
Establishment of a Cell Suspension Culture of Eysenhardtia platycarpa: Phytochemical Screening of Extracts and Evaluation of Antifungal Activity

 , , and
, , and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Obtaining Plantlets and Callus Induction
2.2. Cell Suspension Cultures
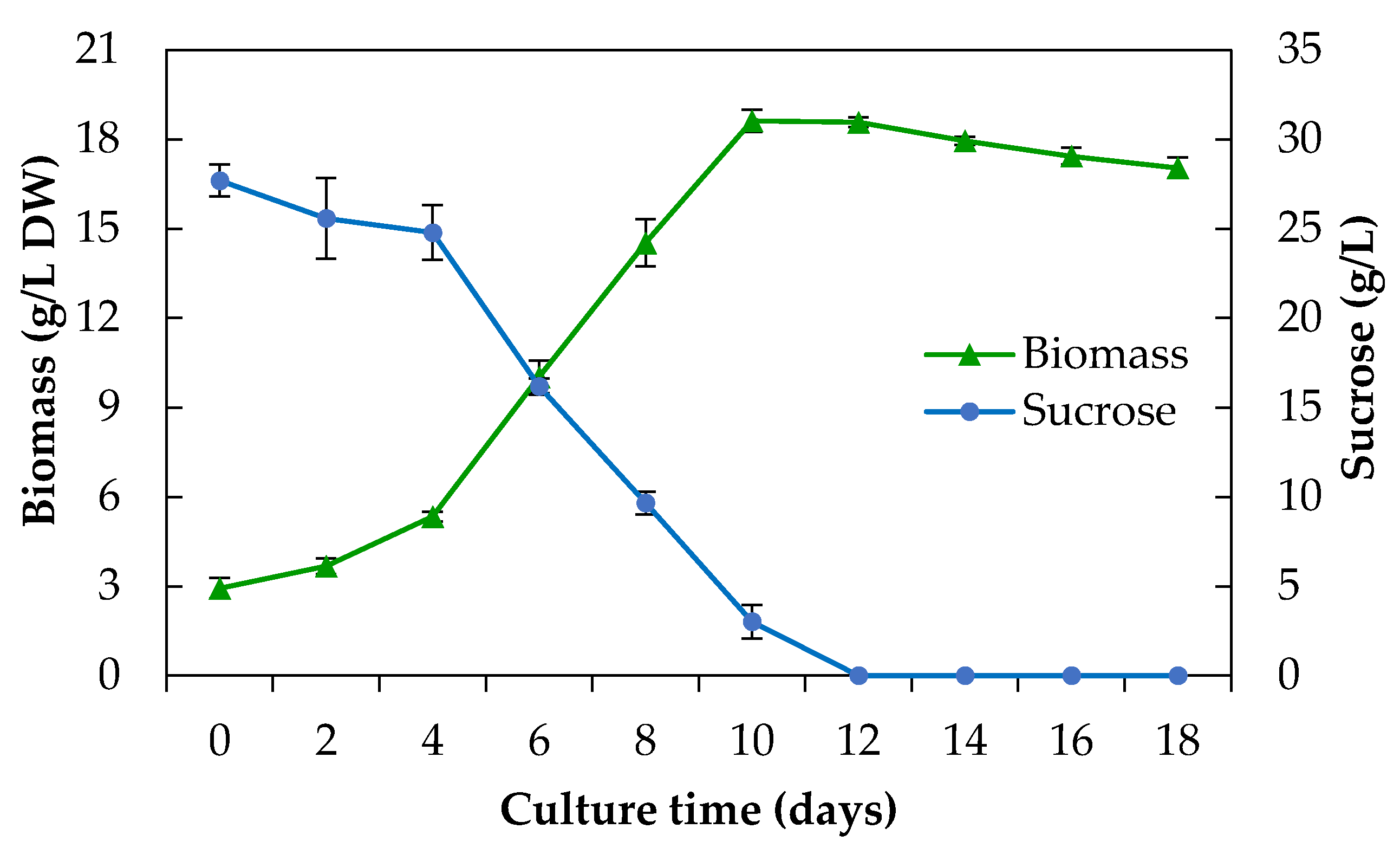
2.2.1. Growth Kinetics and Sucrose Consumption
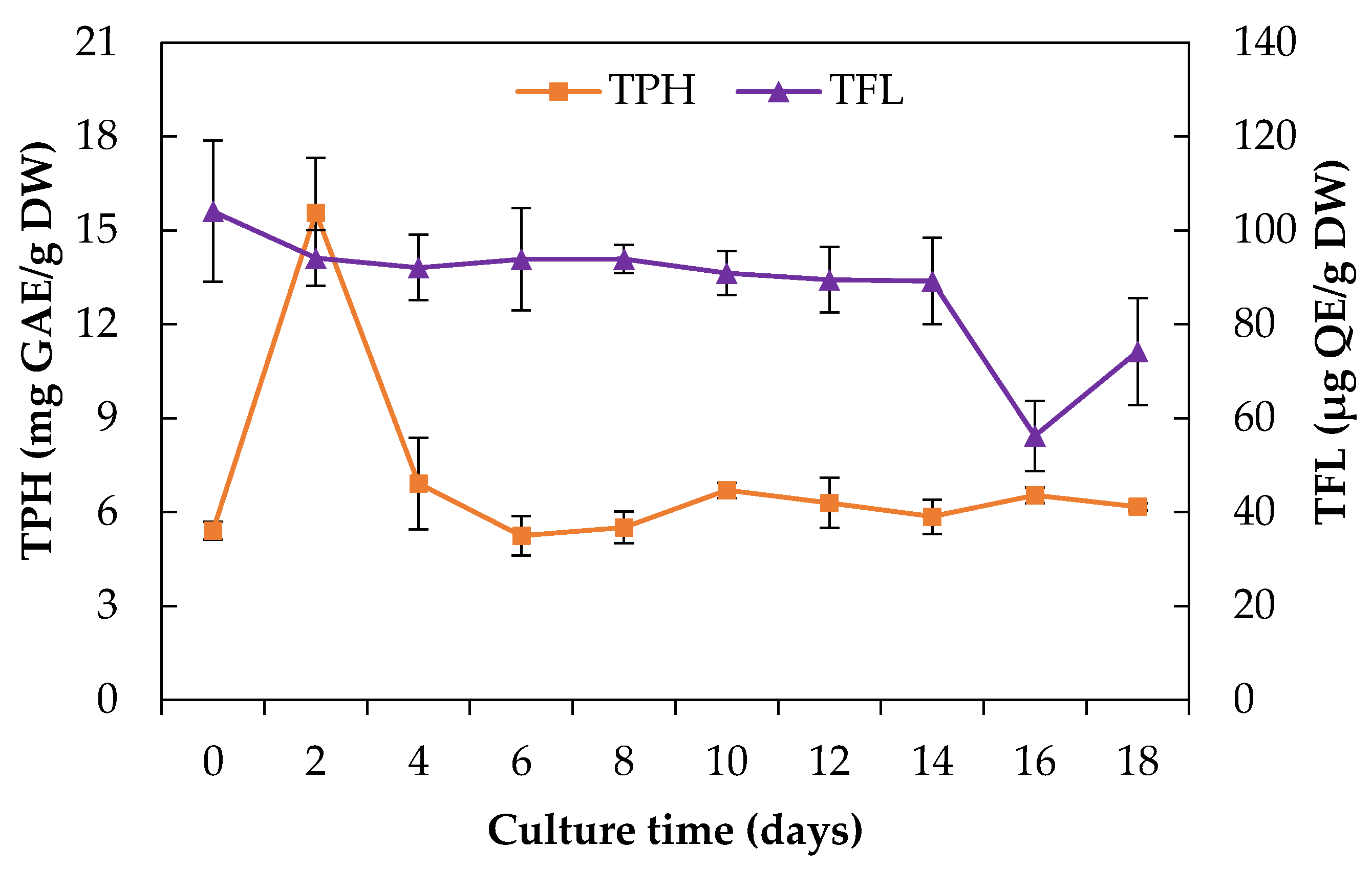
2.2.2. Total Phenolics and Flavonoids Content
2.3. Yield of Extracts of Cell Suspension Culture and Intact Plant
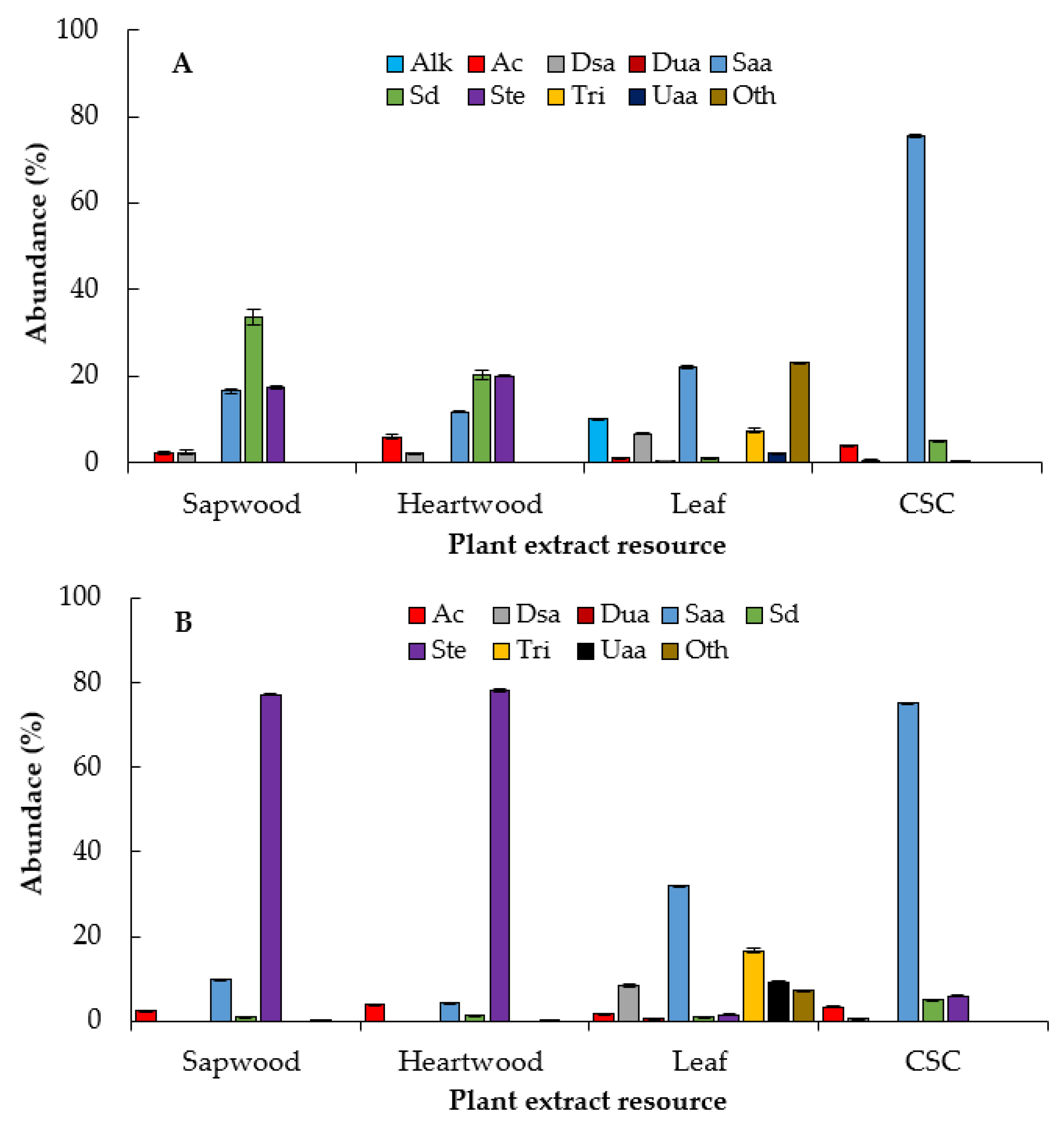
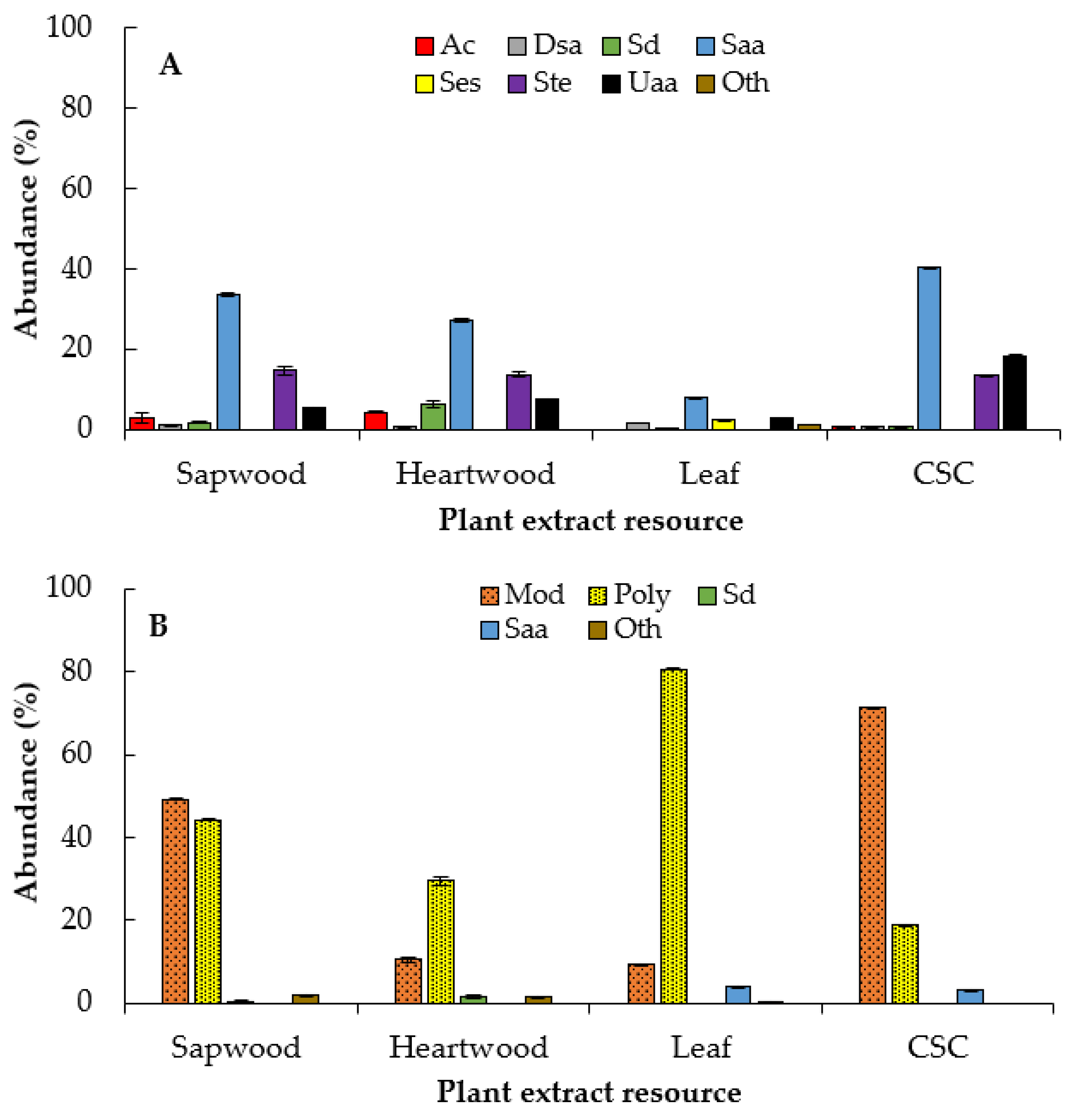
2.4. Compounds Identified by GC‒MS
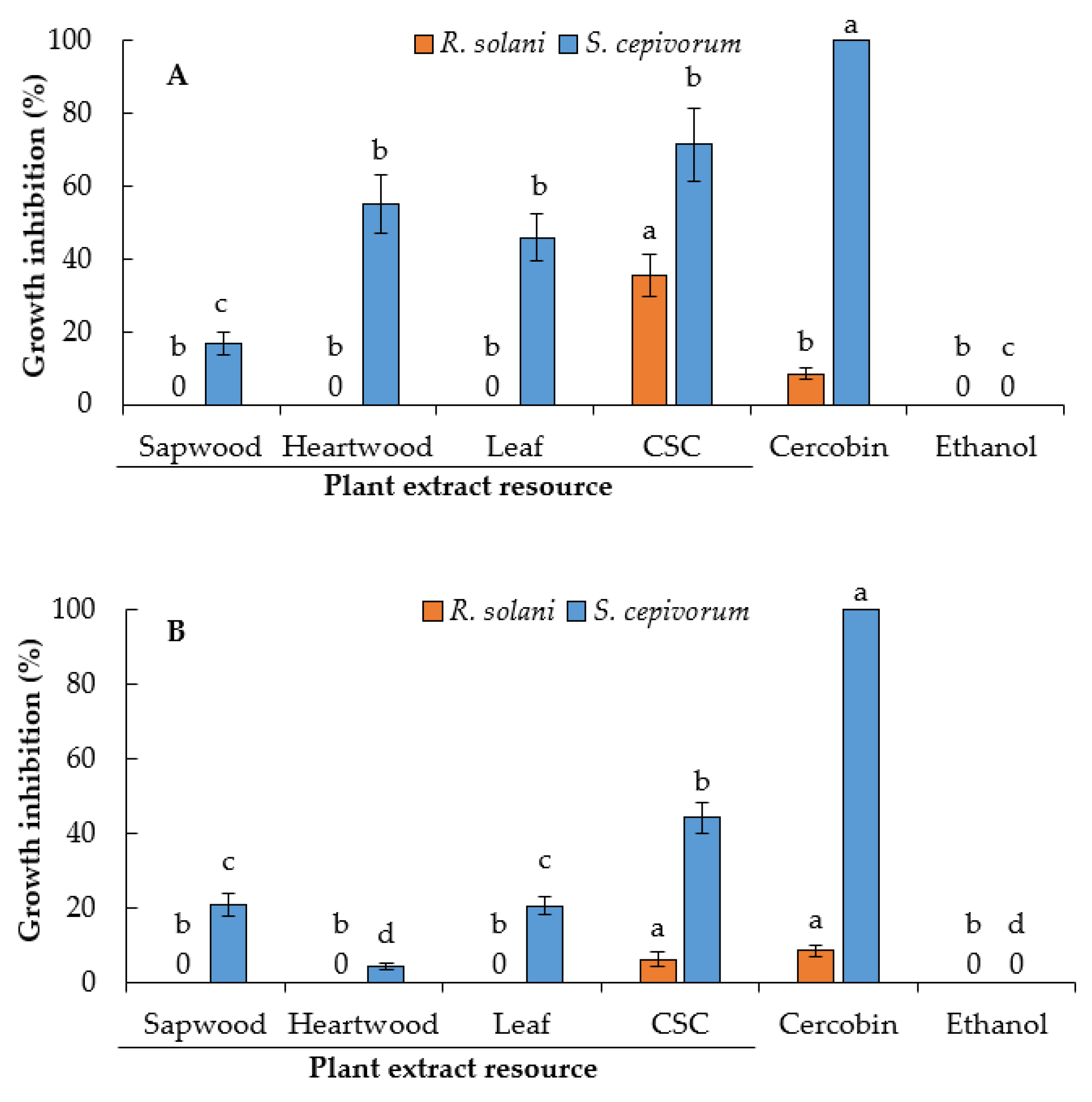
2.5. Antifungal Activity
3. Materials and Methods
3.1. Collection of Plant Material
3.2. Obtaining Plantlets and Incubation Conditions
3.3. Establishment of Callus Cultures
3.4. Cell Suspension Cultures
3.4.1. Growth Kinetics
3.4.2. Sucrose Consumption Determination
3.4.3. Determination of the Total Phenolics and Flavonoids Content
3.5. Phytochemical Analysis of Cell Suspension Cultures and Intact Plants
3.5.1. Extraction and Sample Preparation
3.5.2. Analysis of Extracts by Gas Chromatography‒Mass Spectrometry (GC‒MS)
3.6. In Vitro Antifungal Evaluation of Extracts
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Silva, N.C.C.; Fernandes Júnior, A. Biological properties of medicinal plants: A review of their antimicrobial activity. J. Venom. Anim. Toxins 2010, 16, 402–413. [Google Scholar] [CrossRef]
- Borges, D.F.; Lopes, E.A.; Moraes, F.A.R.; Soares, M.S.; Visôtto, L.E.; Oliveira, C.R.; Valente, V.M.M. Formulation of botanicals for the control of plant-pathogens: A review. Crop Prot. 2018, 110, 135–140. [Google Scholar] [CrossRef]
- Mulabagal, V.; Hsin-Sheng, T. Plant cell cultures—An alternative and efficient source for the production of biologically important secondary metabolites. Int. J. Appl. Sci. Eng. 2004, 2, 29–48. [Google Scholar]
- McCoy, E.; O’Connor, S.E. Natural products from plant cell cultures. Prog. Drug Res. 2008, 65, 329–370. [Google Scholar] [PubMed]
- Smetanska, I. Production of secondary metabolites using plant cell cultures. Adv. Biochem. Eng. Biotechnol. 2008, 111, 187–228. [Google Scholar]
- Haida, Z.; Nakasha, J.J.; Hakiman, M. In vitro responses of plant growth factors on growth, yield, phenolics content and antioxidant activities of Clinacanthus nutans (Sabah Snake Grass). Plants 2020, 9, 1030. [Google Scholar] [CrossRef]
- Rzedowsky, J.; Equihua, M. Atlas Cultural de México; De Planeta: Ciudad de México, México, 1987. [Google Scholar]
- Martínez, M. Las Plantas Medicinales de México; Editorial Botas: Ciudad de México, México, 1996. [Google Scholar]
- Villavicencio, N.M.A.; Pérez, E.B.E.; Ramírez, A.A. Plantas útiles del Estado de Hidalgo II; Universidad Autónoma del Estado de Hidalgo: Pachuca, Hidalgo, 2002. [Google Scholar]
- Garcia-Campoy, A.; Garcia, E.; Muñiz-Ramirez, A. Phytochemical and Pharmacological Study of the Eysenhardtia Genus. Plants 2020, 9, 1124. [Google Scholar] [CrossRef]
- Narváez-Mastache, J.M.; Garduño-Ramírez, M.L.; Álvarez, L.; Delgado, G. Antihyperglycemic activity and chemical constituents of Eysenhardtia platycarpa. J. Nat. Prod. 2006, 69, 1687–1691. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Villegas, V.; Clares-Naveros, B.; García-López, M.L.; Calpena-Campmany, A.C.; Bustos-Zagal, P.; Garduño-Ramírez, M.L. Development and characterization of two nano-structured systems for topical application of flavanones isolated from Eysenhardtia platycarpa. Colloids Surf. B 2014, 116, 183–192. [Google Scholar] [CrossRef]
- Bernabé-Antonio, A.; Maldonado-Magaña, A.; Ramírez-López, C.B.; Salcedo-Pérez, E.; Meza-Contreras, J.C.; González-García, Y.; López-Dellamary Toral, F.A.; Cruz-Sosa, F. Establishment of callus and cell suspension cultures of Eysenhardtia polystachya (Ortega) and fungistatic activity of their extracts. S. Afr. J. Bot. 2017, 112, 40–47. [Google Scholar] [CrossRef]
- Trejo-Espino, J.L.; Rodríguez-Monroy, M.; Vernon-Carter, E.J.; Cruz-Sosa, F. Establishment and characterization of Prosopis laevigata (Humb. & Bonpl. ex Willd) M.C. Johnst. cell suspension culture: A biotechnology approach for mesquite gum production. Acta Physiol. Plant 2011, 33, 1687–1695. [Google Scholar]
- Maldonado-Magaña, A.; Orozco-Villafuerte, J.; Buendía-González, L.; Estrada-Zúñiga, M.E.; Bernabé-Antonio, A.; Cruz-Sosa, F. Establishment of cell suspension cultures of Prosopis laevigata (Humb. & Bonpl. Ex Willd) M.C. Johnst to determine the effect of zinc on the uptake and accumulation of lead. Rev. Mex. Ing. Quím. 2013, 12, 489–498. [Google Scholar]
- Su, Y.H.; Liu, Y.B.; Zhang, X.S. Auxin-cytokinin interaction regulates meristem development. Mol. Plant 2011, 4, 616–625. [Google Scholar] [CrossRef]
- Pal, A.K.; Acharya, K.; Ahuja, P.S. Endogenous auxin level is a critical determinant for in vitro adventitious shoot regeneration in potato (Solanum tuberosum L.). J. Plant Biochem. Biot. 2012, 21, 205–212. [Google Scholar] [CrossRef]
- Machakova, I.; Zazimolova, E.; George, E.F. Plant growth regulators I: Introduction, auxins, their analogues and inhibitors. In Plant Propagation by Tissue Culture; George, E.F., Hall, M.A., De Klerk, G.J., Eds.; Springer: Dordrecht, The Netherlands, 2008; p. 175204. [Google Scholar]
- George, E.F. Plant Tissue Culture Procedure—Background. In Plant Propagation by Tissue Culture; George, E.F., Hall, M.A., De Klerk, G.J., Eds.; Springer: Dordrecht, The Netherlands, 2008; p. 128. [Google Scholar]
- Carmona, R.L.; Rojas, L.M.; Aura, I.U.T.; Atehortúa, G.L. Effect of nutrients starvation in the cell cycle synchronization of cell suspension cultures of Jatropha curcas L. Curr. Plant Biol. 2018, 13, 23–31. [Google Scholar]
- Hussein, S.; Halmi, M.I.E.; Ling, A.P.K. Modelling the growth kinetics of callus cultures from the seedling of Jatropha curcas L. according to the modified Gompertz model. J. Microbiol. Biotechn. 2016, 4, 20–23. [Google Scholar]
- Paine, C.E.T.; Marthews, T.R.; Vogt, D.R.; Purves, D.; Rees, M.; Hector, A.; Turnbull, L.A. How to fit nonlinear plant growth models and calculate growth rates: And update for ecologists. Methods Ecol. Evol. 2012, 3, 245–256. [Google Scholar] [CrossRef]
- Cisneros-Torres, D.; Cruz-Sosa, F.; Nicasio-Torres, M.P.; González-Cortazar, M.; Romero-Estrada, A.; Bernabé-Antonio, A. Establishment of a cell suspension culture from Calophyllum brasiliense and evaluation of its antioxidant and anti-inflammatory activity. Rev. Mex. Ing. Quím. 2020, 19, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Mohamad Puad, N.I.; Abdullah, T.A. Monitoring the growth of plant cells in suspension culture. In Multifaceted Protocol in Biotechnology; Amid, A., Sulaiman, S., Jimat, D.N., Azmin, N.F., Eds.; Springer: Singapore, 2018; pp. 203–214. [Google Scholar]
- Benjamins, R.; Scheres, B. Auxin: The looping star in plant development. Annu. Rev. Plant Biol. 2008, 59, 443–465. [Google Scholar] [CrossRef]
- Ammar, M.I.; Nenaah, G.E.; Mohamed, A.H.H. Antifungal activity of prenylated flavonoids isolated from Tephrosia apollinea L. against four phytopathogenic fungi. Crop Prot. 2013, 49, 21–25. [Google Scholar] [CrossRef]
- Wang, K.; Jiang, S.; Pu, T.; Fan, L.; Su, F.; Ye, M. Antifungal activity of phenolic monoterpenes and structure related compounds against plant pathogenic fungi. Nat. Prod Res. 2019, 33, 1423–1430. [Google Scholar] [CrossRef] [PubMed]
- Giri, L.; Dhyani, P.; Rawat, S.; Bhatt, I.D.; Nandi, S.K.; Rawal, R.S.; Pande, V. In vitro production of phenolic compounds and antioxidant activity in callus suspension cultures of Habenaria edgeworthii: A rare Himalayan medicinal orchid. Ind. Crops Prod. 2012, 39, 1–6. [Google Scholar] [CrossRef]
- Liu, C.Z.; Saxena, P.K. Saussurea medusa cell suspension cultures for flavonoid production. In Protocols for In Vitro Cultures and Secondary Metabolite Analysis of Aromatic and Medicinal Plants; Jain, S.M., Saxena, P.K., Eds.; Springer: Dordrecht, The Netherlands, 2009; p. 5359. [Google Scholar]
- Torres-Andrade, P.A.; Lomelí-Ramírez, M.G.; López-Dellamary, F.; Fuentes-Talavera, F.J.; Richter, H.G.; Silva-Guzmán, J.A. Natural decay resistance of Eysenhardtia polystachya (Ortega) Sarg. Int. Wood Prod. J. 2010, 1, 81–84. [Google Scholar] [CrossRef]
- Ypushima, P.A.L. Caracterización Biológica y Química de la Madera de teca (Tectona grandis) en dos Condiciones Edafoclimáticas y su Relación con el Crecimiento. Master’s Thesis, University de Guadalajara, Guadalajara, Jalisco, Mexico, 2015. [Google Scholar]
- Santiago, M.G. Actividad Antifúngica de Extractos de las Maderas de Caesalpinia platyloba, Lysiloma latisquum, Metopium brownei y Piscidia piscipula. Master’s Thesis, Universidad de Guadalajara, Guadalajara, Jalisco, Mexico, 2017. [Google Scholar]
- Dieu-Hien, T.; Dinh, H.N.; Nhat, T.A.T.; Anh, V.B.; Tuong, H.D.; Hoang, C.N. Evaluation of the use of different solvents for phytochemical constituents, antioxidants, and in vitro anti-inflammatory activities of Severinia buxifolia. J. Food Qua. 2019, 2019, 1–9. [Google Scholar]
- Bukke, A.N.; Hadi, F.N.; Produtur, C.S. Comparative study of in vitro antibacterial activity of leaves, bark, heart wood and seed extracts of Caesalpinia sappan L. Asian Pac. J. Trop. Dis. 2015, 5, 903–907. [Google Scholar] [CrossRef]
- Do, Q.D.; Angkawijaya, A.E.; Tran-Nguyen, P.L.; Huynh, L.H.; Soetaredjo, F.E.; Ismadji, S.; Ju, Y.H. Effect of extraction solvent on total phenol content, total flavonoid content, and antioxidant activity of Limnophila aromatica. J. Food Drug Anal. 2014, 22, 296–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arda, N.; Onay, E.; Koz, O.; Kirmizigul, S. Monosaccharides and polyols from mistletoes (Viscum album L.) growing on two different host species. Biologia (Bratislava) 2003, 58, 1037–1042. [Google Scholar]
- Wink, M. Evolution of secondary metabolites in legumes (Fabaceae). S. Afr. J. Bot. 2013, 89, 164–175. [Google Scholar] [CrossRef] [Green Version]
- Musa, A.M.; Ibrahim, M.A.; Aliyu, A.B.; Abdullahi, M.S.; Tajuddeen, N.; Ibrahim, H.; Oyewale, A.O. Chemical composition and antimicrobial activity of hexane leaf extract of Anisopus mannii (Asclepiadaceae). J. Intercul. Ethnopharmacol. 2015, 4, 129–133. [Google Scholar] [CrossRef]
- Hosseinihashemi, S.K.; Safdari, V.; Kanani, S. Comparative chemical composition of n-hexane and ethanol extractives from the heartwood of black locust. Asian J. Chem. 2013, 25, 929–933. [Google Scholar] [CrossRef]
- Miranda, I.; Sousa, V.; Ferreira, J.; Pereira, H. Chemical characterization and extractives composition of heartwood and sapwood from Quercus faginea. PLoS ONE 2017, 12, e0179268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-González, M.Z.; Nieto-Trujillo, A.; Gutiérrez-Rebolledo, G.A.; García-Martínez, I.; Estrada-Zúñiga, M.E.; Bernabé-Antonio, A.; Cruz-Sosa, F. Lupeol acetate production and antioxidant activity of a cell suspension culture from Cnidoscolus chayamansa leaves. S. Afr. J. Bot. 2019, 125, 30–38. [Google Scholar] [CrossRef]
- Regasini, L.O.; Vieira-Júnior, G.M.; Fernandes, D.C.; Da Silva Bolzani, V.; Cavalheiro, A.J.; Siqueira Silva, D.H. Identification of triterpenes and sterols from Pterogyne nitens (Fabaceae-caesalpinioideae) using high-resolution gas chromatography. J. Chil. Chem. Soc. 2009, 54, 218–221. [Google Scholar] [CrossRef] [Green Version]
- Meela, M.M.; Mdee, L.K.; Masoko, P.; Eloff, J.N. Acetone leaf extracts of seven invasive weeds have promising activity against eight important plant fungal pathogens. S. Afr. J. Bot. 2019, 121, 442–446. [Google Scholar] [CrossRef]
- Bhagwat, M.K.; Datar, A.G. Antifungal activity of herbal extracts against plant pathogenic fungi. Arch. Phytopathol. Plant Prot. 2014, 47, 959–965. [Google Scholar] [CrossRef]
- Liu, S.; Ruan, W.; Li, J.; Xu, H.; Wang, J.; Gao, Y.; Wang, J. Biological control of phytopathogenic fungi by fatty acids. Mycopathologia 2008, 166, 93–102. [Google Scholar] [CrossRef]
- Gołębiowski, M.; Cerkowniak, M.; Boguś, M.I.; Włóka, E.; Dawgul, M.; Kamysz, W.; Stepnowski, P. Free fatty acids in the cuticular and internal lipids of Calliphora vomitoria and their antimicrobial activity. J. Insect Physiol. 2013, 59, 416–429. [Google Scholar] [CrossRef]
- Bergsson, G.; Arnfinnsson, J.; Steingrímsson, Ó.; Thormar, H. In vitro killing of Candida albicans by fatty acids and monoglycerides. Antimicrob. Agents Chemother. 2001, 45, 3209–3212. [Google Scholar] [CrossRef] [Green Version]
- Li, X.C.; Jacob, M.R.; ElSohly, H.N.; Nagle, D.G.; Smillie, T.J.; Walker, L.A.; Clark, A.M. Acetylenic acids inhibiting Azole-resistant Candida albicans from Pentagonia gigantifolia. J. Nat. Prod. 2003, 66, 1132–1135. [Google Scholar] [CrossRef] [PubMed]
- Pohl, C.H.; Kock, J.L.; Thibane, V.S. Antifungal free fatty acids: A review. Sci. Microb. Pathog. Commun. Curr. Res. Technol. Adv. 2011, 1, 61–71. [Google Scholar]
- Calvo-Garrido, C.; Elmer, P.A.; Parry, F.J.; Viñas, I.; Usall, J.; Torres, R.; Agnew, R.H.; Teixidó, N. Mode of action of a fatty acid–based natural product to control Botrytis cinerea in grapes. J. Appl. Microbiol. 2014, 116, 967–979. [Google Scholar] [CrossRef]
- Ueda, T.; Kai, H.; Taniguchi, E. Growth inhibition of soil-borne pathogenic fungi by typical sterols. Soil Biol. Biochem. 1990, 22, 987–991. [Google Scholar] [CrossRef]
- Choi, N.H.; Jang, J.Y.; Choi, G.J.; Choi, Y.H.; Jang, K.S.; Min, B.S.; Dang, Q.L.; Kim, J.C. Antifungal activity of sterols and dipsacus saponins isolated from Dipsacus asper roots against phytopathogenic fungi. Pestic. Biochem. Physiol. 2017, 141, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Yenjit, P.; Issarakraisila, M.; Intana, W.; Chantrapromma, K. Fungicidal activity of compounds extracted from the pericarp of Areca catechu against Colletotrichum gloeosporioides in vitro and in mango fruit. Postharvest Biol. Technol. 2010, 55, 129–132. [Google Scholar] [CrossRef]
- Canzi, E.F.; Marques, F.A.; Teixeira, S.D.; Tozzi, A.M.G.A.; Silva, M.J.; Duarte, R.M.T.; Duarte, M.C.T.; Ruiz, A.L.T.G.; Monteiro, P.A.; de Carvalho, J.E.; et al. Prenylated flavonoids from roots of Dahlstedtia glaziovii (Fabaceae). J. Braz. Chem. Soc. 2014, 25, 995–1001. [Google Scholar]
- Cursino, L.M.C.; Lima, N.M.; Murillo, R.; Nunez, C.V.; Merfort, I.; Humar, M. Isolation of flavonoids from Deguelia duckeana and their effect on cellular viability, AMPK, eEF2, eIF2 and eIF4E. Molecules 2016, 21, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lucca, A.J. Harmful fungi in both agriculture and medicine. Rev. Iberoam. Micol. 2007, 24, 3–13. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Nicasio-Torres, M.P.; Pérez-Hernández, J.; González-Cortazar, M.; Meckes-Fischer, M.; Tortoriello, J.; Cruz-Sosa, F. Production of potential anti-inflammatory compounds in cell suspension cultures of Sphaeralcea angustifolia (Cav.) G. Don. Acta Physiol. Plant. 2016, 38, 209. [Google Scholar] [CrossRef]
- Zwietering, M.H.; Jongenburger, I.; Rombouts, F.M.; van’t Riet, K. Modeling of the bacterial growth curve. Appl. Environ. Microbiol. 1990, 56, 1875–1881. [Google Scholar] [CrossRef] [Green Version]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Wu, T.T.; Shiau, S.Y.; Chang, R.C. Effects of drying temperature and extraction solvent on the antioxidant, color and sensory properties of Angelica sinensis leaves. J. Food Process. Preserv. 2013, 37, 371–379. [Google Scholar] [CrossRef]
- Liu, M.; Li, X.Q.; Weber, C.; Lee, C.Y.; Brown, J.; Liu, R.H. Antioxidant and antiproliferative activities of raspberries. J. Agric. Food Chem. 2002, 50, 2926–2930. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Castro, A.J.; Zapata-Morales, J.R.; Arana-Argáez, V.; Torres-Romero, J.C.; Ramírez-Villanueva, E.; Pérez-Medina, S.E.; Ramírez-Morales, M.A.; Juárez-Méndez, M.A.; Infante-Barrios, Y.P.; Martínez-Gutiérrez, F.; et al. Pharmacological and toxicological study of a chemical-standardized ethanol extract of the branches and leaves from Eysenhardtia polystachya (Ortega) Sarg. (Fabaceae). J. Ethnopharmacol. 2018, 224, 314–322. [Google Scholar] [CrossRef]
- Garland, S.; Goheen, S.; Donald, P.; McDonald, L.; Campbell, J. Application of derivatization gas chromatography/mass spectrometry for the identification and quantitation of pinitol in plant roots. Anal. Lett. 2009, 42, 2096–2105. [Google Scholar] [CrossRef]
- Amadioha, A.C. Controlling rice blast in vitro and in vivo with extracts of Azadirachta indica. Crop Prot. 2000, 19, 287–290. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PGRs (mg/L) | Callus Induction (%) | PGRs (mg/L) | Callus Induction (%) | ||
|---|---|---|---|---|---|
| 2,4-D | KIN | NAA | KIN | ||
| 0.0 | 0.0 | 50.0 ± 0.0 b | 0.0 | 0.0 | 50 ± 0.0 c |
| 0.0 | 0.1 | 66.7 ± 7.2 ab | 0.0 | 0.1 | 58.3 ± 19.1 bc |
| 0.0 | 1.0 | 93.8 ± 6.3 a | 0.0 | 1.0 | 100.0 ± 0.0 a |
| 0.0 | 2.0 | 75.0 ± 10.2 ab | 0.0 | 2.0 | 100.0 ± 0.0 a |
| 0.1 | 0.0 | 81.3 ± 12.0 ab | 0.1 | 0.0 | 75.0 ± 10.2 ab |
| 0.1 | 0.1 | 81.3 ± 12.0 ab | 0.1 | 0.1 | 93.8 ± 6.3 a |
| 0.1 | 1.0 | 93.8 ± 6.3 a | 0.1 | 1.0 | 93.8 ± 6.3 a |
| 0.1 | 2.0 | 87.5 ± 12.5 ab | 0.1 | 2.0 | 100.0 ± 0.0 a |
| 1.0 | 0.0 | 75.0 ± 10.2 ab | 1.0 | 0.0 | 93.8 ± 6.3 a |
| 1.0 | 0.1 | 100.0 ± 0.0 a | 1.0 | 0.1 | 100.0 ± 0.0 a |
| 1.0 | 1.0 | 83.3 ± 14.4 ab | 1.0 | 1.0 | 100.0 ± 0.0 a |
| 1.0 | 2.0 | 100.0 ± 0.0 a | 1.0 | 2.0 | 100.0 ± 0.0 a |
| 2.0 | 0.0 | 83.3 ± 14.4 ab | 2.0 | 0.0 | 100.0 ± 0.0 a |
| 2.0 | 0.1 | 100.0 ± 0.0 a | 2.0 | 0.1 | 100.0 ± 0.0 a |
| 2.0 | 1.0 | 91.7 ± 7.2 a | 2.0 | 1.0 | 100.0 ± 0.0 a |
| 2.0 | 2.0 | 100.0 ± 0.0 a | 2.0 | 2.0 | 100.0 ± 0.0 a |
| Extract | Yield of Extract (%) | |||
|---|---|---|---|---|
| Sapwood | Heartwood | Leaf | Cell Suspension Culture | |
| Fatty hexane | 4.0 | 6.7 | 8.0 | 0.1 |
| Defatted hexane | 5.0 | 8.6 | 10.1 | 0.5 |
| Dichloromethane | 15.0 | 22.8 | 20.0 | 2.4 |
| Methanolic | 21.1 | 36.9 | 30.8 | 23.8 |
| Total | 45.1 | 75.0 | 68.9 | 26.8 |
| Compound Name | Chemical Formula | RT (min) | Abundance (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Fatty Hexane Extract | Defatted Hexane Extract | |||||||||
| Sapwood | Heartwood | Leaf | CSC | Sapwood | Heartwood | Leaf | CSC | |||
| Alkanes | ||||||||||
| Heptacosane | C27H56 | 38.38 | - | - | 2.46 ± 0.02 | - | - | - | - | - |
| Nonacosane | C29H60 | 42.98 | - | - | 6.36 ± 0.01 | - | - | - | - | - |
| Octacosane | C28H58 | 47.26 | - | - | 1.31 ± 0.02 | - | - | - | - | - |
| Aromatic compounds | ||||||||||
| Isophthalic acid | C8H6O4 | 13.9 | 1.19 ± 0.09 | - | - | - | - | - | - | - |
| Terephthalic acid | C8H6O4 | 14.72 | 1.30 ± 0.23 | - | - | - | - | - | - | - |
| Dibutyl phthalate | C16H22O4 | 19.14 | - | 5.49 ± 0.60 | 0.69 ± 0.06 | 3.91 ± 0.03 | 2.51 ± 0.05 | 3.91 ± 0.01 | 1.05 ± 0.05 | 2.48 ± 0.13 |
| 1,2-Dihydroxyanthraquinone | C14H8O4 | 31.87 | - | - | - | - | - | - | 0.28 ± 0.04 | - |
| Bis(2-ethylhexyl) phthalate | C24H38O4 | 34.69 | - | - | - | - | - | - | - | 0.79 ± 0.02 |
| 4,5-Dihydroxyanthraquinone-2-carboxylic acid | C15H8O6 | 41.37 | - | - | 0.47 ± 0.05 | - | - | - | - | - |
| Derivatives from saturated alkanoic acids | ||||||||||
| 2-Hydroxyheptanoic acid | C7H14O3 | 6.41 | 1.23 ± 0.50 | 1.40 ± 0.05 | - | - | - | - | - | - |
| 3-Phenylpropanoic acid | C9H10O2 | 7.12 | - | - | 4.38 ± 0.09 | - | - | - | 6.28 ± 0.33 | - |
| 3-Hydroxyoctanoic acid | C8H16O3 | 7.98 | 0.78 ± 0.06 | 0.80 ± 0.02 | - | - | - | - | - | - |
| 3-(4-Methoxyphenyl) propionic acid | C10H12O3 | 11.49 | - | - | 2.34 ± 0.11 | - | - | - | 2.48 ± 0.13 | - |
| 2,3-Dihydroxypropyl palmitate | C19H38O4 | 36.21 | - | - | - | 0.60 ± 0.04 | - | - | - | 0.53 ± 0.00 |
| Derivatives from unsaturated alkanoic acids | ||||||||||
| 3-Phenylprop-2-enoic acid | C9H8O2 | 9.2 | - | - | 0.55 ± 0.02 | - | - | - | 0.76 ± 0.06 | - |
| Saturated alkanoic acids | ||||||||||
| Tetradecanoic acid | C14H28O2 | 16.07 | - | - | 1.06 ± 0.02 | 0.39 ± 0.00 | 0.20 ± 0.01 | - | 1.56 ± 0.07 | 0.42 ± 0.00 |
| Pentadecanoic acid | C15H30O2 | 18.74 | - | - | - | 0.42 ± 0.01 | - | - | - | 0.33 ± 0.03 |
| Hexadecanoic acid | C16H32O2 | 21.49 | 14.32 ± 0.45 | 10.06 ± 0.13 | 11.48 ± 0.12 | 62.58 ± 0.36 | 6.25 ± 0.08 | 3.37 ± 0.03 | 19.02 ± 0.02 | 63.01 ± 0.07 |
| Heptadecanoic acid | C17H34O2 | 24.22 | - | - | - | 0.85 ± 0.01 | 0.32 ± 0.00 | 0.17 ± 0.02 | - | 0.80 ± 0.00 |
| Octadecanoic acid | C18H36O2 | 26.93 | 1.90 ± 0.09 | 1.49 ± 0.07 | 2.93 ± 0.08 | 6.09 ± 0.00 | 3.11 ± 0.05 | 0.70 ± 0.01 | 5.14 ± 0.10 | 6.91 ± 0.06 |
| Eicosanoic acid | C20H40O2 | 32.17 | - | - | 0.55 ± 0.00 | 1.14 ± 0.00 | - | 0.14 ± 0.01 | 0.92 ± 0.03 | 1.00 ± 0.01 |
| Saturated alkanoic acids | ||||||||||
| Docosanoic acid | C22H44O2 | 37.08 | - | - | 1.19 ± 0.01 | 0.84 ± 0.00 | - | - | 1.51 ± 0.05 | 0.61 ± 0.01 |
| Tetracosanoic acid | C24H48O2 | 41.71 | - | - | 5.09 ± 0.03 | 2.81 ± 0.01 | - | - | 3.95 ± 0.05 | 2.15 ± 0.05 |
| Saturated diacids | ||||||||||
| Hexanedioic acid | C6H10O4 | 8.49 | 0.91 ± 0.01 | 1.79 ± 0.53 | - | - | - | 0.19 ± 0.04 | - | - |
| Heptanedioic acid | C7H12O4 | 10.33 | 1.54 ± 0.02 | - | - | - | - | - | - | - |
| Octanedioic acid | C8H14O4 | 12.44 | 5.41 ± 0.36 | 2.90 ± 0.34 | - | 0.96 ± 0.09 | 0.18 ± 0.00 | 0.21 ± 0.02 | - | 0.82 ± 0.05 |
| Nonanedioic acid | C9H16O4 | 14.87 | 27.93 ± 1.48 | 16.22 ± 1.27 | 1.02 ± 0.02 | 4.18 ± 0.03 | 0.94 ± 0.04 | 1.03 ± 0.01 | 0.92 ± 0.05 | 3.97 ± 0.10 |
| Steroids | ||||||||||
| Campesterol | C28H48O | 50.45 | - | - | - | - | 8.76 ± 0.07 | 9.15 ± 0.20 | - | - |
| Stigmasterol | C29H48O | 51.14 | - | 1.19 ± 0.14 | - | 0.55 ± 0.08 | 23.56 ± 0.19 | 23.50 ± 0.11 | 1.55 ± 0.02 | 2.58 ± 0.08 |
| β-Sitosterol | C29H50O | 52.32 | - | - | - | - | 43.76 ± 0.03 | 43.43 ± 0.14 | - | 1.87 ± 0.01 |
| Stigmastanol | C29H52 | 52.43 | - | - | - | - | 1.30 ± 0.12 | 1.04 ± 0.08 | - | 1.42 ± 0.02 |
| Stigmasta-3,5-dien-7-one | C29H46O | 53.27 | 17.48 ± 0.35 | 19.00 ± 0.29 | - | - | - | - | - | - |
| Sitostenone | C29H48O | 54.05 | - | - | - | - | - | 0.57 ± 0.04 | - | - |
| Triterpenoids | ||||||||||
| β-Amyrin | C30H50O | 52.16 | - | - | 7.13 ± 0.46 | - | - | - | 16.30 ± 0.49 | - |
| Unsaturated alkanoic acids | ||||||||||
| (Z,Z)-9,12-Octadecadienoic acid | C18H32O2 | 25.99 | - | - | 0.67 ± 0.01 | - | - | - | 2.46 ± 0.06 | - |
| (Z,Z,Z)-9,12,15-Octadecatrienoic acid | C18H30O2 | 26.17 | - | - | 1.49 ± 0.01 | - | - | - | 6.88 ± 0.16 | - |
| (Z)-9-Octadecenoic acid | C18H34O2 | 26.18 | - | - | - | - | 0.25 ± 0.01 | 0.25 ± 0.01 | - | - |
| Others | ||||||||||
| 6,10,14-Trimethylpentadecan-2-one | C18H36O | 15.91 | - | - | 0.53 ± 0.03 | - | - | - | 0.66 ± 0.03 | - |
| 3,7,11,15-Tetramethyl-2-hexadecen-1-ol | C20H40O | 25.11 | - | - | - | - | - | - | 2.89 ± 0.15 | - |
| Octacosanol | C28H58O | 48.27 | - | - | 22.68 ± 0.01 | - | - | - | 3.85 ± 0.13 | - |
| Compound Name | Chemical Formula | RT(min) | Abundance (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Dichloromethane Extract | Methanolic Extract | |||||||||
| Sapwood | Heartwood | Leaf | CSC | Sapwood | Heartwood | Leaf | CSC | |||
| Aromatic compounds | ||||||||||
| Vanillin | C8H8O3 | 9.09 | 0.53 ± 0.32 | - | - | - | - | - | - | - |
| Terephthalic acid | C8H6O4 | 14.72 | 0.39 ± 0.12 | - | - | - | - | - | - | - |
| Dibutyl phthalate | C16H22O4 | 19.14 | 3.05 ± 0.82 | 2.97 ± 0.16 | - | 0.56 ± 0.01 | - | - | - | - |
| 1,2-Dihydroxyanthraquinone | C14H8O4 | 31.87 | - | 1.58 ± 0.01 | - | 0.43 ± 0.03 | - | - | - | - |
| Derivatives from saturated alkanoic acids | ||||||||||
| 3-Phenylpropanoic acid | C9H10O2 | 7.12 | - | - | 1.02 ± 0.00 | - | - | - | - | - |
| 3-(4-Methoxyphenyl) propionic acid | C10H12O3 | 11.49 | - | - | 0.65 ± 0.02 | - | - | - | - | - |
| 2,3-Dihydroxypropyl palmitate | C19H38O4 | 36.21 | 1.19 ± 0.00 | 0.72 ± 0.01 | - | 0.96 ± 0.01 | - | - | - | - |
| Mono and disaccharides | ||||||||||
| Ketohexoses | C6H12O6 | * | - | - | - | - | 10.12 ± 0.10 | 3.14 ± 0.08 | 3.61 ± 0.02 | 27.77 ± 0.03 |
| Aldohexoses | C6H12O6 | * | - | - | - | - | 6.30 ± 0.32 | 3.49 ± 0.24 | 2.88 ± 0.07 | 2.79 ± 0.00 |
| Furanoses | C6H12O6 | * | - | - | - | - | 4.67 ± 0.08 | - | 0.96 ± 0.02 | - |
| Disaccharides (glucose with fructose) | C12H22O11 | * | - | - | - | - | 28.24 ± 0.44 | 3.55 ± 0.12 | 1.82 ± 0.00 | 40.91 ± 0.09 |
| Polyols | ||||||||||
| L-threitol | C4H10O4 | 8.77 | - | - | - | - | - | - | - | 0.42 ± 0.00 |
| Xylitol | C5H12O5 | 13.52 | - | - | - | - | 0.40 ± 0.00 | 4.23 ± 0.15 | - | 2.12 ± 0.00 |
| D-pinitol | C7H14O6 | 16.38 | - | - | - | - | 37.00 ± 0.22 | 15.56 ± 0.47 | 74.28 ± 0.01 | - |
| Ribitol | C5H12O5 | 18.11 | - | - | - | - | - | - | - | 9.22 ± 0.01 |
| Sorbitol | C6H14O6 | 19.21 | - | - | - | - | - | 2.07 ± 0.08 | - | 1.84 ± 0.00 |
| Inositol | C6H12O6 | * | - | - | - | - | 7.07 ± 0.05 | 7.00 ± 0.22 | 6.46 ± 0.02 | 5.22 ± 0.01 |
| Saturated diacids | ||||||||||
| Hexanedioic acid | C6H10O4 | 8.49 | - | 0.48 ± 0.02 | - | - | - | - | - | - |
| Octanedioic acid | C8H14O4 | 12.44 | - | 0.74 ± 0.10 | - | - | - | - | - | - |
| Nonanedioic acid | C9H16O4 | 14.87 | 1.94 ± 0.09 | 4.53 ± 0.73 | 0.51 ± 0.01 | 0.63 ± 0.01 | 0.44 ± 0.09 | 1.17 ± 0.49 | - | - |
| Saturated alkanoic acids | ||||||||||
| Tetradecanoic acid | C14H28O2 | 16.07 | - | - | 0.36 ± 0.01 | - | - | - | - | - |
| Hexadecanoic acid | C16H32O2 | 21.49 | 27.42 ± 0.26 | 22.40 ± 0.26 | 5.20 ± 0.00 | 35.02 ± 0.44 | - | - | 2.06 ± 0.00 | 2.05 ± 0.00 |
| Heptadecanoic acid | C17H34O2 | 24.22 | 0.77 ± 0.13 | 0.87 ± 0.02 | - | 0.36 ± 0.00 | - | - | - | - |
| Octadecanoic acid | C18H36O2 | 26.93 | 3.37 ± 0.21 | 3.21 ± 0.04 | 1.44 ± 0.00 | 2.75 ± 0.22 | - | - | 1.88 ± 0.01 | 1.16 ± 0.03 |
| Eicosanoic acid | C20H40O2 | 32.17 | 0.58 ± 0.04 | - | - | 0.42 ± 0.00 | - | - | - | - |
| Docosanoic acid | C22H44O2 | 37.08 | 0.54 ± 0.01 | - | - | 0.29 ± 0.01 | - | - | - | - |
| Tetracosanoic acid | C24H48O2 | 41.71 | 1.25 ± 0.10 | 0.90 ± 0.00 | 0.96 ± 0.03 | 1.27 ± 0.01 | - | - | - | - |
| Sesquiterpenoids | ||||||||||
| β-Selinene | C15H24 | 8.31 | - | - | 0.46 ± 0.01 | - | - | - | - | - |
| γ-Muurolene | C15H24 | 8.74 | - | - | 0.32 ± 0.01 | - | - | - | - | - |
| β-Cadinene | C15H24 | 11.2 | - | - | 0.77 ± 0.03 | - | - | - | - | - |
| 11-Hydroxy-4βH,5α-eremophil-1(10)-ene | C15H26O | 13.42 | - | - | 0.60 ± 0.02 | - | - | - | - | - |
| Trans, trans-farnesol | C15H26O | 15.07 | - | - | 0.49 ± 0.01 | - | - | - | - | - |
| Steroids | ||||||||||
| Campesterol | C28H48O | 50.45 | 1.61 ± 0.11 | 1.76 ± 0.11 | - | 0.82 ± 0.11 | - | - | - | - |
| Stigmasterol | C29H48O | 51.14 | 4.06 ± 0.41 | 3.96 ± 0.16 | - | 6.72 ± 0.05 | - | - | - | - |
| β-Sitosterol | C29H50O | 52.32 | 8.19 ± 0.63 | 7.50 ± 0.55 | - | 6.00 ± 0.07 | - | - | - | - |
| Unsaturated alkanoic acids | ||||||||||
| (Z,Z)-9,12-Octadecadienoic acid | C18H32O2 | 25.99 | 3.99 ± 0.06 | 5.20 ± 0.05 | 0.72 ± 0.03 | 13.63 ± 0.16 | - | - | - | - |
| (Z,Z,Z)-9,12,15-Octadecatrienoic acid, | C18H30O2 | 26.17 | - | - | 2.29 ± 0.00 | 3.79 ± 0.06 | - | - | - | - |
| (Z)-9-Octadecenoic acid | C18H34O2 | 26.18 | 1.62 ± 0.04 | 2.49 ± 0.05 | - | 0.82 ± 0.01 | - | - | - | - |
| Others | ||||||||||
| Malic acid | C4H6O5 | 8.3 | - | - | - | - | 1.95 ± 0.01 | - | - | - |
| L-Threonic acid | C4H8O5 | 9.79 | - | - | - | - | - | - | 0.42 ± 0.00 | - |
| Galactaric acid | C6H10O8 | 20.16 | - | - | - | - | - | 1.68 ± 0.06 | - | - |
| 3,7,11,15-Tetramethyl-2-hexadecen-1-ol | C20H40O | 25.11 | - | - | 1.06 ± 0.00 | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernabé-Antonio, A.; Sánchez-Sánchez, A.; Romero-Estrada, A.; Meza-Contreras, J.C.; Silva-Guzmán, J.A.; Fuentes-Talavera, F.J.; Hurtado-Díaz, I.; Alvarez, L.; Cruz-Sosa, F. Establishment of a Cell Suspension Culture of Eysenhardtia platycarpa: Phytochemical Screening of Extracts and Evaluation of Antifungal Activity. Plants 2021, 10, 414. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020414
Bernabé-Antonio A, Sánchez-Sánchez A, Romero-Estrada A, Meza-Contreras JC, Silva-Guzmán JA, Fuentes-Talavera FJ, Hurtado-Díaz I, Alvarez L, Cruz-Sosa F. Establishment of a Cell Suspension Culture of Eysenhardtia platycarpa: Phytochemical Screening of Extracts and Evaluation of Antifungal Activity. Plants. 2021; 10(2):414. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020414
Chicago/Turabian StyleBernabé-Antonio, Antonio, Alejandro Sánchez-Sánchez, Antonio Romero-Estrada, Juan Carlos Meza-Contreras, José Antonio Silva-Guzmán, Francisco Javier Fuentes-Talavera, Israel Hurtado-Díaz, Laura Alvarez, and Francisco Cruz-Sosa. 2021. "Establishment of a Cell Suspension Culture of Eysenhardtia platycarpa: Phytochemical Screening of Extracts and Evaluation of Antifungal Activity" Plants 10, no. 2: 414. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020414