different for these benchmark functions. For the other 12 cases, the median change from MBBO algorithm to DEsPA algorithm is very different from zero.
- MBBO algorithm and MVMO algorithm: A Wilcoxon signed-rank test showed that the MBBO algorithm did not elicit a statistically significant change in , F2 (z = −1.823, and . In these three functions, the median change from MBBO algorithm to SPS-L-SHADE-EIG algorithm is not significantly different from zero; for the other 12 cases, the two algorithms are different with a 95 % confidence level.
3.3. Effect of the HIR on the Performance of MBBO
3.3.1. Experiment Setting
3.3.2. Experimental Results and Analysis
3.4. Experimental Results Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- MacArthur, R.; Wilson, E. The Theory of Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Simon, D. Biogeography-based optimization. IEEE Trans. Evol. Comput. 2008, 12, 702–713. [Google Scholar] [CrossRef]
- Ma, H.; Simon, D. Analysis of migration models of biogeography-based optimization using Markov theory. Eng. Appl. Artif. Intell. 2011, 24, 1052–1060. [Google Scholar] [CrossRef]
- Ma, H.; Simon, D.; Fei, M.; Xie, Z. Variations of biogeography-based optimization and Markov analysis. Inf. Sci. 2013, 220, 492–506. [Google Scholar] [CrossRef]
- Yang, G.P.; Liu, S.Y.; Zhang, J.K.; Feng, Q.X. Control and synchronization of chaotic systems by an improved biogeography-based optimization algorithm. Appl. Intell. 2013, 39, 132–143. [Google Scholar] [CrossRef]
- Gong, W.; Cai, Z.; Ling, C.X.; Li, H. A real-coded biogeography-based optimization with mutation. Appl. Math. Comput. 2010, 216, 2749–2758. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Chattopadhyay, P. Hybrid Differential Evolution With Biogeography-Based Optimization for Solution of Economic Load Dispatch. IEEE Trans. Power Syst. 2010, 25, 1955–1964. [Google Scholar] [CrossRef]
- Frey, B.J.; Dueck, D. Clustering by passing messages between data points. Science 2007, 315, 972–976. [Google Scholar] [CrossRef] [PubMed]
- Moscato, P.; Norman, M.G. A memetic approach for the traveling salesman problem implementation of a computational ecology for combinatorial optimization on message-passing systems. Parallel Comput. Transput. Appl. 1992, 1, 177–186. [Google Scholar]
- Merz, P.; Zell, A. Clustering gene expression profiles with memetic algorithms. In Parallel Problem Solving from Nature-PPSN VII; Springer: Berlin, Germany, 2002; pp. 811–820. [Google Scholar]
- Kannan, S.S.; Ramaraj, N. A novel hybrid feature selection via Symmetrical Uncertainty ranking based local memetic search algorithm. Knowl. Based Syst. 2010, 23, 580–585. [Google Scholar] [CrossRef]
- Banos, R.; Gil, C.; Reca, J.; Montoya, F. A memetic algorithm applied to the design of water distribution networks. Appl. Soft Comput. 2010, 10, 261–266. [Google Scholar] [CrossRef]
- Kirkpatrick, S.; Gelatt, C.D.; Vecchi, M.P. Optimization by simulated annealing. Science 1983, 220, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.M.; Tsai, J.S.H.; Yang, C.C.; Hsu, P.H. A self-optimization approach for L-SHADE incorporated with eigenvector-based crossover and successful-parent-selecting framework on CEC 2015 benchmark set. In Proceedings of the 2015 IEEE Congress on Evolutionary Computation, Sendai, Japan, 25–28 May 2015.
- Awad, N.; Ali, M.Z.; Reynolds, R.G. A differential evolution algorithm with success-based parameter adaptation for CEC2015 learning-based optimization. In Proceedings of the 2015 IEEE Congress on Evolutionary Computation, Sendai, Japan, 25–28 May 2015; pp. 1098–1105.
- Rueda, J.L.; Erlich, I. Testing MVMO on learning-based real-parameter single objective benchmark optimization problems. In Proceedings of the 2015 IEEE Congress on Evolutionary Computation (CEC), Sendai, Japan, 25–28 May 2015.
- Wesche, T.A.; Goertler, C.M.; Hubert, W.A. Modified habitat suitability index model for brown trout in southeastern Wyoming. N. Am. J. Fish. Manag. 1987, 7, 232–237. [Google Scholar] [CrossRef]
- Hanski, I.; Gilpin, M.E. Metapopulation Biology; Academic Press: Waltham, MA, USA, 1997. [Google Scholar]
- Liang, J.; Qu, B.; Suganthan, P.; Chen, Q. Problem Definitions and Evaluation Criteria for the CEC 2015 Competition on Learning-Based Real-Parameter Single Objective Optimization; Technical Report 201411A; Computational Intelligence Laboratory, Zhengzhou University: Zhengzhou, China; Nanyang Technological University: Singapore, 2014. [Google Scholar]
- Tanabe, R.; Fukunaga, A. Success-history based parameter adaptation for differential evolution. In Proceedings of the 2013 IEEE Congress on Evolutionary Computation, Cancun, Mexico, 20–23 June 2013; pp. 71–78.
- Tanabe, R.; Fukunaga, A. Evaluating the performance of SHADE on CEC 2013 benchmark problems. In Proceedings of the 2013 IEEE Congress on Evolutionary Computation, Cancun, Mexico, 20–23 June 2013; pp. 1952–1959.
- Tanabe, R.; Fukunaga, A.S. Improving the search performance of SHADE using linear population size reduction. In Proceedings of the 2014 IEEE Congress on Evolutionary Computation (CEC), Beijing, China, 6–11 July 2014; pp. 1658–1665.
- Guo, S.M.; Yang, C.C. Enhancing differential evolution utilizing eigenvector-based crossover operator. IEEE Trans. Evol. Comput. 2015, 19, 31–49. [Google Scholar]
- Guo, S.M.; Yang, C.C.; Hsu, P.H.; Tsai, J.S.H. Improving Differencial Evolution with a Successful Parent Selecting Framework. IEEE Trans. Evol. Comput. 2015, 19, 717–730. [Google Scholar] [CrossRef]
- Zhang, J.; Sanderson, A.C. JADE: adaptive differential evolution with optional external archive. IEEE Trans. Evol. Comput. 2009, 13, 945–958. [Google Scholar] [CrossRef]
- Holland, J.H. Building blocks, cohort genetic algorithms, and hyperplane-defined functions. Evol. Comput. 2000, 8, 373–391. [Google Scholar] [CrossRef] [PubMed]
- Booker, L.B.; Goldberg, D.E.; Holland, J.H. Classifier systems and genetic algorithms. Artif. Intell. 1989, 40, 235–282. [Google Scholar] [CrossRef]
- Kennedy, J. Particle swarm optimization. In Encyclopedia of machine learning; Springer: New York, NY, USA, 2011; pp. 760–766. [Google Scholar]
- Shi, Y.; Eberhart, R. A modified particle swarm optimizer. In Proceedings of the 1998 IEEE International Conference on Evolutionary Computation, Anchorage, AK, USA, 4–9 May 1998; pp. 69–73.
- Reynolds, R.G. An introduction to cultural algorithms. In Proceedings of the Third Annual Conference on Evolutionary Programming, Singapore, 24–26 February 19949.




{kind=link}
{kind=link}
{kind=link}
| Test Functions | ||
|---|---|---|
| Unimodal Functions | ||
| F1 | Rotated High Conditioned Elliptic Function | 100 |
| F2 | Rotated Cigar Function | 200 |
| Simple Multimodal Functions | ||
| F3 | Shifted and Rotated Ackley’s Function | 300 |
| F4 | Shifted and Rotated Rastrigin’s Function | 400 |
| F5 | Shifted and Rotated Schwefel’s Function | 500 |
| Hybrid Functions | ||
| F6 | Hybrid Function 1 (N = 3) | 600 |
| F7 | Hybrid Function 2 (N = 4) | 700 |
| F8 | Hybrid Function 3(N = 5) | 800 |
| Composition Functions | ||
| F9 | Composition Function 1 (N = 3) | 900 |
| F10 | Composition Function 2 (N = 3) | 1000 |
| F11 | Composition Function 3 (N = 5) | 1100 |
| F12 | Composition Function 4 (N = 5) | 1200 |
| F13 | Composition Function 5 (N = 5) | 1300 |
| F14 | Composition Function 6 (N = 7) | 1400 |
| F15 | Composition Function 7 (N = 10) | 1500 |
| Search Range: | ||
| MBBO | SPS-L-SHADE-EIG | DEsPA | MVMO | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Best | Median | Mean | Distance | Best | Median | Mean | Distance | Best | Median | Mean | Distance | Best | Median | Mean | Distance | ||||
| 10-D | |||||||||||||||||||
| F1 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | |||
| F2 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | |||
| F3 | 1.10E+01 | 1.50E+01 | 1.70E+01 | 1.10E+01 | 2.00E+01 | 2.20E+01 | 2.60E+01 | 2.00E+01 | 1.91E+01 | 2.19E+01 | 1.90E+01 | 1.91E+01 | 2.01E+01 | 2.33E+01 | 1.40E+01 | 2.01E+01 | |||
| F4 | 1.34E-01 | 6.25E-01 | 1.23E+00 | 1.34E-01 | 7.83E-01 | 8.72E-01 | 1.95E+00 | 7.83E-01 | 8.93E-01 | 9.95E-01 | 1.17E+00 | 8.93E-01 | 9.83E-01 | 1.23E+00 | 9.76E-01 | 9.83E-01 | |||
| F5 | 9.93E+00 | 1.34E+01 | 2.14E+01 | 9.93E+00 | 1.24E+01 | 1.64E+01 | 2.35E+01 | 1.24E+01 | 1.43E+01 | 1.86E+01 | 2.56E+01 | 1.43E+01 | 2.19E+01 | 2.35E+01 | 2.98E+01 | 2.19E+01 | |||
| F6 | 0.00E+00 | 0.00E+00 | 1.01E-01 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 1.23E-01 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 8.79E-01 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 7.98E-01 | 0.00E+00 | |||
| F7 | 6.70E-03 | 1.02E-02 | 2.23E-02 | 6.70E-03 | 9.76E-03 | 1.94E-02 | 2.78E-02 | 9.76E-03 | 1.23E-02 | 2.31E-02 | 3.21E-02 | 1.23E-02 | 2.35E-02 | 3.12E-02 | 5.37E-02 | 2.35E-02 | |||
| F8 | 7.63E-05 | 1.26E-04 | 7.32E-04 | 7.63E-05 | 1.01E-04 | 1.37E-04 | 8.81E-04 | 1.01E-04 | 2.13E-04 | 4.34E-04 | 9.86E-04 | 2.13E-04 | 3.98E-04 | 6.98E-04 | 9.98E-04 | 3.98E-04 | |||
| F9 | 2.39E+01 | 8.69E+01 | 9.51E+01 | 2.39E+01 | 6.92E+01 | 1.00E+02 | 1.50E+02 | 6.92E+01 | 9.21E+01 | 1.02E+02 | 1.02E+02 | 9.21E+01 | 1.02E+02 | 1.97E+02 | 1.28E+02 | 1.02E+02 | |||
| F10 | 2.03E+02 | 2.21E+02 | 2.04E+02 | 2.03E+02 | 1.98E+02 | 3.68E+02 | 3.53E+02 | 1.98E+02 | 2.58E+02 | 2.76E+02 | 2.37E+02 | 2.58E+02 | 2.98E+02 | 3.97E+02 | 3.15E+02 | 2.98E+02 | |||
| F11 | 3.28E-02 | 7.52E-02 | 4.37E-01 | 3.28E-02 | 7.82E-02 | 9.79E-02 | 9.27E-01 | 7.82E-02 | 8.27E-02 | 9.86E-01 | 7.78E-01 | 8.27E-02 | 5.24E-02 | 1.00E-01 | 3.33E-01 | 5.24E-02 | |||
| F12 | 2.39E+01 | 8.96E+01 | 1.00E+02 | 2.39E+01 | 9.82E+01 | 1.32E+02 | 1.06E+02 | 9.82E+01 | 1.92E+02 | 2.94E+02 | 2.36E+02 | 1.92E+02 | 2.01E+02 | 2.96E+02 | 2.75E+02 | 2.01E+02 | |||
| F13 | 3.30E-03 | 1.23E-02 | 2.12E-02 | 3.30E-03 | 3.46E-02 | 8.26E-02 | 2.31E-02 | 3.46E-02 | 8.72E-02 | 9.26E-02 | 7.89E-02 | 8.72E-02 | 2.05E-02 | 2.83E-02 | 7.23E-02 | 2.05E-02 | |||
| F14 | 1.03E+02 | 1.98E+02 | 3.94E+02 | 1.03E+02 | 1.95E+02 | 2.35E+02 | 5.21E+02 | 1.95E+02 | 4.32E+02 | 8.92E+02 | 7.26E+02 | 4.32E+02 | 3.95E+02 | 4.77E+02 | 5.67E+02 | 3.95E+02 | |||
| F15 | 3.61E+01 | 8.70E+01 | 9.50E+01 | 3.61E+01 | 9.32E+01 | 1.02E+02 | 1.67E+02 | 9.32E+01 | 1.01E+02 | 1.36E+02 | 2.34E+02 | 1.01E+02 | 2.51E+02 | 3.55E+02 | 2.45E+02 | 2.51E+02 | |||
| 50-D | |||||||||||||||||||
| F1 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | |||
| F2 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 0.00E+00 | |||
| F3 | 1.09E+01 | 1.83E+01 | 1.93E+01 | 1.09E+01 | 2.91E+01 | 3.22E+01 | 2.32E+01 | 2.91E+01 | 2.81E+01 | 3.11E+01 | 2.82E+01 | 2.81E+01 | 4.15E+01 | 6.51E+01 | 2.82E+01 | 4.15E+01 | |||
| F4 | 9.23E-01 | 1.23E+00 | 1.98E+00 | 9.23E-01 | 1.02E+00 | 3.52E+00 | 2.55E+00 | 1.02E+00 | 1.92E+00 | 2.46E+00 | 3.26E+00 | 1.92E+00 | 2.24E+00 | 3.86E+00 | 4.62E+00 | 2.24E+00 | |||
| F5 | 1.70E+03 | 1.92E+03 | 1.87E+03 | 1.70E+03 | 1.53E+03 | 2.98E+03 | 1.75E+03 | 1.53E+03 | 2.42E+03 | 2.99E+03 | 2.68E+03 | 2.42E+03 | 2.73E+03 | 3.82E+03 | 3.00E+03 | 2.73E+03 | |||
| F6 | 1.35E+02 | 1.98E+02 | 1.75E+02 | 1.35E+02 | 1.87E+02 | 2.11E+02 | 2.39E+02 | 1.87E+02 | 2.06E+02 | 2.67E+02 | 2.49E+02 | 2.06E+02 | 2.94E+02 | 3.22E+02 | 3.15E+02 | 2.94E+02 | |||
| F7 | 2.31E+01 | 3.92E+01 | 2.56E+01 | 2.31E+01 | 3.52E+01 | 4.05E+01 | 2.74E+01 | 3.52E+01 | 3.94E+01 | 4.96E+01 | 3.16E+01 | 3.94E+01 | 4.52E+01 | 5.81E+01 | 4.87E+01 | 4.52E+01 | |||
| F8 | 9.23E+00 | 2.16E+01 | 4.58E+01 | 9.23E+00 | 1.81E+01 | 2.79E+01 | 5.57E+01 | 1.81E+01 | 2.02E+01 | 2.62E+01 | 6.58E+01 | 2.02E+01 | 3.31E+01 | 6.32E+01 | 5.18E+01 | 3.31E+01 | |||
| F9 | 7.20E+01 | 1.06E+02 | 1.02E+02 | 7.20E+01 | 1.36E+02 | 2.56E+02 | 2.37E+02 | 1.36E+02 | 1.84E+02 | 3.35E+02 | 3.02E+00 | 1.84E+02 | 4.89E+02 | 6.51E+02 | 5.56E+02 | 4.89E+02 | |||
| F10 | 3.29E+02 | 7.95E+02 | 6.87E+02 | 3.29E+02 | 7.23E+02 | 8.09E+02 | 8.35E+02 | 7.23E+02 | 7.84E+02 | 9.86E+02 | 8.76E+02 | 7.84E+02 | 5.91E+02 | 6.98E+02 | 7.92E+02 | 5.91E+02 | |||
| F11 | 1.36E+02 | 2.78E+02 | 2.69E+02 | 1.36E+02 | 2.47E+02 | 3.00E+02 | 3.00E+02 | 2.47E+02 | 2.84E+02 | 3.76E+02 | 3.56E+02 | 2.84E+02 | 3.34E+02 | 5.87E+02 | 4.98E+02 | 3.34E+02 | |||
| F12 | 4.50E+01 | 1.00E+02 | 1.10E+02 | 4.50E+01 | 9.32E+01 | 1.04E+02 | 1.23E+02 | 9.32E+01 | 1.07E+02 | 1.98E+02 | 1.79E+02 | 1.07E+02 | 2.04E+02 | 3.01E+02 | 2.96E+02 | 2.04E+02 | |||
| F13 | 1.32E-02 | 6.54E-02 | 5.77E-02 | 1.32E-02 | 3.73E-02 | 7.57E-02 | 7.62E-02 | 3.73E-02 | 6.25E-02 | 8.27E-02 | 7.26E-02 | 6.25E-02 | 6.13E-02 | 7.60E-02 | 6.67E-02 | 6.13E-02 | |||
| F14 | 3.47E+04 | 5.98E+04 | 4.79E+04 | 3.47E+04 | 5.95E+04 | 6.83E+04 | 6.35E+04 | 5.95E+04 | 5.74E+04 | 7.89E+04 | 7.12E+04 | 5.74E+04 | 6.95E+04 | 8.63E+04 | 8.01E+04 | 6.95E+04 | |||
| F15 | 3.72E+01 | 9.60E+01 | 1.01E+02 | 3.72E+01 | 1.03E+02 | 1.56E+02 | 1.35E+02 | 1.03E+02 | 1.62E+02 | 2.32E+02 | 2.17E+02 | 1.62E+02 | 3.74E+02 | 6.19E+02 | 5.97E+02 | 3.74E+02 | |||
| MBBO and SPS-L-SHADE-EIG | MBBO and DEsPA | MBBO and MVMO | ||||||
|---|---|---|---|---|---|---|---|---|
| z | p-Value (2-Tailed) | z | p-Value (2-Tailed) | z | p-Value (2-Tailed) | |||
| F1 | −0.367 | 0.713 | −1.547 | 0.122 | −1.249 | 0.212 | ||
| F2 | −0.920 | 0.357 | −0.178 | 0.859 | −1.823 | 0.068 | ||
| F3 | −2.077 | 0.038 | −2.505 | 0.012 | −2.716 | 0.007 | ||
| F4 | −1.134 | 0.254 | −1.932 | 0.061 | −1.899 | 0.058 | ||
| F5 | −2.139 | 0.032 | −2.660 | 0.008 | −2.634 | 0.008 | ||
| F6 | −3.696 | 0.000 | −3.705 | 0.000 | −3.702 | 0.000 | ||
| F7 | −2.996 | 0.003 | −2.820 | 0.004 | −2.492 | 0.013 | ||
| F8 | −2.323 | 0.025 | −2.432 | 0.014 | −2.814 | 0.005 | ||
| F9 | −2.928 | 0.003 | −2.413 | 0.015 | −3.203 | 0.001 | ||
| F10 | −3.222 | 0.001 | −2.995 | 0.003 | −3.220 | 0.001 | ||
| F11 | −2.746 | 0.006 | −2.395 | 0.017 | −2.466 | 0.014 | ||
| F12 | −2.640 | 0.008 | −3.324 | 0.001 | −5.244 | 0.000 | ||
| F13 | −5.376 | 0.000 | −4.278 | 0.000 | −4.437 | 0.000 | ||
| F14 | −2.934 | 0.003 | −2.666 | 0.008 | −2.936 | 0.003 | ||
| F15 | −1.090 | 0.091 | −2.936 | 0.003 | −6.232 | 0.000 | ||
| MBBO () | MBBO | MBBO | MBBO | MBBO | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Std. | Mean | Std. | Mean | Std. | Mean | Std. | Mean | Std. | |
| F1 | 1.78E+00 | 6.27E-01 | 2.30E+00 | 9.89E-01 | 2.13E+00 | 8.51E-01 | 2.84E+00 | 7.58E-01 | 2.59E+00 | 1.26E+00 |
| F2 | 4.68E+03 | 1.62E+03 | 4.76E+03 | 1.39E+03 | 4.93E+03 | 1.33E+03 | 5.19E+03 | 1.34E+03 | 4.82E+03 | 1.92E+03 |
| F3 | 4.11E+01 | 8.11E+00 | 4.84E+01 | 1.04E+01 | 4.45E+01 | 9.63E+00 | 4.13E+01 | 1.02E+01 | 4.41E+01 | 1.15E+01 |
| F4 | 5.91E+00 | 1.56E+00 | 7.56E+00 | 2.31E+00 | 6.37E+00 | 1.14E+00 | 7.27E+00 | 1.60E+00 | 7.03E+00 | 1.69E+00 |
| F5 | 9.97E+01 | 2.91E+01 | 1.14E+02 | 3.65E+01 | 1.13E+02 | 4.02E+01 | 1.14E+02 | 2.50E+01 | 1.18E+02 | 3.82E+01 |
| F6 | 6.04E+02 | 2.63E+02 | 8.86E+02 | 3.03E+02 | 7.77E+02 | 2.65E+02 | 1.02E+03 | 4.75E+02 | 1.06E+03 | 4.16E+02 |
| F7 | 4.65E-02 | 3.60E-02 | 1.07E-01 | 1.14E-01 | 7.23E-02 | 6.15E-02 | 1.13E-01 | 8.00E-02 | 9.99E-02 | 1.11E-01 |
| F8 | 7.31E+02 | 2.09E+02 | 8.49E+02 | 2.21E+02 | 8.74E+02 | 2.33E+02 | 9.65E+02 | 2.34E+02 | 8.67E+02 | 2.79E+02 |
| F9 | 3.02E+01 | 6.21E+00 | 3.45E+01 | 5.78E+00 | 3.16E+01 | 5.81E+00 | 3.46E+01 | 7.08E+00 | 3.45E+01 | 6.64E+00 |
| F10 | 7.26E+00 | 1.09E+00 | 8.47E+00 | 9.48E-01 | 7.66E+00 | 9.36E-01 | 8.22E+00 | 1.34E+00 | 8.33E+00 | 1.01E+00 |
| F11 | 7.22E+00 | 2.41E+00 | 9.63E+00 | 2.82E+00 | 7.70E+00 | 2.93E+00 | 1.04E+01 | 3.11E+00 | 9.70E+00 | 3.79E+00 |
| F12 | 6.18E+04 | 2.72E+04 | 6.92E+04 | 2.41E+04 | 6.39E+04 | 1.97E+04 | 7.28E+04 | 3.49E+04 | 6.42E+04 | 1.91E+04 |
| F13 | 2.13E+00 | 4.68E-01 | 3.52E+00 | 3.14E-01 | 3.40E+00 | 4.55E-01 | 6.23E+02 | 5.39E+02 | 3.23E+03 | 2.41E+03 |
| F14 | 6.40E+01 | 1.47E+01 | 2.77E+02 | 1.03E+02 | 1.71E+02 | 2.01E+02 | 4.32E+02 | 2.43E+01 | 5.62E+02 | 3.31E+02 |
| F15 | 1.02E+02 | 7.31E+00 | 1.07E+02 | 9.03E+00 | 1.23E+02 | 1.46E+01 | 5.11E+03 | 3.21E+03 | 6.42E+04 | 1.91E+04 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Liu, P.; Ren, M.; Yang, Y.; Tian, X. Improved Biogeography-Based Optimization Based on Affinity Propagation. ISPRS Int. J. Geo-Inf. 2016, 5, 129. https://0-doi-org.brum.beds.ac.uk/10.3390/ijgi5080129
Wang Z, Liu P, Ren M, Yang Y, Tian X. Improved Biogeography-Based Optimization Based on Affinity Propagation. ISPRS International Journal of Geo-Information. 2016; 5(8):129. https://0-doi-org.brum.beds.ac.uk/10.3390/ijgi5080129
Chicago/Turabian StyleWang, Zhihao, Peiyu Liu, Min Ren, Yuzhen Yang, and Xiaoyan Tian. 2016. "Improved Biogeography-Based Optimization Based on Affinity Propagation" ISPRS International Journal of Geo-Information 5, no. 8: 129. https://0-doi-org.brum.beds.ac.uk/10.3390/ijgi5080129