Shoot and Root Traits Underlying Genotypic Variation in Early Vigor and Nutrient Accumulation in Spring Wheat Grown in High-Latitude Light Conditions

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
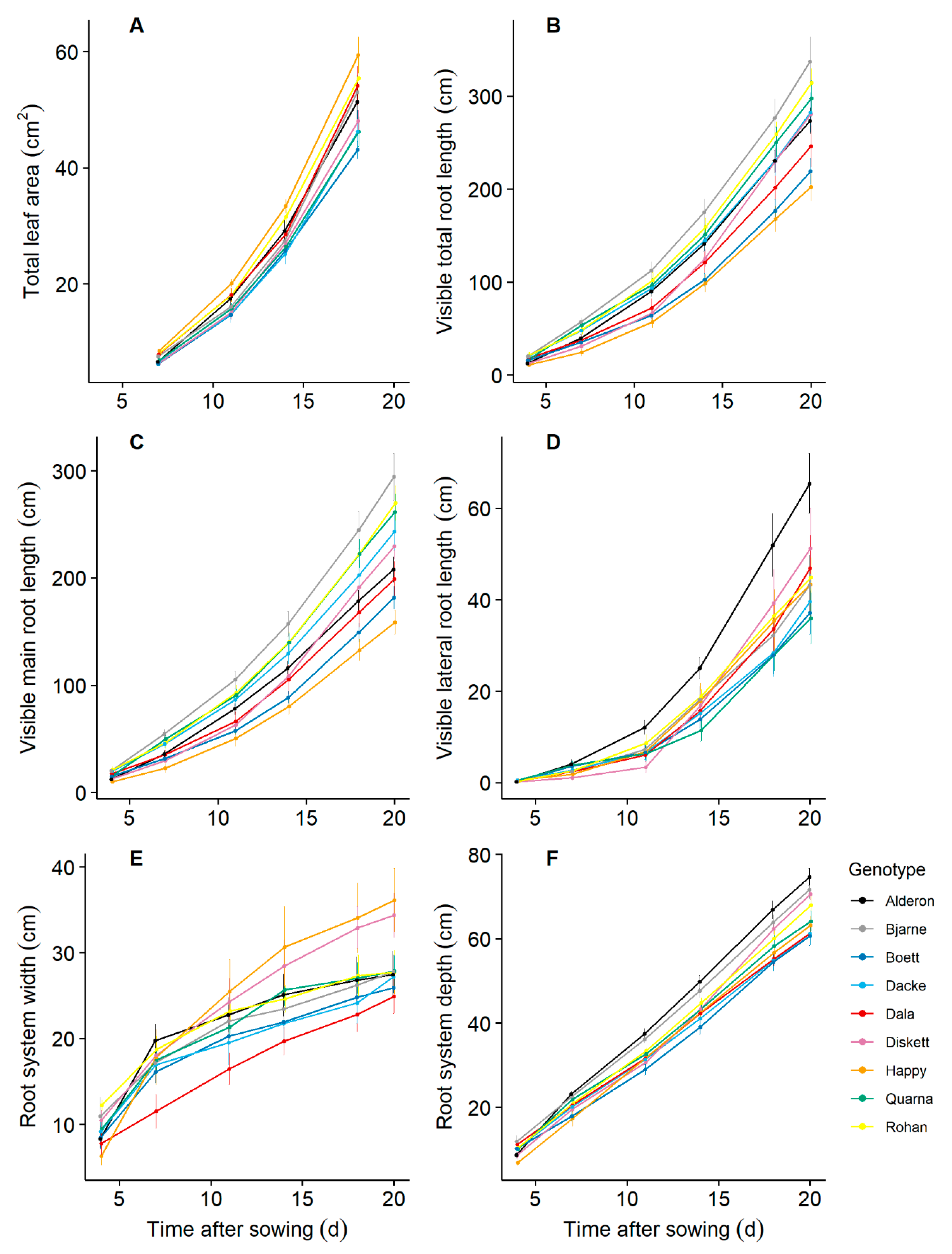
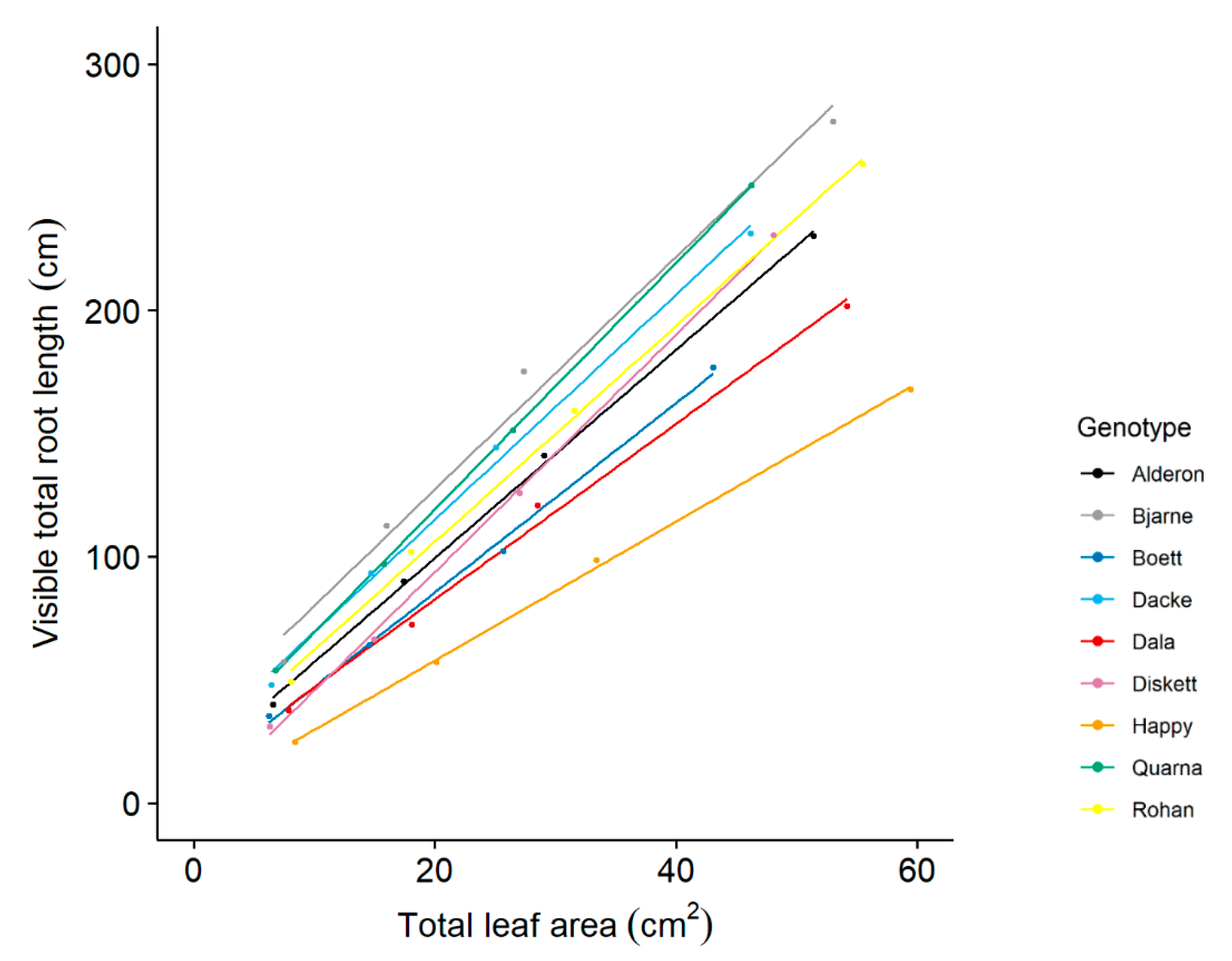
2.1. There Is Considerable Trait Variation across the Nine Genotypes
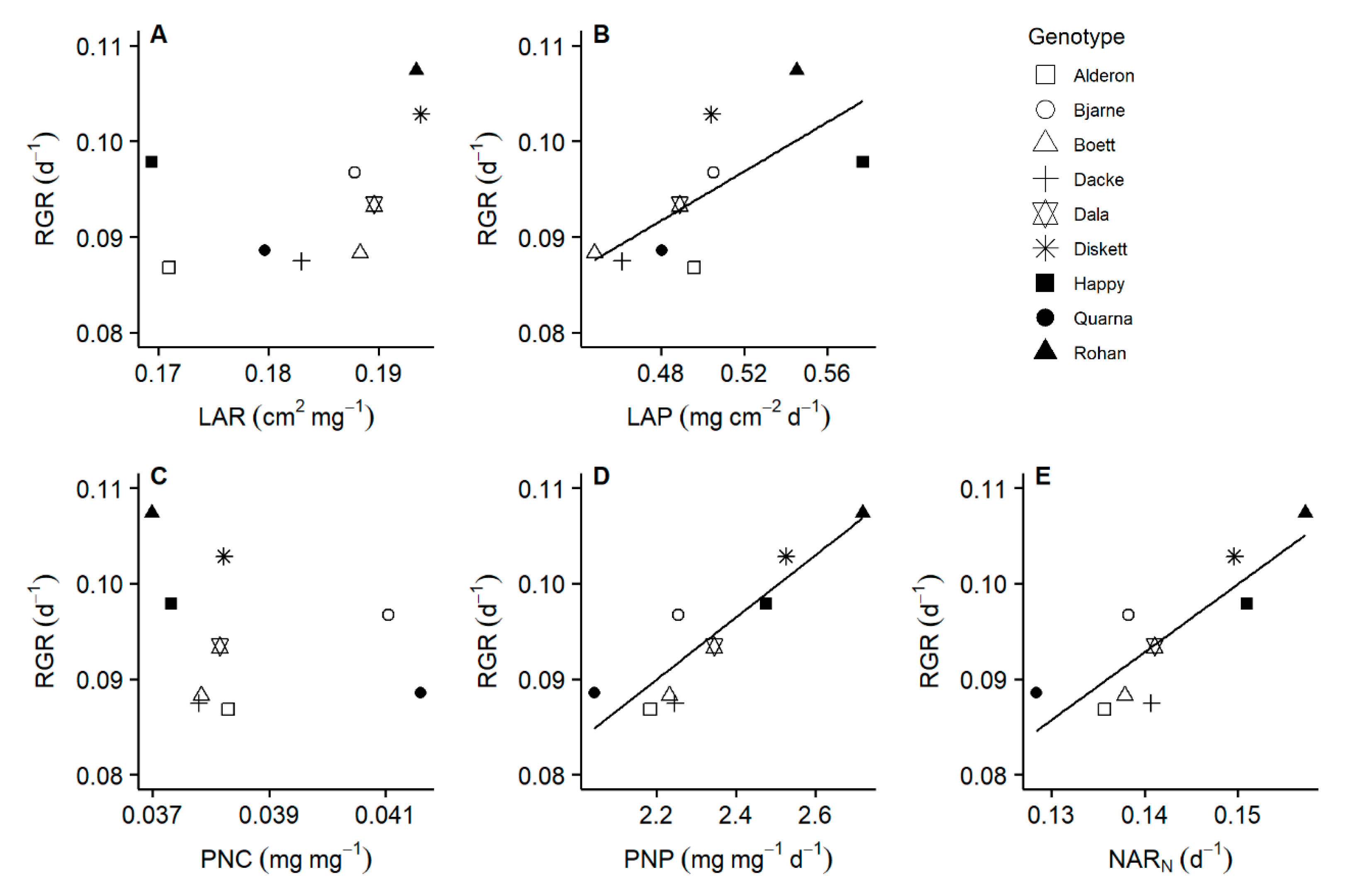
2.2. Leaf Area Productivity and Plant Nitrogen Productivity Drive the Variation in Relative Growth Rate of Whole Plant Biomass among Genotypes
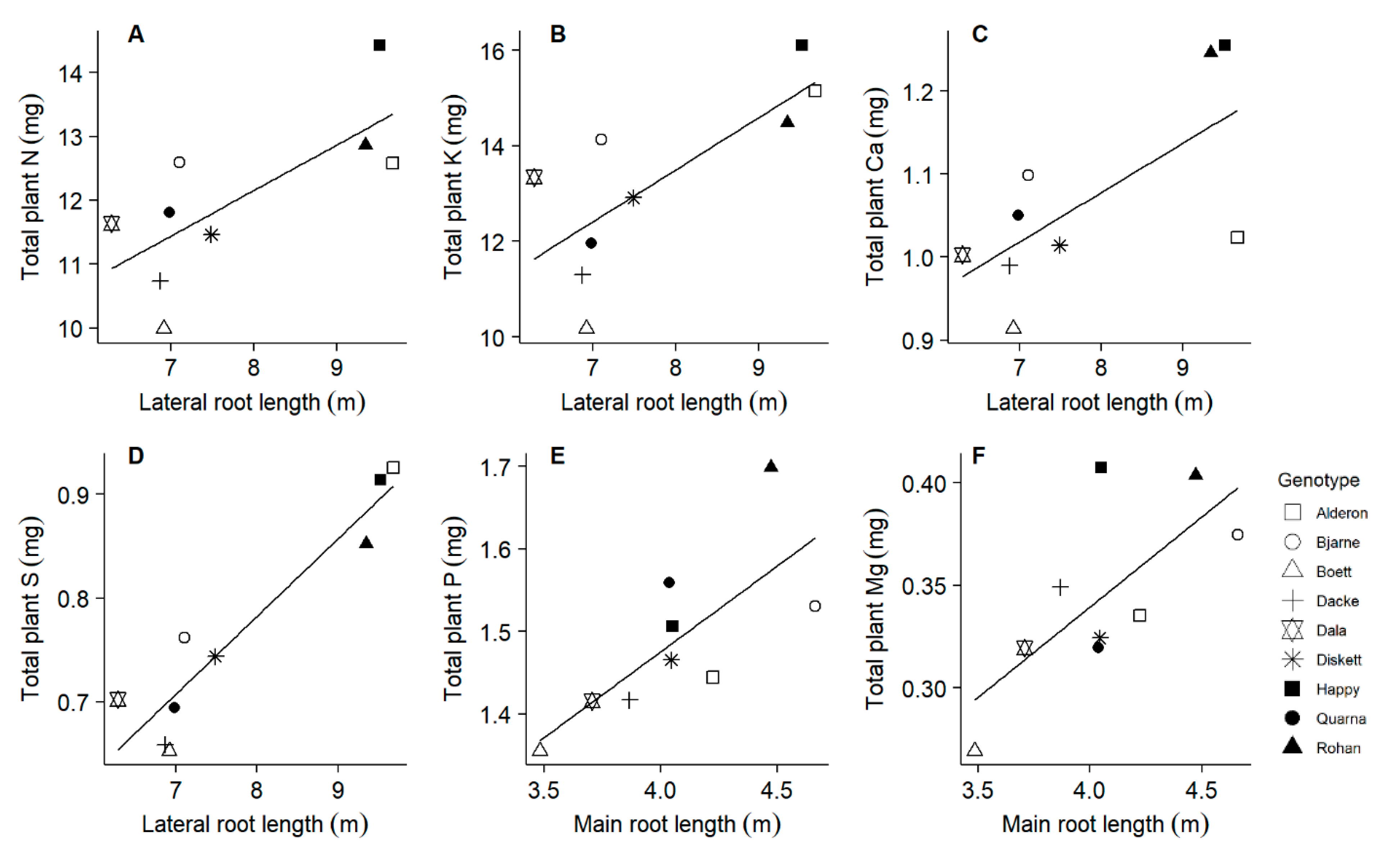
2.3. Main and Lateral Root Length Are Associated with Accumulation of Different Nutrients
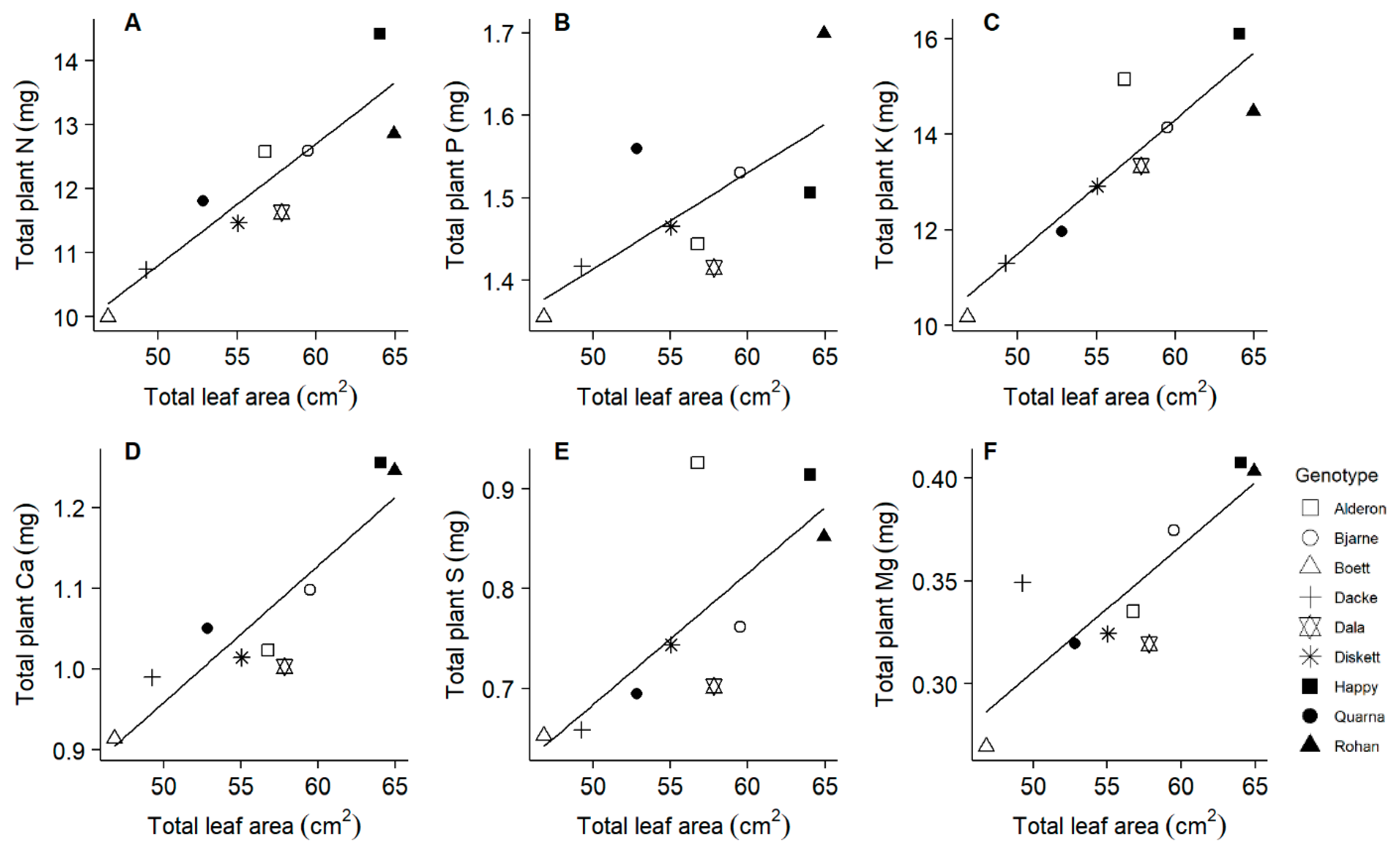
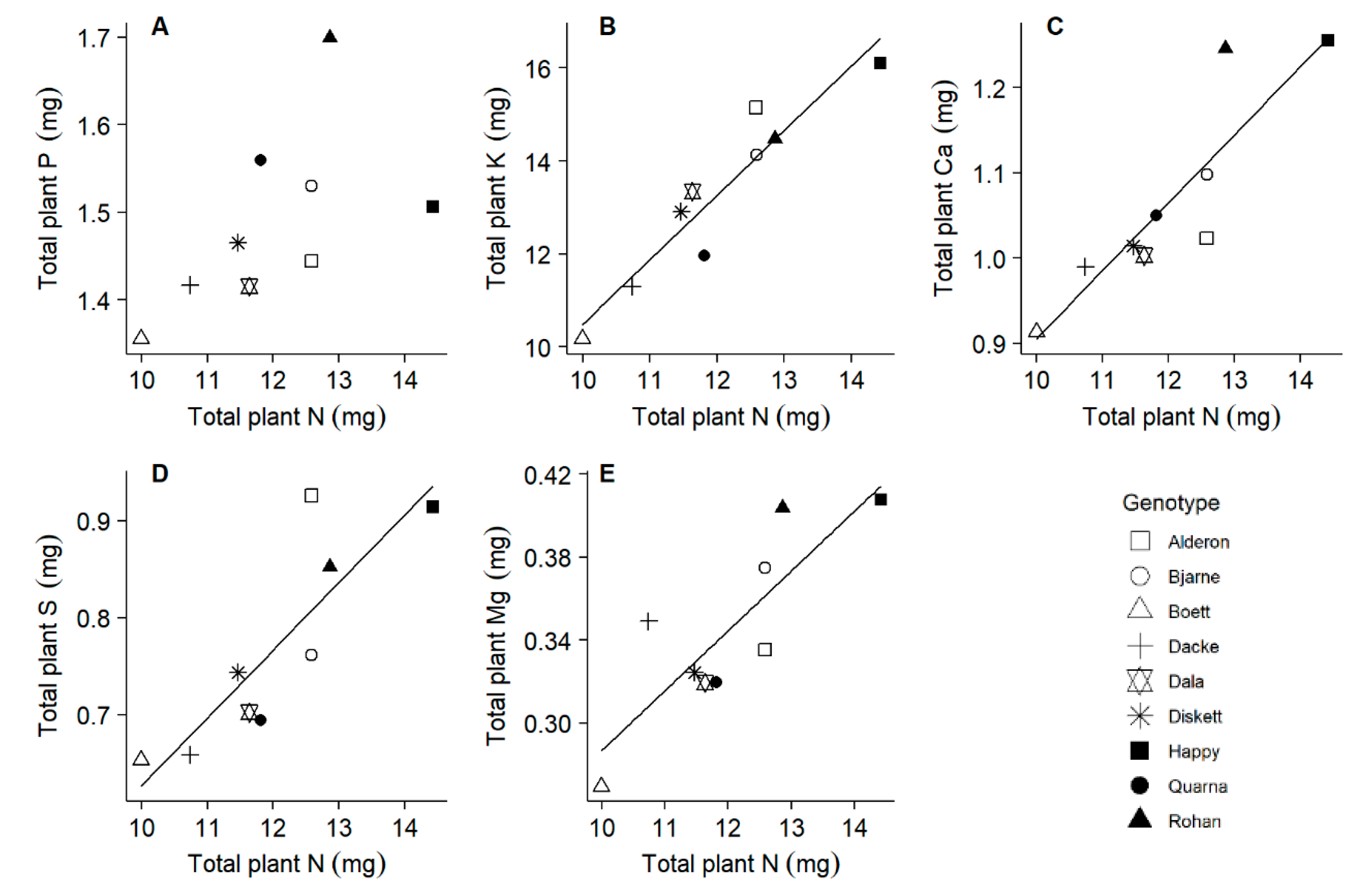
2.4. All Macronutrients Except Phosphorus Scale Similar to Nitrogen
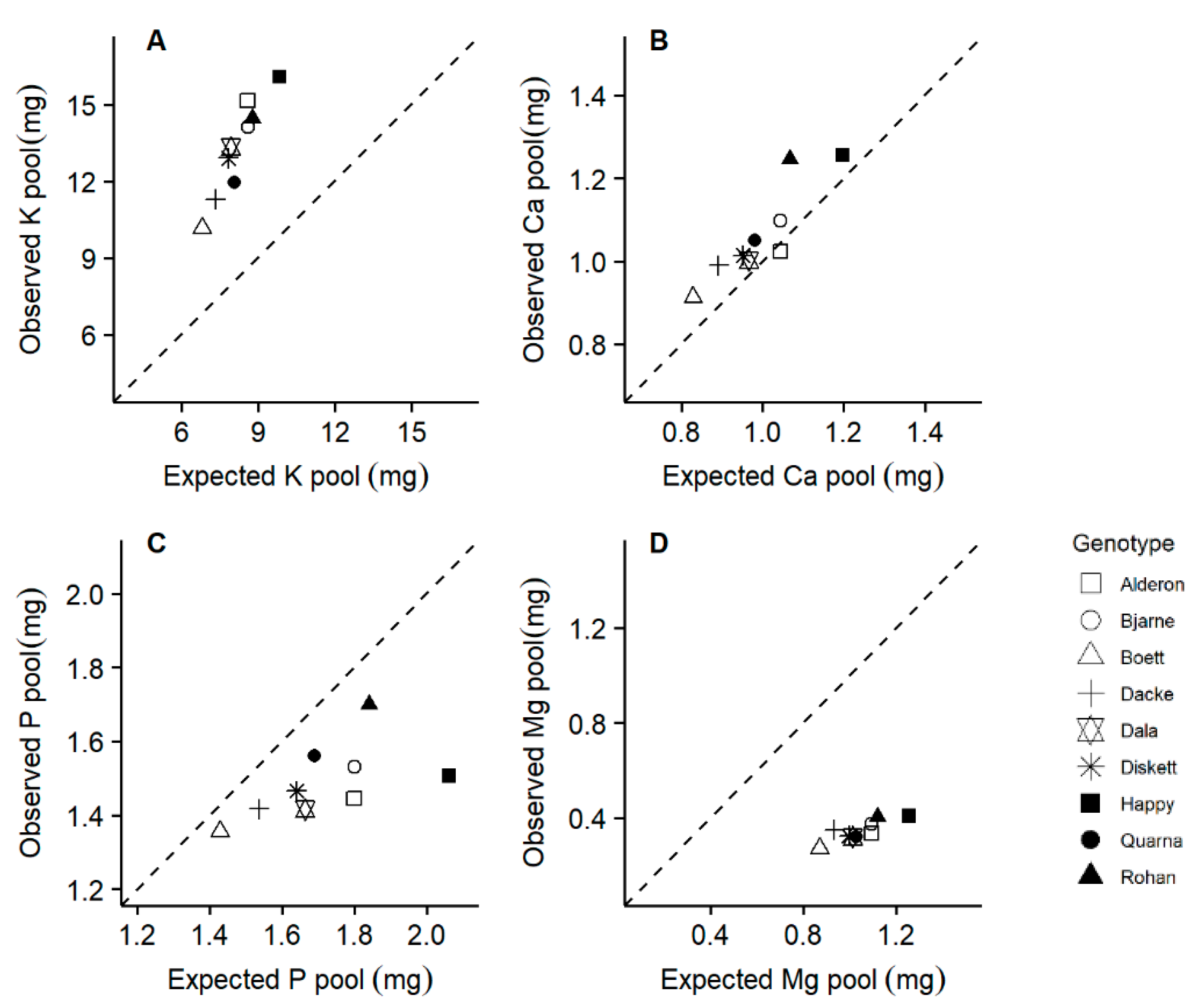
2.5. Suboptimal P-to-N and Mg-to-N Ratios Were Observed in Some Genotypes
3. Discussion
3.1. Genotypic Variation in Relative Growth Rate of Whole Plant Biomass and Its Determinants
3.2. Association of Shoot and Root Traits with the Accumulation of Individual Nutrient Elements
3.3. Nutrient Accumulation and Stoichiometry, and Optimum Nutrient Ratios
3.4. Implications for Breeding
4. Materials and Methods
4.1. Plant Material and Experimental Setup
4.2. Quantification of Phenotypic Traits
4.3. Nutrient Analyses
4.4. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| LAP | leaf area productivity |
| LAR | leaf area ratio |
| PNC | plant nitrogen concentration |
| PNP | plant nitrogen productivity |
| RGR | relative growth rate of whole plant biomass |
References
- López-Arredondo, D.L.; Leyva-González, M.A.; González-Morales, S.I.; López-Bucio, J.; Herrera-Estrella, L. Phosphate nutrition: Improving low-phosphate tolerance in crops. Annu. Rev. Plant Biol. 2014, 65, 95–123. [Google Scholar] [CrossRef] [PubMed]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, M.T.; Fillery, I.R.P.; Palta, J.A. Early vigorous growth is a major factor influencing nitrogen uptake in wheat. Funct. Plant Biol. 2004, 31, 121–129. [Google Scholar] [CrossRef]
- Liao, M.; Palta, J.A.; Fillery, I.R.P. Root characteristics of vigorous wheat improve early nitrogen uptake. Aust. J. Agric. Res. 2006, 57, 1097–1107. [Google Scholar] [CrossRef]
- An, D.; Su, J.; Liu, Q.; Zhu, Y.; Tong, Y.; Li, J.; Jing, R.; Li, B.; Li, Z. Mapping QTLs for nitrogen uptake in relation to the early growth of wheat (Triticum aestivum L.). Plant Soil 2006, 284, 73–84. [Google Scholar] [CrossRef]
- Asseng, S.; Turner, N.C.; Botwright, T.; Condon, A.G. Evaluating the impact of a trait for increased specific leaf area on wheat yields using a crop simulation model. Agron. J. 2003, 95, 10–19. [Google Scholar] [CrossRef]
- Li, J.; Wan, H.; Wei, H.; Wang, Q.; Zhou, Y.; Yang, W. QTL mapping for early vigor related traits in an elite wheat-breeding parent Chuanmai 42 derived from synthetic hexaploid wheat. Pakistan J. Agric. Sci. 2018, 55, 33–45. [Google Scholar]
- Palta, J.A.; Fillery, I.R.; Rebetzke, G.J. Restricted-tillering wheat does not lead to greater investment in roots and early nitrogen uptake. Field Crops Res. 2007, 104, 52–59. [Google Scholar] [CrossRef]
- Pang, J.; Palta, J.A.; Rebetzke, G.J.; Milroy, S.P. Wheat genotypes with high early vigour accumulate more nitrogen and have higher photosynthetic nitrogen use efficiency during early growth. Funct. Plant Biol. 2014, 41, 215–222. [Google Scholar] [CrossRef]
- Richards, R.; Lukacs, Z. Seedling vigour in wheat-sources of variation for genetic and agronomic improvement. Aust. J. Agric. Res. 2002, 53, 41–50. [Google Scholar] [CrossRef]
- Bishopp, A.; Lynch, J.P. The hidden half of crop yields. Nat. Plants 2015, 1, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Lynch, J.P. Reduced crown root number improves water acquisition under water deficit stress in maize (Zea mays L.). J. Exp. Bot. 2016, 67, 4545–4557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangarajan, H.; Postma, J.A.; Lynch, J.P. Co-optimization of axial root phenotypes for nitrogen and phosphorus acquisition in common bean. Ann. Bot. 2018, 122, 485–499. [Google Scholar] [CrossRef]
- Saengwilai, P.; Tian, X.; Lynch, J.P. Low crown root number enhances nitrogen acquisition from low-nitrogen soils in maize. Plant Physiol. 2014, 166, 581–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, A.; Lynch, J.P. Reduced frequency of lateral root branching improves N capture from low-N soils in maize. J. Exp. Bot. 2015, 66, 2055–2065. [Google Scholar] [CrossRef] [Green Version]
- Chimungu, J.G.; Brown, K.M.; Lynch, J.P. Large root cortical cell size improves drought tolerance in maize. Plant Physiol. 2014, 166, 2166–2178. [Google Scholar] [CrossRef] [Green Version]
- Chimungu, J.G.; Maliro, M.F.; Nalivata, P.C.; Kanyama-Phiri, G.; Brown, K.M.; Lynch, J.P. Utility of root cortical aerenchyma under water limited conditions in tropical maize (Zea mays L.). Field Crops Res. 2015, 171, 86–98. [Google Scholar] [CrossRef]
- Saengwilai, P.; Nord, E.A.; Chimungu, J.G.; Brown, K.M.; Lynch, J.P. Root cortical aerenchyma enhances nitrogen acquisition from low-nitrogen soils in maize. Plant Physiol. 2014, 166, 726–735. [Google Scholar] [CrossRef] [Green Version]
- Lynch, J.P.; Brown, K.M. Root strategies for phosphorus acquisition. In The Ecophysiology of Plant-Phosphorus Interactions; White, P., Hammond, J., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 83–116. [Google Scholar]
- Chawade, A.; Armoniene, R.; Berg, G.; Brazauskas, G.; Frostgard, G.; Geleta, M.; Gorash, A.; Henriksson, T.; Himanen, K.; Ingver, A.; et al. A transnational and holistic breeding approach is needed for sustainable wheat production in the Baltic Sea region. Physiol. Plant. 2018, 164, 442–451. [Google Scholar] [CrossRef] [Green Version]
- Fiorani, F.; Schurr, U. Future scenarios for plant phenotyping. Annu. Rev. Plant Biol. 2013, 64, 267–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagel, K.A.; Putz, A.; Gilmer, F.; Heinz, K.; Fischbach, A.; Pfeifer, J.; Faget, M.; Blossfeld, S.; Ernst, M.; Dimaki, C.; et al. GROWSCREEN-Rhizo is a novel phenotyping robot enabling simultaneous measurements of root and shoot growth for plants grown in soil-filled rhizotrons. Funct. Plant Biol. 2012, 39, 891–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, A.; Liebisch, F.; Hund, A. Plant phenotyping: From bean weighing to image analysis. Plant Methods 2015, 11, 14. [Google Scholar] [CrossRef] [Green Version]
- Watt, M.; Fiorani, F.; Usadel, B.; Rascher, U.; Muller, O.; Schurr, U. Phenotyping: New windows into the plant for breeders. Annu. Rev. Plant Biol. 2020, 71, 689–712. [Google Scholar] [CrossRef] [PubMed]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Growth and allocation. In Plant Physiological Ecology; Lambers, H., Chapin, F.S., Pons, T.L., Eds.; Springer: New York, NY, USA, 2008; pp. 321–374. [Google Scholar]
- Lambers, H.; Freijsen, A.; Poorter, H.; Hirose, T.; Van der Werf, A.; Cambridge, M.; Konings, H.; Pons, T. Analyses of growth based on net assimilation rate and nitrogen productivity: Their physiological background. In Causes and Consequences of Variation in Growth Rate and Productivity of Higher Plants; Lambers, H., Cambridge, M., Konings, H., Pons, T., Eds.; SPB Academic Publishing: Hague, The Netherlands, 1990; pp. 1–17. [Google Scholar]
- Weih, M. Evidence for increased sensitivity to nutrient and water stress in a fast-growing hybrid willow compared with a natural willow clone. Tree Physiol. 2001, 21, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Ågren, G.I. Theory for growth of plants derived from the nitrogen productivity concept. Physiol. Plant. 1985, 64, 17–28. [Google Scholar] [CrossRef]
- Poorter, H.; Remkes, C. Leaf area ratio and net assimilation rate of 24 wild species differing in relative growth rate. Oecologia 1990, 83, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Shipley, B. Net assimilation rate, specific leaf area and leaf mass ratio: Which is most closely correlated with relative growth rate? A meta-analysis. Funct. Ecol. 2006, 20, 565–574. [Google Scholar] [CrossRef]
- Shipley, B. Trade-offs between net assimilation rate and specific leaf area in determining relative growth rate: Relationship with daily irradiance. Funct. Ecol. 2002, 16, 682–689. [Google Scholar] [CrossRef]
- Adams, S.; Langton, F. Photoperiod and plant growth: A review. J. Hortic. Sci. Biotechnol. 2005, 80, 2–10. [Google Scholar] [CrossRef]
- Cormier, F.; Foulkes, J.; Hirel, B.; Gouache, D.; Moënne-Loccoz, Y.; Le Gouis, J. Breeding for increased nitrogen-use efficiency: A review for wheat (T. aestivum L.). Plant Breed. 2016, 135, 255–278. [Google Scholar] [CrossRef] [Green Version]
- López-Arredondo, D.L.; Sánchez-Calderón, L.; Yong-Villalobos, L. Molecular and genetic basis of plant macronutrient use efficiency: Concepts, opportunities, and challenges. In Plant Macronutrient Use Efficiency; Hossain, M., Kamiya, T., Burritt, D., Tran, L., Fujiwara, T., Eds.; Academic Press: Amsterdam, The Netherlands, 2017; pp. 1–29. [Google Scholar]
- Reich, M.; Aghajanzadeh, T.; De Kok, L.J. Physiological basis of plant nutrient use efficiency–concepts, opportunities and challenges for its improvement. In Nutrient Use Efficiency in Plants; Hawkesford, M., Kopriva, S., De, K., Eds.; Springer: Cham, Switzerland, 2014; pp. 1–27. [Google Scholar]
- Weih, M.; Hamnér, K.; Pourazari, F. Analyzing plant nutrient uptake and utilization efficiencies: Comparison between crops and approaches. Plant Soil 2018, 430, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Mengel, K.; Kirkby, E.A.; Kosegarten, H.; Appel, T. Plant nutrients. In Principles of Plant Nutrition; Mengel, K., Kirkby, E., Kosegarten, H., Appel, T., Eds.; Springer: Dordrecht, The Netherlands, 2001; pp. 1–13. [Google Scholar]
- Hamnér, K.; Weih, M.; Eriksson, J.; Kirchmann, H. Influence of nitrogen supply on macro-and micronutrient accumulation during growth of winter wheat. Field Crops Res. 2017, 213, 118–129. [Google Scholar] [CrossRef]
- Asplund, L.; Bergkvist, G.; Weih, M. Functional traits associated with nitrogen use efficiency in wheat. Acta Agric. Scand. Sect. B Soil Plant Sci. 2016, 66, 153–169. [Google Scholar] [CrossRef]
- Knecht, M.F.; Göransson, A. Terrestrial plants require nutrients in similar proportions. Tree Physiol. 2004, 24, 447–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ågren, G.I.; Weih, M. Multi-dimensional plant element stoichiometry—Looking beyond carbon, nitrogen, and phosphorus. Front. Plant Sci. 2020, 11, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gioia, T.; Nagel, K.A.; Beleggia, R.; Fragasso, M.; Ficco, D.B.M.; Pieruschka, R.; De Vita, P.; Fiorani, F.; Papa, R. Impact of domestication on the phenotypic architecture of durum wheat under contrasting nitrogen fertilization. J. Exp. Bot. 2015, 66, 5519–5530. [Google Scholar] [CrossRef] [Green Version]
- Heide, O.M. Effects of photoperiod and temperature on growth and flowering in Norwegian and British timothy cultivars (Phleum pratense L.). Acta Agric. Scand. 1982, 32, 241–252. [Google Scholar] [CrossRef]
- Golzarian, M.R.; Frick, R.A.; Rajendran, K.; Berger, B.; Roy, S.; Tester, M.; Lun, D.S. Accurate inference of shoot biomass from high-throughput images of cereal plants. Plant Methods 2011, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Møller, I.S.; White, P. Functions of macronutrients. In Marschner’s Mineral Nutrition of Higher Plants; Marschner, P., Ed.; Academic Press: Amsterdam, The Netherlands, 2012; pp. 135–189. [Google Scholar]
- Rychter, A.M.; Rao, I. Role of phosphorus in photosynthetic carbon metabolism. In Handbook of Photosynthesis; Pessarakli, M., Ed.; CRC Press: Boca Raton, FL, USA, 2005; pp. 123–148. [Google Scholar]
- Carstensen, A.; Szameitat, A.E.; Frydenvang, J.; Husted, S. Chlorophyll a fluorescence analysis can detect phosphorus deficiency under field conditions and is an effective tool to prevent grain yield reductions in spring barley (Hordeum vulgare L.). Plant Soil 2019, 434, 79–91. [Google Scholar] [CrossRef]
- Novák, V.; Vidovič, J. Transpiration and nutrient uptake dynamics in maize (Zea mays L.). Ecol. Model. 2003, 166, 99–107. [Google Scholar] [CrossRef]
- Reid, R.; Hayes, J. Mechanisms and control of nutrient uptake in plants. Int. Rev. Cytol. 2003, 229, 73–115. [Google Scholar] [PubMed]
- Lynch, J.P. Root phenes for enhanced soil exploration and phosphorus acquisition: Tools for future crops. Plant Physiol. 2011, 156, 1041–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombi, T.; Kirchgessner, N.; Walter, A.; Keller, T. Root Tip Shape Governs Root Elongation Rate under Increased Soil Strength. Plant Physiol. 2017, 174, 2289–2301. [Google Scholar] [CrossRef] [Green Version]
- Colombi, T.; Herrmann, A.M.; Vallenback, P.; Keller, T. Cortical cell diameter is key to energy costs of root growth in wheat. Plant Physiol. 2019, 180, 2049–2060. [Google Scholar] [CrossRef] [Green Version]
- Weih, M.; Pourazari, F.; Vico, G. Nutrient stoichiometry in winter wheat: Element concentration pattern reflects developmental stage and weather. Sci. Rep. 2016, 6, 35958. [Google Scholar] [CrossRef] [Green Version]
- Cakmak, I.; Yazici, A.M. Magnesium: A forgotten element in crop production. Better Crops 2010, 94, 23–25. [Google Scholar]
- Wang, Y.; Thorup-Kristensen, K.; Jensen, L.S.; Magid, J. Vigorous root growth is a better indicator of early nutrient uptake than root hair traits in spring wheat grown under low fertility. Front. Plant Sci. 2016, 7, 865. [Google Scholar] [CrossRef] [Green Version]
- Reich, M. The significance of nutrient interactions for crop yield and nutrient use efficiency. In Plant Macronutrient Use Efficiency; Elsevier: Amsterdam, The Netherlands, 2018; pp. 65–82. [Google Scholar]
- Rose, T.; Liu, L.; Wissuwa, M. Improving phosphorus efficiency in cereal crops: Is breeding for reduced grain phosphorus concentration part of the solution? Front. Plant Sci. 2013, 4, 444. [Google Scholar] [CrossRef] [Green Version]
- van de Wiel, C.C.; van der Linden, C.G.; Scholten, O.E. Improving phosphorus use efficiency in agriculture: Opportunities for breeding. Euphytica 2016, 207, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Gransee, A.; Führs, H. Magnesium mobility in soils as a challenge for soil and plant analysis, magnesium fertilization and root uptake under adverse growth conditions. Plant Soil 2013, 368, 5–21. [Google Scholar] [CrossRef] [Green Version]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circular. Calif. Agric. Exp. Station. 1950, 347, 32. [Google Scholar]
- Ren, Y.; He, X.; Liu, D.; Li, J.; Zhao, X.; Li, B.; Tong, Y.; Zhang, A.; Li, Z. Major quantitative trait loci for seminal root morphology of wheat seedlings. Mol. Breed. 2012, 30, 139–148. [Google Scholar] [CrossRef]
- Hoffmann, W.A.; Poorter, H. Avoiding bias in calculations of relative growth rate. Ann. Bot. 2002, 90, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Hunt, R. Plant Growth Curves. The Functional Approach to Plant Growth Analysis; Edward Arnold Ltd.: London, UK, 1982. [Google Scholar]
- Niklas, K.J. Plant allometry, leaf nitrogen and phosphorus stoichiometry, and interspecific trends in annual growth rates. Ann. Bot. 2006, 97, 155–163. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait (Unit) | p Value | Description |
|---|---|---|
| Non-destructive measurements | ||
| Leaf number (-) | <0.001 *** | Amount of leaves per plant |
| SPAD leaf1 (-) | <0.001 *** | Leaf chlorophyll content measured for the first leaf |
| SPAD leaf3 (-) | <0.001 *** | Leaf chlorophyll content measured for the third leaf |
| Visible total root length (cm) | <0.001 *** | Length of all visible roots in image |
| Visible main root length (cm) | <0.001 *** | Length of visible main roots in image |
| Visible lateral root length (cm) | 0.021 * | Length of visible lateral roots in image |
| Visible root system depth (cm) | <0.001 *** | Maximal vertical depth of a visible root system in image |
| Visible root system width (cm) | 0.009 ** | Maximal horizontal width of a visible root system in image |
| Measurements after destructive sampling | ||
| Shoot biomass (mg) | 0.008 ** | Dry weight of all shoots including leaves |
| Root biomass (mg) | 0.031 * | Dry weight of all roots |
| Whole plant biomass (mg) | 0.013 * | Sum of shoot and root biomass |
| Shoot fraction (%) | <0.001 *** | Shoot biomass per whole plant biomass |
| Total leaf area (cm2) | 0.006 ** | Total area of all leaves |
| Root fraction (%) | <0.001 *** | Root biomass per whole plant biomass |
| Total root length (m) | 0.006 ** | Total length of all roots |
| Total root surface area (m2) | 0.019 * | Total area of all roots |
| Main root length (m) | 0.179 | Length of roots with diameter > 0.2 mm |
| Lateral root length (m) | <0.001 *** | Length of roots with diameter < 0.2 mm |
| Specific root length (m mg−1) | 0.469 | Total root length per root biomass |
| Total root length: total leaf area (m cm−2) | 0.015 ** | Root length per leaf area |
| Seminal root number (-) | <0.001 *** | Amount of seminal roots per plant |
| Nodal root number (-) | <0.001 *** | Amount of nodal roots per plant |
| Cross-sectional root area (µm2) | <0.001 *** | Cross-sectional area of a nodal root |
| Cross-sectional stele area (µm2) | 0.001 ** | Cross-sectional stele area of a nodal root |
| Cross-sectional cortex area (µm2) | <0.001 *** | Cross-sectional cortex area of a nodal root |
| Cross-sectional aerenchyma area (µm2) | 0.642 | Cross-sectional aerenchyma area of a nodal root |
| Aerenchyma proportion (%) | 0.788 | Cross-sectional aerenchyma area per cortex area of a nodal root |
| Cortical cell diameter (µm) | 0.078 | Mean diameter of 15 cortical cells in a nodal root |
| N | P | K | S | Ca | Mg | Shoot Biomass | Total Leaf Area | Total Root Length | Main Root Length | Lateral Root Length | Aerenchyma Proportion | Cortical Cell Diameter | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | 0.102 | <0.001 *** | 0.003 ** | 0.001 *** | 0.005 ** | <0.001 *** | <0.001 *** | 0.013 * | 0.075 | 0.026 * | 0.929 | 0.592 | |
| P | 0.580 | 0.206 | 0.253 | 0.010 ** | 0.033 * | 0.109 | 0.034 * | 0.107 | 0.021 * | 0.219 | 0.578 | 0.721 | |
| K | 0.944 | 0.466 | <0.001 *** | 0.015 * | 0.016 * | <0.001 *** | 0.001 *** | 0.009 ** | 0.070 | 0.018 * | 0.906 | 0.641 | |
| S | 0.857 | 0.426 | 0.920 | 0.036 * | 0.054 | 0.004 ** | 0.016 * | <0.001 *** | 0.126 | <0.001 *** | 0.658 | 0.188 | |
| Ca | 0.889 | 0.799 | 0.770 | 0.701 | <0.001 *** | <0.001 *** | <0.001 *** | 0.023 * | 0.070 | 0.045 * | 0.611 | 0.679 | |
| Mg | 0.839 | 0.707 | 0.768 | 0.658 | 0.926 | 0.005 ** | 0.004 ** | 0.035 * | 0.029 * | 0.080 | 0.804 | 0.896 | |
| Shoot biomass | 0.987 | 0.569 | 0.944 | 0.845 | 0.905 | 0.841 | <0.001 *** | 0.018 * | 0.116 | 0.029 | 0.890 | 0.666 | |
| Total leaf area | 0.900 | 0.706 | 0.902 | 0.766 | 0.901 | 0.847 | 0.935 | 0.032 * | 0.050 * | 0.067 | 0.916 | 0.844 | |
| Total root length | 0.781 | 0.573 | 0.807 | 0.949 | 0.739 | 0.702 | 0.759 | 0.709 | 0.067 | <0.001 *** | 0.413 | 0.127 | |
| Main root length | 0.619 | 0.747 | 0.628 | 0.549 | 0.628 | 0.718 | 0.562 | 0.667 | 0.634 | 0.218 | 0.620 | 0.794 | |
| Lateral root length | 0.730 | 0.455 | 0.757 | 0.942 | 0.678 | 0.611 | 0.720 | 0.633 | 0.977 | 0.456 | 0.270 | 0.086 | |
| Aerenchyma proportion | 0.035 | 0.215 | −0.046 | 0.172 | 0.197 | 0.097 | 0.054 | −0.041 | 0.312 | −0.192 | 0.412 | 0.729 | |
| Cortical cell diameter | 0.207 | 0.139 | 0.181 | 0.482 | 0.161 | −0.051 | 0.168 | 0.077 | 0.547 | 0.102 | 0.602 | 0.135 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Fiorani, F.; Jäck, O.; Colombi, T.; Nagel, K.A.; Weih, M. Shoot and Root Traits Underlying Genotypic Variation in Early Vigor and Nutrient Accumulation in Spring Wheat Grown in High-Latitude Light Conditions. Plants 2021, 10, 174. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10010174
Liu H, Fiorani F, Jäck O, Colombi T, Nagel KA, Weih M. Shoot and Root Traits Underlying Genotypic Variation in Early Vigor and Nutrient Accumulation in Spring Wheat Grown in High-Latitude Light Conditions. Plants. 2021; 10(1):174. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10010174
Chicago/Turabian StyleLiu, Hui, Fabio Fiorani, Ortrud Jäck, Tino Colombi, Kerstin A. Nagel, and Martin Weih. 2021. "Shoot and Root Traits Underlying Genotypic Variation in Early Vigor and Nutrient Accumulation in Spring Wheat Grown in High-Latitude Light Conditions" Plants 10, no. 1: 174. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10010174