Pectoral Fin Anomalies in tbx5a Knockdown Zebrafish Embryos Related to the Cascade Effect of N-Cadherin and Extracellular Matrix Formation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Ethics
2.2. Maintenance and Breeding of Zebrafish and Embryo Collection
2.3. Microinjections and Morpholino Treatment
2.4. Validation of the Efficacy, Traumatic Effects, and Off-Target Effects of tbx5a MO
2.5. Histology
2.6. Alcian Blue Staining
2.7. Immunohistochemistry Assay
2.8. RNA Isolation
2.9. Microarray Analysis
2.10. Quantitative Reverse-Transcriptase Polymerase Chain Reaction (qPCR)
2.11. Statistical Analysis
3. Results
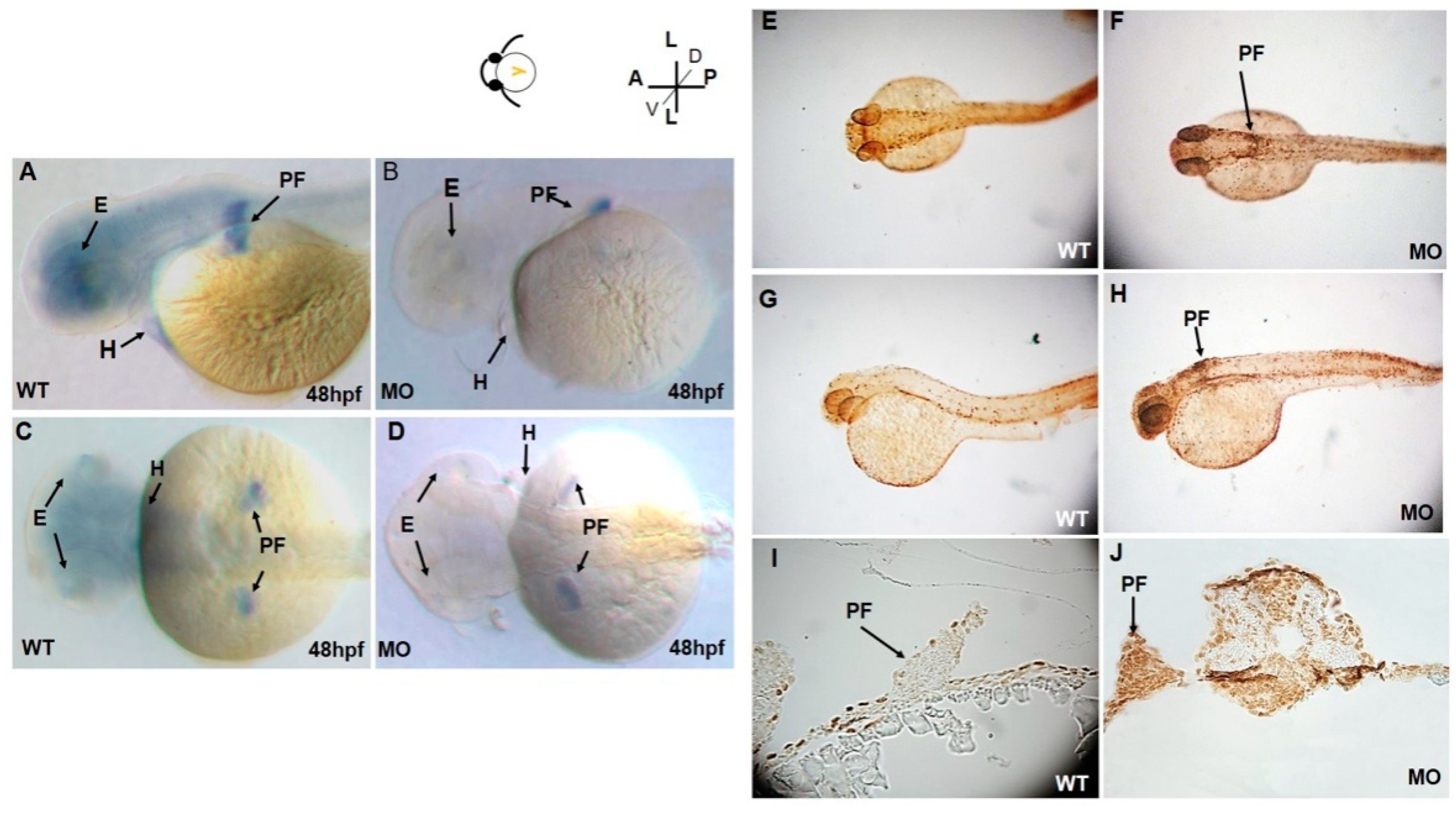
3.1. Abnormal Phenotypes of Pectoral Fins
3.2. Depressed tbx5a Gene Expression
3.3. Microarray Screening
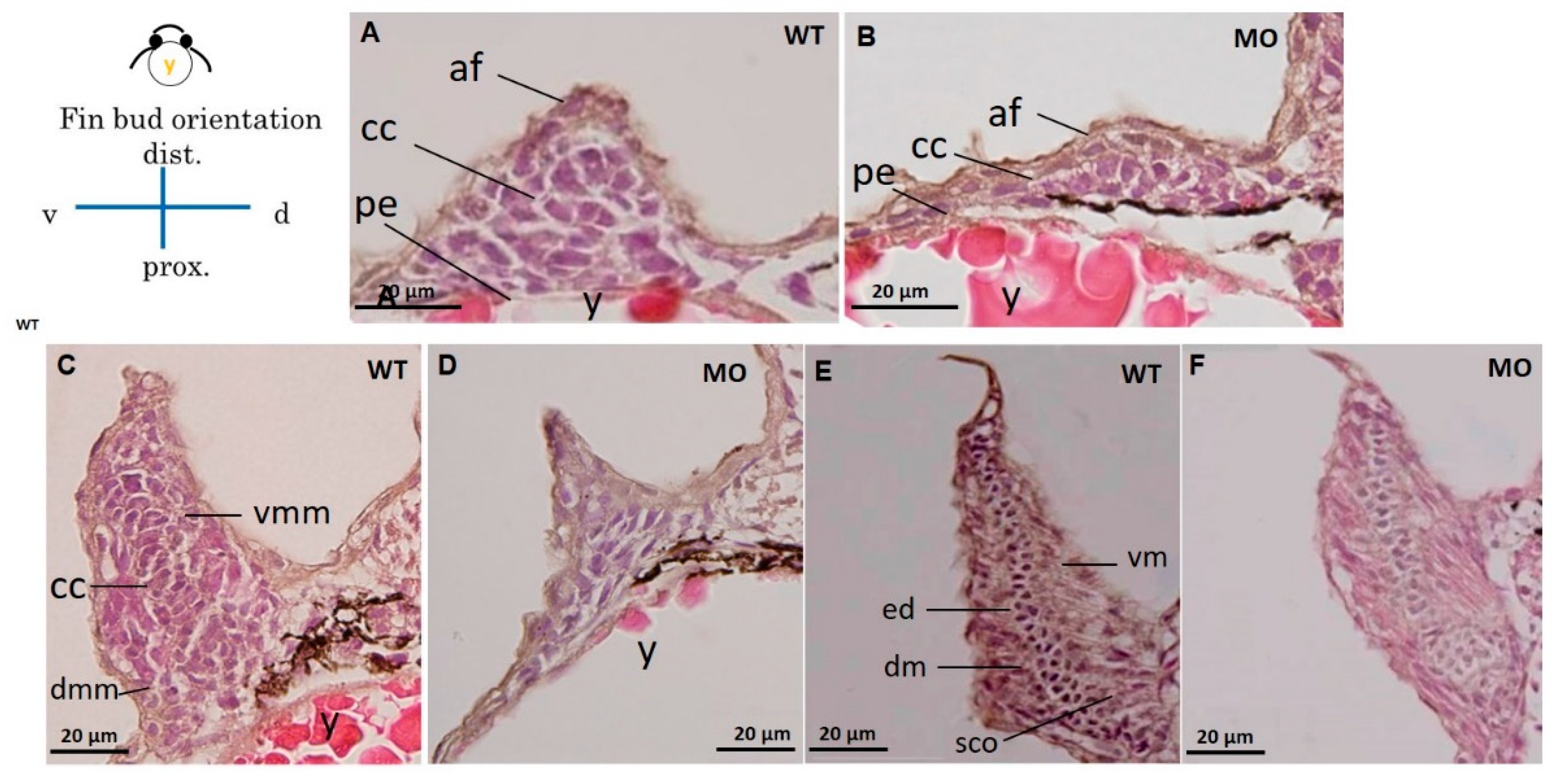
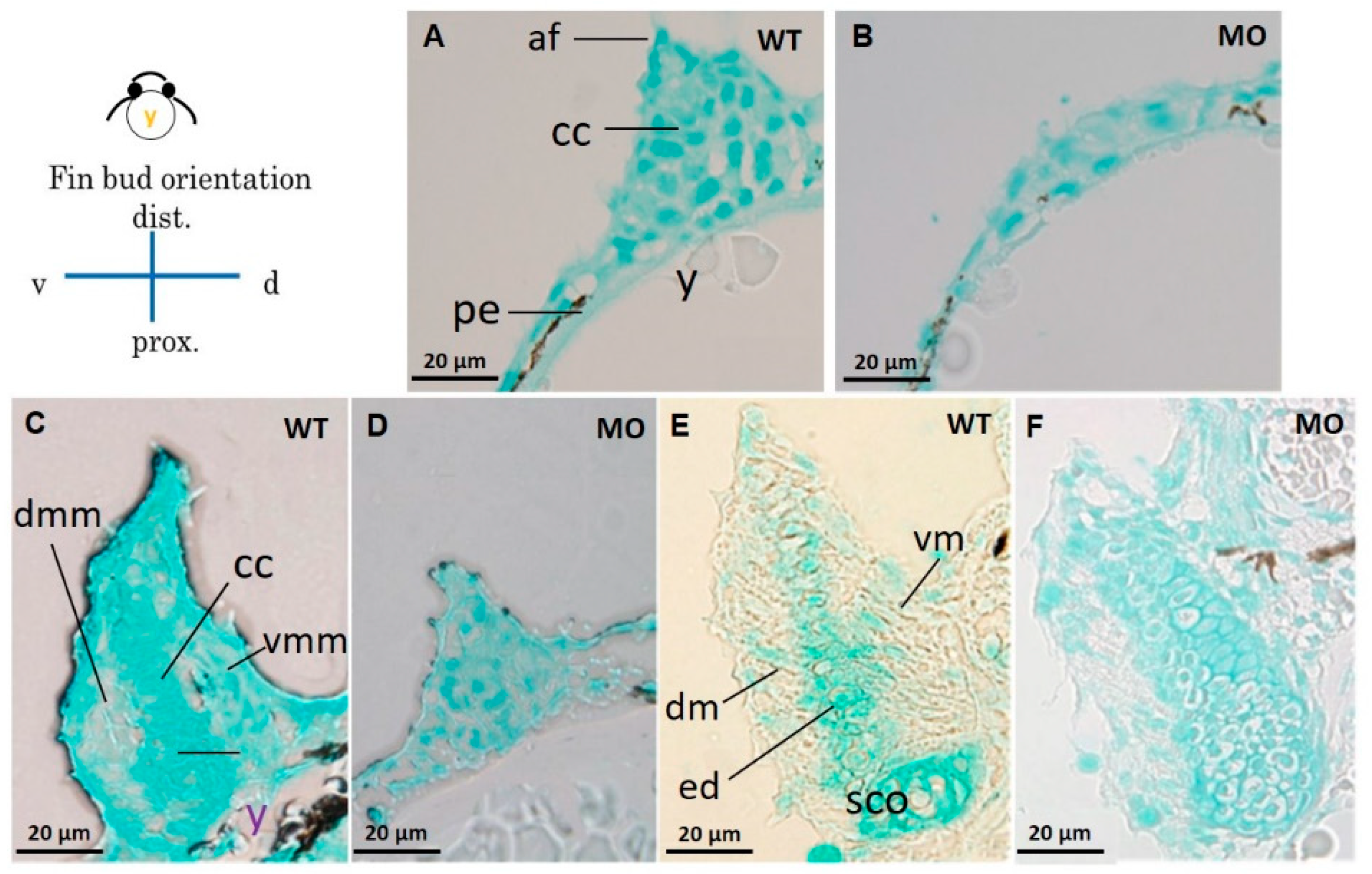
3.4. Chondrocytes and Glycosaminoglycans in Pectoral Fins
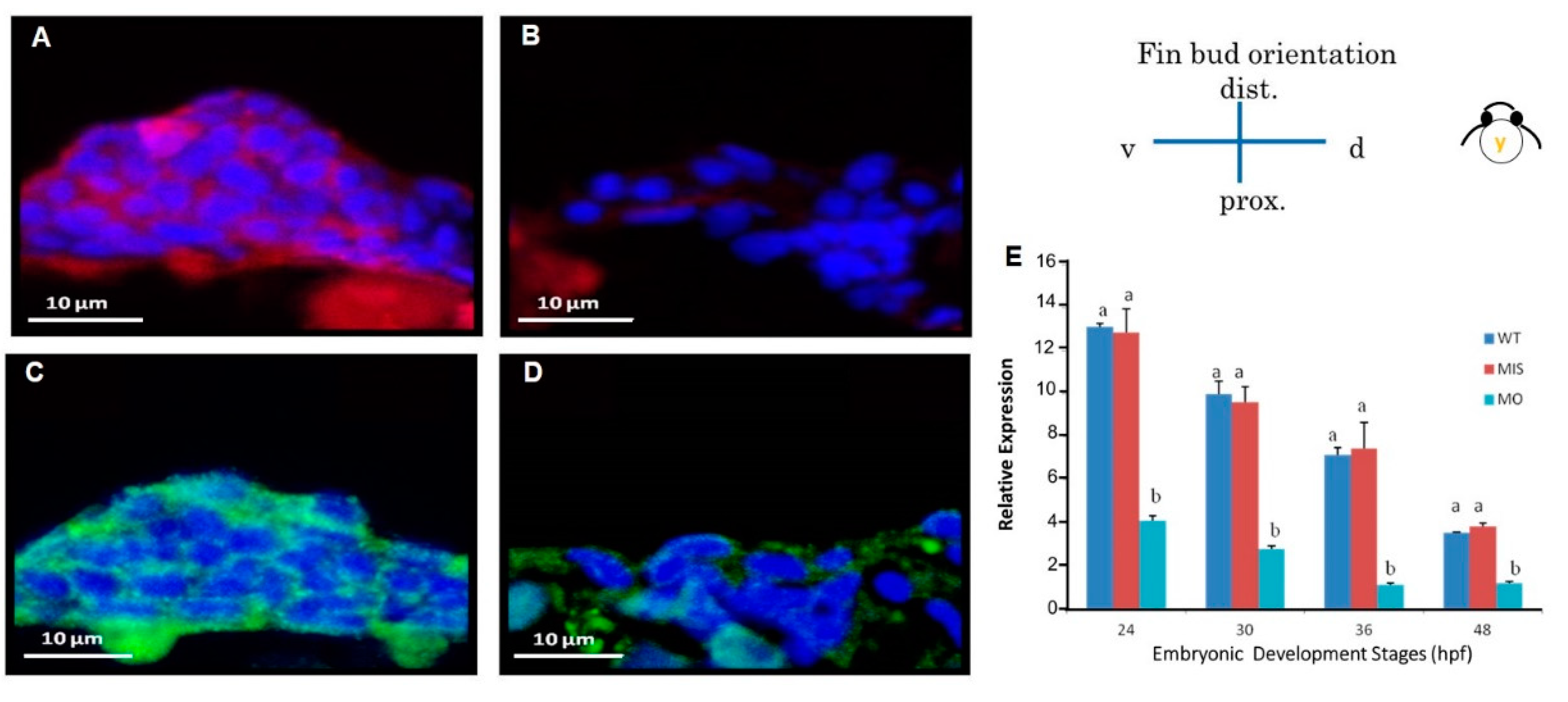
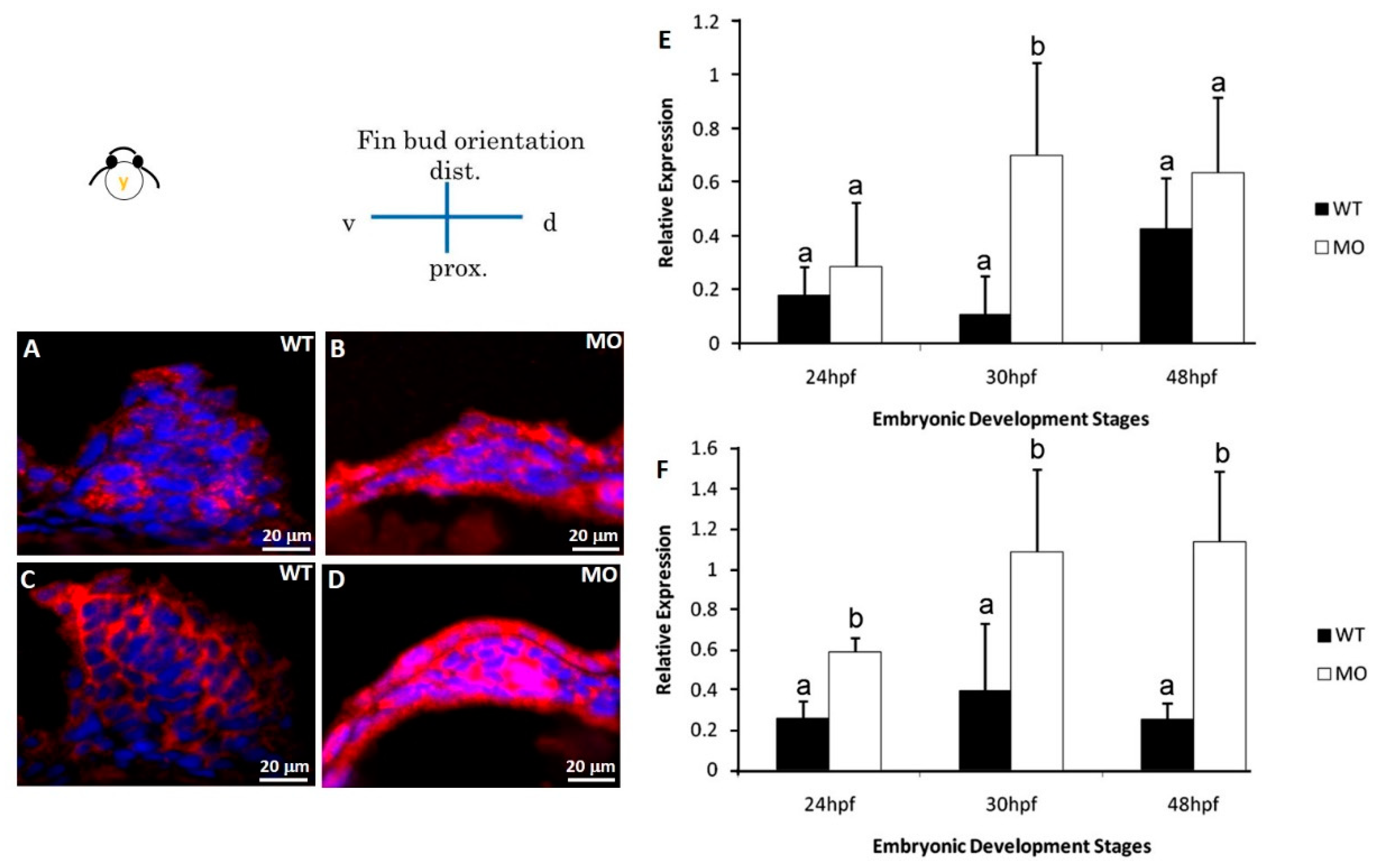
3.5. Reduced cdh2 Gene Expression
3.6. Reduction in N-Cadherin Protein Expression in Pectoral Fins
3.7. Reduction in ECM Protein and Cx4 Expression
3.8. Increase in Apoptosis-Related Proteins in Pectoral Fins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hall, B.K.; Miyake, T. All for one and one for all: Condensations and the initiation of skeletal development. Bioessays 2000, 22, 138–147. [Google Scholar] [CrossRef]
- Mercader, N. Early steps of paired fin development in zebrafish compared with tetrapod limb development. Dev. Growth Differ. 2007, 49, 421–437. [Google Scholar] [CrossRef] [PubMed]
- Hasson, P.; Del Buono, J.; Logan, M.P. Tbx5 is dispensable for forelimb outgrowth. Development 2007, 134, 85–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasson, P.; DeLaurier, A.; Bennett, M.; Grigorieva, E.; Naiche, L.A.; Papaioannou, V.E.; Mohun, T.J.; Logan, M.P. Tbx4 and tbx5 acting in connective tissue are required for limb muscle and tendon patterning. Dev. Cell 2010, 18, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Basson, C.T.; Bachinsky, D.R.; Lin, R.C.; Levi, T.; Elkins, J.A.; Soults, J.; Grayzel, D.; Kroumpouzou, E.; Traill, T.A.; Leblanc-Straceski, J.; et al. Mutations in human TBX5 [corrected] cause limb and cardiac malformation in Holt-Oram syndrome. Nat. Genet. 1997, 15, 30–35. [Google Scholar] [CrossRef]
- Gibson-Brown, J.J.; Agulnik, S.I.; Chapman, D.L.; Alexiou, M.; Garvey, N.; Silver, L.M.; Papaioannou, V.E. Evidence of a role for T-box genes in the evolution of limb morphogenesis and the specification of forelimb/hindlimb identity. Mech. Dev. 1996, 56, 93–101. [Google Scholar] [CrossRef]
- Yano, T.; Abe, G.; Yokoyama, H.; Kawakami, K.; Tamura, K. Mechanism of pectoral fin outgrowth in zebrafish development. Development 2012, 139, 2916–2925. [Google Scholar] [CrossRef] [Green Version]
- Ryan, K.; Chin, A.J. T-box genes and cardiac development. Birth Defects Res. Part C Embryo Today Rev. 2003, 69, 25–37. [Google Scholar] [CrossRef]
- Begemann, G.; Ingham, P.W. Developmental regulation of Tbx5 in zebrafish embryogenesis. Mech. Dev. 2000, 90, 299–304. [Google Scholar] [CrossRef]
- Lee, B.C.; Roy, S. Blimp-1 is an essential component of the genetic program controlling development of the pectoral limb bud. Dev. Biol. 2006, 300, 623–634. [Google Scholar] [CrossRef] [Green Version]
- Grandel, H.; Schulte-Merker, S. The development of the paired fins in the zebrafish (Danio rerio). Mech. Dev. 1998, 79, 99–120. [Google Scholar] [CrossRef]
- Thorsen, D.H.; Hale, M.E. Development of zebrafish (Danio rerio) pectoral fin musculature. J. Morphol. 2005, 266, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Kerstetter, A.E.; Azodi, E.; Marrs, J.A. Cadherin-1, -2, and -11 expression and cadherin-2 function in the pectoral limb bud and fin of the developing zebrafish. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2003, 228, 734–739. [Google Scholar] [CrossRef] [PubMed]
- Garrity, D.M.; Childs, S.; Fishman, M.C. The heartstrings mutation in zebrafish causes heart/fin Tbx5 deficiency syndrome. Development 2002, 129, 4635–4645. [Google Scholar] [PubMed]
- Ng, J.K.; Kawakami, Y.; Buscher, D.; Raya, A.; Itoh, T.; Koth, C.M.; Rodriguez Esteban, C.; Rodriguez-Leon, J.; Garrity, D.M.; Fishman, M.C.; et al. The limb identity gene Tbx5 promotes limb initiation by interacting with Wnt2b and Fgf10. Development 2002, 129, 5161–5170. [Google Scholar] [PubMed]
- Rallis, C.; Bruneau, B.G.; Del Buono, J.; Seidman, C.E.; Seidman, J.G.; Nissim, S.; Tabin, C.J.; Logan, M.P. Tbx5 is required for forelimb bud formation and continued outgrowth. Development 2003, 130, 2741–2751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, Q.; Stinnett, H.K.; Ho, R.K. Asymmetric cell convergence-driven zebrafish fin bud initiation and pre-pattern requires Tbx5a control of a mesenchymal Fgf signal. Development 2015, 142, 4329–4339. [Google Scholar] [CrossRef] [PubMed]
- Ahn, D.G.; Kourakis, M.J.; Rohde, L.A.; Silver, L.M.; Ho, R.K. T-box gene tbx5 is essential for formation of the pectoral limb bud. Nature 2002, 417, 754–758. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, P.; Wylie, J.N.; Galceran, J.; Arkhitko, O.; Li, C.; Deng, C.; Grosschedl, R.; Bruneau, B.G. Tbx5 is essential for forelimb bud initiation following patterning of the limb field in the mouse embryo. Development 2003, 130, 623–633. [Google Scholar] [CrossRef]
- Rodriguez-Esteban, C.; Tsukui, T.; Yonei, S.; Magallon, J.; Tamura, K.; Izpisua Belmonte, J.C. The T-box genes Tbx4 and Tbx5 regulate limb outgrowth and identity. Nature 1999, 398, 814–818. [Google Scholar] [CrossRef]
- Tsai, T.C.; Lu, J.K.; Choo, S.L.; Yeh, S.Y.; Tang, R.B.; Lee, H.Y.; Lu, J.H. The paracrine effect of exogenous growth hormone alleviates dysmorphogenesis caused by tbx5 deficiency in zebrafish (Danio rerio) embryos. J. Biomed. Sci. 2012, 19, 63. [Google Scholar] [CrossRef]
- Tsai, T.C.; Shih, C.C.; Chien, H.P.; Yang, A.H.; Lu, J.K.; Lu, J.H. Anti-apoptotic effects of IGF-I on mortality and dysmorphogenesis in tbx5-deficient zebrafish embryos. BMC Dev. Biol. 2018, 18, 5. [Google Scholar] [CrossRef]
- Lu, J.H.; Lu, J.K.; Choo, S.L.; Li, Y.C.; Yeh, H.W.; Shiue, J.F.; Yeh, V.C. Cascade effect of cardiac myogenesis gene expression during cardiac looping in tbx5 knockdown zebrafish embryos. J. Biomed. Sci. 2008, 15, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Goldring, M.B.; Tsuchimochi, K.; Ijiri, K. The control of chondrogenesis. J. Cell. Biochem. 2006, 97, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Hall, B.K.; Miyake, T. The membranous skeleton: The role of cell condensations in vertebrate skeletogenesis. Anat. Embryol. 1992, 186, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Mundlos, S.; Olsen, B.R. Heritable diseases of the skeleton. Part I: Molecular insights into skeletal development-transcription factors and signaling pathways. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1997, 11, 125–132. [Google Scholar] [CrossRef]
- Lee, J.K.; Hu, J.C.; Yamada, S.; Athanasiou, K.A. Initiation of Chondrocyte Self-Assembly Requires an Intact Cytoskeletal Network. Tissue Eng. Part A 2016, 22, 318–325. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Tsai, T.; Choo, S.; Yeh, S.; Tang, R.; Yang, A.; Lee, H.; Lu, J. Induction of apoptosis and inhibition of cell growth by tbx5 knockdown contribute to dysmorphogenesis in Zebrafish embryos. J. Biomed. Sci. 2011, 18, 73. [Google Scholar] [CrossRef] [PubMed]
- Jessen, J.R. Recent advances in the study of zebrafish extracellular matrix proteins. Dev. Biol. 2015, 401, 110–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govindan, J.; Tun, K.M.; Iovine, M.K. Cx43-Dependent Skeletal Phenotypes Are Mediated by Interactions between the Hapln1a-ECM and Sema3d during Fin Regeneration. PLoS ONE 2016, 11, e0148202. [Google Scholar] [CrossRef] [PubMed]
- Michigami, T. Current understanding on the molecular basis of chondrogenesis. Clin. Pediatr. Endocrinol. 2014, 23, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Oberlender, S.A.; Tuan, R.S. Spatiotemporal profile of N-cadherin expression in the developing limb mesenchyme. Cell Adhes. Commun. 1994, 2, 521–537. [Google Scholar] [CrossRef] [PubMed]
- Tuan, R.S. Biology of developmental and regenerative skeletogenesis. Clin. Orthop. Related Res. 2004, 427, S105–S117. [Google Scholar] [CrossRef] [PubMed]
- Hulpiau, P.; van Roy, F. Molecular evolution of the cadherin superfamily. Int. J. Biochem. Cell Biol. 2009, 41, 349–369. [Google Scholar] [CrossRef] [PubMed]
- Lele, Z.; Folchert, A.; Concha, M.; Rauch, G.J.; Geisler, R.; Rosa, F.; Wilson, S.W.; Hammerschmidt, M.; Bally-Cuif, L. parachute/n-cadherin is required for morphogenesis and maintained integrity of the zebrafish neural tube. Development 2002, 129, 3281–3294. [Google Scholar] [PubMed]
- Provot, S.; Schipani, E. Molecular mechanisms of endochondral bone development. Biochem. Biophys. Res. Commun. 2005, 328, 658–665. [Google Scholar] [CrossRef] [PubMed]
- Tuan, R.S. Cellular signaling in developmental chondrogenesis: N-cadherin, Wnts, and BMP-2. J. Bone Joint Surg. 2003, 85 (Suppl. 2), 137–141. [Google Scholar] [CrossRef]
- Halbleib, J.M.; Nelson, W.J. Cadherins in development: Cell adhesion, sorting, and tissue morphogenesis. Genes Dev. 2006, 20, 3199–3214. [Google Scholar] [CrossRef]
- Oberlender, S.A.; Tuan, R.S. Expression and functional involvement of N-cadherin in embryonic limb chondrogenesis. Development 1994, 120, 177–187. [Google Scholar]
- Lefort, C.T.; Wojciechowski, K.; Hocking, D.C. N-cadherin cell-cell adhesion complexes are regulated by fibronectin matrix assembly. J. Biol. Chem. 2011, 286, 3149–3160. [Google Scholar] [CrossRef]
- Dewit, J.; Witten, P.; Willems, M.; Huysseune, A. The distribution of fibronectin in developing zebrafish (Danio rerio) cartilage. J. Appl. Ichthyol. 2010, 26, 205–209. [Google Scholar] [CrossRef]
- Goetz, S.C.; Brown, D.D.; Conlon, F.L. TBX5 is required for embryonic cardiac cell cycle progression. Development 2006, 133, 2575–2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govindan, J.; Iovine, M.K. Hapln1a is required for connexin43-dependent growth and patterning in the regenerating fin skeleton. PLoS ONE 2014, 9, e88574. [Google Scholar] [CrossRef] [PubMed]
- Iovine, M.K.; Higgins, E.P.; Hindes, A.; Coblitz, B.; Johnson, S.L. Mutations in connexin43 (GJA1) perturb bone growth in zebrafish fins. Dev. Biol. 2005, 278, 208–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genbank Accession | Gene Symbol | Gene Name | Biological Process | Log2 Expression Ratio | Stages |
|---|---|---|---|---|---|
| NM_131038 | cx43 | connexin 43 | fin development | −1.98 | 24 |
| NM_001080012 | hhip | hedgehog interacting protein | fin development | −1.63 | 24 |
| NM_001077145 | bbs7 | Bardet-Biedl syndrome 7 | fin development | −2.02 | 24 |
| NM_194416 | mmp14a | matrix metalloproteinase 14 (membrane-inserted) alpha | cartilage development | −1.67 | 24 |
| NM_199777 | sec23b | Sec23 homolog B (S. cerevisiae) | cartilage development | −1.88 | 24 |
| NM_176859 | tfap2a | transcription factor AP-2 alpha | cartilage development | −2.12 | 24 |
| NM_131503 | axin 1 | axin 1 | fin development | −1.71 | 30 |
| NM_131260 | msxb | muscle segment homeobox B | fin development | −1.61 | 30 |
| NM_001005771 | hoxc8a | homeo box C8a | fin development | −1.93 | 30 |
| NM_205635 | slc35b2 | solute carrier family 35, member B2 | cartilage development | −1.89 | 30 |
| NM_131306 | dlx5a | distal-less homeobox gene 5a | cartilage development | −1.52 | 30 |
| NM_131305 | dlx1a | distal-less homeobox gene 1a | cartilage development | −1.84 | 30 |
| NM_001024665 | tfap2b | transcription factor AP-2 beta | cartilage development | −1.77 | 30 |
| NM_152963 | fmr1 | fragile X mental retardation 1 | cartilage development | −1.53 | 30 |
| BC163560 | runx3 | runt-related transcription factor 3 | cartilage development | −1.65 | 30 |
| NM_131081 | cdh2 | cadherin 2, neuronal | cartilage condensation | −1.94 | 30 |
| NM_001126448 | lect1 | leukocyte cell derived chemotaxin 1 | cartilage development | −1.75 | 30 |
| NM_001110278 | acvr2a | activin receptor IIa | cartilage development | −1.99 | 30 |
| NM_194414 | mmp14b | matrix metalloproteinase 14 (membrane-inserted) beta | cartilage development | −1.87 | 30 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, J.-K.; Tsai, T.-C.; Lee, H.; Hsia, K.; Lin, C.-H.; Lu, J.-H. Pectoral Fin Anomalies in tbx5a Knockdown Zebrafish Embryos Related to the Cascade Effect of N-Cadherin and Extracellular Matrix Formation. J. Dev. Biol. 2019, 7, 15. https://0-doi-org.brum.beds.ac.uk/10.3390/jdb7030015
Lu J-K, Tsai T-C, Lee H, Hsia K, Lin C-H, Lu J-H. Pectoral Fin Anomalies in tbx5a Knockdown Zebrafish Embryos Related to the Cascade Effect of N-Cadherin and Extracellular Matrix Formation. Journal of Developmental Biology. 2019; 7(3):15. https://0-doi-org.brum.beds.ac.uk/10.3390/jdb7030015
Chicago/Turabian StyleLu, Jenn-Kan, Tzu-Chun Tsai, Hsinyu Lee, Kai Hsia, Chih-Hsun Lin, and Jen-Her Lu. 2019. "Pectoral Fin Anomalies in tbx5a Knockdown Zebrafish Embryos Related to the Cascade Effect of N-Cadherin and Extracellular Matrix Formation" Journal of Developmental Biology 7, no. 3: 15. https://0-doi-org.brum.beds.ac.uk/10.3390/jdb7030015