Distribution Pattern of Red Fox (Vulpes vulpes) Dens and Spatial Relationships with Sea Turtle Nests, Recreation, and Environmental Characteristics



Abstract
:1. Introduction
- Are fox den entrances clustered, random, or evenly distributed across Masonboro Island? If they are clustered, are there particular hot spots? Is the pattern consistent from year to year?
- Are the sea turtle nests clustered, random, or evenly distributed? Is there a difference in the spatial pattern for nests that have been predated vs. not predated?
- Is there a spatial relationship between the fox den entrances and sea turtle nests?
- Is there a spatial relationship between the fox den entrances and the location of recreational activity?
- Are the locations of fox den entrances related to the topography of the island?
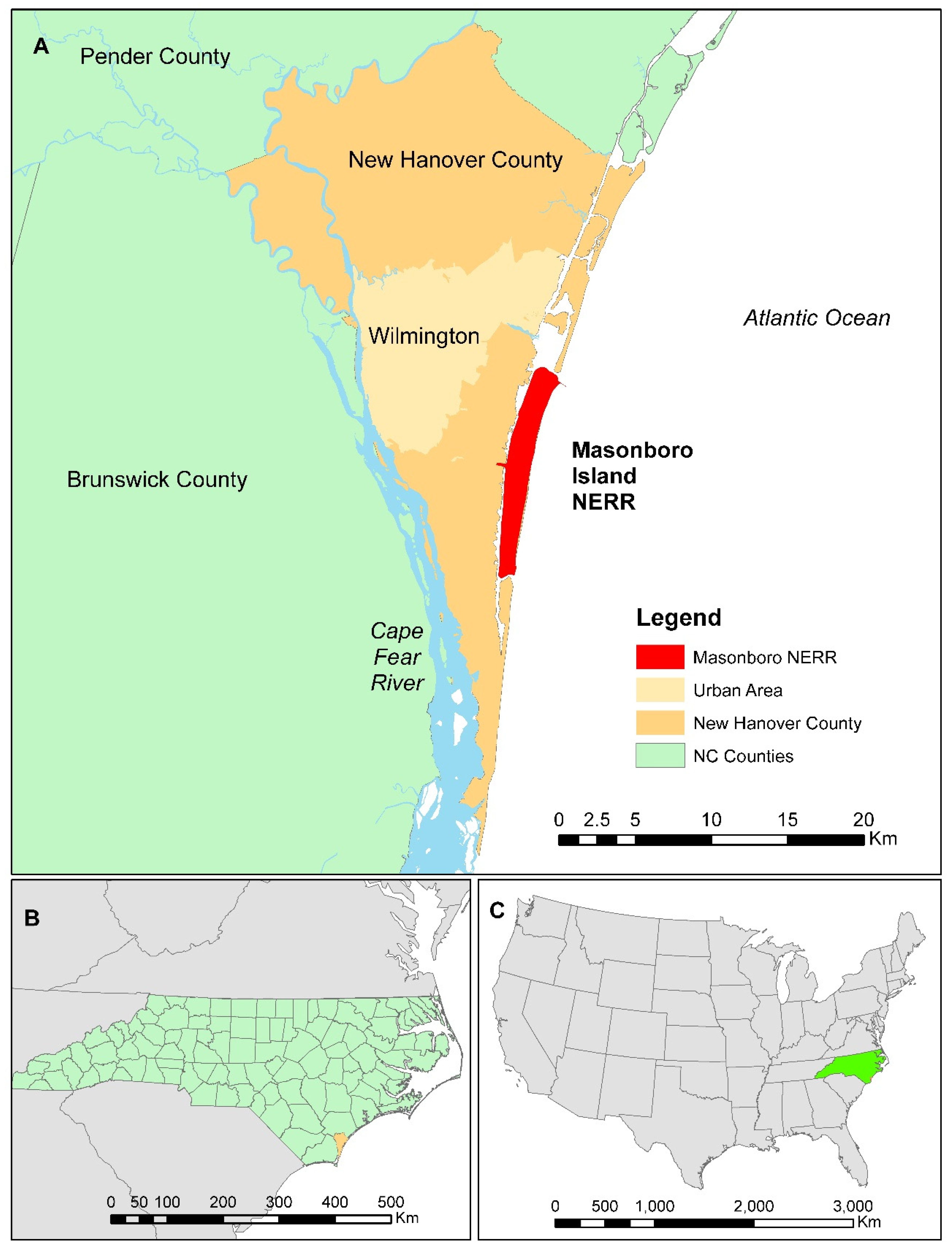
1.1. Masonboro Island Study Area
2. Methods
2.1. Field Work
2.2. Data Processing
2.3. Spatial Analysis
2.3.1. Point Pattern Analysis
2.3.2. Spatial Relationships between Fox Den Entrances, Sea Turtle Nests, and Boat Access Sites
2.3.3. Island Topography and Fox Den Entrance Locations
2.3.4. Predictive Model for Fox Den Entrance Locations
3. Results
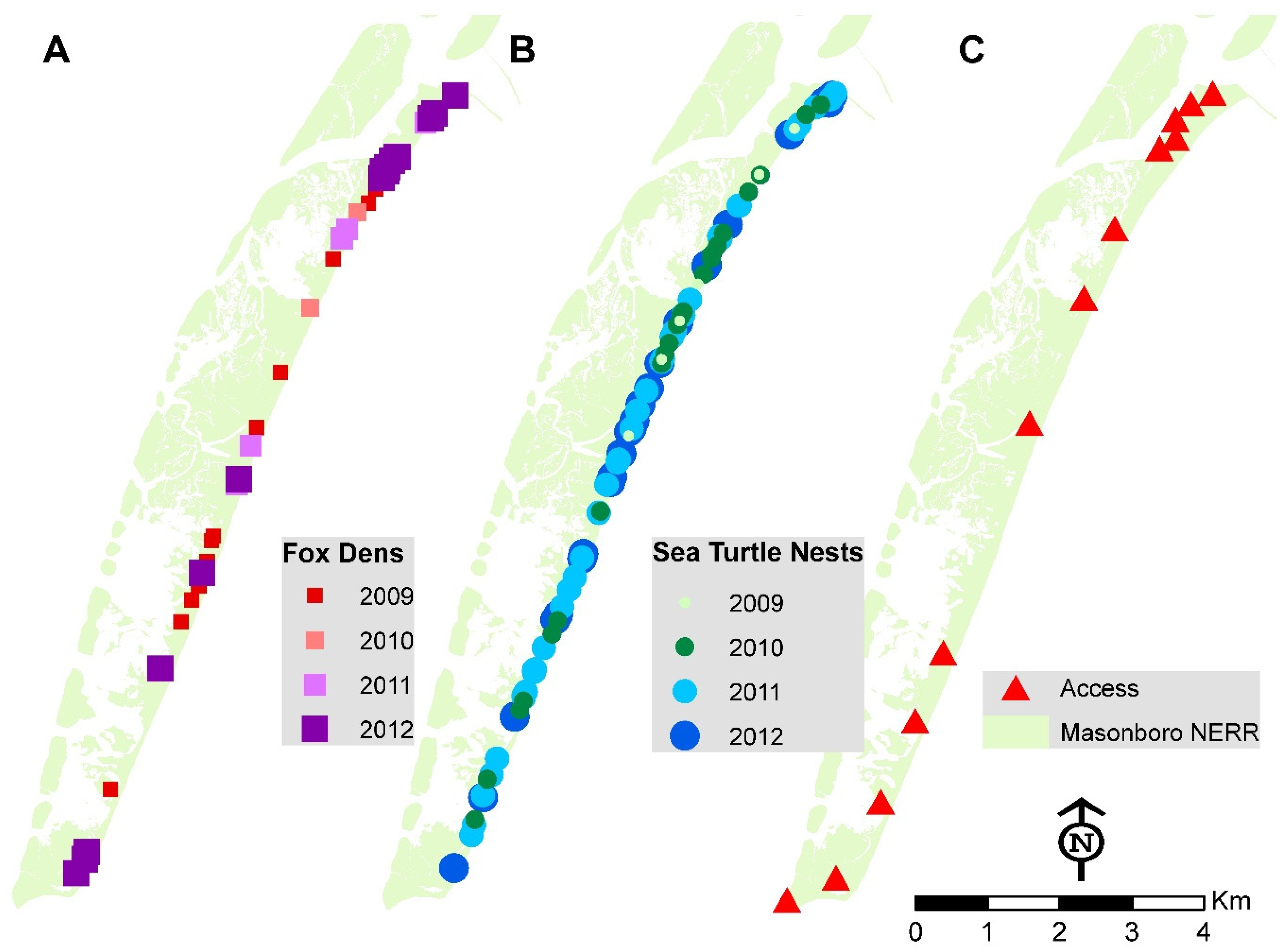
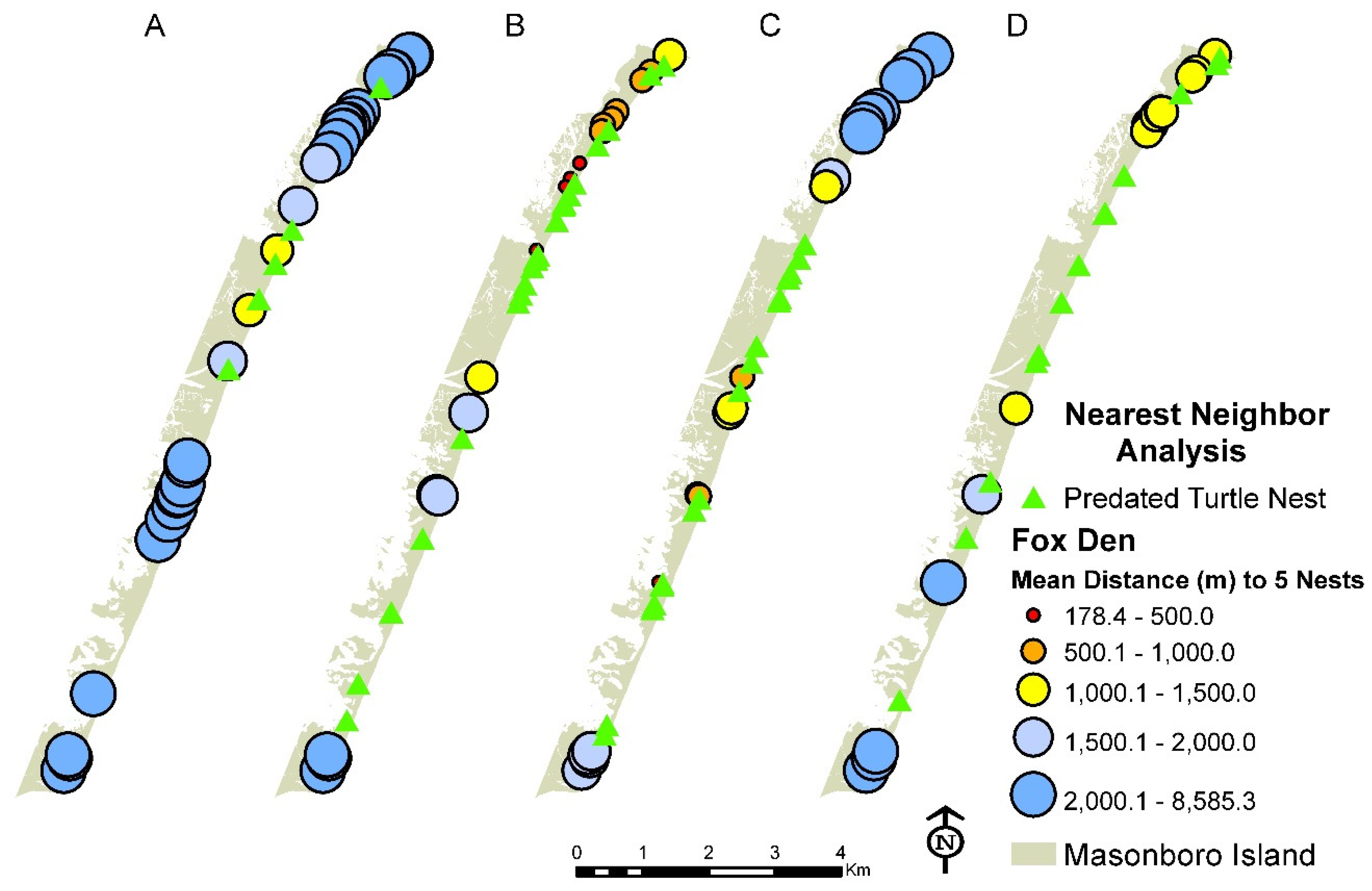
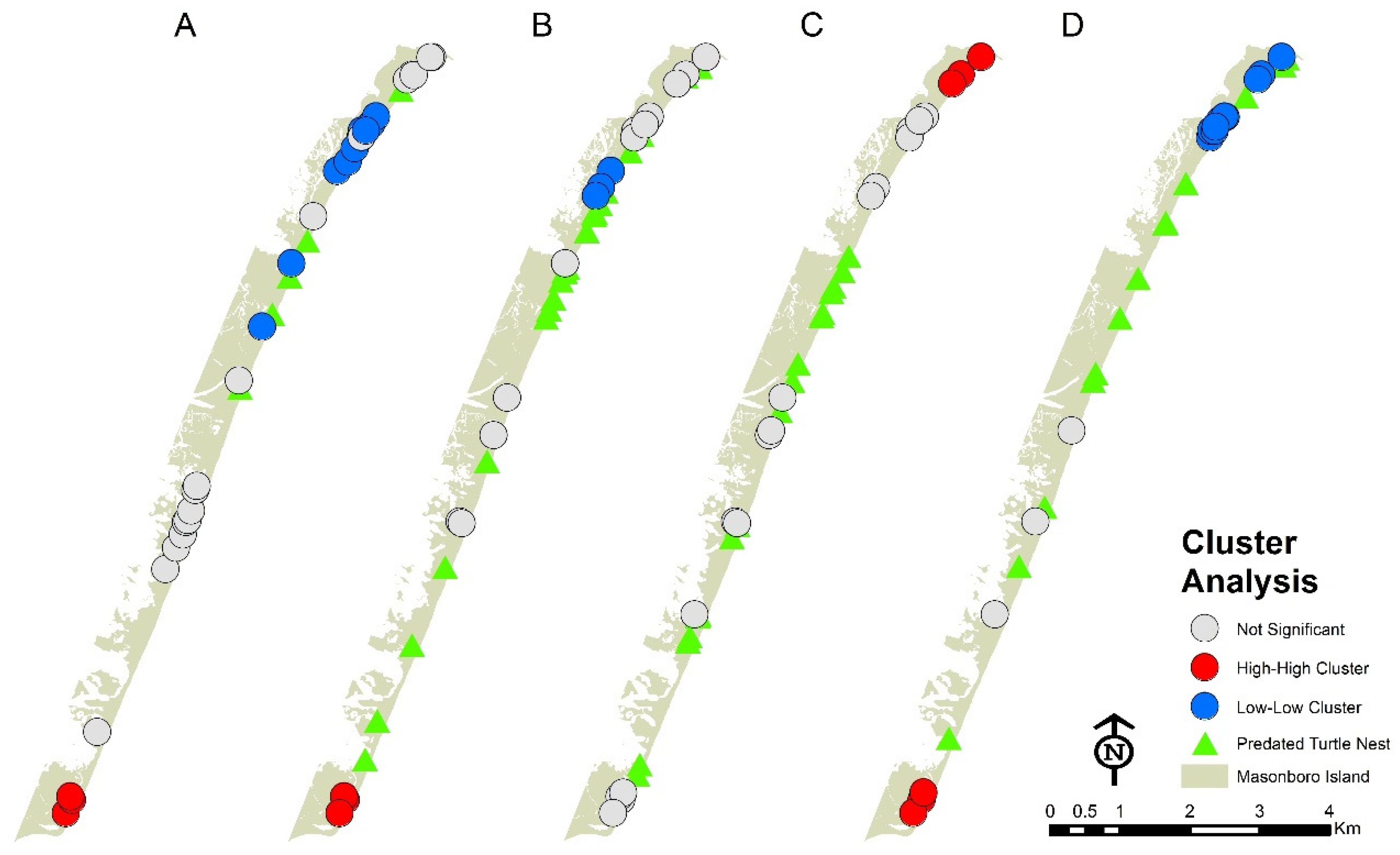
3.1. Distribution of Fox Den Entrances and Sea Turtle Nests
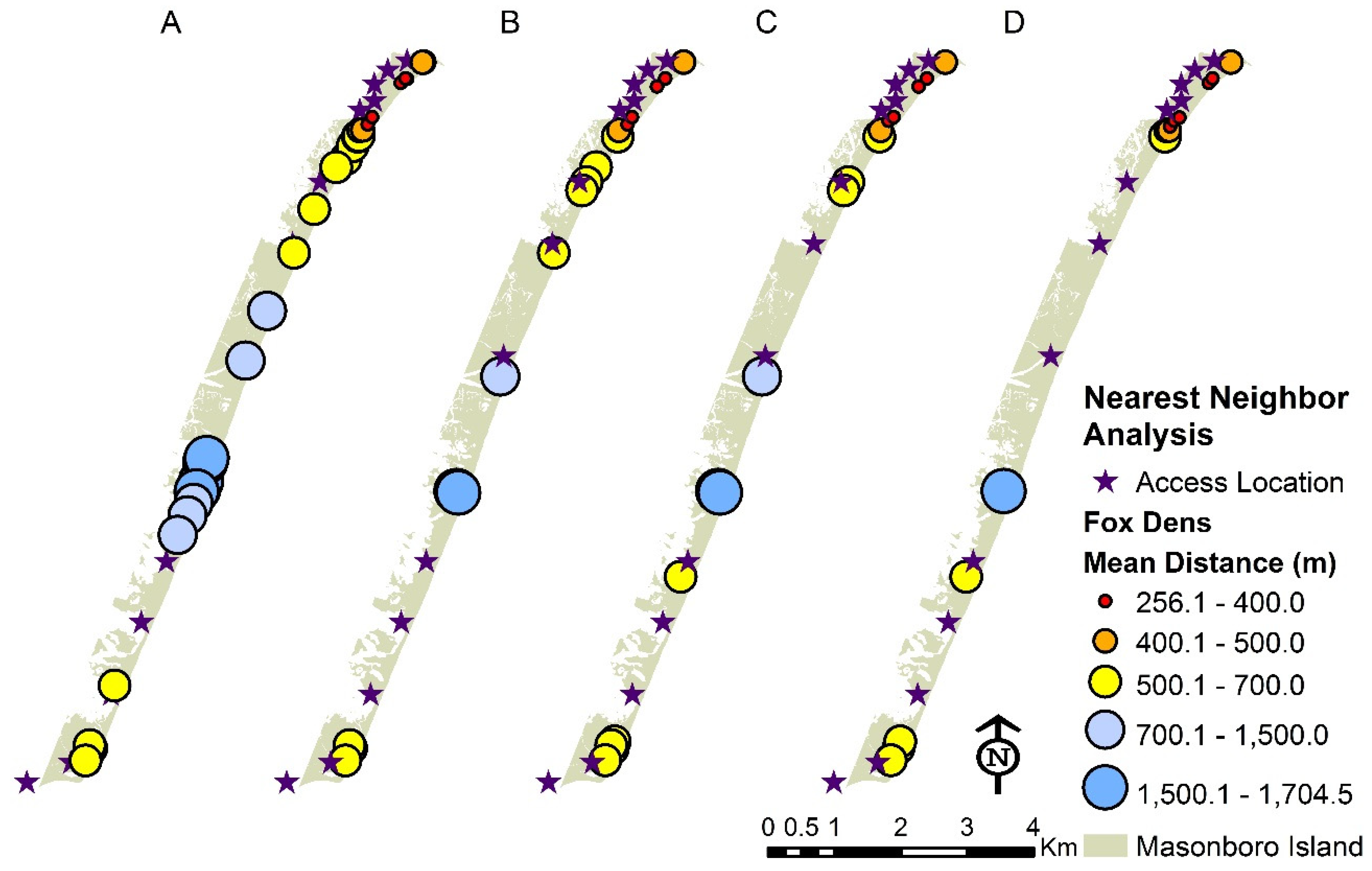
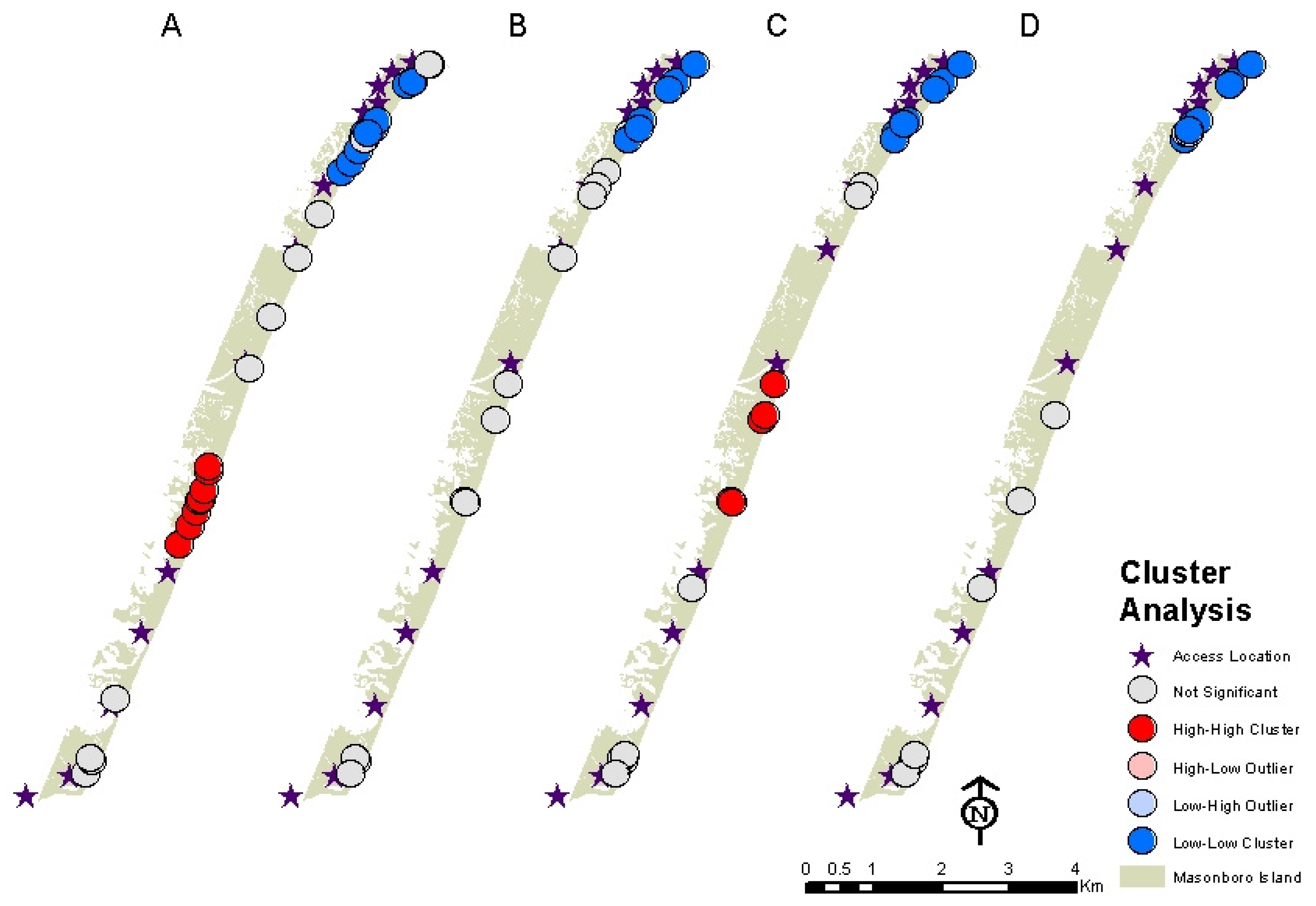
3.2. Distribution of Fox Dens and Boating Access Sites
3.3. Distribution of Fox Den Entrances Versus Island Topography
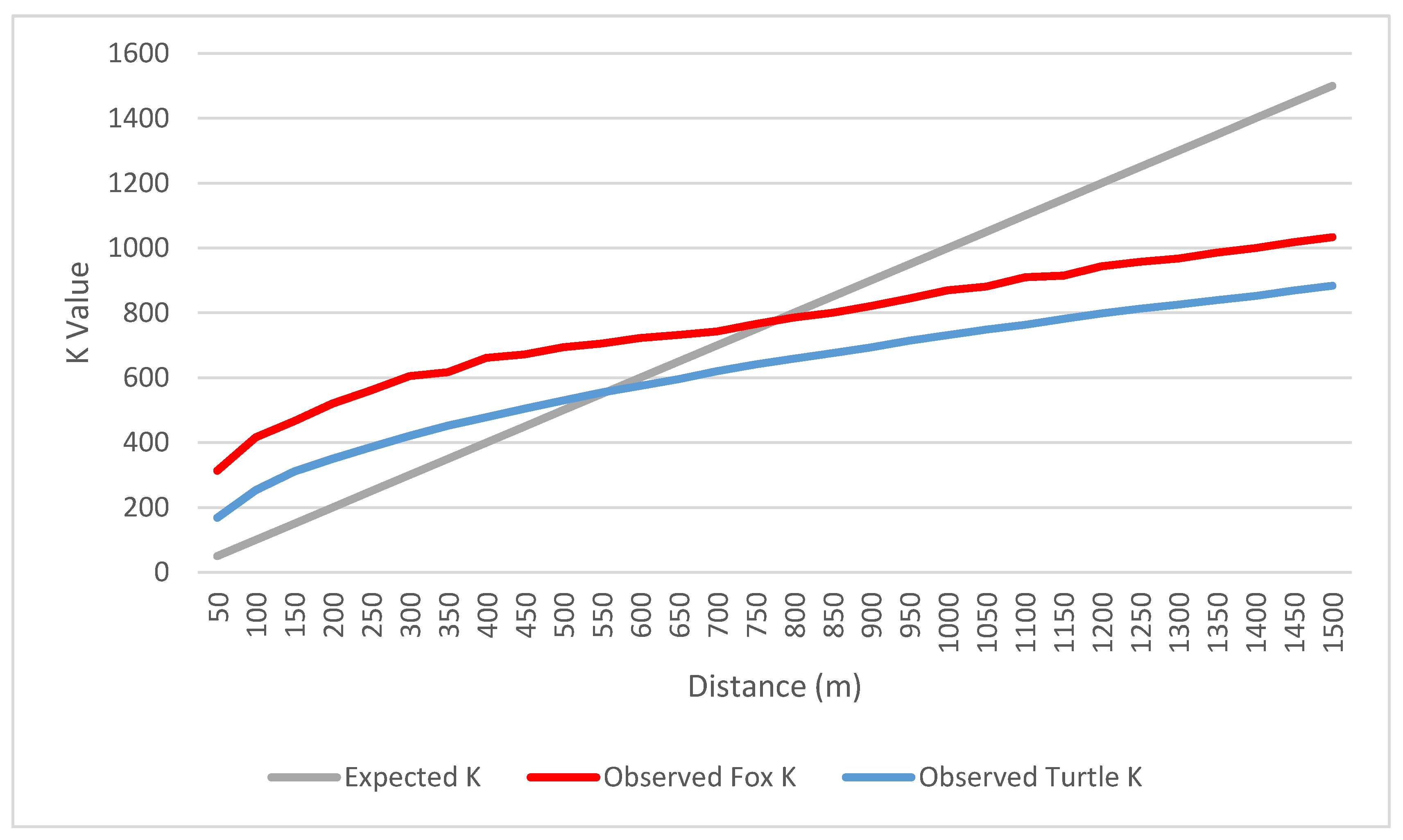
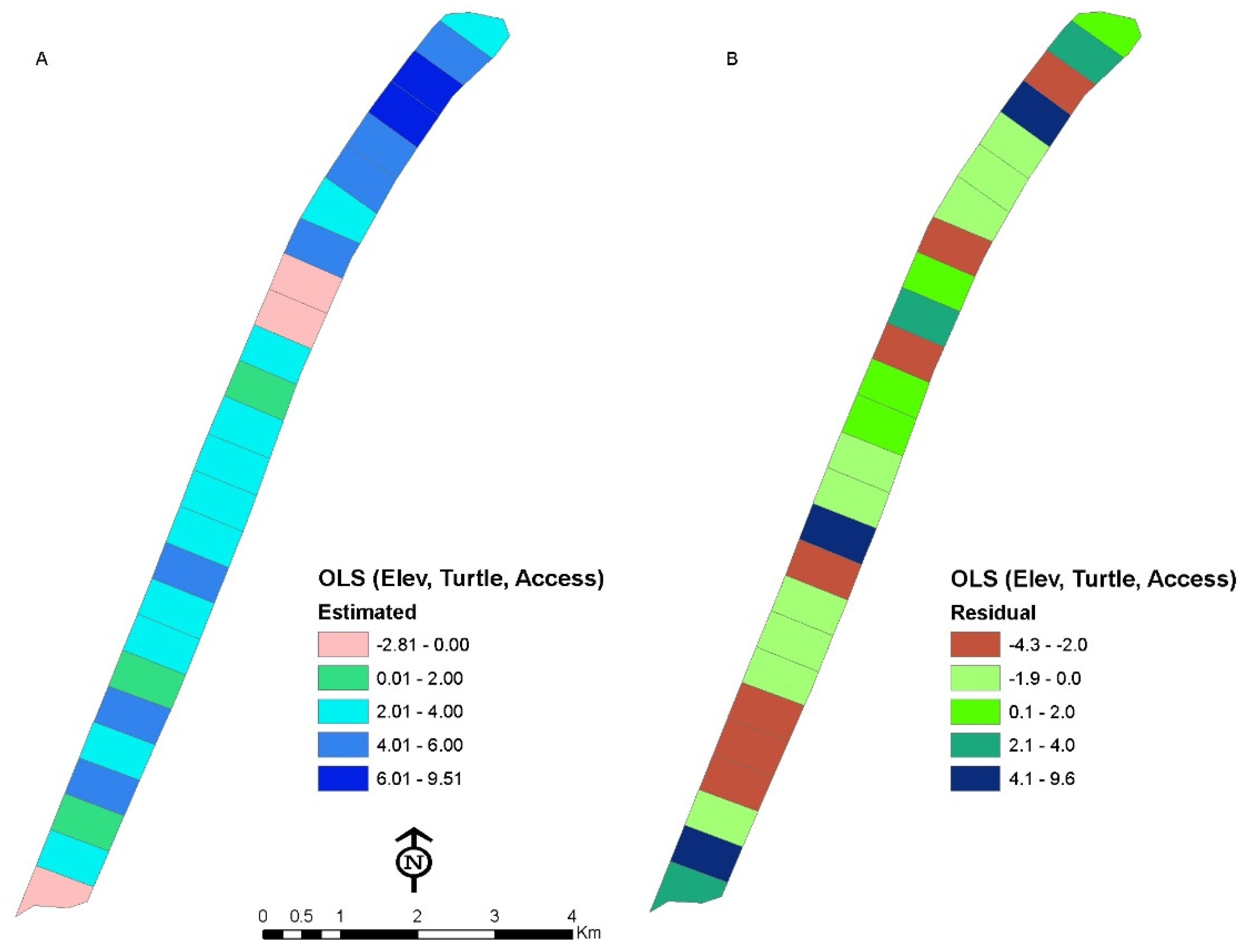
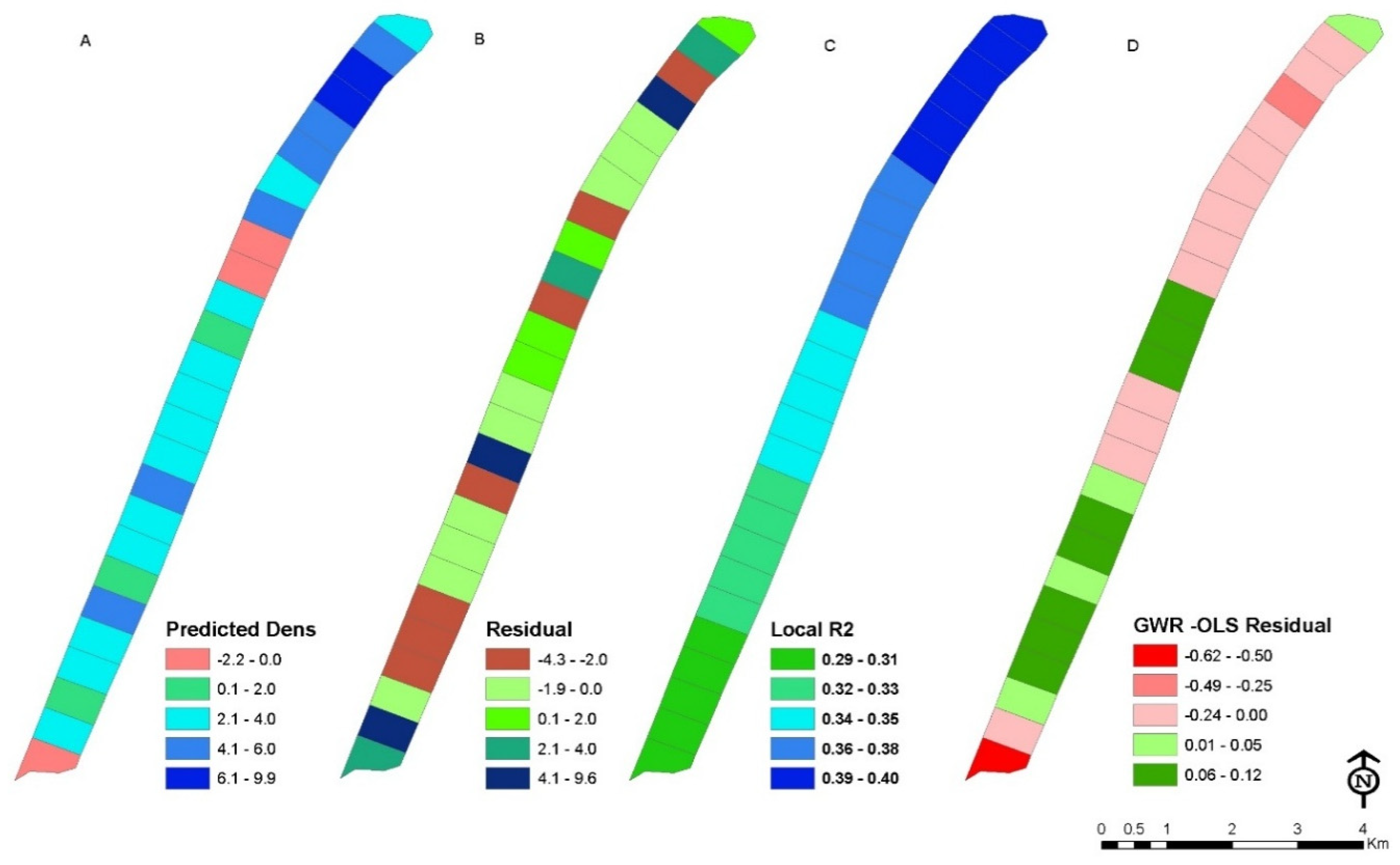
3.4. Relationship between Dependent and Independent Variables
4. Discussion and Conclusions
4.1. Wildlife Patterns and Predictions
4.2. Assumptions
4.3. Management Implications
5. Conclusions and Future Work
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kamler, J.F.; Ballard, W.B. A review of native and nonnative red foxes in North America. Wildl. Soc. Bulletin 2002, 30, 370–379. [Google Scholar]
- Statham, M.; Sacks, B.; Aubry, K.; Perrine, J.; Wisely, S. The origin of recently established red fox populations in the united states: Translocations or natural range expansions? J. Mammal. 2012, 93, 52–65. [Google Scholar] [CrossRef]
- NCWRC. Fox and Coyote Populations Study Final Report; North Carolina Wildlife Resources Commission: Raleigh, NC, USA, 2012. [Google Scholar]
- Albers, G.; Day, K.A.; Olfenbuttel, C.; Sawyer, D.; Sumner, P. Red Fox: North Carolina Wildlife Profiles. North Carolina Wildlife Resources Commission: Raleigh, NC, USA, 2017. Available online: http://www.ncwildlife.org/Portals/0/Learning/documents/Profiles/RedFox_1.pdf (accessed on 15 September 2017).
- Sheldon, W.G. Denning habits and home range of red foxes in new-york state. J. Wildl. Manag. 1950, 14, 33–42. [Google Scholar] [CrossRef]
- Soulsbury, C.D.; Baker, P.J.; Iossa, G.; Harris, S. Red foxes (vulpes vulpes). In Urban Carnivores: Ecology, Conflict, and Conservation; Gehrt, S.D., Riley, S.P.D., Cypher, B.L., Eds.; John Hopkins University Press: Baltimore, MD, USA, 2010; pp. 63–75. [Google Scholar]
- Buerger, R.; Hill, J.; Hertsine, J.; Taggart, J. The impact of recreation on barrier islands: A case study on masonboro island. Coast. Manag. 2000, 28, 249–259. [Google Scholar]
- Buerger, R.; Hill, J.; Herstine, J.; Auger, D.; Taggart, J. Re-emerging recreational use patterns on an undeveloped barrier island following the impact of hurricanes: A North Carolina case study. Soc. Nat. Resour. 2003, 16, 527–539. [Google Scholar] [CrossRef]
- Rabon, D. Green Sea Turtles in North Carolina. Available online: https://www.fws.gov/nc-es/reptile/greensea.html (accessed on 18 May 2017).
- Rabon, D. Loggerhead Sea Turtles in North Carolina. Available online: https://www.fws.gov/nc-es/reptile/logger.html (accessed on 18 May 2017).
- Kurz, D.; Straley, K.; DeGregorio, B. Out-foxing the red fox: How best to protect the nests of the endangered loggerhead marine turtle caretta caretta from mammalian predation? Oryx 2012, 46, 223–228. [Google Scholar] [CrossRef]
- Esri. Arcgis; 10.5.1; Esri: Redlands, California, USA, 2017. [Google Scholar]
- Bailey, T.C.; Gatrell, A.C. Interactive Spatial Data Analysis; J. Wiley: New York, NY, USA, 1995; p. 413. [Google Scholar]
- Fritz, C.E.; Schuurman, N.; Robertson, C.; Lear, S. A scoping review of spatial cluster analysis techniques for point-event data. Geospat. Health 2013, 7, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Zuckerberg, B.; Porter, W.F.; Zhang, L. Use of localized descriptive statistics for exploring the spatial pattern changes of bird species richness at multiple scales. Appl. Geogr. 2012, 32, 185–194. [Google Scholar] [CrossRef]
- Wilkins, D.E.; Ford, R.L. Nearest neighbor methods applied to dune field organization: The coral pink sand dunes, Kane County, Utah, USA. Geomorphology 2007, 83, 48–57. [Google Scholar] [CrossRef]
- Law, R.; Illian, J.; Burslem, D.; Gratzer, G.; Gunatilleke, C.V.S.; Gunatilleke, I. Ecological information from spatial patterns of plants: Insights from point process theory. J. Ecol. 2009, 97, 616–628. [Google Scholar] [CrossRef]
- Getis, A. A history of the concept of spatial autocorrelation: A geographer’s perspective. Geographical Analysis 2008, 40, 297–309. [Google Scholar] [CrossRef]
- Anselin, L. Local indicators of spatial association—Lisa. Geogr. Anal. 1995, 27, 93–115. [Google Scholar] [CrossRef]
- Medeiros, S.; Hagen, S.; Weishampel, J.; Angelo, J. Adjusting lidar-derived digital terrain models in coastal marshes based on estimated aboveground biomass density. Remote Sens. 2015, 7, 3507–3525. [Google Scholar] [CrossRef]
- Mongus, D.; Zalik, B. Parameter-free ground filtering of lidar data for automatic dtm generation. ISPRS J. Photogramm. Remote Sens. 2012, 67, 1–12. [Google Scholar] [CrossRef]
- Pietro, L.S.; O’Neal, M.A.; Puleo, J.A. Developing terrestrial-lidar-based digital elevation models for monitoring beach nourishment performance. J. Coast. Res. 2008, 24, 1555–1564. [Google Scholar] [CrossRef]
- Rayburg, S.; Thoms, M.; Neave, M. A comparison of digital elevation models generated from different data sources. Geomorphology 2009, 106, 261–270. [Google Scholar] [CrossRef]
- Rogers, J.N.; Parrish, C.E.; Ward, L.G.; Burdick, D.M. Assessment of elevation uncertainty in salt marsh environments using discrete-return and full-waveform lidar. J. Coast. Res. 2016, 76, 107–122. [Google Scholar] [CrossRef]
- Smeeckaert, J.; Mallet, C.; David, N.; Chehata, N.; Ferraz, A. Large-scale classification of water areas using airborne topographic lidar data. Remote Sens. Environ. 2013, 138, 134–148. [Google Scholar] [CrossRef]
- Pe’eri, S.; Long, B. Lidar technology applied in coastal studies and management. J. Coast. Res. 2011. [Google Scholar] [CrossRef]
- Gosselink, T.E.; Van Deelen, T.R.; Warner, R.E.; Joselyn, M.G. Temporal habitat partitioning and spatial use of coyotes and red foxes in east-central Illinois. J. Wildl. Manag. 2003, 67, 90–103. [Google Scholar] [CrossRef]
- Hamilton, L. Regression with Graphics: A Second Course in Applied Statistics; Duxbury Press: Belmont, CA, USA, 1992. [Google Scholar]
- Fotheringham, A.S.; Brunsdon, C.; Charlton, M. Geographically Weighted Regression: The Analysis of Spatially Varying Relationships; Wiley: Hoboken, NJ, USA, 2002. [Google Scholar]
- Crawford, T.W.; Marcucci, D.J.; Bennett, A. Impacts of residential development on vegetation cover for a remote coastal barrier in the outer banks of North Carolina, USA. J. Coast. Conserv. 2013, 17, 431–443. [Google Scholar] [CrossRef]
- Kim, J.; Nicholls, S. Using geographically weighted regression to explore the equity of public open space distributions. J. Leis. Res. 2016, 48, 105–133. [Google Scholar] [CrossRef]
- Trewhella, W.; Harris, S.; McAllister, F. Dispersal distance, home-range size and population-density in the red fox (vulpes vulpes)—A quantitative analysis. J. Appl. Ecol. 1988, 25, 423–434. [Google Scholar] [CrossRef]
- Ables, E.D. Ecology of the red fox in america. In The Wild Canids; Fox, M.W., Ed.; Van Nostrand Reinhold: New York, NY, USA, 1975; pp. 216–235. [Google Scholar]
- Voight, D.R. Red fox. In Wild Furbearer Management and Conservation I North America; Novak, M., Baker, J.A., Obbard, M.E., Malloch, B., Eds.; Ontario Trappers Association and Ontario Ministry of Natural Resources: Ottawa, ON, Canada, 1987; pp. 379–392. [Google Scholar]
- Newsome, T.; Ripple, W. A continental scale trophic cascade from wolves through coyotes to foxes. J. Anim. Ecol. 2015, 84, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Panek, M. Factors affecting predation of red foxes vulpes vulpes on brown hares lepus europaeus during the breeding season in Poland. Wildl. Biol. 2009, 15, 345–349. [Google Scholar] [CrossRef]
- Hill, J.; Herstine, J.; Buerger, R.; Snider, A. North Carolina Coastal Reserve Program Initiatives, Unpublished data. 2008.
- Fowler, L.E. Hatching success and nest predation in the green sea turtle, chelonia-mydas, at tortuguero, costa rica. Ecology 1979, 60, 946–955. [Google Scholar] [CrossRef]
- Hitipeuw, C.; Dutton, P.; Benson, S.; Thebu, J.; Bakarbessy, J. Population status and internesting movement of leatherback turtles, dermochelys coriacea, nesting on the Northwest Coast of Papua, Indonesia. Chelonian Conserv. Biol. 2007, 6, 28–36. [Google Scholar] [CrossRef]
- Ordonez, C.; Troeng, S.; Meylan, A.; Meylan, P.; Ruiz, A. Chiriqui beach, panama, the most important leatherback nesting beach in central America. Chelonian Conserv. Biol. 2007, 6, 122–126. [Google Scholar] [CrossRef]
- Patino-Martinez, J.; Marco, A.; Quinones, L.; Godley, B. Globally significant nesting of the leatherback turtle (dermochelys coriacea) on the caribbean coast of Colombia and Panama. Biol.Conserv. 2008, 141, 1982–1988. [Google Scholar] [CrossRef]
- Ruiz-Olmo, J.; Blanch, F.; Vidal, F. Relationships between the red fox and waterbirds in the ebro delta natural park, ne spain. Waterbirds 2003, 26, 217–225. [Google Scholar] [CrossRef]
- Jarnevich, C.; Evangelista, P.; Stohlgren, T.; Morisette, J. Improving national-scale invasion maps: Tamarisk in the western United States. W. N. Am. Nat. 2011, 71, 164–175. [Google Scholar] [CrossRef]
- Pearson, R.G. Species’ distribution modeling for conservation educators and practitioners. Synth. Am. Mus. Nat. Hist. 2007, 3, 54–89. [Google Scholar]
- Morisette, J.; Jarnevich, C.; Holcombe, T.; Talbert, C.; Ignizio, D.; Talbert, M.; Silva, C.; Koop, D.; Swanson, A.; Young, N. Vistrails sahm: Visualization and workflow management for species habitat modeling. Ecography 2013, 36, 129–135. [Google Scholar] [CrossRef]
- Ramsey, D.S.L.; Caley, P.A.; Robley, A. Estimating population density from presence-absence data using a spatially explicit model. J. Wildl. Manag. 2015, 79, 491–499. [Google Scholar] [CrossRef]
- West, A.; Evangelista, P.; Jarnevich, C.; Kumar, S.; Swallow, A.; Luizza, M.; Chignell, S. Using multi-date satellite imagery to monitor invasive grass species distribution in post-wildfire landscapes: An iterative, adaptable approach that employs open-source data and software. Int. J. Appl. Earth Observ. Geoinf. 2017, 59, 135–146. [Google Scholar] [CrossRef]
- Daniel, C.J.; Frid, L.; Sleeter, B.M.; Fortin, M.J. State-and-transition simulation models: A framework for forecasting landscape change. Methods Ecol. Evolut. 2016, 7, 1413–1423. [Google Scholar] [CrossRef]
- Vall-llosera, M.; Woolnough, A.; Anderson, D.; Cassey, P. Improved surveillance for early detection of a potential invasive species: The alien rose-ringed parakeet Psittacula krameri in Australia. Biol. Invasions 2017, 19, 1273–1284. [Google Scholar] [CrossRef]
- Brock, J.C.; Krabill, W.B.; Sallenger, A.H. Barrier island morphodynamic classification based on lidar metrics for North Assateague Island, Maryland. J. Coast. Res. 2004, 20, 498–509. [Google Scholar] [CrossRef]
- Halls, J.N. Measuring habitat changes in barrier island marshes: An example from Southeastern North Carolina. In Remote Sensing and Geospatial Technologies for Coastal Ecosystem Assessment and Management: Principles and Applications; Yang, X., Ed.; Lecture Notes in Geoinformation and Cartography; Springer: Berlin/Heidelberg, Germany, 2009; pp. 391–413. [Google Scholar]
- Houser, C. Feedback between ridge and swale bathymetry and barrier island storm response and transgression. Geomorphology 2012, 173–174, 1–16. [Google Scholar] [CrossRef]
- Mitasova, H.; Overton, M.F.; Recalde, J.J.; Bernstein, D.J.; Freeman, C.W. Raster-based analysis of coastal terrain dynamics from multitemporal lidar data. J. Coast. Res. 2009, 25, 507–514. [Google Scholar] [CrossRef]
- Halls, J.N.; Frishman, M.A.; Hawkes, A.D. An Automated Model to Classify Barrier Island Geomorphology Using Lidar Data. In Proceedings of the 2nd International Electronic Conference on Remote Sensing, Online. 22 March–5 April 2018; p. 336. [Google Scholar]
- McCarthy, M.; Halls, J. Habitat mapping and change assessment of coastal environments: An examination of worldview-2, quickbird, and ikonos satellite imagery and airborne lidar for mapping barrier island habitats. ISPRS Int. J. Geo-Inf. 2014, 3, 297–325. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Count | Observed Mean Distance (m) | Expected Mean Distance (m) | z-Score | p-Value | Index | Result |
|---|---|---|---|---|---|---|---|
| all | 84 | 43.2 | 128.5 | −11.64 | 0.000 | 0.336 | clustered |
| 2009 | 31 | 193.2 | 211.5 | −0.92 | 0.357 | 0.914 | random |
| 2010 | 18 | 242.7 | 277.5 | −1.02 | 0.307 | 0.875 | random |
| 2011 | 19 | 210.3 | 270.1 | −1.85 | 0.065 | 0.778 | clustered |
| 2012 | 16 | 352 | 294.3 | 1.49 | 0.137 | 1.194 | random |
| Year | Count | Observed Mean Distance (m) | Expected Mean Distance (m) | z-Score | p-Value | Index | Result |
|---|---|---|---|---|---|---|---|
| All years (nests & false crawls) | 177 | 43 | 88.5 | −13.08 | 0.000 | 0.486 | significantly clustered |
| All years (false crawls) | 84 | 79.9 | 128.5 | −6.62 | 0.000 | 0.622 | significantly clustered |
| All years (nests) | 93 | 70.8 | 122.1 | −7.76 | 0.000 | 0.579 | significantly clustered |
| All Years (predated nests) | 58 | 107 | 154.6 | −4.48 | 0.000 | 0.662 | significantly clustered |
| 2009 nests | 6 | 749.9 | 480.7 | 2.62 | 0.0087 | 1.560 | dispersed |
| 2010 nests | 24 | 242.7 | 240.3 | 0.093 | 0.9255 | 1.010 | random |
| 2011 nests | 38 | 147.03 | 190.99 | −2.71 | 0.0066 | 0.769 | significantly clustered |
| 2012 nests | 25 | 295 | 235.48 | 2.44 | 0.0149 | 1.255 | dispersed |
| 2009 false crawls | 9 | 326.5 | 392.5 | −0.965 | 0.3346 | 0.832 | random |
| 2010 false crawls | 11 | 874.8 | 354.99 | 9.29 | 0.000 | 2.464 | dispersed |
| 2011 false crawls | 29 | 208.3 | 218.6 | −0.485 | 0.6276 | 0.953 | random |
| 2012 false crawls | 35 | 184.85 | 199.01 | −0.805 | 0.4207 | 0.929 | random |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Halls, J.N.; Hill, J.M.; Urbanek, R.E.; Sutton, H. Distribution Pattern of Red Fox (Vulpes vulpes) Dens and Spatial Relationships with Sea Turtle Nests, Recreation, and Environmental Characteristics. ISPRS Int. J. Geo-Inf. 2018, 7, 247. https://0-doi-org.brum.beds.ac.uk/10.3390/ijgi7070247
Halls JN, Hill JM, Urbanek RE, Sutton H. Distribution Pattern of Red Fox (Vulpes vulpes) Dens and Spatial Relationships with Sea Turtle Nests, Recreation, and Environmental Characteristics. ISPRS International Journal of Geo-Information. 2018; 7(7):247. https://0-doi-org.brum.beds.ac.uk/10.3390/ijgi7070247
Chicago/Turabian StyleHalls, Joanne N., Jeffery M. Hill, Rachael E. Urbanek, and Hope Sutton. 2018. "Distribution Pattern of Red Fox (Vulpes vulpes) Dens and Spatial Relationships with Sea Turtle Nests, Recreation, and Environmental Characteristics" ISPRS International Journal of Geo-Information 7, no. 7: 247. https://0-doi-org.brum.beds.ac.uk/10.3390/ijgi7070247