Differentiation of Hedyotis diffusa and Common Adulterants Based on Chloroplast Genome Sequencing and DNA Barcoding Markers

 ,
, 
Abstract
:1. Introduction
2. Results
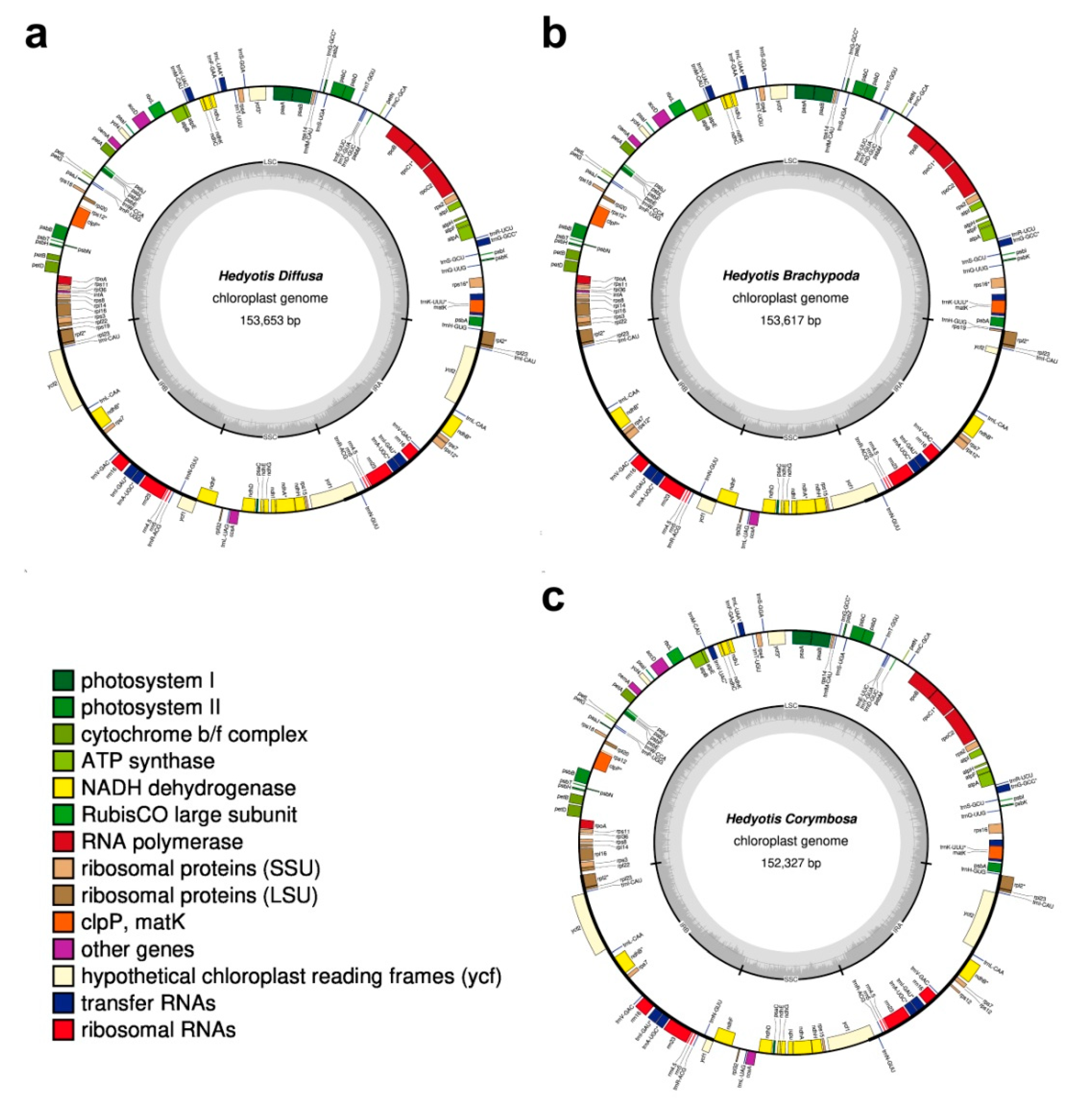
2.1. Chloroplast Genome Organization
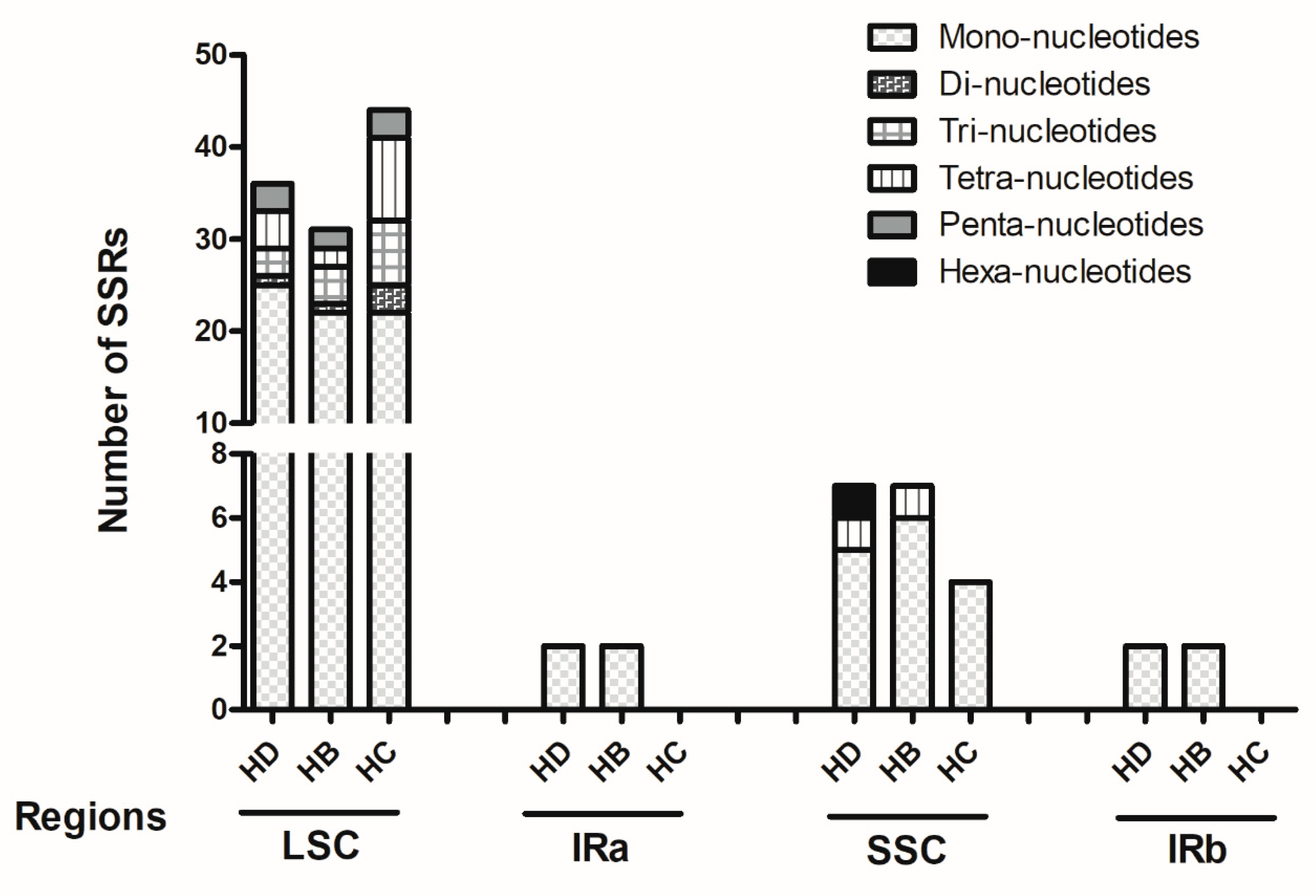
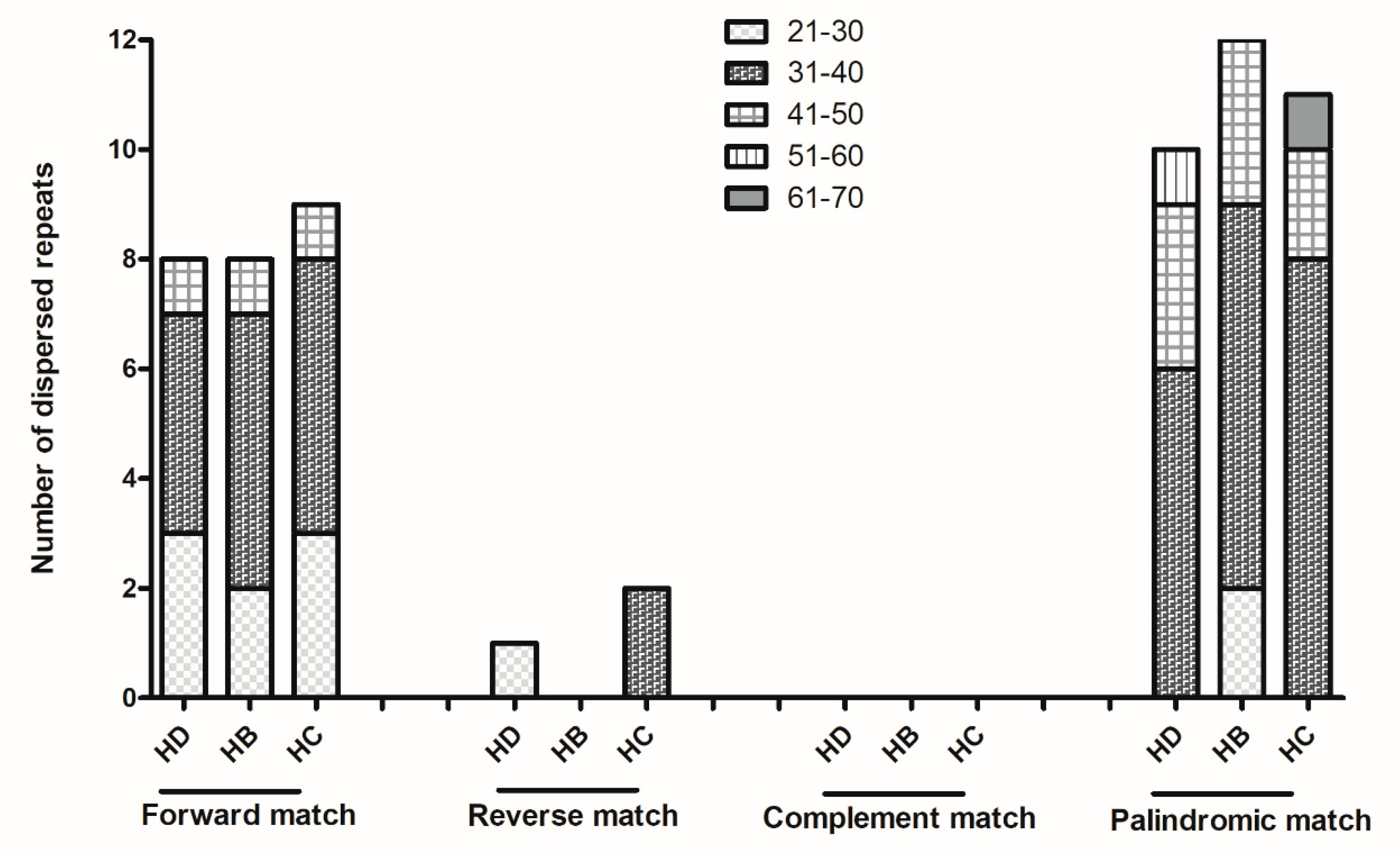
2.2. Analysis of Repeat Sequences
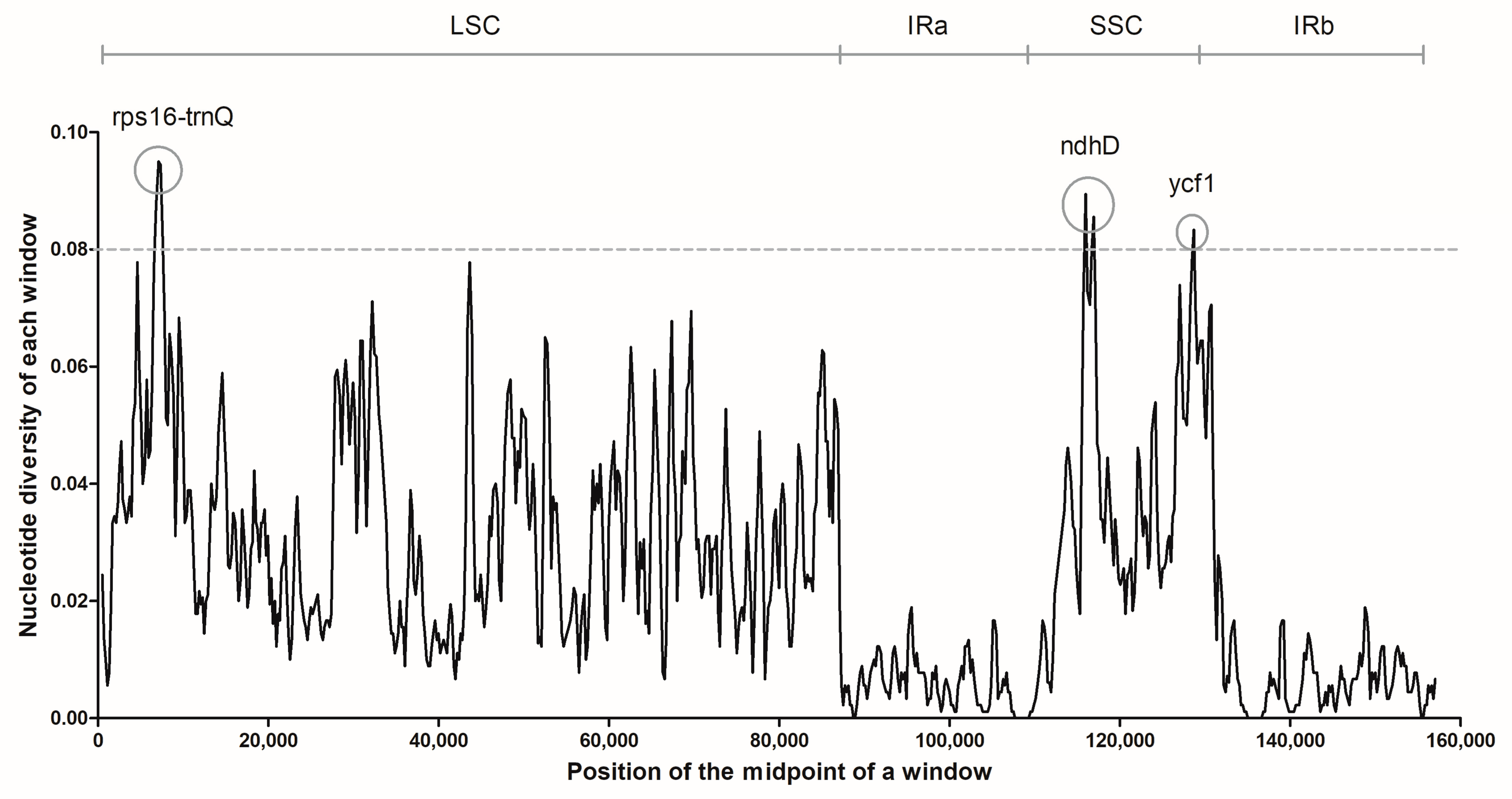
2.3. Genome Sequence Divergence Analysis
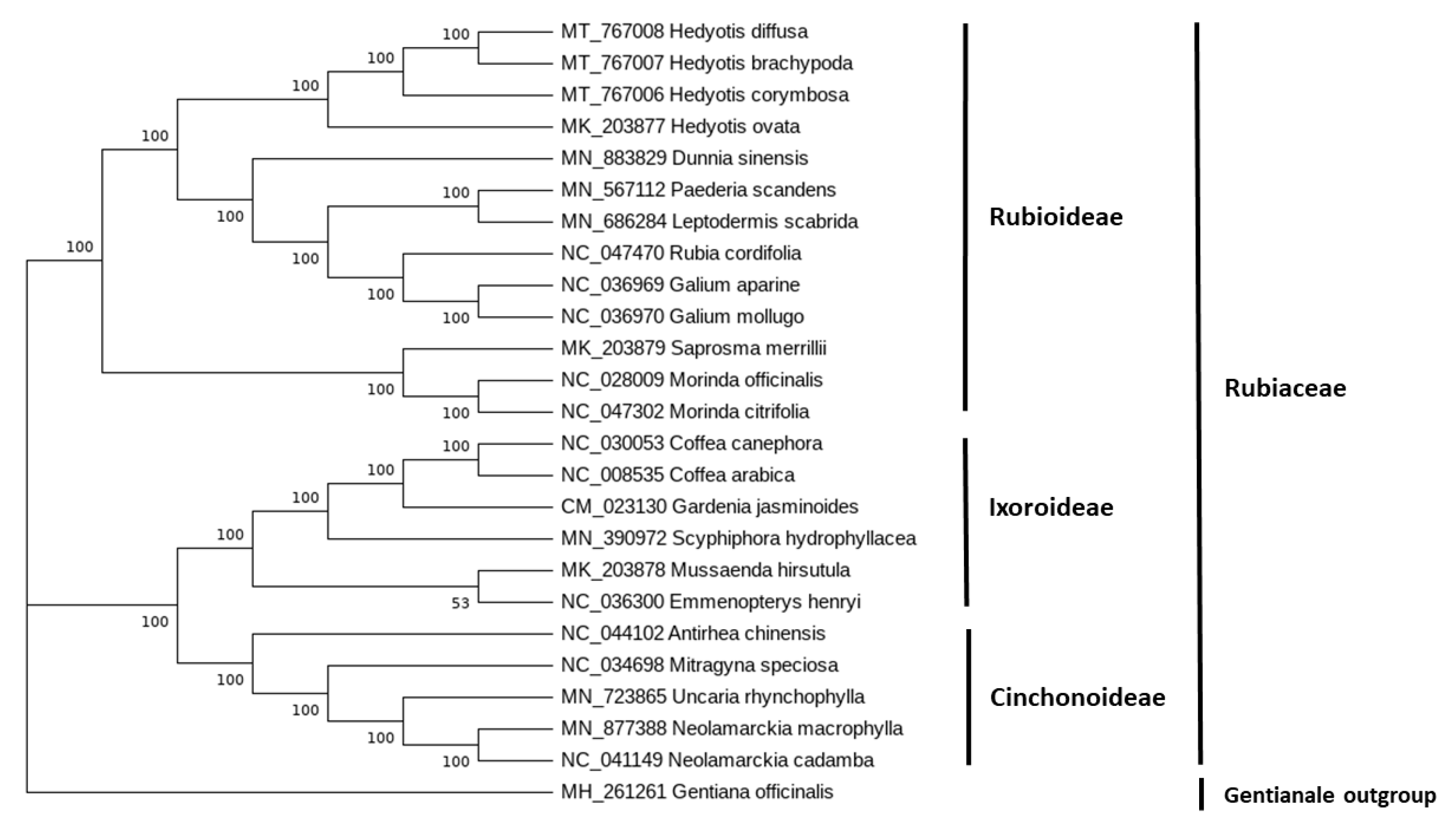
2.4. Phylogenetic Analysis
2.5. Development of DNA Barcoding Markers
3. Discussion
3.1. Chloroplast Genome Features
3.2. Sequence Divergence, Phylogenetic Relationships and Molecular Markers for Authentication
4. Materials and Methods
4.1. Collection of Plant Samples
4.2. DNA Extraction
4.3. Chloroplast Genome Sequencing, Assembling and Annotation
4.4. Repeat Elements Analysis
4.5. Sequence Divergence Analysis
4.6. Phylogenetic Analysis
4.7. Development of DNA Barcoding Markers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, Y.-C.; Lu, F.-H.; Wu, J.-S.; Wu, C.-H.; Chang, C.-J. The Protective Effect of Habitual Tea Consumption on Hypertension. Arch. Intern. Med. 2004, 164, 1534–1540. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Taylor, C.M. Hedyotis. Flora China 2011, 19, 147–174. [Google Scholar]
- Groeninckx, I.; Dessein, S.; Ochoterena, H.; Persson, C.; Motley, T.J.; Kårehed, J.; Bremer, B.; Huysmans, S.; Smets, E. Phylogeny of the Herbaceous Tribe Spermacoceae (Rubiaceae) Based on Plastid DNA Data1. Ann. Mo. Bot. Gard. 2009, 96, 109–132. [Google Scholar] [CrossRef]
- Nimal Christhudas, I.V.; Praveen Kumar, P.; Sunil, C.; Vajravijayan, S.; Lakshmi Sundaram, R.; Jenifer Siril, S.; Agastian, P. In vitro studies on alpha-glucosidase inhibition, antioxidant and free radical scavenging activities of Hedyotis biflora L. Food Chem. 2013, 138, 1689–1695. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; Ali, A.M.; Israf, D.A.; Ismail, N.H.; Shaari, K.; Lajis, N.H. Antioxidant, radical-scavenging, anti-inflammatory, cytotoxic and antibacterial activities of methanolic extracts of some Hedyotis species. Life Sci. 2005, 76, 1953–1964. [Google Scholar] [CrossRef]
- Ye, M.; Su, J.J.; Liu, S.T.; Cao, L.; Xiong, J.; Zhao, Y.; Fan, H.; Yang, G.X.; Xia, G.; Hu, J.F. (24S)-ergostane-3beta,5alpha,6beta-triol from Hedyotis chrysotricha with inhibitory activity on migration of SK-HEP-1 human hepatocarcinoma cells. Nat. Prod. Res. 2013, 27, 1136–1140. [Google Scholar] [CrossRef]
- Shan, B.E.; Zhang, J.Y.; Du, X.N. Immunomodulatory activity and anti-tumor activity of Oldenlandia diffusa in vitro. Zhongguo Zhong xi yi jie he za zhi Zhongguo Zhongxiyi jiehe zazhi Chin. J. Integr. Tradit. West. Med. 2001, 21, 370–374. [Google Scholar]
- Yan, Z.; Feng, J.; Peng, J.; Lai, Z.; Zhang, L.; Jin, Y.; Yang, H.; Chen, W.; Lin, J. Chloroform extract of Hedyotis diffusa Willd inhibits viability of human colorectal cancer cells via suppression of AKT and ERK signaling pathways. Oncol. Lett. 2017, 14, 7923–7930. [Google Scholar] [CrossRef] [Green Version]
- Yue, G.G.-L.; Lee, J.K.-M.; Chan, B.C.-L.; Kwok, H.-F.; Hoi, S.W.-H.; Sze, D.M.-Y.; Fung, K.-P.; Leung, P.-C.; Lau, C.B.-S. An innovative anti-cancer Chinese herbal formula exhibited multi-targeted efficacies in metastatic breast cancer mouse model. Chin. Med. 2018, 13, 64. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Lai, Z.; Yan, Z.; Peng, J.; Jin, Y.; Wei, L.; Lin, J. Hedyotis diffusa Willd inhibits proliferation and induces apoptosis of 5FU resistant colorectal cancer cells by regulating the PI3K/AKT signaling pathway. Mol. Med. Rep. 2018, 17, 358–365. [Google Scholar]
- Lau, C.B.S.; Cheng, L.; Cheng, B.W.H.; Yue, G.G.L.; Wong, E.C.W.; Lau, C.-P.; Leung, P.-C.; Fung, K.-P. Development of a simple chromato-graphic method for distinguishing between two easily confused species, Hedyotis diffusa and Hedyotis corymbosa. Nat. Prod. Res. 2012, 26, 1446–1450. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Jiang, R.-W.; Hon, P.-M.; Cheng, L.; Li, L.-L.; Zhou, J.-R.; Shaw, P.-C.; But, P.P.-H. Authentication of the anti-tumor herb Baihuash-eshecao with bioactive marker compounds and molecular sequences. Food Chem. 2010, 119, 1239–1245. [Google Scholar] [CrossRef]
- Song, Y.; Chen, Y.; Lv, J.; Xu, J.; Zhu, S.; Li, M. Comparative Chloroplast Genomes of Sorghum Species: Sequence Divergence and Phylogenetic Relationships. BioMed Res. Int. 2019, 2019, 5046958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, W.; Xu, C.; Wu, P.; Cheng, T.; Yu, J.; Zhou, S.-L.; Hong, D.-Y. Resolving the systematic positions of enigmatic taxa: Manipulating the chloroplast genome data of Saxifragales. Mol. Phylogenetics Evol. 2018, 126, 321–330. [Google Scholar] [CrossRef]
- Moore, M.J.; Bell, C.D.; Soltis, P.S.; Soltis, D.E. Using plastid genome-scale data to resolve enigmatic relationships among basal angi-osperms. Proc. Natl. Acad. Sci. USA 2007, 104, 19363–19368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, W.; Liu, J.; Yu, J.; Wang, L.; Zhou, S. Highly Variable Chloroplast Markers for Evaluating Plant Phylogeny at Low Taxonomic Levels and for DNA Barcoding. PLoS ONE 2012, 7, e35071. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Xu, C.; Cheng, T.; Lin, K.; Zhou, S. Sequencing Angiosperm Plastid Genomes Made Easy: A Complete Set of Universal Primers and a Case Study on the Phylogeny of Saxifragales. Genome Biol. Evol. 2013, 5, 989–997. [Google Scholar] [CrossRef] [Green Version]
- Raman, G.; Park, S. The Complete Chloroplast Genome Sequence of Ampelopsis: Gene Organization, Comparative Analysis, and Phylogenetic Relationships to Other Angiosperms. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Wang, J.; Jia, Y.; Li, W.; Xu, F.; Wang, X. Comparative Chloroplast Genome Analyses of Species in Gentiana section Cruciata (Gentianaceae) and the Development of Authentication Markers. Int. J. Mol. Sci. 2018, 19, 1962. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Zuo, L.; Lu, D.; Lu, B.; Yang, M.; Wang, J. Comparative analysis of chloroplast genomes of five Robinia species: Genome comparative and evolution analysis. Gene 2019, 689, 141–151. [Google Scholar] [CrossRef]
- Lu, R.-S.; Li, P.; Qiu, Y. The Complete Chloroplast Genomes of Three Cardiocrinum (Liliaceae) Species: Comparative Genomic and Phylogenetic Analyses. Front. Plant Sci. 2017, 7, 2054. [Google Scholar] [CrossRef] [PubMed]
- Maier, R.M.; Neckermann, K.; Igloi, G.L.; Kössel, H. Complete Sequence of the Maize Chloroplast Genome: Gene Content, Hotspots of Divergence and Fine Tuning of Genetic Information by Transcript Editing. J. Mol. Biol. 1995, 251, 614–628. [Google Scholar] [CrossRef] [PubMed]
- Hupfer, H.; Swiatek, M.; Hornung, S.; Herrmann, R.G.; Maier, R.M.; Chiu, W.-L.; Sears, B. Complete nucleotide sequence of the Oenothera elata plastid chromosome, representing plastome I of the five distinguishable Euoenothera plastomes. Mol. Genet. Genom. 2000, 263, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Menezes, A.P.A.; Resende-Moreira, L.C.; Buzatti, R.S.O.; Nazareno, A.G.; Carlsen, M.; Lobo, F.P.; Kalapothakis, E.; Lovato, M.B. Chloroplast genomes of Byrsonima species (Malpighiaceae): Comparative analysis and screening of high divergence sequences. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raman, G.; Park, S. Analysis of the Complete Chloroplast Genome of a Medicinal Plant, Dianthus superbus var. longicalyncinus, from a Comparative Genomics Perspective. PLoS ONE 2015, 10, e0141329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, M.W.; Covello, P.S. RNA editing in plant mitochondria and chloroplasts. FASEB J. 1993, 7, 64–71. [Google Scholar] [CrossRef]
- Weng, M.L.; Blazier, J.C.; Govindu, M.; Jansen, R.K. Reconstruction of the ancestral plastid genome in Geraniaceae reveals a corre-lation between genome rearrangements, repeats, and nucleotide substitution rates. Mol. Biol. Evol. 2014, 31, 645–659. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Dong, W.; Li, W.; Lu, Y.; Xie, X.; Jin, X.; Shi, J.; He, K.; Suo, Z. Comparative Analysis of Six Lagerstroemia Complete Chloroplast Genomes. Front. Plant Sci. 2017, 8, 15. [Google Scholar] [CrossRef] [Green Version]
- Nazareno, A.G.; Carlsen, M.; Lohmann, L.G. Complete Chloroplast Genome of Tanaecium tetragonolobum: The First Bignoniaceae Plastome. PLoS ONE 2015, 10, e0129930. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Du, L.; Liu, A.; Chen, J.; Wu, L.; Hu, W.; Zhang, W.; Kim, K.; Lee, S.C.; Yang, T.J.; et al. The Complete Chloroplast Genome Se-quences of Five Epimedium Species: Lights into Phylogenetic and Taxonomic Analyses. Front Plant Sci. 2016, 7, 306. [Google Scholar]
- Khakhlova, O.; Bock, R. Elimination of deleterious mutations in plastid genomes by gene conversion. Plant J. 2006, 46, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, Y.; Henry, R.J.; Rossetto, M.; Wang, Y.; Chen, S. Plant DNA barcoding: From gene to genome. Biol. Rev. Camb. Philos. Soc. 2015, 90, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-B.; Tang, M.; Li, H.; Zhang, Z.; Li, D.Z. Complete chloroplast genome of the genus Cymbidium: Lights into the species identification, phylogenetic implications and population genetic analyses. BMC Evol. Biol. 2013, 13, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollingsworth, P.; Graham, S.W.; Little, D.P. Choosing and Using a Plant DNA Barcode. PLoS ONE 2011, 6, e19254. [Google Scholar] [CrossRef]
- But, G.W.-C.; Wu, H.-Y.; Shaw, P.C. Identification of fish species of sushi products in Hong Kong. Food Control 2019, 98, 164–173. [Google Scholar] [CrossRef]
- Lo, Y.-T.; Li, M.; Shaw, P.C. Identification of constituent herbs in ginseng decoctions by DNA markers. Chin. Med. 2015, 10, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Sivarajan, V.V.; Biju, S.D. Taxonomic and nomenclatural notes on the Hedyotis corymbosa-diffusa complex (Rubiaceae) in India. Taxonomy 1990, 39, 665–674. [Google Scholar] [CrossRef]
- Lohse, M.; Drechsel, O.; Kahlau, S.; Bock, R. OrganellarGenomeDRAW—a suite of tools for generating physical maps of plastid and mitochondrial genomes and visualizing expression data sets. Nucleic Acids Res. 2013, 41, W575–W581. [Google Scholar] [CrossRef]
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-web: A web server for microsatellite prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef] [Green Version]
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Plat-forms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Hedyotis diffusa (MT767008) | Hedyotis brachypoda (MT767007) | Hedyotis corymbosa (MT767006) |
|---|---|---|---|
| Specimen Voucher | H2200 | H2174 | H2106 |
| Total Size (bp) | 153,653 | 153,617 | 152,327 |
| Large Single Copy (LSC, bp) | 83,863 | 83,780 | 83,609 |
| Small Single Copy (SSC, bp) | 17,218 | 17,195 | 18,146 |
| Inverted Repeat (IR, bp) | 26,286 | 26,321 | 25,286 |
| Total number of genes | 130 | 129 | 128 |
| Protein-coding genes | 85 | 84 | 83 |
| tRNA genes | 37 | 37 | 37 |
| rRNA genes | 8 | 8 | 8 |
| GC content (%) | 37.62 | 37.61 | 37.41 |
| Category | Gene Group | Gene Name |
|---|---|---|
| Protein synthesis and DNA-replication | Ribosomal RNA genes | rrn4.5, rrn5, rrn16, rrn23 |
| Transfer RNA genes | trnH-GUG, trnL-UAG, trnQ-UUG, trnS-GCU, trnR-UCU, trnC-GCA, trnD-GUC, trnY-GUA, trnE-UUC, trnT-GGU, trnS-UGA, trnfM-CAU, trnS-GGA, trnT-UGU, trnF-GAA, trnM-CAU, trnW-CCA, trnP-UGG, trnV-UAC, trnL-UAA *, trnK-UUU *, trnG-GCC *, trnI-CAU, trnL-CAA, trnV-GAC, trnI-GAU *, trnA-UGC *, trnR-ACG, trnN-GUU | |
| Small subunit of ribosome | rps2, rps3, rps4, rps7, rps8, rps11, rps12 *, rps14, rps15, rps16 *, rps18, rps19 δ | |
| Large subunit of ribosome | rpl2 *, rpl14, rpl16, rpl20, rpl22, rpl23, rpl32, rpl36 | |
| RNA polymerase subunits | rpoA, rpoB, rpoC1 *, rpoC2 | |
| Photosynthesis | NADH dehydrogenase | ndhA *, ndhB *, ndhC, ndhD, ndhE, ndhF, ndhG, ndhH, ndhI, ndhJ, ndhK |
| Photosystem I | psaA, psaB, psaC, psaI, psaJ, ycf3 ** | |
| Photosystem II | psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbN, psbT, psbZ | |
| Cytochrome b/f complex | petA, petB, petD, petG, petL, petN | |
| ATP synthase | atpA, atpB, atpE, atpF, atpH, atpI | |
| Large subunit of rubisco | rbcL | |
| Miscellaneous group | Maturase | matK |
| Protease | clpP ** | |
| Envelope membrane protein | cemA | |
| Subunit of acetyl-CoA-carboxylase | accD | |
| c-type cytochrome synthesis | ccsA | |
| Component of TIC complex | ycf1Ψ | |
| Translation initiation factor | infA ω | |
| Hypothetical chloroplast reading frames | ycf2σ | |
| Pseudogene unknown function | ORFs | ycf4 |
| Hedyotis diffusa | Hedyotis brachypoda | Hedyotis corymbosa | |
|---|---|---|---|
| Hedyotis diffusa | 1051 | 4996 | |
| Hedyotis brachypoda | 198 | 5061 | |
| Hedyotis corymbosa | 684 | 677 |
| Specimen Voucher | Percentage Identity with ndhD in Chloroplast Genome | Percentage Identity with rps16-trnQ in Chloroplast Genome | Sample Identity | ||||
|---|---|---|---|---|---|---|---|
| H. corymbosa | H. brachypoda | H. diffusa | H. corymbosa | H. brachypoda | H. diffusa | ||
| T5084 | 100% | 97% | 97% | 99% | 85% | 85% | H. corymbosa |
| T5089 | 97% | 100% | 99% | 86% | 97% | 96% | H. brachypoda |
| T5093 | 100% | 97% | 97% | 99% | 87% | 86% | H. corymbosa |
| T5097 | 100% | 97% | 97% | 99% | 87% | 87% | H. corymbosa |
| T5101 | 97% | 100% | 99% | 87% | 98% | 97% | H. brachypoda |
| T5106 | 100% | 97% | 97% | 100% | 87% | 87% | H. corymbosa |
| T5110 | 97% | 100% | 99% | 87% | 98% | 97% | H. brachypoda |
| T5114 | 97% | 100% | 99% | 87% | 98% | 97% | H. brachypoda |
| T5121 | 97% | 100% | 99% | 87% | 98% | 97% | H. brachypoda |
| T5126 | 97% | 100% | 99% | 87% | 98% | 97% | H. brachypoda |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yik, M.H.-Y.; Kong, B.L.-H.; Siu, T.-Y.; Lau, D.T.-W.; Cao, H.; Shaw, P.-C. Differentiation of Hedyotis diffusa and Common Adulterants Based on Chloroplast Genome Sequencing and DNA Barcoding Markers. Plants 2021, 10, 161. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10010161
Yik MH-Y, Kong BL-H, Siu T-Y, Lau DT-W, Cao H, Shaw P-C. Differentiation of Hedyotis diffusa and Common Adulterants Based on Chloroplast Genome Sequencing and DNA Barcoding Markers. Plants. 2021; 10(1):161. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10010161
Chicago/Turabian StyleYik, Mavis Hong-Yu, Bobby Lim-Ho Kong, Tin-Yan Siu, David Tai-Wai Lau, Hui Cao, and Pang-Chui Shaw. 2021. "Differentiation of Hedyotis diffusa and Common Adulterants Based on Chloroplast Genome Sequencing and DNA Barcoding Markers" Plants 10, no. 1: 161. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10010161