
Elevated Atmospheric CO2 Concentration Improved C4 Xero-Halophyte Kochia prostrata Physiological Performance under Saline Conditions


Abstract
:1. Introduction
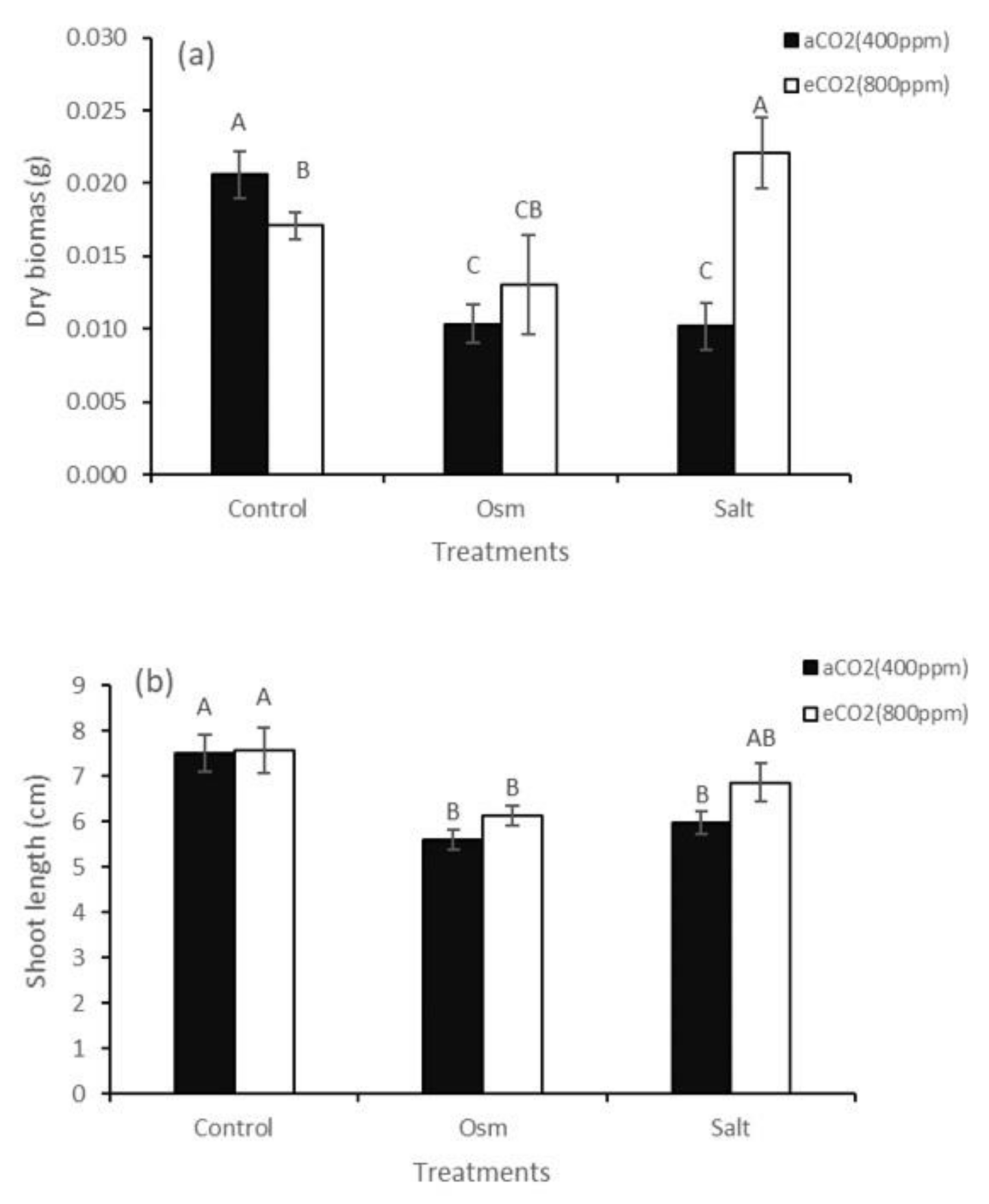
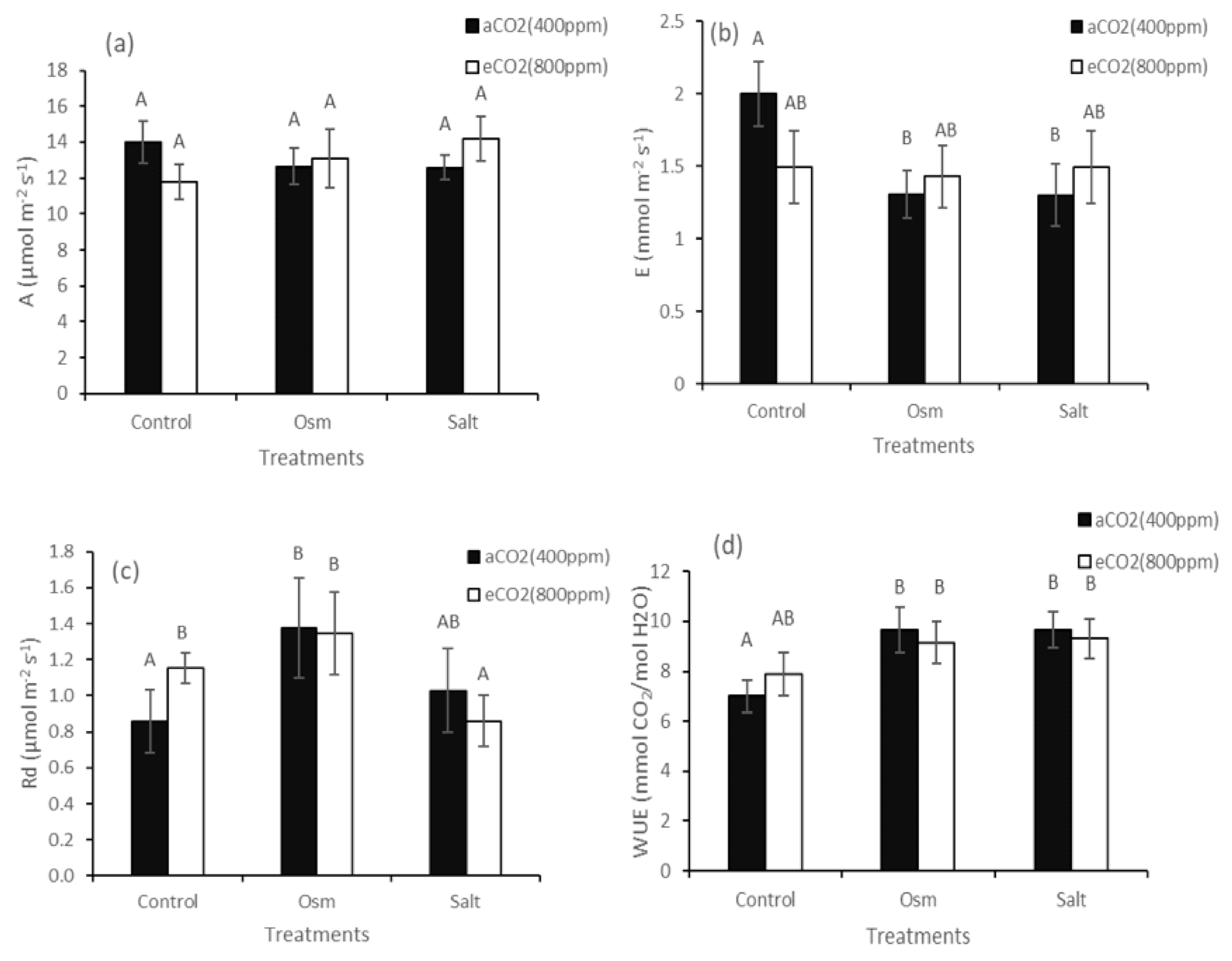
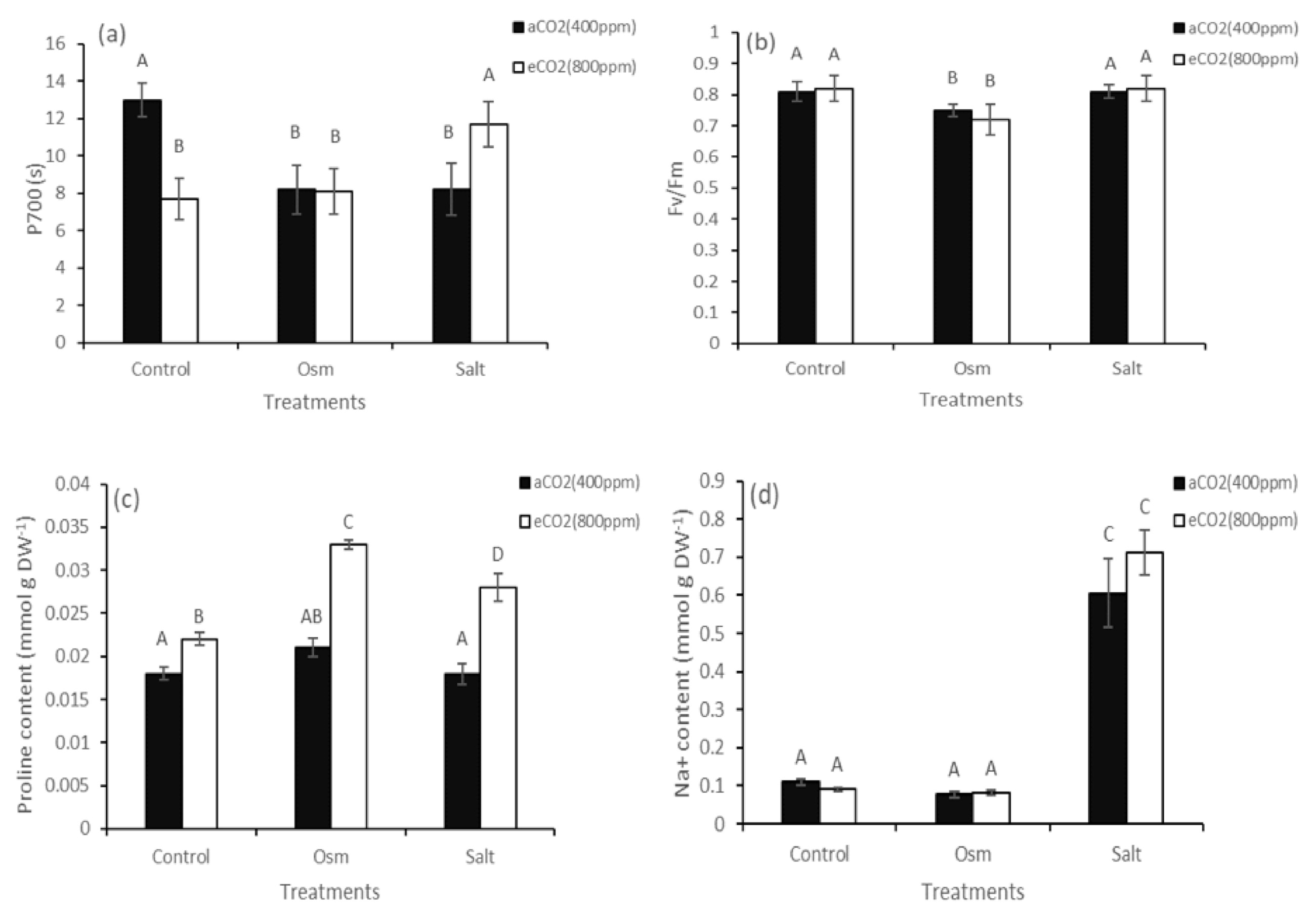
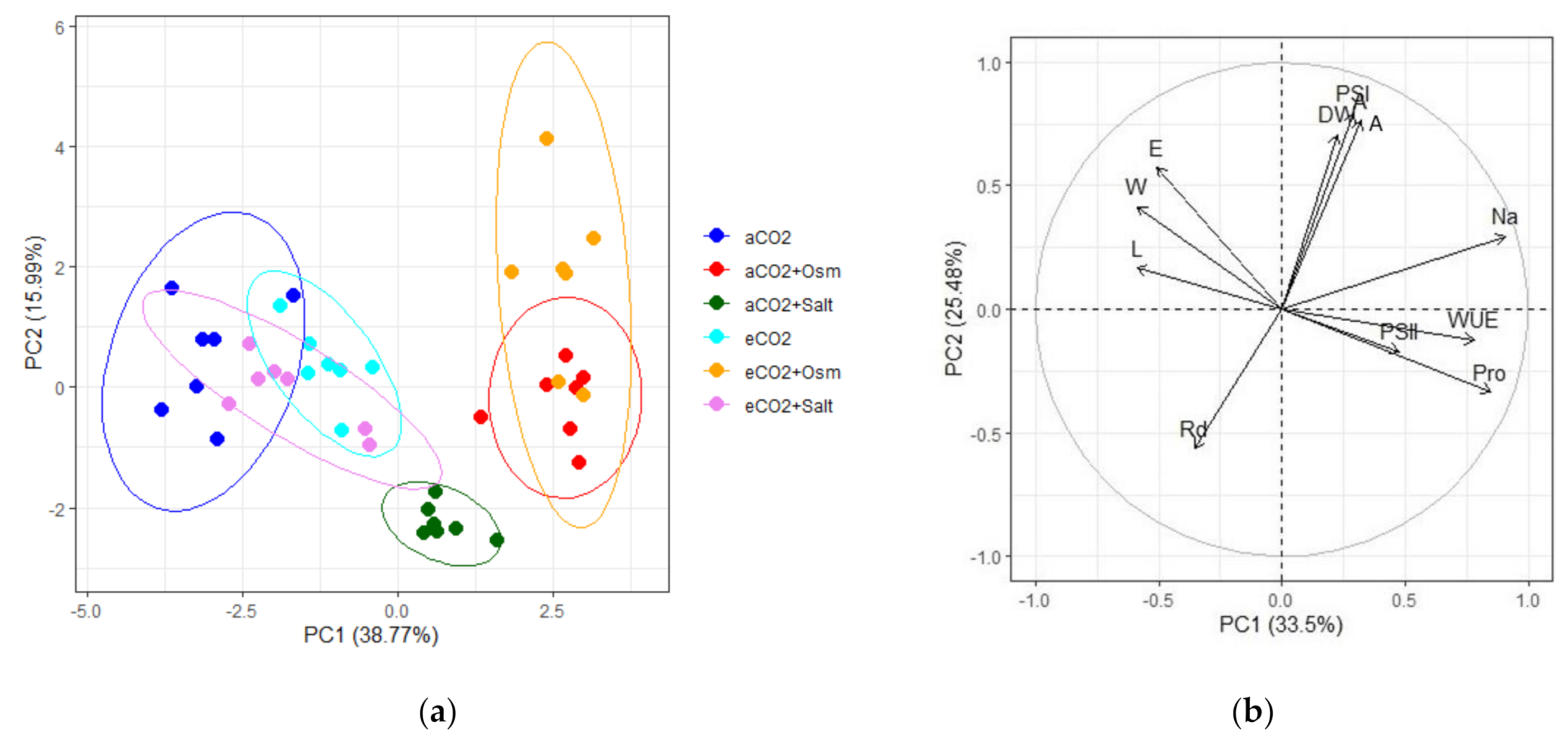
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Growth Conditions
4.3. Dry Biomass and Water Content
4.4. Proline and Na+ Ion Contents
4.5. CO2/H2O Gas Exchange
4.6. Photosystem I
4.7. Photosystem II
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References and Note
- Pérez-Romero, J.A.; Idaszkin, Y.; Barcia-Piedras, J.M.; Duarte, B.; Redondo-Gomez, S.; Caçador, I.; Mateos-Naranjo, E. Disentangling the effect of atmospheric CO2 enrichment on the halophyte Salicornia ramosissima J. Woods physiological performance under optimal and suboptimal saline conditions. Plant. Physiol. Biochem. 2018, 127, 617–629. [Google Scholar] [CrossRef]
- De Faria, A.P.; Marabesi, M.A.; Gaspar, M.; França, M.G.C. The increase of current atmospheric CO2 and temperature can benefit leaf gas exchanges, carbohydrate content and growth in C4 grass invaders of the Cerrado biome. Plant. Physiol. Biochem. 2018, 127, 608–616. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, F.; Hao, L.; Yu, J.; Guo, L.; Zhou, H.; Ma, C.; Zhang, X.; Xu, M. Elevated CO2 concentration induces photosynthetic down-regulation with changes in leaf structure, non-structural carbohydrates and nitrogen content of soybean. BMC Plant. Biol. 2019, 19, 255. [Google Scholar] [CrossRef]
- Sage, R.F.; Kubien, D.S. The temperature response of C3 and C4 photosynthesis. Plant Cell Environ. 2007, 30, 1086–1106. [Google Scholar] [CrossRef]
- Reich, P.B.; Hobbie, S.E.; Lee, T.D.; Pastore, M.A. Unexpected reversal of C3 versus C4 grass response to elevated CO2 during a 20-year field experiment. Science 2018, 360, 317–320. [Google Scholar] [CrossRef] [Green Version]
- Harper, A.B.; Cox, P.M.; Friedlingstein, P.; Wiltshire, A.J.; Jones, C.D.; Sitch, S.; Mercado, L.M.; Groenendijk, M.; Robertson, E.; Kattge, J. Improved representation of plant functional types and physiology in the Joint UK Land Environment Simulator (JULES v4.2) using plant trait information. Geosci. Model. Dev. 2016, 9, 2415–2440. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Gerten, D.; Le Maire, G.; Parton, W.J.; Weng, E.; Zhou, X.; Keough, C.; Beier, C.; Ciais, P.; Cramer, W.; et al. Modeled interactive effects of precipitation, temperature, and [CO2] on ecosystem carbon and water dynamics in different climatic zones. Glob. Chang. Biol. 2008, 14, 1986–1999. [Google Scholar] [CrossRef]
- Jothiramshekar, S.; Benjamin, J.J.; Krishnasamy, R.; Pal, A.K.; George, S.; Swaminathan, R.; Parida, A.K. Responses of selected C3 and C4 halophytes to elevated CO2 concentration under salinity. Curr. Sci. 2018, 115, 129–135. [Google Scholar] [CrossRef]
- Watling, J.R.; Press, M.C.; Quick, W.P. Elevated CO2 induces biochemical and ultrastructural changes in leaves of the C4 cereal Sorghum. Plant. Physiol. 2019, 123, 1143–1151. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Xie, B.; Fu, Y.; Dong, C.; Hui, L.; Guanghui, L.; Liu, H. Effects of different elevated CO2 concentrations on chlorophyll contents, gas exchange, water use efficiency, and PSII activation C3 and C4 cereal crops in a closed artificial ecosystem. Photosynth Res. 2015, 126, 351–362. [Google Scholar] [CrossRef]
- Boretti, A.; Florentine, S. Atmospheric CO2 concentration and other limiting factors in the growth of C3 and C4 plants. Plants 2019, 8, 92. [Google Scholar] [CrossRef] [Green Version]
- Hamim, H. Photosynthesis of C3 and C4 species in response to increased CO2 concentration and drought stress. HAYATI 2005, 12, 131–138. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, H.; Wang, J.; Wu, X.; Ma, S.; Xu, Z.; Zhou, T.; Xu, N.; Tang, X.; An, B. Increased CO2 concentrations increasing water use efficiency and improvement PSII function of mulberry seedling leaves under drought stress. J. Plant. Interact. 2019, 14, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Wand, S.J.E.; Midgley, G.F.; Jones, M.H.; Curtis, P.S. Responses of wild C2 and C3 grass (Poaceae) species to elevated atmospheric CO2 concentration: Ameta-analytic test of current theories and perceptions. Glob. Chang. Biol. 1999, 5, 723–741. [Google Scholar] [CrossRef] [Green Version]
- Lara, M.V.; Andreo, C.S. C4 Plants adaptation to high levels of CO2 and to drought environments. In Abiotic Stress in Plants—Mechanisms and Adaptations; Shanker, A., Ed.; InTech: Vienna, Austria, 2011; Volume 18, pp. 415–428. ISBN 978-953-307-394-1. Available online: http://www.intechopen.com/books/abiotic-stress-in-plantsmechanisms-and-adaptations/c4-plants-adaptation-to-high-levels-of-co2-and-to-drought-environments (accessed on 1 October 2020).
- Leakey, A.D.B. Rising atmospheric carbon dioxide concentration and the future of C4 crops for food and fuel. Proc. R. Soc. B 2009, 276, 2333–2343. [Google Scholar] [CrossRef] [Green Version]
- Quirk, J.; Bellasio, C.; Johnson, D.A.; Beerling, D.J. Response of photosynthesis, growth and water relations of a savannah-adapted tree and grass grown across high to low CO2. Ann. Bot. 2019, 124, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Romero, J.A.; Duarte, B.; Barcia-Piedras, J.-M.; Matos, A.R.; Redondo-Gómeza, S.; Caçadorc, I.; Mateos-Naranjo, E. Investigating the physiological mechanisms underlying Salicorniaramosissima response to atmospheric CO2 enrichment under coexistence of prolonged soil flooding and saline excess. Plant. Physiol. Biochem. 2019, 135, 149–159. [Google Scholar] [CrossRef]
- Nikalje, G.C.; Nikam, T.D.; Suprasanna, P. Looking at halophytic adaptation to high salinity through genomics landscape. Curr. Genom. 2017, 18, 542–552. [Google Scholar] [CrossRef]
- Hamed, K.B.; Ellouzi, H.; Talb, O.Z.; Hessini, K.; Slama, I.; Ghnaya, T.; Bosch, S.M.; Savouré, A.; Abdelly, C. Physiological response of halophytes to multiple stresses. Funct. Plant. Biol. 2013, 40, 883–896. [Google Scholar] [CrossRef]
- Panta, S.; Flowers, T.; Lane, P.; Doyle, R.; Haros, G.; Shabala, S. Halophyte agriculture: Success stories. Environ. Exp. Bot. 2014, 107, 71–83. [Google Scholar] [CrossRef]
- Lauterbach, M.; Schmidt, H.; Billakurthi, K.; Hankeln, T.; Westhoff, P.; Gowik, U.; Kadereit, G. Denovo transcriptome assembly and comparison of C3, C3-C4, and C4 species of tribe Salsoleae (Chenopodiaceae). Front. Plant. Sci. 2017, 8, 1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sage, R.F.; Sage, T.L.; Kocacinar, F. Photorespiration and the evolution of C4 photosynthesis. Annu. Rev. Plant. Biol. 2012, 63, 17.1–17.29. [Google Scholar] [CrossRef]
- Yorimitsu, Y.; Kadosono, A.; Hatakeyama, Y.; Yabiku, T.; Ueno, O. Transition from C3 to proto-Kranz to C3–C4 intermediate type in the genus Chenopodium (Chenopodiaceae). J. Plant. Res. 2019, 132, 839–855. [Google Scholar] [CrossRef] [PubMed]
- Qadir, M.; Tubeileh, A.; Akhtar, J.; Larbi, A.; Minhas, P.; Khan, M. Productivity enhancement of salt-affected environments through crop diversification. Land Degrad. Dev. 2008, 19, 429–453. [Google Scholar] [CrossRef]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant Salinity Stress: Many Unanswered Questions Remain. Front. Plant. Sci. 2019, 10, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flowers, T.J.; Munns, R.; Colmer, T.D. Sodium chloride toxicity and the cellular basis of salt tolerance in halophytes. Ann. Bot. 2015, 115, 419–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghannoum, O.; Von Caemmerer, S.; Ziska, L.H.; Conroy, J.P. The growth response of C4 plants to rising atmospheric CO2 partial pressure: A reassessment. Plant. Cell Environ. 2000, 23, 931–942. [Google Scholar] [CrossRef] [Green Version]
- Reddy, A.R.; Rasineni, G.K.; Raghavendra, A.S. The impact of global elevated CO2 concentration on photosynthesis and plant productivity. Curr. Sci. 2010, 99, 46–57. [Google Scholar]
- Gonzàlez-Meler, M.A.; Blanc-Betes, E.; Flower, C.E.; Ward, J.K.; Gomez-Casanovas, N. Plastic and adaptive responses of plant respiration to changes in atmospheric CO2 concentration. Physiol. Plant. 2009, 137, 473–484. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant. Signal. Behav. 2012, 7, 1456. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, N.; Iwano, M.; Havaux, M.; Yokota, A.; Munekage, Y.N. Promotion of cyclic electron transportaround photosystem I during the evolution of NADP malic enzyme-type C photosynthesis in the genus Flaveria. New Phytol. 2013, 199, 832–842. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Elansary, H.O.; Mattar, M.A.; MElhindi, K.A.; Alotaibi, M.; Mishra, A. Differential Accumulation of Metabolites in Suaeda Species Provides New Insights into Abiotic Stress Tolerance in C4-Halophytic Species in Elevated CO2 Conditions. Agronomy 2021, 11, 131. [Google Scholar] [CrossRef]
- Balyan, G. Kochia prostrata in Kyrgyzstan. Frunze, Kyrgyzstan. 1972.
- Gintzburger, G.; Toderich, K.N.; Mardonov, B.K.; Makhmudov, M.M. Rangelands of the Arid and Semi-Arid Zones in Uzbekistan; Centre de Cooperation Internationale en Resherche Agronomique pour le Development (CIRAD): Monpellier, France, 2003. [Google Scholar]
- Waldron, B.; Eun, J.; ZoBell, D.; Olson, K. Forage kochia (Kochia prostrata) for fall and winter grazing. Small Rumin. Res. 2010, 91, 47–55. [Google Scholar] [CrossRef]
- Karimi, G.; Ghorbanli, M.; Heidari, H.; Khavari Nejad, R.; Assareh, M. The effects of NaCl on growth, water relations, osmolytes and ion content in Kochia prostrata. Biol. Plant. 2005, 49, 301–304. [Google Scholar] [CrossRef]
- Orlovsky, N.S.; Japakova, U.N.; Shulgina, I.; Volis, S. Comparative study of seed germination and growth of Kochia prostrata and Kochia scoparia (Chenopodiaceae) under salinity. J. Arid Environ. 2011, 75, 532–537. [Google Scholar] [CrossRef]
- Bailey, D.; Al Tabini, R.; Waldron, B.; Libbin, J.; Al-Khalidi Kh Alqadi, A.; Al Oun, M.; Jensen, K. Potential of Kochia prostrata and Perennial Grasses for Rangeland Restoration in Jordan. Rangel. Ecol. Manag. 2010, 63, 707–711. [Google Scholar] [CrossRef]
- Malinovsky, A.V.; Akanov, E.N.; Voronin, P.Y. A Vegetation Climatic Unit for Studying the Impact on Higher Plants of an Increased CO2 Concentration in Comparison with the Atmospheric CO2 Concentration. Russ. J. Plant. Physiol. 2020, 67, 194–200. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant. Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Klughammer, C.; Schreiber, U. Measuring P700 absorbance changes in the near infrared spectral region with a dual wavelength pulse modulation system. In Photosynthesis: Mechanisms and Effects; Garab, G., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998; pp. 4357–4360. [Google Scholar]
- Schreiber, U. Chlorophyll Fluorescence and Photosynthetic Energy Conversion: Simple Introductory Experiments with the TEACHING-PAM Chlorophyll Fluorometer; Heinz Walz GmbH: Effeltrich, Germany, 1997. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | PC1 | PC2 |
|---|---|---|
| Photosystem II (PSII) | −0.3219 | −0.3105 |
| Photosystem I (PSI) | −0.2527 | −0.1142 |
| Shoot length (L) | −0.3614 | 0.2312 |
| Dry biomass (DW) | −0.3445 | 0.3182 |
| Water content (W) | −0.2508 | −0.3807 |
| Proline (Pro) | 0.1590 | 0.3675 |
| Na+ ions (Na) | −0.1063 | −0.3773 |
| Apparent photosynthesis (A) | −0.1480 | 0.2795 |
| Transpiration (E) | −0.2643 | 0.3570 |
| Water-use efficiency (WUE) | 0.2540 | −0.0673 |
| Dark respiration (Rd) | 0.3146 | 0.2831 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rakhmankulova, Z.; Shuyskaya, E.; Toderich, K.; Voronin, P. Elevated Atmospheric CO2 Concentration Improved C4 Xero-Halophyte Kochia prostrata Physiological Performance under Saline Conditions. Plants 2021, 10, 491. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030491
Rakhmankulova Z, Shuyskaya E, Toderich K, Voronin P. Elevated Atmospheric CO2 Concentration Improved C4 Xero-Halophyte Kochia prostrata Physiological Performance under Saline Conditions. Plants. 2021; 10(3):491. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030491
Chicago/Turabian StyleRakhmankulova, Zulfira, Elena Shuyskaya, Kristina Toderich, and Pavel Voronin. 2021. "Elevated Atmospheric CO2 Concentration Improved C4 Xero-Halophyte Kochia prostrata Physiological Performance under Saline Conditions" Plants 10, no. 3: 491. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030491