Identification of Tomato Ve1 Homologous Proteins in Flax and Assessment for Race-Specific Resistance in Two Fiber FlaxCultivars against Verticillium dahliae Race 1

 ,
,  ,
,
Abstract
:1. Introduction
2. Methods
2.1. Ve1 Homologous Proteins Screening, Phylogeny and Primary Structure Analysis
2.2. Ve1 Homologous Proteins Phylogeny
2.3. Ve1 Homologous Proteins Primary Structure Analysis
2.4. Plant Material
2.5. Pathogen Strain
2.6. Inoculation Procedure and Plants Cultivation
2.7. Symptoms and Pathogen Observation in Plants
2.8. Plant DNA Extraction and Quantification
2.9. Plant RNA Extraction, Quality Control and cDNA Synthesis
2.10. Detection of the Ve1 Homologous Genes Using PCR
2.11. LuVe11 Relative Expression by Real-Time PCR
3. Results
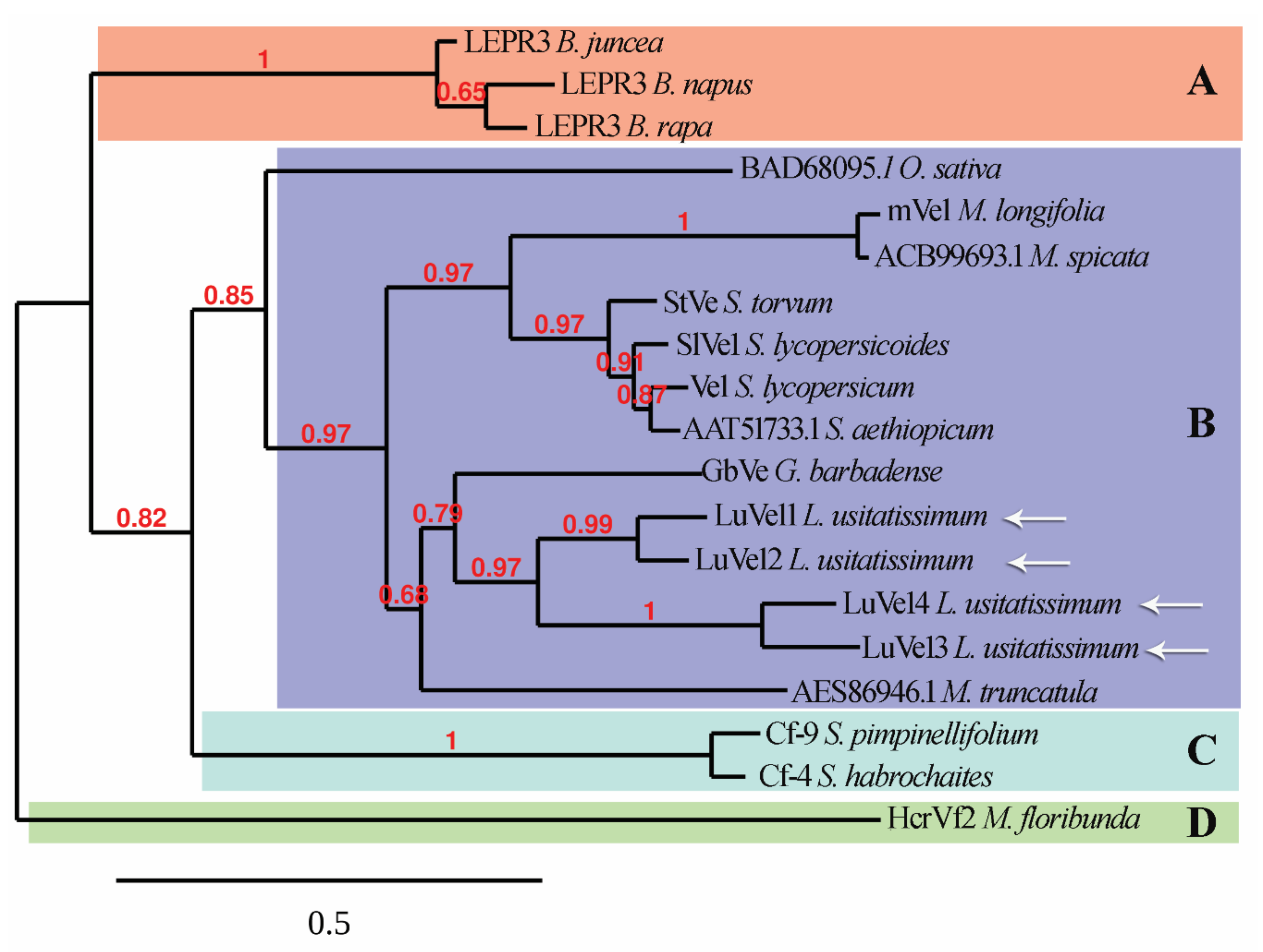
3.1. In Silico Analysis Reveals Ve1 Homologs in Flax
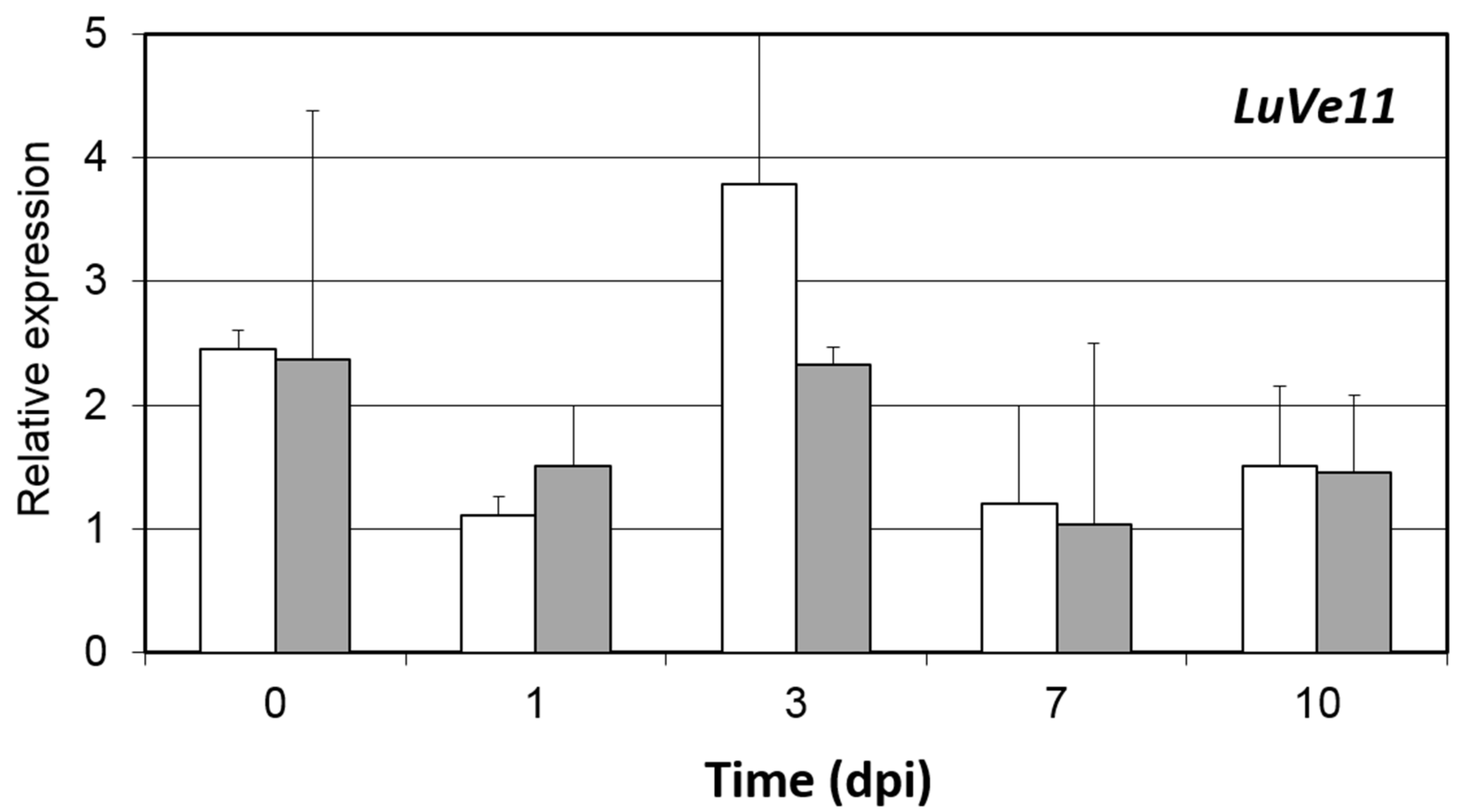
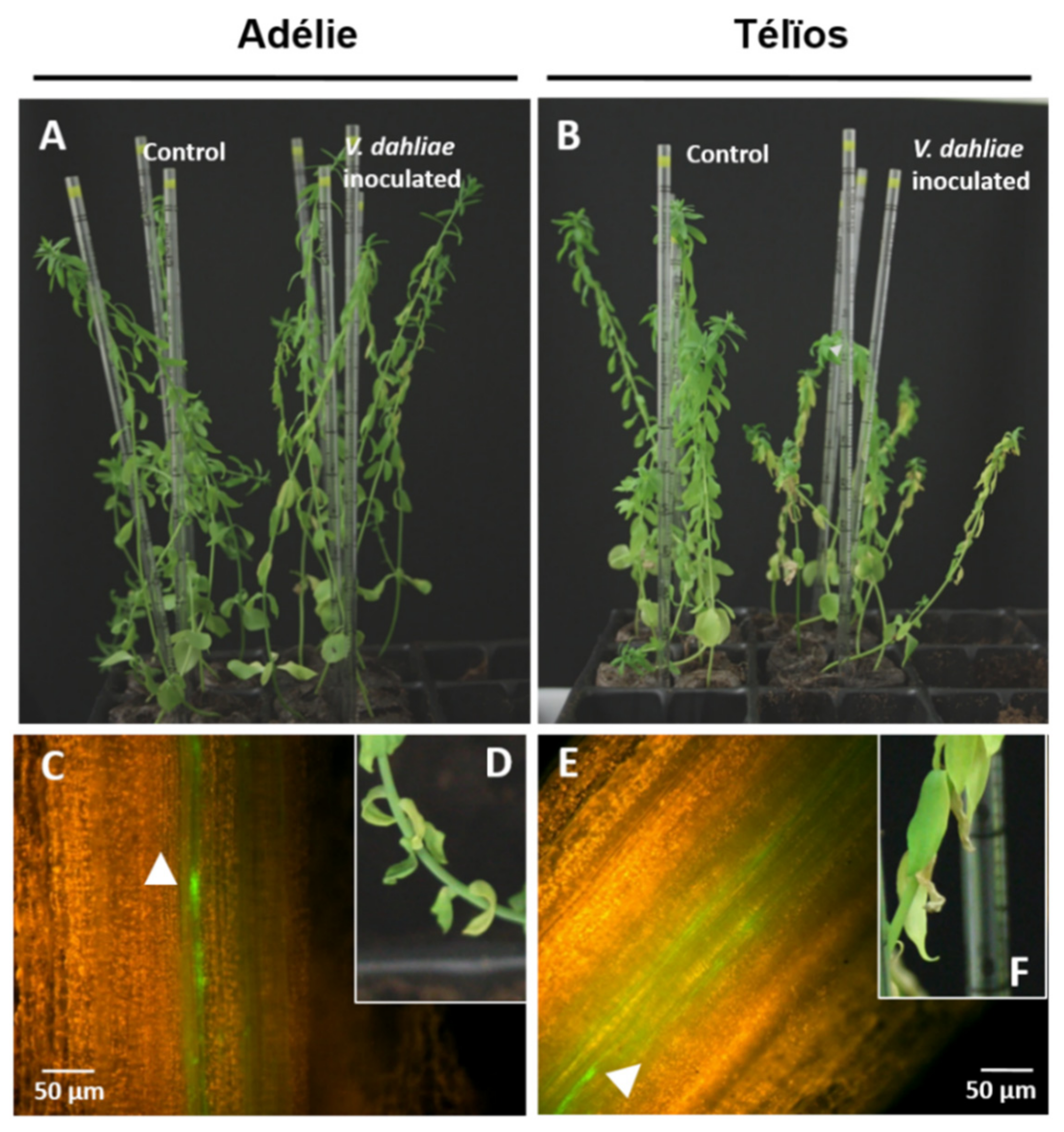
3.2. Disease Onset and Pathogen Colonization despite the Presence of Ve1 Homologous Genes and the Expression of LuVe11
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Agrios, G.N. Plant Pathology; Academic Press: San Diego, CA, USA, 1997; 635p. [Google Scholar]
- Belfanti, E.; Silfverberg-Dilworth, E.; Tartarini, S.; Patocchi, A.; Barbieri, M.; Zhu, J.; Vinatzer, B.A.; Gianfranceschi, L.; Gessler, C.; Sansavini, S. The HcrVf2 gene from a wild apple confers scab resistance to a transgenic cultivated variety. Proc. Natl. Acad. Sci. USA 2004, 101, 886–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, A.; Bressan, M.; Zahid, A.; Trinsoutrot-Gattin, I.; Driouich, A.; Laval, K. Verticillium Wilt on fiber flax: Symptoms and pathogen development in planta. Plant Dis. 2018, 102, 2421–2429. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.; Zhao, L.; Liao ZSun, X.; Zuo, K.; Zhang, L.; Wang, S.; Tang, K. Molecular cloning of a potential Verticillium dahliae resistance gene SlVe1 with multi-site polyadenylation from Solanum lycopersicoides. DNA Seq. 2003, 14, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Chinchilla, D.; Zipfel, C.; Robatzek, S.; Kemmerling, B.; Nürnberger, T.; Jones, J.D.G.; Felix, G.; Boller, T. A flagellin-induced complex of the receptor FLS2 and BAK1 initiates plant defence. Nature 2007, 448, 497–500. [Google Scholar] [CrossRef] [PubMed]
- De Jonge, R.; van Esse, H.P.; Maruthachalam, K.; Bolton, M.D.; Santhanam, P.; Saber, M.K.; Zhang, Z.; Usami, T.; Lievens, B.; Subbarao, K.V. Tomato immune receptor Ve1 recognizes effector of multiple fungal pathogens uncovered by genome and RNA sequencing. Proc. Natl. Acad. Sci. USA 2012, 109, 5110–5115. [Google Scholar] [CrossRef] [Green Version]
- Dereeper, A.; Audic, S.; Claverie, J.M.; Blanc, G. BLAST-EXPLORER helps you building datasets for phylogenetic analysis. BMC Evol. Biol. 2010, 12, 8–10. [Google Scholar] [CrossRef] [Green Version]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef]
- Diederichsen, A. Ex situ collections of cultivated flax (Linum usitatissimum L.) and other species of the genus Linum L. Genet. Resour. Crop. Evol. 2007, 54, 661–678. [Google Scholar] [CrossRef]
- Diwan, N.; Fluhr, R.; Eshed, Y.; Zamir, D.; Tanksley, S.D. Mapping of Ve in tomato: A gene conferring resistance to the broad-spectrum pathogen, Verticillium dahliae race 1. Theor. Appl. Genet. 1999, 98, 315–319. [Google Scholar] [CrossRef]
- Fei, J.; Chai, Y.R.; Wang, J.; Lin, J.; Sun, X.F.; Sun, C.; Zuo, K.J.; Tang, K.X. cDNA cloning and characterization of the Ve homologue gene StVe from Solanum torvum Swartz. DNA Seq. 2004, 15, 88–95. [Google Scholar] [CrossRef]
- Fradin, E.F.; Abd-El-Haliem, A.; Masini, L.; van den Berg, G.C.; Joosten, M.H.; Thomma, B.P. Interfamily transfer of tomato Ve1 mediates Verticillium resistance in Arabidopsis. Plant Physiol. 2011, 156, 2255–2265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fradin, E.F.; Thomma, B.P.H.J. Physiology and molecular aspects of Verticillium wilt diseases caused by V. dahliae and V. alboatrum. Mol. Plant Pathol. 2006, 7, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Fradin, E.F.; Zhang, Z.; Ayala, J.C.J.; Castroverde, C.D.M.; Nazar, R.N.; Robb, J.; Liu, C.M.; Thomma, B.P.H.J. Genetic dissection of Verticillium wilt resistance mediated by tomato Ve1. Plant Physiol. 2009, 150, 320–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fradin, E.F.; Zhang, Z.; Rovenich, H.; Song, Y.; Liebrand, T.W.H.; Masini, L.; van den Berg, G.C.M.; Joosten, M.H.A.J.; Thomma, B.P.H.J. Functional analysis of the tomato immune receptor Ve1 through domain swaps with its non-functional homolog Ve2. PLoS ONE 2014, 9, e88208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grogan, R.; Ioannou, N.; Schneider, R.; Sall, M.; Kimble, K. Verticillium wilt on resistant tomato cultivars in California: Virulence of isolates from plants and soil and relationship of inoculum density to disease incidence. Phytopathology 1979, 69, 1176–1180. [Google Scholar] [CrossRef] [Green Version]
- Hayes, R.J.; McHale, L.K.; Vallad, G.E.; Truco, M.J.; Michelmore, R.W.; Klosterman, S.J.; Maruthachalam, K.; Subbarao, K.V. The inheritance of resistance to Verticillium wilt caused by race 1 isolates of Verticillium dahliae in the lettuce cultivar La Brillante. Theor Appl. Genet. 2011, 123, 509–517. [Google Scholar] [CrossRef]
- Huis, R.; Hawkins, S.; Neutelings, G. Selection of reference genes for quantitative gene expression normalization in flax (Linum usitatissimum L.). BMC Plant Boil. 2010, 10, 71. [Google Scholar] [CrossRef] [Green Version]
- Inderbitzin PSubbarao, K.V. Verticillium Systematics and Evolution: How confusion impedes Verticillium Wilt management and how to resolve it. Phytopathology 2014, 104, 564–574. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.; Trotochaud, A.E.; Clark, S.E. The Arabidopsis CLAVATA2 gene encodes a receptor-like protein required for the stability of the CLAVATA1 receptor-like kinase. Plant Cell 1999, 11, 1925–1933. [Google Scholar]
- Jones, D.A.; Thomas, C.M.; Hammond-Kosack, K.E.; Balint-Kurti, P.J.; Jones, J.D.G. Isolation of the tomato Cf-9 gene for resistance to Cladosporium fulvum by transposon tagging. Science 1994, 266, 789–793. [Google Scholar] [CrossRef]
- Käll, L.; Krogh, A.; Sonnhammer, E.L. A combined transmembrane topology and signal peptide prediction method. J. Mol. Biol. 2004, 338, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Kawchuk, L.M.; Hachey, J.; Lynch, D.R.; Kulcsar, F.; van Rooijen, G.; Waterer, D.R.; Robertson, A.; Kokko, E.; Byers, R.; Howard, B.J.; et al. Tomato Ve disease resistance genes encode cell surface-like receptors. Proc. Natl. Acad. Sci. USA 2001, 98, 6511–6515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klosterman, S.J.; Atallah, Z.K.; Vallad, G.E.; Subbarao, K.V. Diversity pathogenicity and management of Verticillium species. Annu. Rev. Phytopathol. 2009, 47, 39–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koressaar, T.; Remm, M. Enhancements and modifications of primer design program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruijt, M.; De Kock, M.J.D.; de Wit, P.J.G.M. Receptor-like proteins involved in plant disease resistance. Mol. Plant Pathol. 2005, 6, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Kulpa, W.; Danert, S. Zur Systematik von Linum usitatissimum L. Kulturpflanze 1962, 3, 341–388. [Google Scholar]
- Larkan, N.J.; Lydiate, D.J.; Parkin, I.A.; Nelson, M.N.; Epp, D.J.; Cowling, W.A.; Rimmer, S.R.; Borhan, M.H. The Brassica napus blackleg resistance gene LepR3 encodes a receptor-like protein triggered by the Leptosphaeria maculans effector AVRLM1. New Phytol. 2013, 197, 595–605. [Google Scholar] [CrossRef]
- Liu, S.P.; Zhu, Y.P.; Xie, C.; Jue, D.W.; Hong, Y.B.; Chen, M.; Hubdar, A.K.; Yang, Q. Transgenic potato plants expressing StoVe1 exhibit enhanced resistance to Verticillium dahliae. Plant Mol. Biol. Rep. 2012, 30, 1032–1039. [Google Scholar] [CrossRef]
- Malnoy, M.; Xu, M.; Borejsza-Wysocka, E.; Korban, S.S.; Aldwinckle, H.S. Two receptor-like genes, Vfa1 and Vfa2, confer resistance to the fungal pathogen Venturia inaequalis inciting apple scab disease. Mol. Plant Microbe Interact. 2008, 21, 448–458. [Google Scholar] [CrossRef] [Green Version]
- Nürnberger, T.; Kemmerling, B. Receptor protein kinases: Pattern recognition receptors in plant immunity. Trends Plant Sci. 2006, 11, 519–522. [Google Scholar] [CrossRef]
- Ogawa, M.; Shinohara, H.; Sakagami, Y.; Matsubayashi, Y. Arabidopsis CLV3 peptide directly binds CLV1 ectodomain. Science 2008, 319, 294. [Google Scholar] [CrossRef] [PubMed]
- Pegg, G.; Brady, B. Verticillium Wilts; CAB International: Oxford, UK, 2002. [Google Scholar]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]
- Rashid, K.Y. Principal diseases of flax. In Flax, the Genus Linum; Muir, A., Westcott, N., Eds.; Taylor & Francis: London, UK, 2003; pp. 102–133. [Google Scholar]
- Rick, C.M.; Martin, F.M.; Gentile, A. Linkage of Verticillium resistance (Ve). Tomato Genet. Coop. 1959, 9, 44. [Google Scholar]
- Robb, J.; Shittu, H.; Soman, K.V.; Kurosky, A.; Nazar, R.N. Arsenal of elevated defense proteins fails to protect tomato against Verticillium dahliae. Planta 2012, 236, 623–633. [Google Scholar] [CrossRef]
- Rojo, E.; Sharma VKKovaleva, V.; Raikhel, N.V.; Fletcher, J.C. CLV3 is localized to the extracellular space, where it activates the Arabidopsis CLAVATA stem cell signaling pathway. Plant Cell 2002, 14, 969–977. [Google Scholar] [CrossRef] [Green Version]
- Rowland, G.G.; Hormis, Y.A.; Rashid, K.Y. CDC Bethune flax. Can. J. Plant Sci. 2002, 82, 101–102. [Google Scholar] [CrossRef]
- Schaible, L.; Cannon, O.S.; Waddoups, V. Inheritance of resistance to Verticillium wilt in a tomato cross. Phytopathology 1951, 41, 986–990. [Google Scholar]
- Schulze, B.; Mentzel, T.; Jehle, A.K.; Mueller, K.; Beeler, S.; Boller, T.; Felix, G.; Chinchilla, D. Rapid heteromerization and phosphorylation of ligand-activated plant transmembrane receptors and their associated kinaseBAK1. J. Biol. Chem. 2010, 285, 9444–9451. [Google Scholar] [CrossRef] [Green Version]
- Shepard, K.A.; Purugganan, M.D. Molecular population genetics of the Arabidopsis CLAVATA2 region the genomic scale of variation and selection in a selfing species. Genetics 2003, 163, 1083–1095. [Google Scholar]
- Simko, I.; Costanzo, S.; Haynes, K.G.; Ewing, E.E.; Costanzo, S.; Christ, B.J.; Jones, R.W. Linkage disequilibrium mapping of a Verticillium dahliae resistance quantitative trait locus in tetraploid potato (Solanum tuberosum) through a candidate gene approach. Theor. Appl. Genet. 2004, 108, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zhang, Z.; Seidl, M.F.; Majer, A.; Jakse, J.; Javornik, B.; Thomma, B.P. Broad taxonomic characterization of Verticillium wilt resistance genes reveals an ancient origin of the tomato Ve1 immune receptor. Mol. Plant Pathol. 2017, 18, 195–209. [Google Scholar] [CrossRef] [PubMed]
- Thomma, B.P.H.J.; van Esse, H.P.; Crous, P.W.; de Wit, P.J.G.M. Cladosporium fulvum (syn Passalora fulva), a highly specialized plant pathogen as a model for functional studies on plant pathogenic Mycosphaerellaceae. Mol. Plant Pathol. 2005, 6, 379–393. [Google Scholar] [CrossRef]
- Trotochaud, A.E.; Hao, T.; Wu, G.; Yang, Z.; Clark, S.E. The CLAVATA1 receptor-like kinase requires CLAVATA3 for its assembly into a signaling complex that includes KAPP and a Rho-related protein. Plant Cell 1999, 11, 393–406. [Google Scholar] [PubMed] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaisey-genser, M.; Morris, D.H. Introduction: History of the cultivation and uses of flaxseed. In Flax, the Genus Linum; Muir, A., Westcott, N., Eds.; Taylor & Francis: London, UK, 2003; pp. 102–133. [Google Scholar]
- Valade, R.; Cast, D.; Bert, F. Verticilliose du lin fibre, la prévention comme seule arme. Perspect. Agric. 2015, 424, 30–32. [Google Scholar]
- Vallad, G.E.; Qin, Q.M.; Grube, R.; Hayes, R.J.; Subbarao, K.V. Characterization of race-specific interactions among isolates of Verticillium dahliae pathogenic on lettuce. Phytopathology 2006, 96, 1380–1387. [Google Scholar] [CrossRef] [Green Version]
- Vallad, G.E.; Subbarao, K.V. Colonization of resistant and susceptible lettuce cultivars by a green fluorescent protein-tagged isolate of Verticillium dahliae. Phytopathology 2008, 98, 871–885. [Google Scholar] [CrossRef] [Green Version]
- Van Sumere, C. Retting of flax with special reference to enzyme-retting. In The Biology and Processing of Flax; Sharma, H.C., Van Sumere, C., Eds.; M Publications: Belfast, Northern Ireland, 1992; pp. 157–198. [Google Scholar]
- Vinatzer, B.A.; Patocchi, A.; Gianfranceschi, L.; Tartarini, S.; Zhang, H.B.; Gessler, C.; Sansavini, S. Apple contains receptor-like genes homologous to the Cladosporium fulvum resistance gene family of tomato with a cluster of genes cosegregating with Vf apple scab resistance. Mol. Plant Microbe Interact. 2001, 14, 508–515. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Ellendorff, U.; Kemp, B.; Mansfield, J.W.; Forsyth, A.; Mitchel, K.; Bastas, K.; Liu, C.M.; Woods-Tör, A.; Zipfel, C.; et al. A genome-wide functional investigation into the roles of receptor-like proteins in Arabidopsis. Plant Physiol. 2008, 147, 503–517. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Hobson, N.; Galindo, L.; Zhu, S.; Shi, D.; McDill, J.; Yang, L.; Hawkins, S.; Neutelings, G.; Datla, R.; et al. The genome of flax (Linum usitatissimum) assembled de novo from short shotgun sequence reads. Plant J. 2012, 72, 461–473. [Google Scholar] [CrossRef] [Green Version]
- Yadeta, K.A.; Thomma, B.P.H.J. The xylem as battleground for plant hosts and vascular wilt pathogens. Front. Plant Sci. 2013, 4, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelko, I.; Lux, A.; Sterckeman, T.; Martinka, M.; Kollárová, K.; Lišková, D. An easy method for cutting and fluorescent staining of thin roots. Ann. Bot. 2012, 110, 475–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Jian, G.; Jiang, T.; Wang, S.; Qi, F.; Xu, S. Cotton gene expression profiles in resistant Gossypium hirsutum cv Zhongzhimian KV1 responding to Verticillium dahliae strain V991 infection. Mol. Biol. Rep. 2012, 39, 9765–9774. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Jiang, T.; Cui, X.; Qi, F.; Jian, G. Colonization in cotton plants by a green fluorescent protein labelled strain of Verticillium dahliae. Eur. J. Plant Pathol. 2012, 135, 867–876. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Yang, S.; Chi, J.; Zhang, G.; Ma, Z. Cloning and characterization of a Verticillium wilt resistance gene from Gossypium barbadense and functional analysis in Arabidopsis thaliana. Plant Cell Rep. 2011, 30, 2085–2096. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Song, Y.; Liu, C.M.; Thomma, B.P.H.J. Mutational analysis of the Ve1 immune receptor that mediates Verticillium resistance in tomato. PLoS ONE 2014, 9, e99511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Zhao, Y.; Jin, Y.; Zhang, T.; Guo, H. Colonization process of Arabidopsis thaliana roots by a green fluorescent protein-tagged isolate of Verticillium dahliae. Protein Cell 2014, 5, 94–98. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Name | Primer Sequence (5′ -> 3′) | Amplicon Size (bp) | Reference |
|---|---|---|---|---|
| Ubiquitin extension protein 2 | LuUBI2_1F | 5′- CCAAGATCCAGGACAAGGAA -3′ | 129 | [46] |
| LuUBI2_1R | 5′- GAACCAGGTGGAGAGTCGAT -3′ | |||
| Elongation Factor 1-α | LuEF1A_1F | 5′- GATCGCCTGTCAATCTTGGT -3′ | 140 | |
| LuEF1A_1R | 5′- GCTGCCAACTTCACATCTCA -3′ | |||
| LuVe11 | LuVe11_1F | 5′- TTGGAAGCTTGACTGTCCTG -3′ | 122 | This study |
| LuVe11_1R | 5′- CCCTTGAAGCAATCCGTTAT -3′ | |||
| LuVe11_2F | 5′- GGATTCGGTGCAGTTACCTT -3′ | 69 | ||
| LuVe11_2R | 5′- GATCCTTGGCCAGTACCAGT -3′ | |||
| LuVe11_3F | 5′- TGAAACTTCGGTCTGTCTCG -3′ | 79 | ||
| LuVe11_3R | 5′- AAATCGGCAAAGAAATCAGG -3′ | |||
| LuVe12 | LuVe12_1F | 5′- CTTTCAACGAAACCATCCCT -3′ | 125 | |
| LuVe12_1R | 5′- TACAGCGAGAGAGGAAGCAA -3′ | |||
| LuVe12_2F | 5′- TCGAGGTAAGGCTCCTGAGT -3′ | 95 | ||
| LuVe12_2R | 5′- GAGGGATCGGACCTTGTAGA -3′ | |||
| LuVe12_3F | 5′- CCAAGTTCACCAGTTCATGG -3′ | 116 | ||
| LuVe12_3R | 5′- ATAAGAGCGCCTAAGCCAAA -3′ | |||
| LuVe13 | LuVe13_1F | 5′- CTGCTTGCATCACTGGACTT -3′ | 150 | |
| LuVe13_1R | 5′- AGGGATGGTTTCGTTGAAAG -3′ | |||
| LuVe13_2F | 5′- GGCAGCCTTAGCCAGTTATC -3′ | 146 | ||
| LuVe13_2R | 5′- AAGTCCAGTGATGCAAGCAG -3′ | |||
| LuVe13_3F | 5′- GGAACATTTCCTGCCAAGAT -3′ | 148 | ||
| LuVe13_3R | 5′- CACCGGAGAAGCTTGTGTAA -3′ | |||
| LuVe14 | LuVe14_1F | 5′- GCGAGGTGGGATTCAACTAT -3′ | 126 | |
| LuVe14_1R | 5′- GGAGTTACCAAATCCTCCCA -3′ | |||
| LuVe14_2F | 5′- CCAAATTTATGCGGTAACCC -3′ | 90 | ||
| LuVe14_2R | 5′- GATGCTTGGTTTCCCTGATT -3′ | |||
| LuVe14_3F | 5′- AATCAGGGAAACCAAGCATC -3′ | 110 | ||
| LuVe14_3R | 5′- CTTGTTTGCCTTCTTGCTGA -3′ |
| Protein Name | Gene Sequence ID | Protein Size (in Amino Acids) | Score |
|---|---|---|---|
| LuVe11 | Lus10003389.g | 1039 | 879.0 |
| LuVe12 | Lus10042239.g | 1090 | 821.2 |
| LuVe13 | Lus10026415.g | 1067 | 803.1 |
| LuVe14 | Lus10003387.g | 1061 | 774.2 |
| Protein a | Ve1 | LuVe11 | LuVe12 | LuVe13 | LuVe14 | GbVe | mVe1 | SlVe1 | StVe | AAT51733.1 | ACB99693.1 | AES86946.1 | BAD68095.1 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ve1 | 48.8 | 45.3 | 45.4 | 45.9 | 50.1 | 49.1 | 87.3 | 81.2 | 91.2 | 49.3 | 44.8 | 39.5 | |
| LuVe11 | 56.8 | 56.3 | 74.0 | 50.2 | 42.8 | 49.5 | 48.7 | 48.7 | 44.1 | 27.7 | 47.2 | ||
| LuVe12 | 78.9 | 53.5 | 45.9 | 40.6 | 44.6 | 44.0 | 44.8 | 44.6 | 43.3 | 38.7 | |||
| LuVe13 | 46.1 | 45.6 | 41.2 | 45.8 | 44.7 | 44.9 | 41.6 | 43.6 | 38.3 | ||||
| LuVe14 | 49.8 | 41.4 | 47.7 | 47.1 | 47.4 | 41.6 | 42.7 | 36.3 | |||||
| GbVe | 42.6 | 50.3 | 50.6 | 50.5 | 42.4 | 46.5 | 38.4 | ||||||
| mVe1 | 50.1 | 49.1 | 48.9 | 96.5 | 39.4 | 37.6 | |||||||
| SlVe1 | 81.4 | 84.7 | 50.1 | 44.5 | 39.7 | ||||||||
| StVe | 82.2 | 49.7 | 45.4 | 39.1 | |||||||||
| AAT51733.1 | 48.8 | 44.2 | 39.1 | ||||||||||
| ACB99693.1 | 39.6 | 37.8 | |||||||||||
| AES86946.1 | 36.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blum, A.; Castel, L.; Trinsoutrot-Gattin, I.; Driouich, A.; Laval, K. Identification of Tomato Ve1 Homologous Proteins in Flax and Assessment for Race-Specific Resistance in Two Fiber FlaxCultivars against Verticillium dahliae Race 1. Plants 2021, 10, 162. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10010162
Blum A, Castel L, Trinsoutrot-Gattin I, Driouich A, Laval K. Identification of Tomato Ve1 Homologous Proteins in Flax and Assessment for Race-Specific Resistance in Two Fiber FlaxCultivars against Verticillium dahliae Race 1. Plants. 2021; 10(1):162. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10010162
Chicago/Turabian StyleBlum, Adrien, Lisa Castel, Isabelle Trinsoutrot-Gattin, Azeddine Driouich, and Karine Laval. 2021. "Identification of Tomato Ve1 Homologous Proteins in Flax and Assessment for Race-Specific Resistance in Two Fiber FlaxCultivars against Verticillium dahliae Race 1" Plants 10, no. 1: 162. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10010162