Ethylene and Sulfur Coordinately Modulate the Antioxidant System and ABA Accumulation in Mustard Plants under Salt Stress


 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Effect of Ethephon and Sulfur on Ions Accumulation
2.2. Ethephon and Sulfur Coordinately Optimize Antioxidant System and Redox State and Reduce Oxidative Stress
2.3. Ethephon and Sulfur Regulate ATP-S Activity and S Content
2.4. Effects of Ethephon and Sulfur on ACS Activity and Ethylene Emission
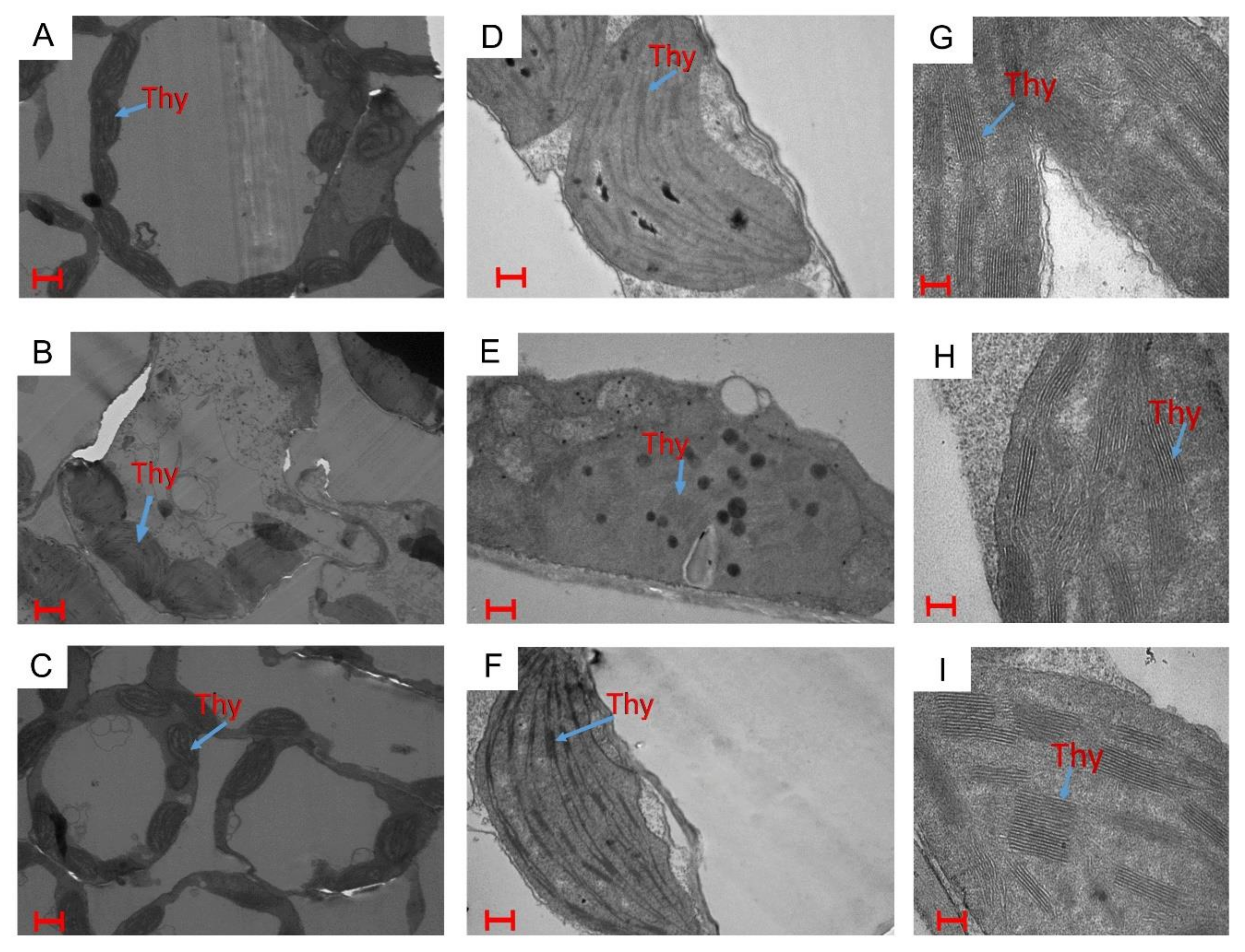
2.5. Effects of Ethylene and Sulfur on Stomatal Response and Chloroplast Ultrastructure
2.6. Supplementation of Ethephon and Sulfur Coordinately Increases Photosynthetic Efficiency, Pigment Content and Growth Parameters
2.7. Effects of Norbornadiene on ABA/H2O2 Content, Stomatal Regulation, and Gas Exchange
3. Discussion
4. Materials and Methods
4.1. Plant Material, Growth Conditions, and Treatments
4.2. Biochemical Analyses
4.2.1. Measurement of Na+ and Cl− Content
4.2.2. Measurement of H2O2 Content and Lipid Peroxidation
4.2.3. Activity of ATP-Sulfurylase and Sulfur Content
4.3. Antioxidant Metabolism
Assay of Antioxidant Enzymes Activities
4.4. Glutathione and Ascorbate Pools
4.5. ACS Activity and Ethylene Evolution
4.6. Abscisic Acid Determination
4.7. Cytological and Histological Analysis
4.7.1. Scanning Electron Microscopy
4.7.2. Transmission Electron Microscopy
4.7.3. Confocal Microscopy
4.8. Pigment Analysis and Leaf Gas Exchange
4.9. Water Use Efficiency and Stomatal Aperture
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hossain, S. Present scenario of global salt affected soils, its management and importance of salinity. Int. Res. J. Biol. Sci. 2019, 1, 1–3. [Google Scholar]
- FAO, Food and Agriculture Organization of the United Nations. How to Feed the World in 2050; Executive Summary—Proceedings of the Expert Meeting on How to Feed the World I 2050; FAO, Food and Agriculture Organization of the United Nations: Rome, Italy, 2009. [Google Scholar]
- Duar, I. Effects of hydro and hormonal priming on quinoa (Chenopodium quinoa willd.) seed germination under salt and drought stress. Pak. J. Bot. 2018, 50, 1669–1673. [Google Scholar]
- Fatma, M.; Masood, A.; Per, T.S.; Khan, N.A. Nitric oxide alleviates salt stress inhibited photosynthetic performance by interacting with sulfur assimilation in mustard. Front. Plant Sci. 2016, 7, 521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehar, Z.; Masood, A.; Khan, N.A. Nitric oxide reverses glucose-mediated photosynthetic repression in wheat (Triticum aestivum L.) under salt stress. Environ. Exp. Bot. 2019, 161, 277–289. [Google Scholar] [CrossRef]
- Hussain, T.; Koyro, H.W.; Zhang, W.; Liu, X.; Gul, B.; Liu, X. Low salinity improves photosynthetic performance in Panicum antidotale under drought stress. Front. Plant Sci. 2020, 11, 481. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Wahid, A.; Siddique, K.H.M. Drought stress in plants: An overview, plant responses to drought stress. In Plant Responses to Drought Stress; Farooq, M., Hussain, M., Wahid, A., Siddique, K.H.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 1–33. [Google Scholar]
- Upchurch, R.G. Fatty acid unsaturation, mobilization, and regulation in the response of plants to stress. Biotechnol. Lett. 2008, 30, 967–977. [Google Scholar] [CrossRef]
- Suo, J.; Zhao, Q.; David, L.; Chen, S.; Dai, S. Salinity response in chloroplasts: Insights from gene characterization. Int. J. Mol. Sci. 2017, 18, 1011. [Google Scholar] [CrossRef]
- Akyol, T.Y.; Yilmaz, O.; UzİLday, B.; UzİLday, R.Ö.; Türkan, İ. Plant response to salinity: An analysis of ROS formation, signaling, and antioxidant defense. Turk. J. Bot. 2020, 44, 1–13. [Google Scholar]
- Riffat, A.; Sajid, M.; Ahmad, A. Alleviation of adverse effects of salt stress on growth of maize (Zea mays) by sulfur supplementation. Pak. J. Bot. 2020, 52, 763–773. [Google Scholar] [CrossRef]
- Riyazuddin, R.; Verma, R.; Singh, K.; Nisha, N.; Keisham, M.; Bhati, K.K.; Kim, S.T.; Gupta, R. Ethylene: A Master regulator of salinity stress tolerance in plants. Biomolecules 2020, 10, 959. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Gulf Professional Publishing, Academic Press: London, UK, 1995; p. 889. [Google Scholar]
- Fatma, M.; Khan, M.I.R.; Masood, A.; Khan, N.A. Coordinate changes in assimilatory Sulfate reduction are correlated to salt tolerance: Involvement of phytohormones. Annu. Rev. Res. Biol. 2013, 267–295. [Google Scholar]
- Khan, N.; Khan, M.I.R.; Asgher, M.; Fatma, M.; Masood, A.; Syeed, S. Salinity tolerance in plants: Revisiting the role of sulfur metabolites. J. Plant Biochem. Physiol. 2014, 2, 2. [Google Scholar]
- Lou, L.; Kang, J.; Pang, H.; Li, Q.; Du, X.; Wu, W.; Chen, J.; Lv, J. Sulfur Protects Pakchoi (Brassica chinensis L.) seedlings against cadmium stress by regulating ascorbate-glutathione metabolism. Int. J. Mol. Sci. 2017, 18, 1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Zelm, E.; Zhang, Y.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [Green Version]
- Fatma, M.; Asgher, M.; Masood, A.; Khan, N.A. Excess sulfur supplementation improves photosynthesis and growth in mustard under salt stress through increased production of glutathione. Environ. Exp. Bot. 2014, 107, 55–63. [Google Scholar] [CrossRef]
- Khan, N.A.; Khan, M.I.R.; Ferrante, A.; Poor, P. Editorial: Ethylene: A key regulatory molecule in plants. Front. Plant. Sci. 2017, 8, 1782. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Belfield, E.J.; Cao, Y.; Smith, J.A.C.; Harberd, N.P. An Arabidopsis soil-salinity tolerance mutation confers ethylene-mediated enhancement of sodium potassium homeostasis. Plant Cell 2013, 25, 3535–3552. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, N.; Khan, N.A.; Ferrante, A.; Trivellini, A.; Francini, A.; Khan, M.I.R. Ethylene role in plant growth, development and senescence: Interaction with other phytohormones. Front. Plant. Sci. 2017, 8, 475. [Google Scholar] [CrossRef] [Green Version]
- Minorsky, P. Ethylene and ABA regulate ascorbic acid and reactive oxygen species. Plant Physiol. Inside 2019, 1. [Google Scholar]
- Nazareno, A.L.; Hernandez, B.S. A mathematical model of the interaction of abscisic acid, ethylene and methyl jasmonate on stomatal closure in plants. PLoS ONE 2017, 12, e0171065. [Google Scholar] [CrossRef]
- Cao, Y.; Wu, Y.; Zheng, Z.; Song, F. Overexpression of the rice EREBP-like gene OsBIERF3 enhances disease resistance and salt tolerance in transgenic tobacco. Physiol. Mol. Plant Pathol. 2005, 67, 202–211. [Google Scholar] [CrossRef]
- Wawrzynska, A.; Moniuszko, G.; Sirko, A. Links between ethephon and Sulfur nutrition—A regulatory interplay or just metabolite association? Front. Plant Sci. 2015, 6, 1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masood, A.; Iqbal, N.; Khan, N.A. Role of ethylene in alleviation of cadmium-induced photosynthetic capacity inhibition by sulphur in mustard. Plant Cell Environ. 2012, 35, 524–533. [Google Scholar] [CrossRef] [PubMed]
- Valerio, M.; Lovelli, S.; Sofo, A.; Perniola, M.; Scopa, A.; Amato, M. Root and leaf ABA concentration impact on gas exchange in tomato (Lycopersicon esculentum Mill.) plants subjected to partial root-zone drying. Ital. J. Agron. 2017, 12, 788. [Google Scholar] [CrossRef] [Green Version]
- Lovelli, S.; Scopa, A.; Perniola, M.; Di Tommaso, T.; Sofo, A. Abscisic acid root and leaf concentration in relation to biomass partitioning in salinized tomato plants. J. Plant Physiol. 2012, 169, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Khorshidi, M.B.; Yarnia, M.; Hassanpanah, D. Salinity effect on nutrients accumulation in alfalfa shoots in hydroponic condition. J. Food Agric. Environ. 2009, 7, 787–790. [Google Scholar]
- Khan, N.; Syeed, S.; Masood, A.; Nazar, R.; Iqbal, N. Application of salicylic acid increases contents of nutrients and antioxidative metabolism in mungbean and alleviates adverse effects of salinity stress. Int. J. Plant Biol. 2010, 1, e1. [Google Scholar] [CrossRef]
- Nazar, R.; Iqbal, N.; Masood, A.; Syeed, S.; Khan, N.A. Understanding the significance of sulfur in improving salinity tolerance in plants. Environ. Exp. Bot. 2011, 70, 80–87. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Sotiras, M.; Papadakis, I.; Landi, M.; Tsaniklidis, G.; Tsiantas, P.; Psychoyou, M. Allocation pattern, photosynthetic performance and sugar metabolism in hydroponically grown seedlings of loquat (Eriobotrya japonica Lindl.) subjected to salinity. Photosynthetica 2019, 57, 258–267. [Google Scholar] [CrossRef]
- Pompeiano, A.; Landi, M.; Meloni, G.; Vita, F.; Guglielminetti, L.; Guidi, L. Allocation pattern, ion partitioning, and chlorophyll a fluorescence in Arundo donax L. in responses to salinity stress. Plant Biosyst. 2017, 151, 613–622. [Google Scholar] [CrossRef]
- Zhu, T.; Deng, X.; Zhou, X.; Zhu, L.; Zou, L.; Li, P.; Zhang, D.; Lin, H. Ethylene and hydrogen peroxide are involved in brassinosteroid-induced salt tolerance in tomato. Sci. Rep. 2016, 6, 35392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2009, 8, 384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foyer, C.H.; Noctor, G. Oxidant and antioxidant signalling in plants: A re-evaluastion of the concept of oxidative stress in a physiological context. Plant Cell Environ. 2005, 28, 1056–1071. [Google Scholar] [CrossRef]
- Singh, R.; Parihar, P.; Prasad, S.M. Sulphur and calcium attenuate arsenic toxicity in Brassica by adjusting ascorbate—Glutathione cycle and sulfur metabolism. Plant Growth Regul. 2020, 91, 221–235. [Google Scholar] [CrossRef]
- Asgher, M.; Khan, N.A.; Khan, M.I.R.; Fatma, M.; Masood, A. Ethylene production is associated with alleviation of cadmium-induced oxidative stress by sulfur in mustard types differing in ethylene sensitivity. Ecotoxicol. Environ. Saf. 2014, 106, 54–61. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. The role of endogenous nitric oxide in salicylic acid-induced up-regulation of ascorbate-glutathione cycle involved in salinity tolerance of pepper (Capsicum annuum L.) plants. Plant Physiol. Biochem. 2020, 147, 10–20. [Google Scholar] [CrossRef]
- Sauter, M.; Moffatt, B.; Saechao, M.C.; Hell, R.; Wirtz, M. Methionine salvage and S-adenosylmethionine: Essential links between sulfur, ethylene and polyamine biosynthesis. Biochem. J. 2013, 451, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Nocito, F.F.; Lancilli, C.; Crema, B.; Fourcroy, P.; Davidian, J.C.; Sacchi, G.A. Heavy metal stress and sulfate uptake in maize roots. Plant Physiol. 2006, 141, 1138–1148. [Google Scholar] [CrossRef] [Green Version]
- Abdalla, M.A.; Mühling, K.H. Plant-derived sulfur containing natural products produced as a response to biotic and abiotic stresses: A review of their structural diversity and medicinal importance. J. Appl. Bot. Food Qual. 2019, 92, 204–215. [Google Scholar]
- Iqbal, N.; Trivellini, A.; Masood, A.; Ferrante, A.; Khan, N.A. Current understanding on ethylene signaling in plants: The influence of nutrient availability. Plant Physiol. Biochem. 2013, 73, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Asgher, M.; Per, T.S.; Verma, S.; Pandith, S.A.; Masood, A.; Khan, N.A. Ethylene supplementation increases PSII efficiency and alleviates chromium-inhibited photosynthesis through increased nitrogen and sulfur assimilation in mustard. J. Plant Growth Regul. 2018, 37, 1300–1317. [Google Scholar] [CrossRef]
- Maruyama-Nakashita, A.; Nakamura, Y.; Tohge, T.; Saito, K.; Takahashi, H. Arabidopsis SLIM1 is a central transcriptional regulator of plant sulfur response and metabolism. Plant Cell 2006, 18, 3235–3251. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.I.R.; Johan, B.; Alajmi, M.F.; Rehman, M.T.; Khan, N.A. Exogenously-sourced ethylene modulates defense mechanisms and promotes tolerance to zinc stress in mustard (Brassica juncea L.). Plants 2019, 8, 540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.A.; Asgher, M.; Per, T.S.; Masood, A.; Fatma, M.; Khan, M.I.R. Ethylene potentiates sulfur-mediated reversal of cadmium inhibited photosynthetic responses in mustard. Front. Plant Sci. 2016, 7, 1628. [Google Scholar] [CrossRef] [Green Version]
- Alves, L.R.; Rodrigues dos Reis, A.; Prado, E.R.; Lavres, J.; Pompeu, G.B.; Azevedo, R.A.; Gratão, P.L. New insights into cadmium stressful-conditions: Role of ethylene on selenium-mediated antioxidant enzymes. Ecotoxicol. Environ. Saf. 2019, 186, 109747. [Google Scholar] [CrossRef]
- Lin, Y.; Wang, J.; Zu, Y.; Tang, Z. Ethylene antagonizes the inhibition of germination in arabidopsis induced by salinity by modulating the concentration of hydrogen peroxide. Acta Physiol. Plant. 2012, 34, 1895–1904. [Google Scholar] [CrossRef]
- Huang, J.Q.; Jiang, H.F.; Zhou, Y.Q.; Lei, Y.; Wang, S.Y.; Liao, B.S. Ethylene inhibited aflatoxin biosynthesis is due to oxidative stress alleviation and related to glutathione redox state changes in Aspergillus flavus. Int. J. Food Microbiol. 2009, 130, 17–21. [Google Scholar] [CrossRef]
- Pallas, J.E.; Kays, S.J. Inhibition of photosynthesis by ethylene—A stomatal effect. Plant Physiol. 1982, 70, 598–601. [Google Scholar] [CrossRef] [Green Version]
- Desikan, R.; Last, K.; Harrett-Williams, R.; Tagliavia, C.; Harter, K.; Hooley, R.; Hancock, J.T.; Neill, S.J. Ethylene-induced stomatal closure in arabidopsis occurs via AtrbohF-mediated hydrogen peroxide synthesis. Plant J. Cell Mol. Biol. 2006, 47, 907–916. [Google Scholar] [CrossRef]
- Iqbal, N.; Nazar, R.; Syeed, S.; Masood, A.; Khan, N.A. Exogenously-sourced ethylene increases stomatal conductance, photosynthesis, and growth under optimal and deficient nitrogen fertilization in mustard. J. Exp. Bot. 2011, 62, 4955–4963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, S.; Guo, S.R.; Sun, J.; Yuan, L.Y. Effects of salt stress on the structure and function of the photosynthetic apparatus in Cucumis sativus and its protection by exogenous putrescine. Physiol. Plant. 2012, 146, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Kirchhoff, H.; Sharpe, R.M.; Herbstova, M.; Yarbrough, R.; Edwards, G.E. Differential mobility of pigment-protein complexes in granal and agranal thylakoid membranes of C3 and C4 plants. Plant Physiol. 2013, 161, 497–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.I.R.; Nazir, F.; Asgher, M.; Per, T.S.; Khan, N.A. Selenium and sulfur influence ethylene formation and alleviate cadmium-induced oxidative stress by improving proline and glutathione production in wheat. J. Plant Physiol. 2015, 173, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Amjad, M.; Akhtar, J.; Anwar-ul-Haq, M.; Yang, A.; Akhtar, S.S.; Jacobsen, S.E. Integrating role of ethylene and ABA in tomato plants adaptation to salt stress. Sci. Horticul. 2014, 172, 109–116. [Google Scholar] [CrossRef]
- Iqbal, N.; Masood, A.; Khan, M.I.R.; Asgher, M.; Fatma, M.; Khan, N.A. Cross-talk between sulfur assimilation and ethylene signaling in plants. Plant Signal. Behav. 2013, 8, e22478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iqbal, N.; Umar, S.; Khan, N.A. Nitrogen availability regulates proline and ethylene production and alleviates salinity stress in mustard (Brassica juncea). J. Plant Physiol. 2015, 178, 84–91. [Google Scholar] [CrossRef]
- Romera, F.J.; Alcántara, E. Ethylene involvement in the regulation of Fe-deficiency stress responses by strategy I plants. Funct. Plant Biol. 2004, 31, 315–328. [Google Scholar] [CrossRef]
- Ibrahim, I.S.; Abdel-Razik, A.B.; Ebeed Naglaa, M. Effect of some plant growth regulators on biochemical gene expression, growth and yield of faba bean. Pak. J. Biotechnol. 2007, 4, 47–64. [Google Scholar]
- Fujita, Y.; Fujita, M.; Satoh, R.; Maruyama, K.; Parvez, M.M.; Seki, M.; Hiratsu, K.; Ohme-Takagi, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AREB1 Is a transcription activator of novel ABRE-dependent ABA signaling that enhances drought stress tolerance in arabidopsis. Plant Cell 2005, 17, 3470–3488. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Nose, T.; Jikumaru, Y.; Kamiya, Y. ABA inhibits entry into stomatal-lineage development in arabidopsis leaves. Plant J. 2013, 74, 448–457. [Google Scholar] [CrossRef] [PubMed]
- She, X.; Song, X. Ethylene inhibits abscisic acid-induced stomatal closure in Vicia faba via reducing nitric oxide levels in guard cells. N. Z. J. Bot. 2012, 50, 203–216. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Sano, T.; Tamaoki, M.; Nakajima, N.; Kondo, N.; Hasezawa, S. Ethylene inhibits abscisic acid-induced stomatal closure in arabidopsis. Plant Physiol. 2005, 138, 2337–2343. [Google Scholar] [CrossRef] [Green Version]
- Daszkowska-Golec, A.; Szarejko, I. Open or close the gate—Stomata action under the control of phytohormones in drought stress conditions. Front. Plant Sci. 2013, 4, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak, J.M.; Mori, I.C.; Pei, Z.M.; Leonhardt, N.; Torres, M.A.; Dangl, J.L.; Bloom, R.E.; Bodde, S.; Jones, J.D.G.; Schroeder, J.I. NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in arabidopsis. EMBO J. 2013, 22, 2623–2633. [Google Scholar] [CrossRef]
- Gudesblat, G.E.; Iusem, N.D.; Morris, P.C. Guard cell-specific inhibition of arabidopsis MPK3 expression causes abnormal stomatal responses to abscisic acid and hydrogen peroxide. New Phytol. 2007, 173, 713–721. [Google Scholar] [CrossRef]
- Beaudoin, N.; Serizet, C.; Gosti, F.; Giraudat, J. Interactions between abscisic acid and ethylene signaling cascades. Plant Cell 2000, 12, 1103–1116. [Google Scholar]
- Ghassemian, M.; Nambara, E.; Cutler, S.; Kawaide, H.; Kamiya, Y.; McCourt, P. Regulation of abscisic acid signaling by the ethylene response pathway in arabidopsis. Plant Cell 2000, 12, 1117–1126. [Google Scholar]
- Yanagisawa, S.; Yoo, S.D.; Sheen, J. Differential regulation of EIN3 stability by glucose and ethylene signalling in plants. Nature 2003, 425, 521–525. [Google Scholar] [CrossRef]
- Beguerisse-Dıaz, M.; Hernández-Gómez, M.C.; Lizzul, A.M.; Barahona, M.; Desikan, R. Compound stress response in stomatal closure: A mathematical model of ABA and ethylene interaction in guard cells. BMC Syst. Biol. 2012, 6, 146. [Google Scholar] [CrossRef] [Green Version]
- Arve, L.E.; Carvalho, D.R.; Olsen, J.E.; Torre, S. ABA induces H2O2 production in guard cells, but does not close the stomata on Vicia faba leaves developed at high air humidity. Plant Signal. Behav. 2014, 9, e29192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, S.; Davies, W.J. Drought, ozone, ABA and ethylene: New insights from cell to plant to community. Plant Cell Environ. 2010, 33, 510–525. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Hazra, S.; Datta, R.; Chattopadhyay, S. Transcriptome analysis of arabidopsis mutants suggests a crosstalk between ABA, ethylene and GSH against combined cold and osmotic stress. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittler, R.; Blumwald, E. The roles of ROS and ABA in systemic acquired acclimation. Plant Cell 2015, 27, 64–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jannat, R.; Uraji, M.; Morofuji, M.; Islam, M.M.; Bloom, R.E.; Nakamura, Y.; McClung, C.R.; Schroeder, J.I.; Mori, I.C.; Murata, Y. Roles of intracellular hydrogen peroxide accumulation in abscisic acid signaling in arabidopsis guard cells. J. Plant Physiol. 2011, 168, 1919–1926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, J.; Song, C.P.; Wang, B.; Zhou, J.; Kangasjärvi, J.; Zhu, J.K.; Gong, Z. Reactive oxygen species signaling and stomatal movement in plant responses to drought stress and pathogen attack. J. Integr. Plant Biol. 2018, 60, 805–826. [Google Scholar] [CrossRef] [Green Version]
- Watkins, J.M.; Hechler, P.J.; Muday, G.K. Ethylene-induced flavonol accumulation in guard cells suppress reactive oxygen species and moderates stomatal aperture. Plant Physiol. 2014, 164, 1707–1717. [Google Scholar] [CrossRef] [Green Version]
- Watkins, J.M.; Chapman, J.M.; Muday, G.K. Abscisic acid-induced reactive oxygen Species are modulated by flavonols to control stomata aperture. Plant Physiol. 2017, 175, 1807–1825. [Google Scholar] [CrossRef] [Green Version]
- Shi, K.; Li, X.; Zhang, H.; Zhang, G.; Liu, Y.; Zhou, Y.; Xia, X.; Chen, Z.; Yu, J. Guard cell hydrogen peroxide and nitric oxide mediate elevated CO2−induced stomatal movement in tomato. New Phytol. 2015, 208, 342–353. [Google Scholar] [CrossRef] [Green Version]
- Munemasa, S.; Hirao, Y.; Tanami, K.; Mimata, Y.; Nakamura, Y.; Murata, Y. Ethylene inhibits methyl jasmonate-induced stomatal closure by modulating guard cell slow-type anion channel activity via the OPEN STOMATA 1/SnRK2.6 kinase-independent pathway in arabidopsis. Plant Cell Physiol. 2019, 60, 2263–2271. [Google Scholar] [CrossRef]
- Schroeder, J.I.; Keller, B.U. Two types of anion channel currents in guard cells with distinct voltage regulation. Proc. Natl. Acad. Sci. USA 1992, 89, 5025–5029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, L.; Liao, W. Hydrogen peroxide signaling in plant development and abiotic responses: Crosstalk with nitric oxide and calcium. Front. Plant Sci. 2016, 7, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.A.; Nazar, R.; Anjum, N.A. Growth, photosynthesis and antioxidant metabolism in mustard (Brassica juncea L.) cultivars differing in ATP-sulfurylase activity under salinity stress. Sci. Hortic. 2009, 122, 455–460. [Google Scholar] [CrossRef]
- Okuda, T.; Matsuda, Y.; Yamanaka, A.; Sagisaka, S. Abrupt increase in the level of hydrogen peroxide in leaves of winter wheat is caused by cold treatment. Plant Physiol. 1991, 97, 1265–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Lappartient, A.G.; Touraine, B. Demand-driven control of root ATP sulfurylase activity and SO42− Uptake in intact canola (the role of phloem-translocated glutathione). Plant Physiol. 1996, 111, 147–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Foyer, C.H.; Halliwell, B. The presence of glutathione and glutathione reductase in chloroplasts: A proposed role in ascorbic acid metabolism. Planta 1976, 133, 21–25. [Google Scholar] [CrossRef]
- Foyer, C.H.; Dujardyn, M.; Lemoine, Y. Responses of photosynthesis and the xanthophyll and ascorbate-glutathione cycles to changes in irradiance, photoinhibition and recovery. Plant Physiol. Biochem. 1989, 27, 751–760. [Google Scholar]
- Hossain, M.A.; Nakano, Y.; Asada, K. Monodehydroascorbate reductase in spinach chloroplasts and its participation in regeneration of ascorbate for scavenging hydrogen peroxide. Plant Cell Physiol. 1984, 25, 385–395. [Google Scholar]
- Griffith, O.W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Law, M.Y.; Charles, S.A.; Halliwell, B. Glutathione and ascorbic acid in spinach (Spinacia oleracea) chloroplasts. The effect of hydrogen peroxide and of Paraquat. Biochem. J. 1983, 210, 899–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avni, A.; Bailey, B.A.; Mattoo, A.K.; Anderson, J.D. Induction of ethylene biosynthesis in Nicotiana tabacum by a Trichoderma viride xylanase is correlated to the accumulation of 1-aminocyclopropane-1-carboxylic acid (ACC) synthase and ACC oxidase transcripts. Plant Physiol. 1994, 106, 1049–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woeste, K.E.; Ye, C.; Kieber, J.J. Two arabidopsis mutants that overproduce ethylene are affected in the posttranscriptional regulation of 1-aminocyclopropane-1-carboxylic acid synthase. Plant Physiol. 1999, 119, 521–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, K.T.; Kao, C.H. Nitric oxide counteracts the senescence of rice leaves induced by abscisic acid. J. Plant Physiol. 2003, 160, 871–879. [Google Scholar] [CrossRef] [Green Version]
- Daud, M.K.; Sun, Y.; Dawood, M. Cadmium-induced functional and ultrastructural alterations in roots of two transgenic cotton cultivars. J. Hazard. Mater. 2009, 161, 463–473. [Google Scholar] [CrossRef]
- Wang, J.; Sun, P.P.; Chen, C.L.; Wang, Y.; Fu, X.Z.; Liu, J.H. An arginine decarboxylase gene PtADC from Poncirus trifoliata confers abiotic stress tolerance and promotes primary root growth in arabidopsis. J. Exp. Bot. 2011, 62, 2899–2914. [Google Scholar] [CrossRef]
- Sandalio, L.M.; Dalurzo, H.C.; Gómez, M.; Romero-Puertas, M.C.; del Río, L.A. Cadmium-induced changes in the growth and oxidative metabolism of pea plants. J. Exp. Bot. 2001, 52, 2115–2126. [Google Scholar] [CrossRef]
- Hiscox, J.D.; Israelstam, G.F. A method for the extraction of chlorophyll from leaf tissue without maceration. Can. J. Bot. 1979, 57, 1332–1334. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Mancinelli, A. Photoregulation of anthocyanin synthesis. VIII. Effects of light pre-treatments. Plant Physiol. 1984, 75, 447–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.I.R.; Khan, N.A. Ethylene reverses photosynthetic inhibition by nickel and zinc in mustard through changes in PSII activity, photosynthetic nitrogen use efficiency, and antioxidant metabolism. Protoplasma 2014, 251, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Dudley, S.A. Differing selection on plant physiological traits in response to environmental water availability: A test of adaptive hypotheses. Evolution 1996, 50, 92–102. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Roots | Leaves | ||
|---|---|---|---|---|
| Na+ | Cl− | Na+ | Cl− | |
| (mg g−1 Leaf Fresh Weight) | ||||
| Control | 9.20 ± 1.03 e | 7.70 ± 0.34 e | 8.44 ± 0.70 e | 6.81 ± 0.58 e |
| NaCl | 27.31 ± 0.41 a | 17.90 ± 0.42 a | 22.36 ± 0.99 a | 15.64 ± 0.76 a |
| Ethephon (E) | 7.74 ± 0.66 f | 5.50 ± 0.29 f | 6.95 ± 0.63 f | 5.22 ± 0.62 f |
| Sulfur (S) | 7.66 ± 0.63 f | 5.21 ± 0.29 f | 6.64 ± 0.60 f | 5.03 ± 0.55 f |
| E + S | 6.61 ± 0.14 g | 4.12 ± 0.09 g | 4.89 ± 0.11 g | 4.17 ± 0.09 g |
| E + NaCl | 15.90 ± 0.73 b | 11.90 ± 0.59 b | 15.30 ± 0.99 b | 11.20 ± 0.65 b |
| S + NaCl | 14.55 ± 0.70 c | 10.90 ± 0.48 c | 12.72 ± 0.75 c | 9.51 ± 0.73 c |
| E + S + NaCl | 10.75 ± 1.07 d | 8.70 ± 0.28 d | 11.00 ± 0.89 d | 8.30 ± 0.65 d |
| Treatments | H2O2 | TBARS | APX | GR | DHAR | MDHAR | GSH | GSSG | Redox State |
|---|---|---|---|---|---|---|---|---|---|
| (nmol g−1 Leaf Fresh Weight) | (U mg−1 Protein min−1) | (nmol g−1 Leaf Fresh Weight) | |||||||
| Control | 16.1 ± 0.9 b | 4.20 ± 0.22 b | 1.22 ± 0.05 g | 0.204 ± 0.007 g | 90.1± 3.7 g | 60 ± 3 g | 58.2 ± 3.3 g | 2.9 ± 0.2 c | 21.0 ± 0.6 f |
| NaCl | 32.4 ± 1.0 a | 8.30 ± 0.23 a | 1.69 ± 0.06 f | 0.248 ± 0.009 f | 124.0 ± 4.1 f | 110 ± 3 f | 71.0 ± 3.6 f | 8.5 ± 0.6 a | 8.4 ± 0.6 g |
| Ethephon (E) | 8.7 ± 0.7 e | 2.70 ± 0.19 e | 3.02 ± 0.09 c | 0.363 ± 0.012 c | 162.0 ± 4.7 c | 144 ± 4 c | 98.6 ± 2.6 c | 3.3 ± 0.2 c | 30.0 ± 0.7 c |
| Sulfur (S) | 8.1 ± 0.8 e | 2.50 ± 0.18 e | 3.09 ± 0.09 c | 0.370 ± 0.013 c | 165.0 ± 4.8 c | 149 ± 4 c | 101.7 ± 3.9 c | 3.2 ± 0.7 c | 31.1 ± 0.7 c |
| E + S | 4.1 ± 0.5 g | 1.64 ± 0.14 g | 3.90 ± 0.11 a | 0.435 ± 0.014 a | 196.0 ± 5.1 a | 172 ± 5 a | 115.4 ± 3.5 a | 3.0 ± 1.0 c | 38.4 ± 1.0 a |
| E + NaCl | 13.2 ± 0.8 c | 3.70 ± 0.20 c | 2.12 ± 0.07 e | 0.311 ± 0.010 e | 135.0 ± 4.5 e | 121 ± 4 e | 80.4 ± 2.5 e | 3.8 ± 0.5 b | 21.3 ± 0.7 e |
| S + NaCl | 11.6 ± 0.6 d | 3.20 ± 0.19 d | 2.43 ± 0.08 d | 0.341 ± 0.011 d | 147.0 ± 4.7 d | 133 ± 4 d | 88.0 ± 2.5 d | 3.7 ± 0.6 b | 24.1 ± 0.7 d |
| E + S + NaCl | 5.7 ± 0.7 f | 2.00 ± 0.17 f | 3.40 ± 0.10 b | 0.400 ± 0.012 b | 180.0 ± 5.0 b | 160 ± 5 b | 110.3 ± 3.2 b | 3.1 ± 0.4 c | 36.1 ± 1.0 b |
| Treatments | ATP-S Activity | S | AsA | DHA |
|---|---|---|---|---|
| (µmol g−1 Protein s−1) | (mg g−1 Fresh Weight) | (nmol g−1 Fresh Weight) | (nmol g−1 Fresh Weight) | |
| Control | 1.24 ± 0.06 g | 5.0 ± 0.2 f | 22.31 ± 0.77 g | 05.28 ± 0.32 g |
| NaCl | 1.44 ± 0.04 f | 4.1 ± 0.1 g | 27.50 ± 0.98 f | 09.20 ± 0.33 f |
| Ethephon (E) | 2.31 ± 0.07 c | 7.7 ± 0.3 c | 37.62 ± 1.10 c | 15.40 ± 0.39 d |
| Sulfur (S) | 2.36 ± 0.07 c | 7.9 ± 0.3 c | 38.28 ± 1.13 c | 15.84 ± 0.41 c |
| E + S | 2.87 ± 0.08 a | 9.1 ± 0.3 a | 46.20 ± 1.34 a | 18.48 ± 0.41 a |
| E + NaCl | 1.88 ± 0.06 e | 6.4 ± 0.3 e | 31.51 ± 1.03 e | 13.11 ± 0.33 e |
| S + NaCl | 2.15 ± 0.06 d | 7.0 ± 0.3 d | 34.27 ± 1.09 d | 14.26 ± 0.37 d |
| E + S + NaCl | 2.54 ± 0.08 b | 8.6 ± 0.3 b | 43.24 ± 1.28 b | 17.71 ± 0.35 b |
| Treatments | Chl a | Chl b | Total Chl | Carotenoids | Anthocyanins |
|---|---|---|---|---|---|
| (mg g−1 Leaf Fresh Weight) | |||||
| Control | 1.31 ± 0.05 f | 0.62 ± 0.02 f | 2.00 ± 0.07 f | 0.57 ± 0.02 f | 0.61 ± 0.03 g |
| NaCl | 1.09 ± 0.05 g | 0.41 ± 0.01 g | 1.56 ± 0.06 g | 0.46 ± 0.02 g | 1.33 ± 0.05 f |
| Ethephon (E) | 1.58 ± 0.06 c | 0.82 ± 0.02 c | 2.40 ± 0.08 c | 0.69 ± 0.03 c | 1.61 ± 0.06 c |
| Sulfur (S) | 1.61 ± 0.06 c | 0.84 ± 0.03 c | 2.43 ± 0.08 c | 0.71 ± 0.03 c | 1.64 ± 0.06 c |
| E + S | 1.81 ± 0.07 a | 1.05 ± 0.03 a | 2.86 ± 0.09 a | 0.83 ± 0.04 a | 1.82 ± 0.07 a |
| E + NaCl | 1.41 ± 0.06 e | 0.68 ± 0.02 e | 2.10 ± 0.07 e | 0.60 ± 0.03 e | 1.46 ± 0.06 e |
| S + NaCl | 1.49 ± 0.04 d | 0.75 ± 0.02 d | 2.24 ± 0.08 d | 0.66 ± 0.03 d | 1.54 ± 0.06 d |
| E + S + NaCl | 1.70 ± 0.07 b | 0.94 ± 0.03 b | 2.64 ± 0.09 b | 0.78 ± 0.03 b | 1.73 ± 0.06 b |
| Treatments | Maximal PSII Photochemical Efficiency | Net Photosynthesis | Intercellular CO2 Concentration | Stomatal Conductance | Water Use Efficiency | Leaf Area | Plant Fresh Weight |
|---|---|---|---|---|---|---|---|
| (µmol CO2 m−2 s−1) | (µmol CO2 mol−1) | (mmol H2O m−2 s−1) | (µmol mol−1) | (cm2 Plant−1) | (g Plant−1) | ||
| Control | 0.78 ± 0.02 f | 13.7 ± 0.6 f | 257 ± 10 f | 377 ± 11 f | 37.1 ± 1.5 f | 117.0 ± 3.9 f | 2.15 ± 0.06 f |
| NaCl | 0.65 ± 0.01 g | 8.1 ± 0.6 g | 176 ± 7 g | 298 ± 9 g | 29.4 ± 1.3 g | 59.4 ± 2.5 g | 1.01 ± 0.05 g |
| Ethephon (E) | 0.90 ± 0.02 c | 19.3 ± 0.7 c | 357 ± 14 c | 481 ± 19 c | 49.1 ± 2.3 c | 164.8 ± 5.1 c | 2.94 ± 0.09 c |
| Sulfur (S) | 0.91 ± 0.02 c | 19.6 ± 0.7 c | 365 ± 15 c | 487 ± 19 c | 50.0 ± 2.3 c | 168.7 ± 5.3 c | 3.00 ± 0.08 c |
| E + S | 0.99 ± 0.02 a | 22.2 ± 0.7 a | 433 ± 16 a | 590 ± 22 a | 60.0 ± 2.4 a | 207.0 ± 5.8 a | 3.81 ± 0.09 a |
| E +NaCl | 0.83 ± 0.02 e | 16.5 ± 0.8 e | 300 ± 12 e | 408 ± 15 e | 40.8 ± 1.6 e | 138.7 ± 4.7 e | 2.56 ± 0.07 e |
| S + NaCl | 0.87 ± 0.02 d | 18.2 ± 0.6 d | 329 ± 13 d | 449 ± 17 d | 43.8 ± 1.7 d | 155.7 ± 4.8 d | 2.78 ± 0.08 d |
| E + S + NaCl | 0.96 ± 0.02 b | 21.4 ± 0.7 b | 398 ± 15 b | 550 ± 21 b | 56.0 ± 2.5 b | 185.0 ± 5.5 b | 3.54 ± 0.09 b |
| Treatments | ABA | H2O2 | Net Photosynthesis | Stomatal Conductance |
|---|---|---|---|---|
| (pmol g−1 Fresh Weight) | (nmol g−1 Fresh Weight) | (µmol CO2 m−2 s−1) | (mmol H2O m−2 s−1) | |
| Control | 70.0 ± 5.2 e | 16.4 ± 0.9 d | 13.0 ± 0.6 c | 373 ± 11 c |
| NaCl | 250.0 ± 8.8 a | 31.9 ± 1.1 a | 7.5 ± 0.6 f | 275 ± 9 e |
| E + S | 54.7 ± 4.4 f | 4.3 ± 0.6 f | 27.6 ± 1.1 a | 582 ± 22 a |
| E + S + NaCl | 101.0 ± 8.3 d | 8.3 ± 0.3 e | 24.0 ± 0.8 b | 540 ± 20 b |
| NBD | 71.2 ± 6.0 e | 22.0 ± 0.8 c | 12.1 ± 0.7 d | 351 ± 12 d |
| E + S + NaCl + NBD | 230.4 ± 8.9 b | 28.0 ± 0.9 b | 8.1 ± 0.6 e | 255 ± 10 f |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fatma, M.; Iqbal, N.; Gautam, H.; Sehar, Z.; Sofo, A.; D’Ippolito, I.; Khan, N.A. Ethylene and Sulfur Coordinately Modulate the Antioxidant System and ABA Accumulation in Mustard Plants under Salt Stress. Plants 2021, 10, 180. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10010180
Fatma M, Iqbal N, Gautam H, Sehar Z, Sofo A, D’Ippolito I, Khan NA. Ethylene and Sulfur Coordinately Modulate the Antioxidant System and ABA Accumulation in Mustard Plants under Salt Stress. Plants. 2021; 10(1):180. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10010180
Chicago/Turabian StyleFatma, Mehar, Noushina Iqbal, Harsha Gautam, Zebus Sehar, Adriano Sofo, Ilaria D’Ippolito, and Nafees A. Khan. 2021. "Ethylene and Sulfur Coordinately Modulate the Antioxidant System and ABA Accumulation in Mustard Plants under Salt Stress" Plants 10, no. 1: 180. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10010180