
A Review and Secondary Analysis of Competition-Related Impacts of Nonindigenous Aquatic Plants in the Laurentian Great Lakes




Abstract
:1. Introduction
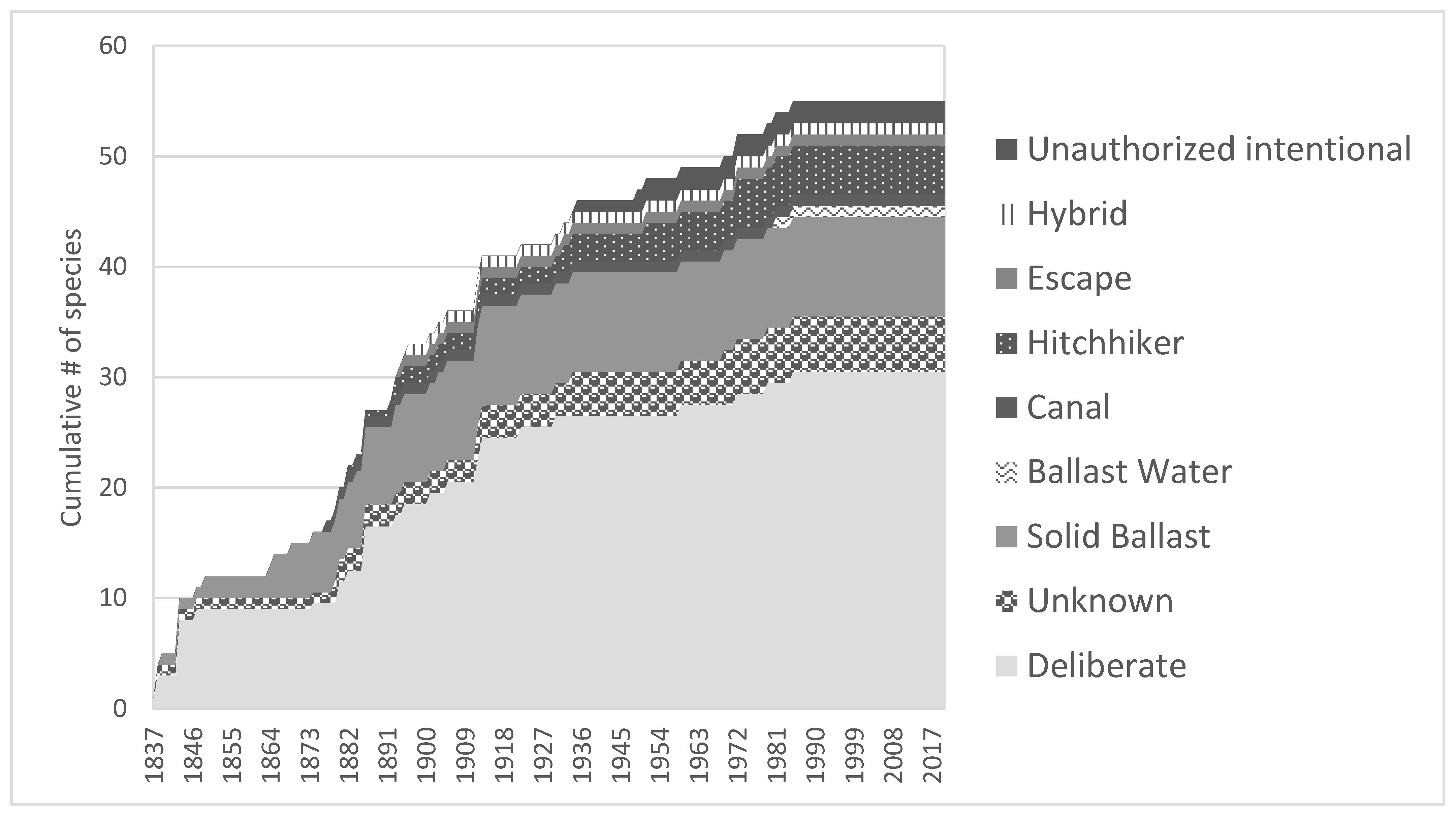
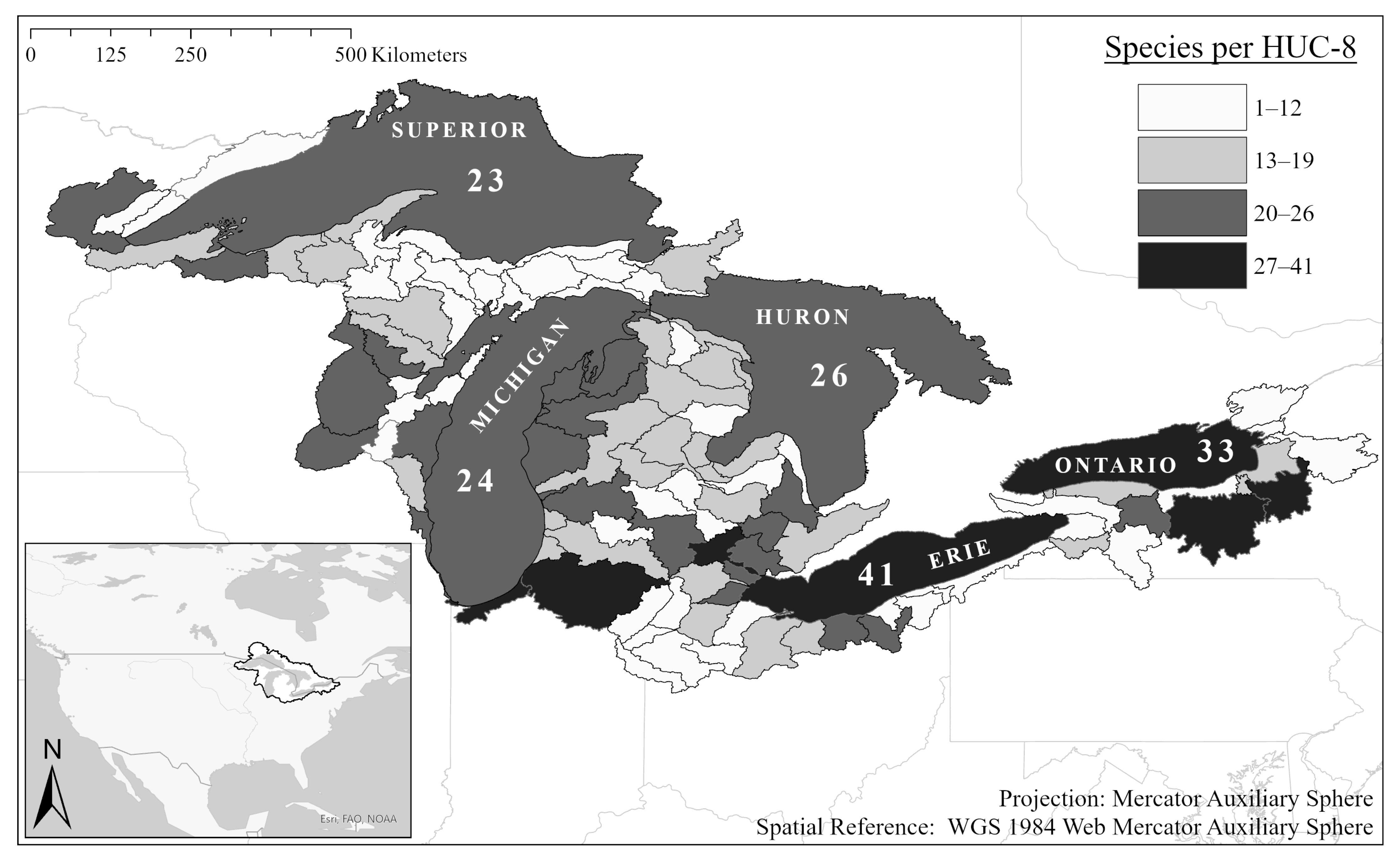
2. Results
- extirpated (Chenopodium capitatum (OH = Ohio), Chenopodium leptophyllum (OH), Juncus greenei (PA = Pennsylvania), and Juncus militaris (IN = Indiana));
- endangered (Chenopodium foggii (PA), Echinochloa walteri (PA), Epilobium angustifolium (IN and OH) Lysimachia radicans (IL = Illinois), Epilobium strictum (PA), Iris brevicaulis (OH), Iris cristata (PA), Iris verna (PA), Iris virginica var. shervei (NY = New York and PA), Rhamnus alnifolia (IL), Rhamnus lanceolata (PA), Myriophyllum sibericum (PA), Juncus alpinus (IL), Juncus ambiguous (NY), Juncus biflorus (NY), Juncus brachycephalus (NY and PA), Juncus dichotomus (OH and PA), Juncus diffusissimus (OH), Juncus ensifolius (NY), Juncus greenei (OH), Juncus interior (OH), Juncus militaris (PA), Juncus scirpoides (NY and PA), Juncus secundus (IN), Juncus stygius (WI = Wisconsin), Juncus subcaudatus (NY), Juncus vaseyi (IL), Lycopus amplectens (IN), and Lycopus rubellus (NY and PA));
- threatened (Chenopodium subglabrum (CAN = Canada), Cirsium pitcheri (US = United States, IL, IN, MI = Michigan, WI, and ONT = Ontario), Epilobium strictum (IL and OH), Iris lacustris (US, MI, and WI), Iris verna (OH), Myriophyllum sibiricum (OH), Juncus alpinus (PA), Juncus biflorus (PA), Juncus brachycephalus (MI), Juncus militaris (MI), Juncus pelocarpus (IN), Juncus scirpoides (IN and MI), Juncus secundus (OH), Juncus stygius (MI), Juncus vaseyi (MI), and Lycopus virginicus (MI));
- special concern (Cirsium pitcheri (CAN), Iris lacustris (CAN and ONT));
- vulnerable (Ilex verticillata (NY)); and
- endemic (Cirsium pitcheri, Iris lacustris, and Iris robusta) species in the Great Lakes region.
3. Discussion
4. Materials and Methods
- Geographic criterion: Only species that are found in the Great Lakes basin below the ordinary high water mark—including connecting channels, wetlands, and waters ordinarily attached to the Lakes—were included in the GLANSIS nonindigenous species list. Species that have been collected from inland lakes within the Great Lakes basin but do not meet this geographic criterion were excluded.
- Aquatic criterion: The United States Department of Agriculture (USDA) wetland indicator status was used as a guideline for determining whether wetland plants should be included in the list. Obligate, facultative wetland, and facultative plants were included as aquatic. Facultative upland and upland plants were excluded, even if found below the ordinary high water mark.
- Nonindigenous criterion: The species included in the GLANSIS nonindigenous list were those that met at least three of the following criteria based on Ricciardi 2006 [18]:
- ⚬
- The species appeared suddenly and had not been recorded in the basin previously.
- ⚬
- It subsequently spreads within the basin.
- ⚬
- Its distribution in the basin is restricted compared with native species.
- ⚬
- Its global distribution is anomalously disjunct (containing widely scattered and isolated population).
- ⚬
- Its global distribution is associated with human vectors of dispersal.
- ⚬
- The basin is isolated from regions possessing the most genetically and morphologically similar species.
- Reproducing and overwintering criterion: A nonindigenous species was considered to be in at least the early stages of establishment if it had a reproducing population within the basin and was capable of overwintering, as inferred from multiple discoveries of adult and juvenile life stages over at least two consecutive years. Given that successful establishment may require multiple introductions, species were excluded if their records of discoveries were based on only one or a few non-reproducing individuals whose occurrence may reflect merely transient species or unsuccessful invasions.
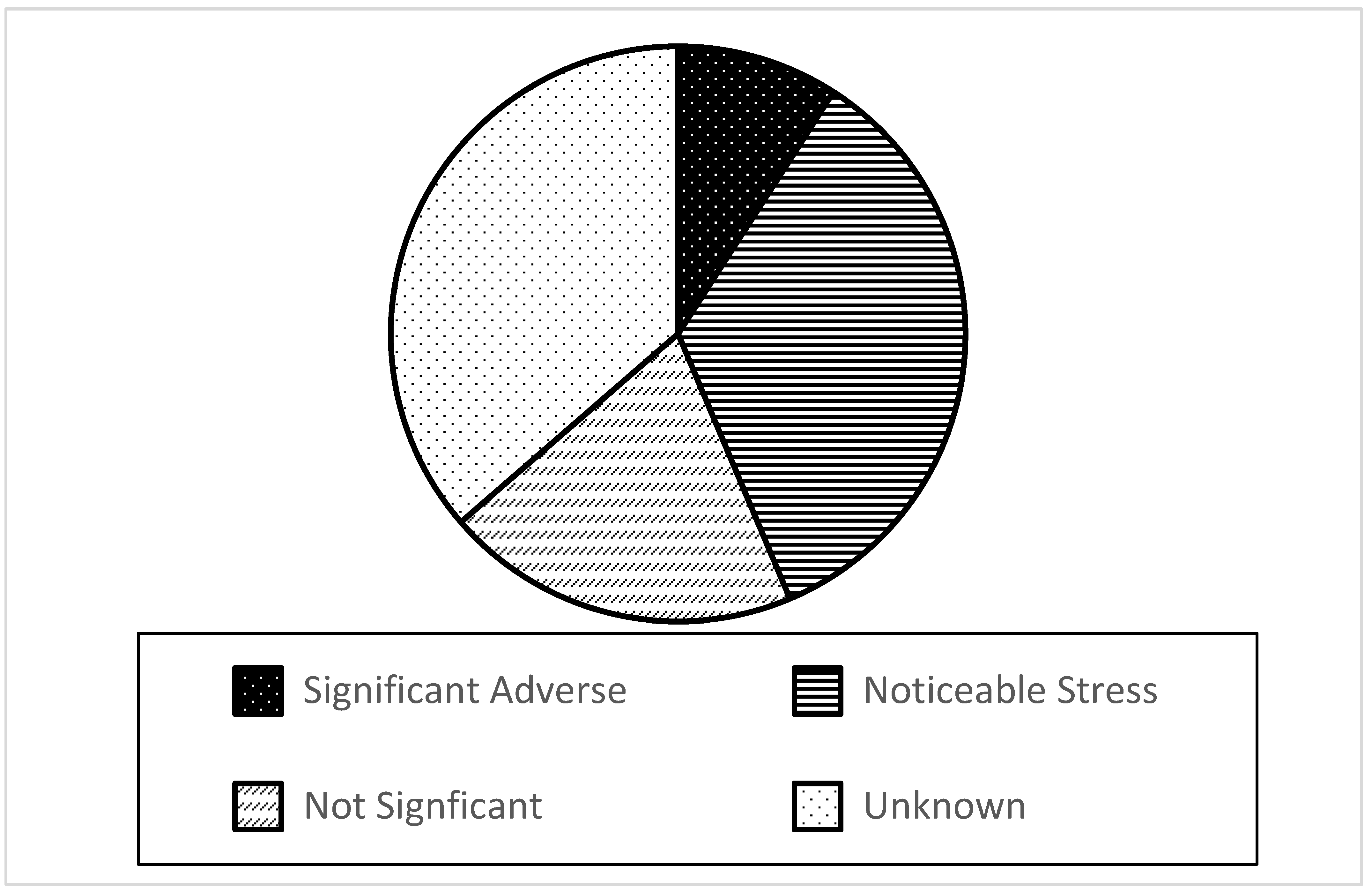
- yes, and it has resulted in significant adverse effects (e.g., critical reduction, extinction, and behavioral changes) on one or more native species populations;
- yes, and it has caused some noticeable stress to or decline of at least one native species population;
- not significantly; or
- unknown.
5. Conclusions
- For 35% of the aquatic nonindigenous plants species in the Great Lakes, it is unknown whether they have an outcome-based competitive impact on native plants. Further study of potential competitive impacts is needed for each of these 20 species.
- While for many nonindigenous aquatic plant species extensive information is available on species traits that may contribute to giving the nonindigenous species a competitive advantage over native species, a direct mechanistic-based examination of the competition between these species is generally lacking and is an area ripe for future research.
- For some nonindigenous aquatic plant species, competitive impacts to native species are documented only at the group level (e.g., impact to native sedges). More work is needed to understand the impacts on particular native plant species within these broader groups, which may include additional at risk natives.
- Competition is only one interaction through which nonindigenous aquatic plants can impact the ecology and economy of the Great Lakes region. More research is needed to place competitive impacts in the context of the broader suite of impact mechanisms.
- The potential alteration of several adaptations (e.g., environmental tolerance and light competition) as a result of climate change should be studied to properly understand how the nature of competition between nonindigenous and native plants may change under various climate scenarios.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Strayer, D.L. Alien species in fresh waters: Ecological effects, interactions with other stressors, and prospects for the future. Freshw. Biol. 2010, 55, 152–174. [Google Scholar] [CrossRef]
- Simberloff, D.; Martin, J.L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Curchamp, F.; Galil, B.; García-Bertou, E.; Pascal, M.; et al. Impacts of biological invasions: what’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Winton, M.; Clayton, J. The impact of invasive submerged weed species on seed banks in lake sediments. Aquat. Bot. 1996, 53, 32–46. [Google Scholar] [CrossRef]
- Hussner, A.; Van de Weyer, K.; Gross, E.M.; Hilts, S. Comments on increasing number and abundance of non-indigenous aquatic macrophyte species in Gremany. Weed Res. 2010, 50, 519–526. [Google Scholar] [CrossRef]
- Sheppard, A.W.; Shaw, R.H.; Sforza, R. Top 20 environmental weeds for classical biological control in Europe: A review of opportunities, regulations and other barriers to adoption. Weed Res. 2006, 46, 93–117. [Google Scholar] [CrossRef]
- Hussner, A. Alien aquatic plant species in European countries. Weed Res. 2012, 52, 297–306. [Google Scholar] [CrossRef]
- Brundu, G. Plant invaders in European and Mediterranean inland waters: Profiles, distribution, and threats. Hydrobiologia 2015, 746, 61–79. [Google Scholar] [CrossRef]
- Vilas, M.P.; Marti, C.L.; Adams, M.P.; Oldham, C.E.; Hipsey, M.R. Invasive macrophytes control the spatial and temporal patterns of temperature and dissolved oxygen in a shallow lake: A proposed feedback mechanism of macrophyte loss. Front. Plant. Sci. 2017, 8, 2097. [Google Scholar] [CrossRef] [Green Version]
- Martin, G.D.; Coetzee, J.A. Pet stores, aquarists and the internet trade as modes of introduction and spread of invasive macrophytes in South Africa. Water 2011, 37, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Thomaz, S.M.; Mormul, R.P.; Michelan, T.S. Propagule pressure, ivasibility of freshwater ecosystems by macrophytes and their ecological impacts: A review of tropical freshwater ecosystems. Hydrobiologia 2014, 746, 39–59. [Google Scholar] [CrossRef]
- Sturtevant, R.A.; Mason, D.M.; Rutherford, E.S.; Elgin, A.K.; Lower, E.K.; Martinez, F.A. Recent history of nonindigenous species in the Laurentian Great Lakes; An update to Mills et al., 1993 (25 years later). J. Great Lakes Res. 2019, 45, 1011–1035. [Google Scholar] [CrossRef]
- Les, D.H.; Mehrhoff, L.J. Introduction of nonindigenous aquatic vascular plants in southern New England: A historical perspective. Biol. Invasions 1999, 1, 281–300. [Google Scholar] [CrossRef]
- Fleming, J.P.; Dibble, E.D. Ecological mechanisms of invasion success in aquatic macrophytes. Hydrobiologia 2015, 746, 23–37. [Google Scholar] [CrossRef]
- Fargione, J.; Tilman, D. Competition and Coexistence in Terrestrial Plants. In Competition and Coexistence. Ecological Studies (Analysis and Synthesis); Springer: Berlin/Heidelberg, Germany, 2002; Volume 161. [Google Scholar]
- Tilman, D. Resource competition between planktonic algae: An experimental and theoretical approach. Ecology 1977, 58, 338–348. [Google Scholar] [CrossRef]
- Great Lakes Restoration Initiative (GLRI) Task Force. GLRI Action Plan FY2010-FY2014; US EPA Great Lakes National Program Office: Chicago, IL, USA, 2010. [Google Scholar]
- Mills, E.L.; Leach, J.H.; Carlton, J.T.; Secor, C.L. Exotic species in the Great Lakes: A history of biotic crises and anthropogenic introductions. J. Great Lakes Res. 1993, 19, 1–54. [Google Scholar] [CrossRef]
- Ricciardi, A. Patterns of invasion in the Laurentian Great Lakes in relation to changes in vector activity. Divers. Distrib. 2006, 12, 425–433. [Google Scholar] [CrossRef]
- Executive Order (E.O.) No. 13112. In Fed. Regist.; 1999; 64, pp. 6183–6186. Available online: https://www.federalregister.gov/presidential-documents/executive-orders (accessed on 15 January 2021).
- Gopal, B.; Goel, U. Competition and Allelopathy in Aquatic Communities. Bot. Rev. 1993, 59, 155–210. [Google Scholar] [CrossRef]
- Wetzel, R.G. Limnology, 2nd ed.; Saunders College Publishing: Philadelphia, PA, USA, 1983. [Google Scholar]
- NOAA. Great Lakes Aquatic Nonindigenous Species Information System (GLANSIS). Available online: https://www.glerl.noaa.gov/glansis/ (accessed on 1 September 2020).
- Clements, F.E. The development and structure of vegetation. Bot. Surv. Nebraska 1904, 7, 5–175. [Google Scholar]
- Grace, J.B.; Tilman, D. Perspectives on Plant Competition; Academic Press: San Diego, CA, USA, 1990. [Google Scholar]
- Chambers, P.A.; Prepas, E.E. Competition and coexistence in submerged aquatic plant communities: The effects of species interactions versus abiotic factors. Freshw. Biol. 1990, 23, 541–550. [Google Scholar] [CrossRef]
- Aschehoug, E.T.; Brooker, R.; Atwater, D.Z.; Maron, J.L.; Callaway, R.M. The Mechanisms and consequences of interspecific competition among plants. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 263–281. [Google Scholar] [CrossRef] [Green Version]
- Mangala, S.; Sheley, R.L.; James, J.J.; Radosevich, S.R. Intra and interspecific competition among invasive and native species during early stages of plant growth. Plant Ecol. 2011, 212, 531–542. [Google Scholar] [CrossRef]
- Reznicek, A.A.; Voss, E.G.; Walters, B.S. Michigan Flora Online; University of Michigan Herbarium, University of Michigan: Ann Arbor, MI, USA, 2011; Available online: http://michiganflora.net/home.aspx (accessed on 1 January 2018).
- Sturtevant, R.; Larson, J.; Berent, L.; McCarthy, M.; Bogdanoff, A.; Fusaro, A.; Rutherford, E. An Impact Assessment of Great Lakes Aquatic Nonindigenous Species; NOAA Technical Memorandum GLERL-161; NOAA-GLERL: Ann Arbor, MI, USA, 2014. [Google Scholar]
- Sturtevant, R.A.; Lower, E.K.; Boucher, N.; Alsip, P.J.; Hopper, K.; Iott, S.; Mason, D.M.; Elgin, A.K.; Martinez, F.A. 2018 Update to “An Impact Assessment of Great Lakes Aquatic Nonindigenous Species.”; NOAA Technical Memorandum GLERL-161b; NOAA-GLERL: Ann Arbor, MI, USA, 2019. [Google Scholar]
- Lower, E.K.; Boucher, N.; Sturtevant, R.A.; Elgin, A.K. 2019 Update to “An Impact Assessment of Great Lakes Aquatic Nonindigenous Species”; NOAA Technical Memorandum GLERL-161c; NOAA-GLERL: Ann Arbor, MI, USA, 2020. [Google Scholar]
- Bhowmik, P.C.; Reddy, K.N. Effects of barnyardgrass (Echinochloa crus-galli) on growth, yield, nutrient status of transplanted tomato (Lycopersicon esculentum). Weed Sci. 1988, 36, 775–778. [Google Scholar] [CrossRef]
- PLANTS Team, United States Department of Agriculture (USDA) Natural Resources Conservation Service (NRCS). The PLANTS Database. Available online: http://plants.usda.gov/threat.html (accessed on 15 January 2021).
- Ohio Department of Natural Resources (OHDNR). Rare Native Ohio Plants 2010–2011 Status List; OHDNR: Columbus, OH, USA, 2012. [Google Scholar]
- Valley, R.D.; Newman, R.M. Competitive interactions between Eurasian watermilfoil and northern watermilfoil in experimental tanks. J. Aquat. Plant Manag. 1998, 36, 121–126. [Google Scholar]
- Lavoie, C.; Jean, M.; Delisle, F.; Létourneau, G. Exotic plant species of the St Lawrence River wetlands: A spatial and historical analysis. J. Biogeogr. 2003, 30, 537–549. [Google Scholar] [CrossRef]
- State of Washington; Department of Ecology. Non-Native Invasive Freshwater Plants: Garden Loosestrife (Lysimachia vulgaris); State of Washington: Olympia, WA, USA, 2013. [Google Scholar]
- Indiana Natural Heritage Database. Illinois Threatened and Endangered Species by County; Indiana Natural Heritage Data Center: Indianapolis, IN, USA, 2011; 126p. [Google Scholar]
- Committee on the Status of Species at Risk in Ontario, (COSSARO). COSSARO Candidate Species at Risk Evaluation form for Pitcher’s Thistle (Cirsium pitcheri); COSSARO: Fergus, ON, Canada, 2011; Available online: http://142.150.190.63/node/10251 (accessed on 15 January 2021).
- Great Lakes Indian Fish & Wildlife Commission, (GLIFWC). Eurasian Marsh Thistle (Cirsium palustre). 2006. Available online: http://www.glifwc.org/invasives/Cirsium_palustre/control.html (accessed on 15 May 2012).
- Sheehan, M. Wisconsin Impact Assessment: Cirsium palustre (L.) Scop; WI DNR: Madison, WI, USA, 2007. [Google Scholar]
- Naylor, M. Water Chestnut (Trapa natans) in the Chesapeake Bay. Md. Dep. Nat. Resour. 2003, 10, 1–35. [Google Scholar]
- Vaccaro, L.E.; Bedford, B.L.; Johnston, C.A. Litter accumulation promotes dominance of invasive species of cattail (Typha spp.) in Lake Ontario wetlands. Wetlands 2009, 29, 1036–1048. [Google Scholar] [CrossRef]
- Weisner, S.E.B. Long-term competitive displacement of Typha latifolia by Typha angustifolia in a eutrophic lake. Oecologia 1993, 94, 451–456. [Google Scholar] [CrossRef]
- Raven, P.H.; Thomas, J.H. Iris pseudacorus in western North America. Madrono 1970, 20, 390–391. [Google Scholar]
- Cox, G.W. Alien species in North America and Hawaii: Impacts on Natural Ecosystems; Island Press: Washington, DC, USA, 1999. [Google Scholar]
- Frappier, B.F.; Eckert, R.T.; Lee, T.D. Potential impacts of invasive exotic shrub Rhamnus frangula L. (glossy buckthorn) on forests of south New Hampshire. Northeast. Nat. 2003, 10, 277–296. [Google Scholar] [CrossRef]
- Hilt, S.; Henschke, I.; Rucker, J.; Nixdorf, B. Can submerged macrophytes influence turbidity and trophic state in deep lakes? Suggestions from a case study. J. Environ. Qual. 2010, 39, 725–733. [Google Scholar] [CrossRef]
- Mulderij, G.; Van Nes, E.H.; Van Donk, E. Macrophyte-phytoplankton interactions: The relative importance of allelopathy versus other factors. Ecol. Model 2007, 204, 85–92. [Google Scholar] [CrossRef]
- Wink, M. Inhibition of seed germination by quinolizidine alkaloids. Planta 1983, 158, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Muzquiz, M.; de la Cuadra, C.; Cuadrado, C.; Burbano, C.; Calvo, R. Herbicide-like effect of Lupinus alkaloids. Ind. Crop. Prod. 1994, 2, 273–280. [Google Scholar] [CrossRef]
- Noxious Weed Control Program. Yellow-Flag Iris (Iris Pseudacorus). Best Management Practices; Department of Natural Resources and Parks; Water and Land Resources Division: King County, WA, USA, 2009. [Google Scholar]
- Mudrák, O.; Hermová, M.; Tesnerová, C.; Rydlová, J.; Frouz, J. Above-ground and below-ground competition between the willow Salix caprea and its understory. J. Veg. Sci. 2016, 27, 156–164. [Google Scholar] [CrossRef]
- Al-Jaber, N.A.; Mujahid, T.G.; Al-Hazimi, H.M.G. Secondary metabolites of Chenopodiaceace species. J. Chem. Soc. Pak. 1992, 14, 76–83. [Google Scholar]
- Larina, S.Y. Chenopodium glaucum. In Interactive Agricultural Ecological Atlas of Russia and Neighboring Countries; Afonin, A.N., Greene, S.L., Dzyubenko, N.I., Frolov, A.N., Eds.; Economic Plants and their Diseases, Pests and Weeds; US Department of Agriculture: Washington, DC, USA, 2008. [Google Scholar]
- Pitcher, D. Element Stewardship Abstract for Conium maculatum L.; The Nature Conservancy: Arlington, VA, USA, 2004. [Google Scholar]
- López, T.A.; Cid, M.S.; Bianchini, M.L. Biochemistry of hemlock (Conium maculatum L.) alkaloids and their acute and chronic toxicity in livestock. A review. Toxicon 1999, 37, 841–865. [Google Scholar] [CrossRef]
- Castells, E.; Berenbaum, M.R. Laboratory rearing of Agonopterix alstroemeriana, the defoliating poison hemlock (Conium maculatum L.) moth, and the effects of piperidine alkaloids on preference and performance. Environ. Entomol. 2006, 35, 607–615. [Google Scholar] [CrossRef] [Green Version]
- Castells, E.; Berenbaum, M.R. Resistance of the generalist moth Trichoplusia ni (Noctuidae) to a novel chemical defense in the invasive plant Conium maculatum. Chemoecology 2008, 18, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Granberg, K.; Gulezian, P.; Ison, J. Keeping the Chicago Region Free of Poison Hemlock: An Interdisciplinary Approach to Studying the Invasion Front of a Known Invasive Plant; Final report to Chicago Wilderness and Illinois Conservation Foundation; University of Illinois at Chicago: Chicago, IL, USA, 2009; 15p. [Google Scholar]
- Cooperrider, T.S.; Galang, M.M. A Pluchea Hybrid from the Pacific. Am. J. Bot. 1965, 52, 1020–1026. [Google Scholar] [CrossRef]
- Flora of North America. Pluchea odorata (Linnaeus) Cassini var. succulenta (Fernald). Available online: http://www.efloras.org/florataxon.aspx?flora_id=1&taxon_id=250068659 (accessed on 2 May 2013).
- Sheahan, C.M. Plant Guide for Seaside Goldenrod (Solidago sempervirens); USDA-Natural Resources Conservation Service, Cape May Plant Materials Center: NJ, USA, 2014. [Google Scholar]
- Peeters, A. Profiles of individual grass species: Alopecurus geniculatus L. In Wild and Sown Grasses: Profiles of a Temperate Species Selection: Ecology, Biodiversity, and Use; Blackwell Publishing for the Food and Agriculture Organization of the United Nations: Rome, Italy, 2004; pp. 82–84. [Google Scholar]
- Virtue, J.G.; Melland, R.L. The Environmental Weed Risk of Revegetation and Foresty Plants; Report DWLBC 2003/02; The Department of Water, Land and Biodiversity Conservation: Urrbrae, South Australia, 2003. [Google Scholar]
- Miklovic, S. Typha angustifolia Management: Implications for Glacial Marsh Restoration; Department of Horticultural Science, University of Minnesota: St. Paul, MN, USA, 2000. [Google Scholar]
- Ohio Environmental Protection Agency (Ohio EPA). Fact Sheet 11: Narrow-leaved and Hybrid Cattail. Invasive Plants of Ohio; Ohio Division of Natural Areas and Preserves, The Nature Conservancy- The Ohio Chapter, Columbus and Franklin County Metro Parks: Columbus, OH, USA, 2001; 2p. [Google Scholar]
- Stevens, M.; Hoag, C. Narrowleaf Cattail, Typha angustifolia L. Plant Guide; United States Department of Agriculture (USDA), Natural Resources Conservation Service (NRCS): Washington, DC, USA, 2006. [Google Scholar]
- Higgins, S.I.; Richardson, D.M. Invasive plants have broader physiological niches. Proc. Natl. Acad. Sci. USA 2014, 111, 10610–10614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, R.; Dibble, E. Effects of invasive macrophytes on freshwater fish and macroinvertebrate communities: The role of invasive plant traits. Hydrobiologia 2012, 684, 1–14. [Google Scholar] [CrossRef]
- Thuiller, W.; Gassó, N.; Pino, J.; Vilà, M. Ecological niche and species traits: Key drivers of regional plant invader assemblages. Biol. Invasions 2012, 14, 1963–1980. [Google Scholar] [CrossRef]
- Mamolos, A.P.; Veresoglou, D.S. Patterns of root activity and responses of species to nutrients in vegetation of fertile alluvial soil. Plant Ecol. 2000, 148, 245–253. [Google Scholar] [CrossRef]
- Wentz, W.A.; Stuckey, R.L. The changing distribution of the genus Najas (Najadaceae) in Ohio. Ohio J. Sci. 1971, 71, 292–302. [Google Scholar]
- Hroudová, Z.; Krahulcová, A.; Zákravský, P.; Jarolímová, V. The biology of Butomus umbellatus in shallow waters with fluctuating water level. Hydrobiologia 1996, 340, 27–30. [Google Scholar]
- Australian Department of the Environment and Heritage. Weeds of National Significance: Weed Management Guide; Department of the Environment and Heritage and the CRC for Australian Weed Management: Adelaide, Australia, 2003. [Google Scholar]
- State of Washington, Department of Ecology. Hairy Willow-Herb (Epilobium hirsutum). Non-Native Invasive Freshwater Plants. 2012. Available online: http://www.ecy.wa.gov/programs/wq/plants/weeds/willowherb.html (accessed on 14 May 2012).
- King County Noxious Weed Control Program. Hairy Willowherb. King County Department of Natural Resources and Parks; Water and Land Resources Division: Seattle, WA, USA, 2008; 2p. [Google Scholar]
- Etherington, J.R. Comparative studies of plant growth and distribution in relation to waterlogging: X. Differential formation of adventitious roots and their experimental excision in Epilobium hirsutum and Chamerion angustifolium. J. Ecol. 1984, 72, 389–404. [Google Scholar] [CrossRef]
- Campbell, S.; Higman, P.; Slaughter, B.; Schools, E. A Field Guide to Invasive Plants of Aquatic and Wetland Habitats for Michigan; Michigan DNRE, Michigan State University Extension, Michigan Natural Features Inventory; MSU: East Lansing, MI, USA, 2010. [Google Scholar]
- King County Noxious Weed Program. Policeman’s Helmet Best Management Practices; Department of Natural Resources and Parks, Water and Land Resources Division: Seattle, WA, USA, 2010; 6p. [Google Scholar]
- Midwest Invasive Species Information Network and Michigan Natural Features Inventory (MISIN and MNFI). Garden Yellow Loosestrife (Lysimachia Vulgaris). 2013. Available online: http://www.misin.msu.edu/facts/detail.php?id=92 (accessed on 3 May 2013).
- Hilty, J. Illinois Wildflowers. 2008. Available online: http://www.illinoiswildflowers.info/weeds/plants/water_chickweed.htm (accessed on 4 September 2020).
- Lansdown, R.V. Veronica Beccabunga. 2013. Available online: http://0-dx-doi-org.brum.beds.ac.uk/10.2305/IUCN.UK.2013-1.RLTS.T167923A13574152.en (accessed on 15 June 2017).
- Les, D.; Stuckey, R. The introduction and spread of Veronica beccabunga (Scrophulariaceae) in Eastern North America. J. N. Engl. Bot. Club 1985, 87, 503–513. [Google Scholar]
- Hilty, J. “Redtop”. Grasses, Sedges, and Non-Flowering Plants of Illinois. 2007. Available online: http://www.illinoiswildflowers.info/grasses/grass_index.htm (accessed on 12 August 2008).
- Long, H.C. Weeds of arable land. Maff Bull. 1938, 108. [Google Scholar]
- Morton Arboretum. White Willow. 2020. Available online: https://www.mortonarb.org/trees-plants/tree-plant-descriptions/white-willow (accessed on 4 September 2020).
- Monk, L.S.; Fagerstedt, K.V.; Crawford, R.M.M. Superoxide dismutase an in anaerobic polypeptide: A key factor in recovery from oxygen deprivation in Iris pseudacorus? Plant Physiol. 1987, 85, 1016–1020. [Google Scholar] [CrossRef] [Green Version]
- Schlüter, U.; Crawford, R.M. Long-term anoxia tolerance in leaves of Acorus calamus L. and Iris pseudacorus L. J. Exp. Bot. 2001, 52, 2213–2225. [Google Scholar] [CrossRef] [PubMed]
- Howard-Williams, C.; Davis, J.; Pickmere, S. The dynamics of growth, the effects of changing area and nitrate uptake by watercress Nasturtium officinale R. Br. in a New Zealand stream. J. Appl. Ecol. 1982, 19, 589–601. [Google Scholar] [CrossRef]
- Aerts, R.; De Caluwe, H. Nitrogen use efficiency of Carex species in relation to nitrogen supply. Ecology 1994, 75, 2362–2372. [Google Scholar] [CrossRef]
- Konings, H.; Verhoeven, J.T.A.; De Groot, R. Growth characteristics and seasonal allocation patterns of biomass and nutrients in Carex species growing in floating fens. Plant Soil 1992, 147, 183–196. [Google Scholar] [CrossRef]
- Akkermans, A.D.L. Root nodule symbioses in non-leguminous N2-fixing plants. In Interactions between Non-Pathogenic Soil Microorganisms and Plants; Dommergues, Y.R., Krupa, S.V., Eds.; Elsevier Scientific Publishing: New York, NY, USA, 1978; pp. 335–372. [Google Scholar]
- Van Dijk, C. Spore formation and endophyte diversity in root nodules of Alnus glutinosa (L.) Vill. New Phytol. 1978, 81, 601–615. [Google Scholar] [CrossRef]
- Hogsden, K.L.; Sager, E.P.S.; Hutchinson, T.C. The impacts of the non-native macrophyte Cabomba caroliniana on littoral biota of Kasshabog Lake, Ontario. J. Great Lakes Res. 2007, 33, 497–504. [Google Scholar] [CrossRef]
- ENSR International. Rapid Response Plan for Fanwort (Cabomba caroliniana) in Massachusetts; Massachusetts Department of Conservation and Recreation: Boston, MA, USA, 2005. [Google Scholar]
- Wilson, J.R.U.; Richardson, D.M.; Rouget, M.; Proches, S.; Amis, M.A.; Henderson, L.; Thuiller, W. Residence time and potential range: Crucial considerations in modelling plant invasions. Divers. Distrib. 2007, 13, 11–22. [Google Scholar] [CrossRef]
- Pullman, G.D.; Crawford, G. A Decade of Starry Stonewort in Michigan. Lake Line 2010, 13, 36–42. [Google Scholar]
- Catling, P.M.; Reznicek, A.A.; Brookes, B.S. The separation of Carex disticha and Carex sartwellii and the status of Carex disticha in North America. Can. J. Bot. 1988, 66, 2323–2330. [Google Scholar] [CrossRef]
- Catling, P.M.; Kostiuk, B. Carex acutiformis dominance of a cryptic invasive sedge at Ottawa. Bot. Electron. News 2003, 315, 1–6. [Google Scholar]
- Grant, M.S. Frogbit. 2013. Available online: http://www.miseagrant.umich.edu/explore/native-and-invasive-species/species/plants/frogbit/ (accessed on 16 April 2013).
- Robinson, M. European naiad: An invasive aquatic plant (Najas minor). In Massachusetts Department of Conservation and Recreation; Lakes and Ponds Program; Office of Water Resources: Boston, MA, USA, 2004; 4p. [Google Scholar]
- Indiana Department of Natural Resources (IN DNR). Aquatic Invasive Species: Water Chestnut; IN DNR: Indianapolis, IN, USA, 2012; 3p. [Google Scholar]
- Ontario’s Invading Species Awareness Program (OISAP). European Water Chestnut Trapa Natans. 2013. Available online: http://www.invadingspecies.com/invaders/plants-aquatic/european-water-chestnut/ (accessed on 2 May 2013).
- Lui, K.; Butler, M.; Allen, M.; Snyder, E.; da Silva, J.; Brownson, B.; Ecclestone, A. Field Guide to Aquatic Invasive Species: Identification, Collection and Reporting of Aquatic Invasive in Ontario Waters; Ontario Ministry of Natural Resources: Peterborough, ON, Canada, 2010; 201p. [Google Scholar]
- Great Lakes Commission (GLC). Lake St. Clair Coastal Habitat Assessment: With recommendations for Conservation and Restoration Planning; Great Lakes Commission: Ann Arbor, MI, USA, 2006. [Google Scholar]
- Natural Resources Conservation Service (NRCS). Buckthorn (Common and Glossy). In Pest Management-Invasive Plant Control; USDA–NRCS: Washington, DC, USA, 2007. [Google Scholar]
- State of Minnesota, Department of Natural Resources. Buckthorn: What You Should Know. What You Can Do; MN DNR: St. Paul, MN, USA, 2009; 4p. [Google Scholar]
- Crack Willows. 2013. Available online: http://invasives.glifwc.org/Salix_spp/control.html (accessed on 12 September 2013).
- Weber, E. Invasive Plant Species of the World: A Reference Guide to Environmental Weeds, 1st ed.; CAB Publishing: Wallingford, UK, 2003. [Google Scholar]
- Kao, W.-K.; Lin, B.-L. Phototropic leaf movements and photosynthetic performance in an amphibious fern, Marsilea quadrifolia. J. Plant Res. 2010, 123, 645–653. [Google Scholar] [CrossRef]
- Sakai, A.K.; Allendorf, F.W.; Holt, J.S.; Lodge, D.M.; Molofsky, J.; With, K.A.; Baughman, S.; Cabin, R.T.; Cohen, J.E.; Ellstrand, N.C.; et al. The Population Biology of Invasive Species. Ann. Rev. Ecol. Syst. 2001, 32, 305–332. [Google Scholar] [CrossRef] [Green Version]
- Mulligan, G.A.; Findlay, J.N. Reproductive systems and colonization in Canadian weeds. Can. J. Bot. 1970, 48, 859–860. [Google Scholar] [CrossRef]
- Honek, A.; Martinková, Z. Geographic variation in seed dormancy among populations of Echinochloa crus-galli. Oecologia 1996, 108, 419–423. [Google Scholar] [CrossRef]
- Bartomeus, I.; Vilà, M.; Steffan-Dewenter, I. Combined effects of Impatiens glandulifera invasion and landscape structure on native plant pollination. J. Ecol. 2010, 98, 440–450. [Google Scholar] [CrossRef] [Green Version]
- Vervoort, A.; Cawoy, V.; Jacuemart, A.-L. Comparative reproductive biology in co-occurring invasive and native Impatiens species. Int. J. Plant Sci. 2011, 172, 366–377. [Google Scholar] [CrossRef]
- Chittka, L.; Schürkens, S. Successful invasion of a floral market. Nature 2001, 411, 653. [Google Scholar] [CrossRef] [PubMed]
- Dieringer, G. The pollination ecology of Orchis spectabilis L. (Orchidaceae). Ohio J. Sci. 1982, 82, 218–225. [Google Scholar]
- Pons, T.L.; During, H.J. Biennial behavior of Cirsium palustre in ash coppice. Holarct. Ecol. 1987, 10, 40–44. [Google Scholar]
- Fraser, N. Cirsium palustre (marsh thistle). Literature search and habitat potential risk analysis. In Prepared for Ministry of Forests; Forest Practices Branch: Kamloops, BC, Canada, 2000. [Google Scholar]
- Agami, M.; Waisel, Y. The role of mallard ducks (Anas platyrhynchos) in distribution and germination of seeds of the submersed hydrophyte Najas marina. Oecologia 1986, 68, 473–475. [Google Scholar] [CrossRef] [PubMed]
- Malecki, R.A.; Blossey, B.; Hight, S.D.; Schroeder, D.; Kok, L.T.; Coulson, J.R. Biological control of purple loosestrife. BioScience 1993, 43, 680–686. [Google Scholar] [CrossRef]
- Reiné, R.; Chocarro, C.; Fillat, F. Soil seed bank and management regimes of semi-natural mountain meadow communities. Agric. Ecosyst. Env. 2004, 104, 567–575. [Google Scholar] [CrossRef]
- MISIN. 2013. Available online: http://www.misin.msu.edu/facts/detail.php?id=77 (accessed on 8 September 2019).
- Madsen, J.D.; Sutherland, J.W.; Bloomfield, J.A.; Eichler, L.W.; Boylen, C.W. The decline of native vegetation under dense Eurasian watermilfoil canopies. J. Aquat. Plant Manag. 1991, 29, 94–99. [Google Scholar]
- Idaho Invasive Species Council Technical Committee, The (IISCTC). Idaho Aquatic Nuisance Species Plant; IISCRC: Boise, ID, USA, 2007; 139p. [Google Scholar]
- Midwest Invasive Species Information Network (MISIN) and Michigan Natural Features Inventory (MNFI). Eurasian watermilfoil (Myriophyllum spicatum), Michigan State University. 2013. Available online: http://www.misin.msu.edu/facts/detail.php?id=44 (accessed on 25 April 2013).
- Illinois Department of Nautral Resources (IL DNR). Aquatic Invasive Species: Eurasian Watermilfoil. 2009. Available online: http://www.in.gov/dnr/files/EURASIAN_WATERMILFOIL.pdf (accessed on 23 April 2013).
- Haynes, R.R. Reproductive biology of selected aquatic plants. Ann. Bot. Gard 1988, 75, 805–810. [Google Scholar] [CrossRef]
- Buttery, B.R.; Lambert, J.M. Competition between Glyceria maxima and Phragmites communis in the region of Surlingham Broad: I. the competition mechanism. J. Ecol. 1965, 53, 163–181. [Google Scholar] [CrossRef]
- King County Noxious Weed Control Program. Reed Sweetgrass: Glyceria Maxima, Noxious Weeds, King County Department of Natural Resources and Parks. 2012. Available online: http://www.kingcounty.gov/environment/animalsAndPlants/noxious-weeds/weed-identification/reed-sweetgrass.aspx (accessed on 6 August 2012).
- Invasive Plant Atlas of New England (IPANE). Glyceria Maxima (Reed mannagrass, Reed sweetgrass), Catalog of Species, University of Connecticut. 2004. Available online: http://www.invasive.org/weedcd/pdfs/ipane/Glyceriamaxima.pdf (accessed on 20 August 2012).
- Wisconsin Department of Natural Resources (WI DNR). Curly-Leaf Pondweed (Potamogeton crispus). 2012. Available online: http://dnr.wi.gov/topic/invasives/fact/curlyleafpondweed.html (accessed on 29 April 2013).
- Nicholls, S.J.; Schloesser, D.W.; Geis, J.W. Seasonal growth of the exotic submersed macrophyte Nitellopsis obtusa in the Detroit River of the Great Lakes. Can. J. Bot. 1988, 66, 116–118. [Google Scholar] [CrossRef]
- Yamane, A.; Nishimura, H.; Mizutani, J. Allelopathy of yellow fieldcress (Rorippa sylvestris): Identification and characterization of phytotoxic constituents. J. Chem. Ecol. 1992, 18, 683–691. [Google Scholar] [CrossRef]
- D’Hertefeldt, T.; Falkengren-Grerup, U. Extensive physiological integration in Carex arenaria and Carex disticha in relation to potassium and water availability. New Phytol. 2002, 156, 469–477. [Google Scholar] [CrossRef]
- Learning Center of the American Southwest. Echinochloa Crus-Galli, Fact Sheets for Invasive Exotic Plants. 2012. Available online: http://www.southwestlearning.org/node/1479/39/2 (accessed on 4 April 2019).
- Open Learning Agency (OLA); British Columbia Ministry of Agriculture, Food, and Fisheries (MAFF). Guide to Weeds in British Columbia; Open Learning Agency: Burnaby, BC, Canada, 2002. [Google Scholar]
- Wisconsin Department of Natural Resources (WIDNR). Invasive Species. 2012. Available online: http://dnr.wi.gov/topic/Invasives/ (accessed on 4 September 2012).
- Francis, R.A.; Hoggart, S.P.G. The flora of urban river wallscapes. River Res. Appl. 2011, 8, 1–17. [Google Scholar] [CrossRef]
- Enescu, C.M.; Durrant, T.H.; Rigo, D.; Caudullo, G. Salix caprea in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., Rigo, D., Caudullo, G., Durrant, H.T., Mauri, A., Eds.; European Commission: Luxembourg, 2016. [Google Scholar]
- Missouri Botanical Garden. Juncus inflexus, Gardening Help. 2012. Available online: http://www.missouribotanicalgarden.org/gardens-gardening/your-garden/plant-finder/plant-details/kc/u310/juncus-inflexus-afro.aspx (accessed on 26 July 2012).
- Benson, A.J.; Richerson, M.M. Nonindigenous aquatic species of the northeastern United States. Aquat. Invaders 2004, 15, 1–9. [Google Scholar]
- Illinois Department of Natural Resources. Illinois Exotic Species: Waterclover Marsilea Quadrifolia. 2013. Available online: http://www.dnr.state.il.us/education/exoticspecies/waterclover.htm (accessed on 8 April 2013).
- Andersson, B. Macrophyte development and habitat characteristics in Sweden’s large lakes. Ambio 2001, 30, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Forest Health Staff. Reed Mannagrass: Glyceria Maxima (Hartman) Holmb; Weed of the Week; U.S. Department of Agriculture Forest Service: Newtown, PA, USA, 2006; 1p. [Google Scholar]
- Boos, T.; Kearns, K.; LeClair, C.; Panke, B.; Scriver, B.; Williams, B. A Field Guide to Terrestrial Invasive Plants in Wisconsin; Wisconsin Department of Natural Resources: Madison, WI, USA, 2010. [Google Scholar]
- Perrins, J.C.; Herwig, R.P. Ozone treatment of seawater mesocosm experiments: Bacterial community enumeration with flow cytometry. In Proceedings of the Third International Conference on Marine Bioinvasions, La Jolla, CA, USA, 16–19 March 2003; p. 105. [Google Scholar]
- Eckel, P.M. Two problems in Betulaceae along the Niagara River: Alnus glutinosa and Betula cordifolia. Clintonia 2003, 18, 3–4. [Google Scholar]
- NatureServe. NatureServe Explorer: An Online Encyclopedia of Life [web Application], Version 7.1. 2010. Available online: http://www.natureserve.org/explorer (accessed on 13 June 2012).
- Mehrhoff, L.J.; Silander, J.A., Jr.; Leicht, S.A.; Mosher, E.S.; Tabak, N.M. IPANE: Invasive Plant Atlas of New England. Department of Ecology and Evolutionary Biology; University of Connecticut: Storrs, CT, USA, 2003; Available online: http://www.ipane.org (accessed on 14 April 2017).
- Aiken, S.G.; Newroth, P.R.; Wile, I. The biology of Canadian weeds. 34. Myriophyllum spicatum L. Can. J. Plant Sci. 1979, 59, 201–215. [Google Scholar] [CrossRef]
- Shamsi, S.R.A.; Whitehead, F.H. Comparative eco-physiology of Epilobium hirsutum L. and Lythrum salicaria L.: II. Growth and development in relation to light. J. Ecol. 1974, 62, 631–645. [Google Scholar] [CrossRef]
- Grime, J.P.; Hodgson, J.G.; Hunt, R. Comparative Plant Ecology: A Functional Approach to Common British Species; Chapman & Hall: London, UK, 1988. [Google Scholar]
- Swearingen, J.; Saltonstall, K. Phragmites Field Guide: Distinguishing Native and Exotic Forms of Common Reed (Phragmites australis) in the United States. Available online: https://www.nrcs.usda.gov/Internet/FSE_PLANTMATERIALS/publications/idpmctn11494.pdf (accessed on 15 January 2021).
- Rudrappa, T.; Choi, Y.S.; Levia, D.F.; Legates, D.R.; Lee, K.H.; Bais, H.P. Phragmites australis root secreted phytotoxin undergoes photo-degradation to execute severe phytotoxicity. Plant Signal. Behav. 2009, 4, 506–513. [Google Scholar] [CrossRef] [Green Version]
- Flora of North America. 2008. Available online: www.eFloras.org (accessed on 8 September 2019).
- O’Neill, C.R., Jr. European Frog-Bit (Hydrocharis Morsus-Ranae)- Floating Invader of Great Lakes Basin Waters; New York Sea Grant: New York, NY, USA, 2007; 4p. [Google Scholar]
- Online Atlas of British & Irish Flora. Juncus compressus (Round-fruited Rush). 2012. Available online: http://www.brc.ac.uk/plantatlas/index.php?q=plant/juncus-compressus (accessed on 23 July 2012).
- Midwest Invasives Species Information Network (MISIN) and Michigan Natural Features Inventory (MNFI). Gingermint (Mentha x gracilis). 2013. Available online: http://www.misin.msu.edu/facts/detail.php?id=96 (accessed on 6 May 2013).
- USDA NRCS Northeast Plant Materials Program. Plant Fact Sheet- Agrostis gigantea Roth. 2002. Available online: http://plants.usda.gov/factsheet/pdf/fs_aggi2.pdf (accessed on 1 November 2018).
- Kennay, J.; Fell, G. Vegetation Management Guideline: Moneywort (Lysimachia nummularia); Illinois Natural History Survey; Prairie Research Institute: Champaign, IL, USA, 2011. [Google Scholar]
- Waggy, M.A. Solanum dulcamara. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory: Ft. Collins, CO, USA, 2009. [Google Scholar]
- Kollmann, J.; Bañuelos, M.J. Latitudinal trends in growth and phenology of the invasive alien plant Impatiens glandulifera (Balsaminaceace). Divers. Distrib. 2004, 10, 377–385. [Google Scholar] [CrossRef]
- Mijnsbrugge, K.V.; Turcsán, A.; Michiels, B. Population differentiation and phenotypic plasticity in temperature response of bud burst in Frangula alnus provenances of different latitude. Plant Syst. Evol. 2016, 302, 257–264. [Google Scholar] [CrossRef]
- Clough, J.M.; Teeri, J.A.; Alberte, R.S. Photosynthetic adaptation of Solanum dulcamara L. to sun and shade environments. I. A comparison of sun and shade populations. Oecologia 1979, 38, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Flynn, D.F.B.; Sudderth, E.A.; Bazzaz, F.A. Effects of aphid herbivory on biomass and leaf-level physiology of Solanum dulcamara under elevated temperature and CO2. Env. Exp. Bot. 2006, 56, 10–18. [Google Scholar] [CrossRef]
- Gauhl, E. Photosynthetic response to varying light intensity in ecotypes of Solanum dulcamara L. from shaded and exposed habitats. Oecologia 1976, 22, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Horvath, I.; Bernath, J.; Tetenyi, P. Effect of the spectral composition of light on dry matter production in Solanum dulcamara ecotypes of different orgin. Acta Agron. Hung. 1977, 26, 346–354. [Google Scholar]
- Chambers, P.A. Light and nutrients in the control of aquatic plant community structure. II. In situ observations. J. Ecol. 1987, 75, 621–628. [Google Scholar] [CrossRef]
- Dailey, K.R.; Welch, K.A.; Lyons, W.B. Evaluating the influence of road salt on water quality of Ohio Rivers over time. Appl. Geochem. 2014, 47, 25–35. [Google Scholar] [CrossRef]
- Foos, A. Spatial distribution of road salt contamination of natural springs and seeps, Cuyahoga Falls, Ohio, USA. Environ. Geol. 2003, 44, 14–19. [Google Scholar] [CrossRef]
- Findlay, S.E.G.; Kelly, V.R. Emerging indirect and long-term road salt effects on ecosystems. Ann. Acad. Sci. 2011, 1223, 58–68. [Google Scholar] [CrossRef]
- Lower, E.; Sturtevant, R.; Iott, S.; Martinez, F.; Rutherford, E.; Mason, D.; Elgin, A. The Great Lakes’ Most Unwanted: Characterizing the Impacts and Traits of the Top Great Lakes Invasive Species. Aquat. Invasions 2020, submitted. [Google Scholar]
- Ontario’s Invading Species Awareness Program (OISAP). Eurasian Water-Milfoil: Myriophyllum spicatum. 2013. Available online: http://www.invadingspecies.com/invaders/plants-aquatic/eurasian-water-milfoil/ (accessed on 25 April 2013).
- IPANE. Invasive Plant Atlas of New England (IPANE) at the University of Connecticut Online Database. 2013. Available online: http://invasives.eeb.uconn.edu/ipane/ (accessed on 1 January 2018).
- Keast, A. The introduced aquatic macrophyte, Myriophyllum spicatum, as habitat for fish and their macroinvertebrate prey. Can. J. Zool. 1984, 62, 1289–1303. [Google Scholar] [CrossRef]
- Jacobs, J.; Mangold, J. Plant Guide for Eurasian Watermilfoil (Myriophyllum spicatum L.); USDA-Natural Resources Conservation Service, Montana State Office: Bozeman, MT, USA, 2009; 3p. [Google Scholar]
- Bowen, D. Eurasian watermilfoil: Factsheet, Minnesota Sea Grant. 2010. Available online: http://www.seagrant.umn.edu/exotics/eurasian.html (accessed on 23 April 2013).
- Waggy, M.A. Phalaris arundinacea in Fire Effects Information System; United States Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory: Ft. Collins, CO, USA, 2010. [Google Scholar]
- Huffman, M.R.; Annala, A.E.; Lattimore, P.T.; Kapustka, L.A. Prairie restorations in southwestern Ohio: Vegetation and soil characteristics after ten years. In The prairie: Past, Present, and Future, Proceedings of the 9th North American Prairie Conference 1984; Moorhead, MN, USA, 29 July–1 August 1984, Clambey, G.K., Pemble, R.H., Eds.; Tri-College University Center for Environmental Studies; North Dakota State University: Fargo, ND, USA, 1986; pp. 185–189. [Google Scholar]
- Reuter, D.D. Sedge meadows of the Upper Midwest: A stewardship summary. Nat. Areas J. 1986, 6, 27–34. [Google Scholar]
- Howe, H.F. Response of Zizia aurea to seasonal mowing and fire in a restored prairie. Am. Mid. Nat. 1999, 141, 373–380. [Google Scholar] [CrossRef]
- Maurer, D.A.; Lindig-Cisneros, R.; Werner, K.J.; Kercher, S.; Miller, R.; Zedler, J.B. The replacement of wetland vegetation by reed canarygrass (Phalaris arundinacea). Ecol. Restor. 2003, 21, 116–119. [Google Scholar] [CrossRef]
- Czarapata, E.J. Invasive Plants of the Upper Midwest: An Illustrated Guide to Their Identification and Control; The University of Wisconsin Press: Madison, WI, USA, 2005. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Nonindigenous Plant Species | Growth Form | Earliest Date | Inhabited HUC8s |
|---|---|---|---|---|
| Amaranthaceae | Chenopodium glaucum | Emergent | 1865 | 38% |
| Apiaceae | Conium maculatum * | Emergent | 1843 | 33% |
| Asteraceae | Cirsium palustre * | Emergent | 1934 | 36% |
| Pluchea odorata | Emergent | 1912 | 3% | |
| Solidago sempervirens | Emergent | 1969 | 6% | |
| Balsaminaceae | Impatiens glandulifera | Emergent | 1912 | 10% |
| Betulaceae | Alnus glutinosa * | Emergent (tree) | 1913 | 15% |
| Boraginaceae | Myosotis scorpiodes | Emergent | 1886 | 50% |
| Brassicaceae | Nasturtium officinale | Floating/Prostrate | 1847 | 37% |
| Rorippa sylvestris | Emergent | 1884 | 25% | |
| Butomaceae | Butomus umbellatus * | Emergent | 1905 | 43% |
| Cabombaceae | Cabomba caroliniana * | Submerged w/Emergent flowers | 1935 | 12% |
| Caryophyllaceae | Myosoton aquaticum | Emergent | 1894 | 10% |
| Characeae | Nitellopsis obtuse * | Submerged | 1981 | 36% |
| Cyperaceae | Carex acutiformis * | Emergent | 1951 | 3% |
| Carex disticha | Emergent | 1972 | 1% | |
| Fabaceae | Lupinus polyphyllus | Emergent | 1959 | 14% |
| Haloragaceae | Myriophyllum spicatum * | Submerged | 1880 | 87% |
| Hydrocharitaceae | Hydrocharis morsus-ranae * | Floating | 1972 | 27% |
| Najas marina | Submerged | 1864 | 12% | |
| Najas minor * | Submerged | 1932 | 28% | |
| Iridaceae | Iris pseudacorus ** | Emergent | 1886 | 56% |
| Juncaceae | Juncus compressus * | Emergent | 1895 | 18% |
| Juncus inflexus * | Emergent | 1922 | 5% | |
| Lamiaceae | Lycopus asper | Emergent | 1892 | 13% |
| Lycopus europaeus | Emergent | 1903 | 9% | |
| Mentha aquatica | Emergent | 1843 | 25% | |
| Mentha spicata | Emergent | 1843 | 26% | |
| Mentha × gracilis | Emergent | 1896 | 10% | |
| Lythraceae | Lythrum salicaria ** | Emergent | 1839 | 91% |
| Trapa natans* | Floating | 1949 | 9% | |
| Marsileaceae | Marsilea quadrifolia | Floating/Emergent | 1893 | 6% |
| Menyanthaceae | Nymphoides peltata * | Floating | 1930 | 12% |
| Onagraceae | Epilobium hirsutum * | Emergent | 1874 | 34% |
| Plantaginaceae | Veronica beccabunga | Emergent | 1849 | 10% |
| Poaceae | Agrostis gigantea | Emergent | 1838 | 70% |
| Alopecurus geniculatus | Emergent | 1882 | 9% | |
| Echinochloa crus-galli | Emergent | 1838 | 51% | |
| Glyceria maxima * | Emergent | 1979 | 8% | |
| Phragmites australis ssp. Australis ** | Emergent | 1869 | 33% | |
| Poa trivialis | Emergent | 1843 | 12% | |
| Puccinellia distans | Emergent | 1893 | 13% | |
| Polygonaceae | Persicaria maculosa | Emergent | 1838 | 41% |
| Rumex longifolius | Emergent | 1901 | 6% | |
| Rumex obtusifolius | Emergent | 1837 | 36% | |
| Potamogetonaceae | Potamogeton crispus * | Submerged | 1879 | 75% |
| Primulaceae | Lysimachia nummularia * | Emergent | 1882 | 38% |
| Lysimachia vulgaris * | Emergent | 1912 | 11% | |
| Rhamnaceae | Frangula alnus ** | Emergent (tree) | 1913 | 56% |
| Salicaceae | Salix alba | Emergent (tree) | 1886 | 23% |
| Salix caprea | Emergent (tree) | 1985 | 1% | |
| Salix fragilis | Emergent (tree) | 1886 | 25% | |
| Salix purpurea | Emergent (tree) | 1880 | 13% | |
| Solanaceae | Solanum dulcamara | Emergent | 1843 | 66% |
| Typhaceae | Typha angustifolia ** | Emergent | 1877 | 82% |
| Growth Form | Nonindigenous Plant Species | Impacted Natives |
|---|---|---|
| Submerged | Myriophyllum spicatum * | M. sibiricum+ |
| Floating | Trapa natans * | Native (emergent) grasses |
| Emergent | Chenopodium glaucum | C. album, C. berlandieri, C. capitatum +, C. foggii +, C. humile, C. leptophyllum +. C. overi, C. pallescens, C. pratericola, C. rubrum, C. salinum, C. simplex, C. standleyanum, C. subglabrum + |
| Emergent | Conium maculatum * | Grasses and forbs |
| Emergent | Cirsium palustre * | C. pitcheri+, C. muticum Carex spp. |
| Emergent | Pluchea odorata | High marsh community ^ |
| Emergent | Butomus umbellatus * | Phalaris arundinacea, Phragmites australis ssp. americanus, Non-specific natives |
| Emergent | Carex acutiformis * | Understory natives ^ |
| Emergent | Iris pseudacorus ** | Typha spp., Peltandra virginica, I. brevicaulis +, I. cristata +, I. lacustris +, I. robusta [versicolor × virginica] +, I. setosa, I. verna +, I. versicolor, I. virginica var. shrevei +, sedges, rushes |
| Emergent | Juncus compressus * | Juncus alpinus +, J. ambiguous +, J. balticus, J. biflorus +, J. marginatus, J. brachycarpus, J. brachycephalus +, J. dichotomus +, J. diffusissimus +, J. ensifolius +, J. greenei +, J. interior +, J. militaris +, J. pelocarpus +, J. scirpoides +, J. secundus +, J. stygius +, J. subcaudatus +, J. vaseyi +, non-rushes |
| Emergent | Juncus inflexus * | J. alpinus +, J. ambiguous +, J. balticus, J. biflorus +, J. marginatus, J. brachycarpus, J. brachycephalus +, J. dichotomus +, J. diffusissimus +, J. ensifolius +, J. greenei +, J. interior +, J. militaris +, J. pelocarpus +, J. scirpoides +, J. secundus +, J. stygius +, J. subcaudatus +, J. vaseyi + |
| Emergent | Lycopus asper | L. americanus, L. amplectens +, L. rubellus +, L. uniflorus, L. virginicus + |
| Emergent | Lycopus europaeus | L. americanus, L. amplectens +, L. rubellus +, L. uniflorus, L. virginicus + |
| Emergent | Lythrum salicaria ** | Grasses, sedges, flowering plants |
| Emergent | Epilobium hirsutum * | E. angustifolium +, E. strictum + |
| Emergent | Echinochloa crus-galli | E. muricata, E. walteri +, |
| Emergent | Phragmites australis ssp. Australis ** | Sedges, rushes, cattails |
| Emergent | Lysimachia nummularia * | L. radicans + |
| Emergent | Lysimachia vulgaris * | L. radicans + |
| Emergent | Typha angustifolia ** | T. latifolia, Campanula aparinoides, Cicuta bulbifera, Galium tinctorium |
| Emergent (tree) | Frangula alnus ** | Rhamnus alnifolia +, Rhamnus lanceolata +, Ilex verticillata +, trees, shrubs and wildflowers |
| Emergent (tree) | Salix purpurea | Native willows |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sturtevant, R.; Lower, E.; Bartos, A.; Elgin, A. A Review and Secondary Analysis of Competition-Related Impacts of Nonindigenous Aquatic Plants in the Laurentian Great Lakes. Plants 2021, 10, 406. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020406
Sturtevant R, Lower E, Bartos A, Elgin A. A Review and Secondary Analysis of Competition-Related Impacts of Nonindigenous Aquatic Plants in the Laurentian Great Lakes. Plants. 2021; 10(2):406. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020406
Chicago/Turabian StyleSturtevant, Rochelle, El Lower, Austin Bartos, and Ashley Elgin. 2021. "A Review and Secondary Analysis of Competition-Related Impacts of Nonindigenous Aquatic Plants in the Laurentian Great Lakes" Plants 10, no. 2: 406. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020406