
Regions of Chromosome 2A of Bread Wheat (Triticum aestivum L.) Associated with Variation in Physiological and Agronomical Traits under Contrasting Water Regimes

 , and
, and
Abstract
:1. Introduction
2. Results
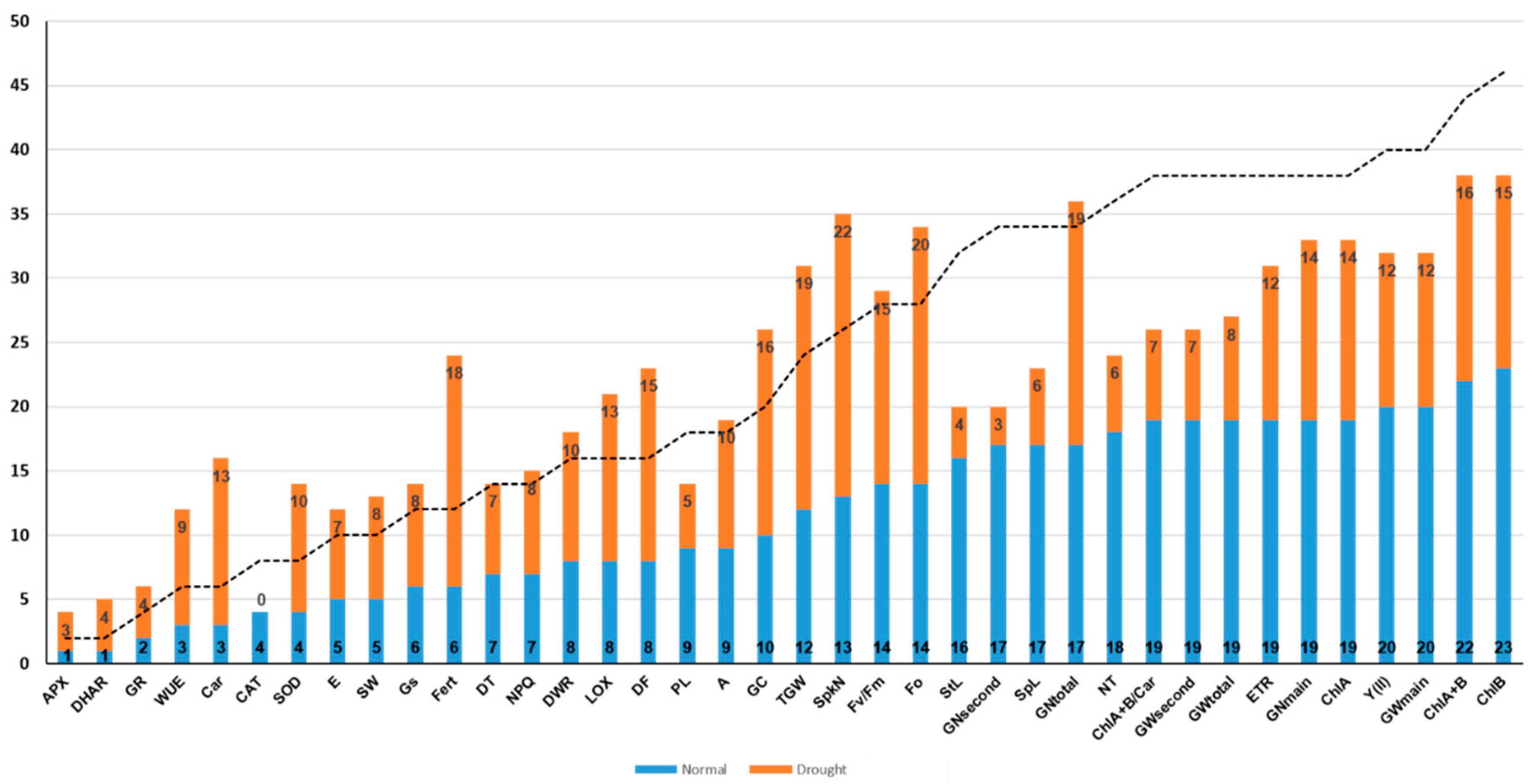
2.1. Trait Variation between S29 and YP and among the SCRDH Lines
2.2. Clustering of SCRDH Lines
2.3. Trait Correlations
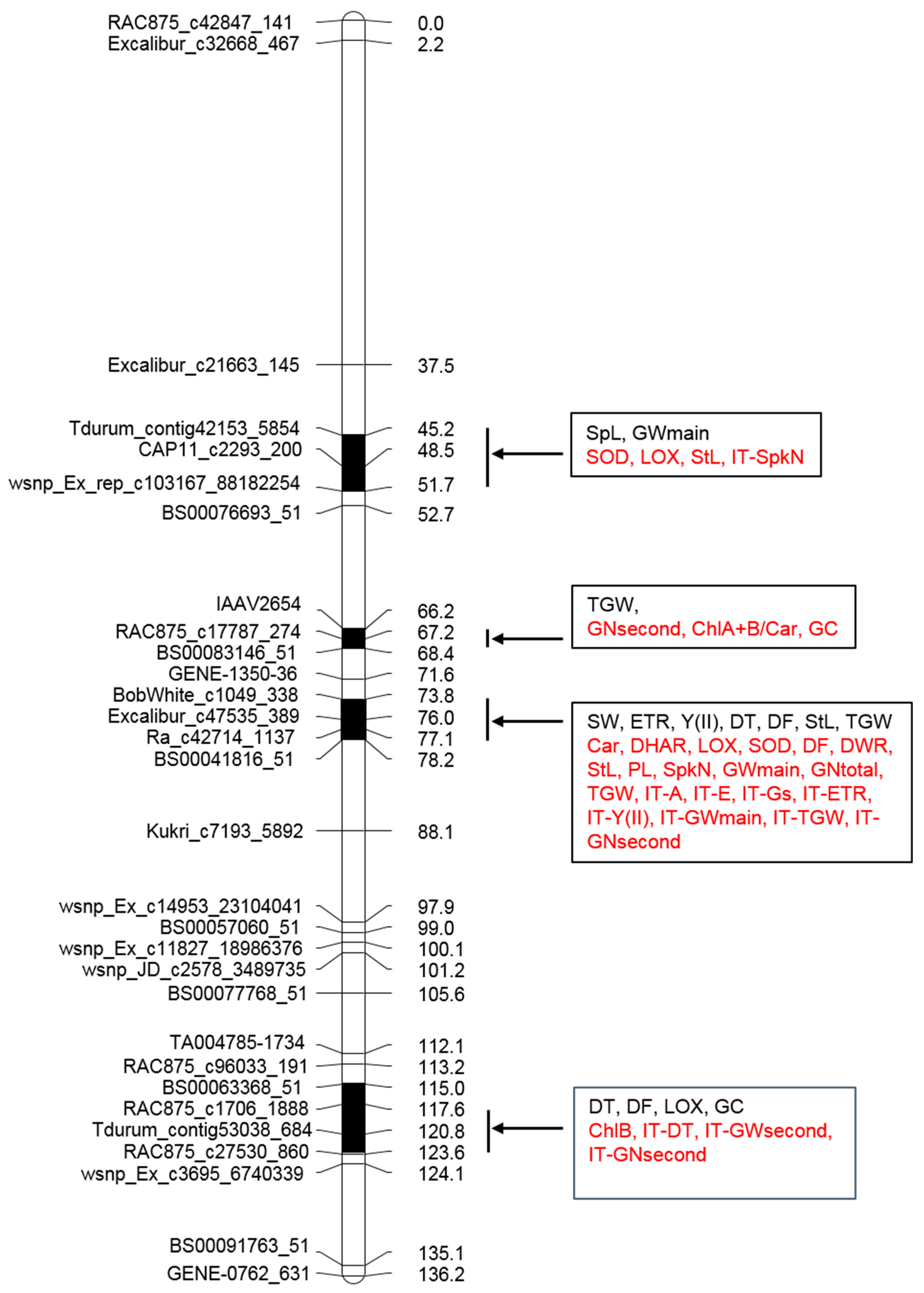
2.4. QTL Analyses
2.5. QTL Clustering
2.6. Candidate Gene Selection
3. Discussion
4. Materials and Methods
4.1. Genetic Material
4.2. Growing Conditions
4.3. Phenotyping
4.4. Genotyping
4.5. Statistical Processing
4.6. Linkage Map Construction and Association Mapping
4.7. Bioinformatic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| A | photosynthetic rate |
| APX | ascorbate peroxidase |
| Car | carotenoids |
| CAT | catalase |
| ChlA, B | chlorophyll A, B |
| DF | days to flowering |
| DHAR | dehydroascorbate reductase |
| DT | days to tillering |
| DWR | days to wax ripening |
| E | transpiration rate |
| ETR | maximum electron transport rate |
| Fert | spike fertility as spikelet number/spike length |
| Fv/Fm | maximum quantum yield of PSII photochemistry |
| F0 | basic chlorophyll fluorescence yield |
| GC | wet gluten content in grain |
| GNmain | grain number in the main spike |
| Gnsecond | grain number in the secondary spikes |
| GNtotal | total grain number from the plant |
| GR | glutathione reductase |
| Gs | stomatal conductance |
| Gwmain | grain weight in the main spike |
| Gwsecond | grain weight in the secondary spikes |
| GWtotal | total grain weight from the plant |
| LOX | lipoxygenase |
| NPQ | non-photochemical quenching |
| NT | number of tillers |
| PL | peduncle length |
| SL | spike length |
| SOD | superoxide dismutase |
| SpkN | spikelets number in the main spike |
| StL | stem length |
| SW | fresh weight of the main shoot |
| TGW | 1000-grain weight as grain weight/grain number |
| WUE | water use efficiency as net photosynthesis/transpiration |
| Y(II) | effective photochemical quantum yield of photosystem II |
| Physiological and Biochemical Traits | Agronomic Traits | ||||
|---|---|---|---|---|---|
| Photosynthesis | Chlorophyll Fluorescence | Leaf Pigments | Enzymes Activity | Developmental | Yield Components |
| A, E, Gs, WUE, SW | F0, Fv/Fm, Y(II), ETR, NPQ | ChlA, ChlB, Car, ChlA+B, ChlA+B/Car | APX, DHAR, SOD, LOX, GR, CAT | DT, DF, DWR | StL, NT, SL, PL, Fert, GNmain, GWmain, GNsecond, GWsecond, GNtotal, GWtotal, TGW, GC |
References
- Touma, D.; Ashfaq, M.; Nayak, M.A.; Kao, S.-C.; Diffenbaugh, N.S. A multi-model and multi-index evaluation of drought characteristics in the 21st century. J. Hydrol. 2015, 526, 196–207. [Google Scholar] [CrossRef] [Green Version]
- Cook, B.I.; Mankin, J.S.; Anchukaitis, K.J. Climate change and drought: From past to future. Curr. Clim. Chang. Rep. 2018, 4, 164–179. [Google Scholar] [CrossRef]
- Asseng, S.; Ewert, F.; Martre, P.; Rötter, R.P.; Lobell, D.B.; Cammarano, D.; Kimball, B.A.; Ottman, M.J.; Wall, G.W.; White, J.W.; et al. Rising temperatures reduce global wheat production. Nat. Clim. Chang. 2015, 5, 143–147. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, S.; Cheng, M.; Jiang, H.; Zhang, X.; Peng, C.; Lu, X.; Zhang, M.; Jin, J. Effect of drought on agronomic traits of rice and wheat: A meta-analysis. Int. J. Environ. Res. Public Health 2018, 15, 839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daryanto, S.; Wang, L.; Jacinthe, P.A. Global synthesis of drought effects on maize and wheat production. PLoS ONE 2016, 11, e0156362. [Google Scholar] [CrossRef]
- McKersie, B. Planning for food security in a changing climate. J. Exp. Bot. 2015, 66, 3435–3450. [Google Scholar] [CrossRef] [Green Version]
- Yasir, T.A.; Wasaya, A.; Hussain, M.; Ijaz, M.; Farooq, M.; Farooq, O.; Nawaz, A.; Hu, Y.G. Evaluation of physiological markers for assessing drought tolerance and yield potential in bread wheat. Physiol. Mol. Biol. Plants 2019, 25, 1163–1174. [Google Scholar] [CrossRef] [PubMed]
- Novaković, L.; Guo, T.; Bacic, A.; Sampathkumar, A.; Johnson, K.L. Hitting the wall-sensing and signaling pathways involved in plant cell wall remodeling in response to abiotic stress. Plants 2018, 7, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, M.; He, C.-Q.; Ding, N.-Z. Abiotic stresses: General defenses of land plants and chances for engineering multistress tolerance. Front. Plant. Sci. 2018, 9, 1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, G.; Sharma, N.; Sahu, P.P.; Prasad, M. Chromatin-based epigenetic regulation of plant abiotic stress response. Curr. Genom. 2016, 17, 490–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapazoglou, A.; Ganopoulos, I.; Tani, E.; Tsaftaris, A. Epigenetics, epigenomics and crop improvement. In Advances in Botanical Research; Kuntz, M., Ed.; Academic Press: London, UK, 2018; Volume 86. [Google Scholar] [CrossRef]
- Sneller, C.H.; Mather, D.E.; Crepieux, S. Analytical approaches and population types for finding and utilizing QTL in complex plant populations. Crop Sci. 2009, 49, 363–380. [Google Scholar] [CrossRef]
- Jannink, J.-L.; Lorenz, A.J.; Iwata, H. Genomic selection in plant breeding: From theory to practice. Briffings Funct. Genom. 2010, 9, 166–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mammadov, J.; Sun, X.; Gao, Y.; Ochsenfeld, C.; Bakker, E.; Ren, R.; Flora, J.; Wang, X.; Kumpatla, S.; Meyer, D.; et al. Combining powers of linkage and association mapping for precise dissection of QTL controlling resistance to gray leaf spot disease in maize (Zea mays L.). BMC Genom. 2015, 16, 916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Li, S.; Wang, Z.; Chang, F.; Kong, J.; Gai, J.; Zhao, T. Identification of major quantitative trait loci for seed oil content in soybeans by combining linkage and genome-wide association mapping. Front. Plant Sci. 2017, 8, 1222. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.; Hao, C.; Zhang, Y.; Cheng, J.; Zhang, Z.; Liu, J.; Yi, X.; Cheng, X.; Sun, D.; Xu, Y.; et al. A combined association mapping and linkage analysis of kernel number per spike in common wheat (Triticum aestivum L.). Front. Plant Sci. 2017, 8, 1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, F.; Wen, W.; Liu, J.; Rasheed, A.; Yin, G.; Xia, X.; Wu, X.; He, Z. Genome-wide linkage mapping of QTL for yield components, plant height and yield-related physiological traits in the Chinese wheat cross Zhou 8425B/Chinese Spring. Front. Plant Sci. 2015, 6, 1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acuña-Galindo, M.A.; Mason, R.E.; Subramanian, N.K.; Hays, D.B. Meta-analysis of wheat QTL regions associated with adaptation to drought and heat stress. Crop Sci. 2015, 55, 477–492. [Google Scholar] [CrossRef]
- Saeed, I.; Chen, X.; Bachir, D.G.; Chen, L.; Hu, Y.-G. Association mapping for photosynthesis and yield traits under two moisture conditions and their drought indices in winter bread wheat (Triticum aestivum L.) using SSR markers. Aust. J. Crop Sci. 2017, 11, 248–257. [Google Scholar] [CrossRef]
- Mwadzingeni, L.; Shimelis, H.; Rees, D.J.G.; Tsilo, T.J. Genome-wide association analysis of agronomic traits in wheat under drought stressed and non-stressed conditions. PLoS ONE 2017, 12, e0171692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Sukumaran, S.; Claverie, E.; Sansaloni, C.; Dreisigacker, S.; Reynolds, M. Genetic dissection of heat and drought stress QTLs in phenology-controlled synthetic-derived recombinant inbred lines in spring wheat. Mol. Breed. 2019, 39, 34. [Google Scholar] [CrossRef]
- Gupta, P.K.; Balyan, H.S.; Sharma, S.; Kumar, R. Genetics of yield, abiotic stress tolerance and biofortification in wheat (Triticum aestivum L.). Theor. Appl. Genet. 2020, 133, 1569–1602. [Google Scholar] [CrossRef] [PubMed]
- International Wheat Genome Sequencing Consortium (IWGSC); Appels, R.; Eversole, K.; Stein, N.; Feuillet, C.; Keller, B.; Rogers, J.; Pozniak, C.J.; Choulet, F.; Distelfeld, A.; et al. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osipova, S.V.; Permyakov, A.V.; Permyakova, M.D.; Pshenichnikova, T.A.; Genaev, M.A.; Börner, A. The antioxidant enzymes activity in leaves of inter-varietal substitution lines of wheat (Triticum aestivum L.) with different tolerance to soil water deficit. Acta Physiol. Plant 2013, 35, 2169–2177. [Google Scholar] [CrossRef]
- Osipova, S.; Permyakov, A.; Permyakova, M.; Rudikovskaya, E.; Pomortsev, A.; Verkhoturov, V.; Pshenichnikova, T. Drought tolerance evaluation of bread wheat (Triticum aestivum L.) lines with the substitution of the second homoeologous group chromosomes. Cereal Res. Commun. 2020, 48, 267–273. [Google Scholar] [CrossRef]
- Ahmad, M.A.; Khan, S.H.; Khan, A.S.; Kazi, A.M.; Basra, S.M.A. Identification of QTLs for drought tolerance traits on wheat chromosome 2A using association mapping. Int. J. Agric. Biol. 2014, 16, 862–870. [Google Scholar]
- Diab, A.A.; Kantety, R.V.; Ozturk, N.Z.; Benscher, D.; Nachit, M.M.; Sorrells, M.E. Drought-inducible genes and differentially expressed sequence tags associated with components of drought tolerance in durum wheat. Sci. Res. Essays 2008, 3, 9–26. [Google Scholar] [CrossRef]
- Yang, D.-L.; Jing, R.-L.; Chang, X.-P.; Li, W. Identification of quantitative trait loci and environmental interactions for accumulation and remobilization of water-soluble carbohydrates in wheat (Triticum aestivum L.) stems. Genetics 2007, 176, 571–575. [Google Scholar] [CrossRef] [Green Version]
- Edae, E.A.; Byrne, P.F.; Haley, S.D.; Lopes, M.S.; Reynolds, M.P. Genome-wide association mapping of yield and yield components of spring wheat under contrasting moisture regimes. Theor. Appl. Genet. 2014, 127, 791–807. [Google Scholar] [CrossRef]
- Yao, J.; Liu, L.; Zheng, Y. Association mapping of agronomic traits on chromosome 2A of wheat. Genetica 2009, 137, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Kabbaj, H.; El Hassouni, K.; Maccaferri, M.; Sanchez-Garcia, M.; Tuberosa, R.; Bassi, F.M. Genomic regions associated with the control of flowering time in durum wheat. Plants 2020, 9, 1628. [Google Scholar] [CrossRef]
- Ilyina, L.G. Breeding of Spring Bread Wheat on Southeast; Saratov University: Saratov, Russia, 1989. (In Russian) [Google Scholar]
- Goltsev, V.N.; Kalaji, H.M.; Paunov, M.; Baba, W.; Horaczek, T.; Mojski, J.; Kociel, H.; Allakhverdiev, S. Variable chlorophyll fluorescence and its use for assessing physiological condition of plant photosynthetic apparatus. Russ. J. Plant Physiol. 2016, 63, 869–893. [Google Scholar] [CrossRef]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef] [PubMed]
- Luna, C.; Pastory, G.; Driscoll, S.; Groten, K. Drought control on H2O2 accumulation, catalase (CAT) activity and CAT gene expression in wheat. J. Exp. Bot. 2005, 56, 417–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, B.N. Role of ascorbic acid in photosynthesis. Biochemistry (Moscow) 2014, 79, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wong, D.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of polyploid wheat genomic diversity using a high-density 90,000 single nucleotide polymorphism array. Plant Biotechnol. J. 2014, 12, 787–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cockram, J.; Mackay, I. Genetic mapping populations for conducting high-resolution trait mapping in plants. In Plant Genetics and Molecular Biology; Varshney, R.K., Pandey, M.R., Chitikineni, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 109–138. [Google Scholar] [CrossRef]
- Hu, X.; Ren, J.; Ren, X.; Huang, S.; Sabiel, S.A.; Luo, M.; Nevo, E.; Fu, C.; Peng, J.; Sun, D. Association of agronomic traits with SNP markers in durum wheat (Triticum turgidum L. durum (Desf.)). PLoS ONE 2015, 10, e0130854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Permiakova, M.D.; Trufanov, V.A.; Pshenichnikova, T.A.; Ermakova, M.F. Role of lipoxygenase in the determination of wheat grain quality. Appl. Biochem. Microbiol. 2010, 46, 87–92. [Google Scholar] [CrossRef]
- Baillo, E.H.; Kimotho, R.N.; Zhang, Z.; Xu, P. Transcription factors associated with abiotic and biotic stress tolerance and their potential for crops improvement. Genes 2019, 10, 771. [Google Scholar] [CrossRef] [Green Version]
- Gahlaut, V.; Jaiswal, V.; Kumar, A.; Gupta, P.K. Transcription factors involved in drought tolerance and their possible role in developing drought tolerant cultivars with emphasis on wheat (Triticum aestivum L.). Theor. Appl. Genet. 2016, 29, 2019–2042. [Google Scholar] [CrossRef]
- Qin, L.; Xue, Y.; Fei, Y.; Zeng, L.; Yang, S.; Deng, X. Identification, evolution and expression analyses of Ribulose-1,5-bisphosphate carboxylase/oxygenase small subunit gene family in wheat (Triticum aestivum L.). Acta Physiol. Plant 2018, 40, 85. [Google Scholar] [CrossRef]
- Wei, Q.; Chen, R.; Wei, X.; Liu, Y.; Zhao, S.; Yin, X.; Xie, T. Genome-wide identification of R2R3-MYB family in wheat and functional characteristics of the abiotic stress responsive gene TaMYB344. BMC Genom. 2020, 21, 792. [Google Scholar] [CrossRef] [PubMed]
- He, Y.-Q.; Huang, W.; Yang, L.; Li, Y.-T.; Lu, C.; Zhu, Y.-X.; Ma, D.-F.; Yin, J.-L. Genome-wide analysis of ethylene-insensitive3 (EIN3/EIL) in Triticum aestivum. Crop Sci. 2020, 60, 2019–2037. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, W. Characterization of the GRAS gene family reveals their contribution to the high adaptability of wheat. PEER J. 2021, 9, e10811. [Google Scholar] [CrossRef] [PubMed]
- Carles, C.; Bies-Etheve, N.; Aspart, L.; Leâon-Kloosterziel, K.M.; Koornneef, M.; Echeverria, M.; Delseny, M. Regulation of Arabidopsis thaliana Em genes: Role of ABI5. Plant J. 2002, 30, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matallana-Ramirez, L.P.; Raufa, M.; Farage-Barhomb, S.; Dortaya, H.; Xue, G.-P.; Dröge-Laser, W.; Lers, A.; Balazadeh, S.; Mueller-Roeber, B. NAC transcription factor ORE1 and senescence-induced BIFUNCTIONAL NUCLEASE1 (BFN1) constitute a regulatory cascade in Arabidopsis. Mol. Plant 2013, 6, 1438–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Nolan, T.M.; Ye, H.; Zhang, M.; Tong, H.; Xin, P.; Chu, J.; Chu, C.; Li, Z.; Yin, Y. Arabidopsis WRKY46, WRKY54, and WRKY70 transcription factors are involved in brassinosteroid-regulated plant growth and drought responses. Plant Cell 2017, 29, 1425–1439. [Google Scholar] [CrossRef]
- Pin, P.A.; Nilsson, O. The multifaceted roles of FLOWERING LOCUS T in plant development. Plant Cell Environ. 2012, 35, 1742–1755. [Google Scholar] [CrossRef]
- Kinoshita, T.; Ono, N.; Hayashi, Y.; Morimoto, S.; Nakamura, S.; Soda, M.; Kato, Y.; Ohnishi, M.; Nakano, T.; Inoue, S.; et al. FLOWERING LOCUS T regulates stomatal opening. Curr. Biol. 2011, 21, 1232–1238. [Google Scholar] [CrossRef] [Green Version]
- Szaker, H.M.; Darkó, É.; Medzihradszky, A.; Janda, T.; Liu, H.; Charng, Y.; Csorba, T. miR824/AGAMOUS-LIKE16 module integrates recurring environmental heat stress changes to fine-tune poststress development. Front. Plant Sci. 2019, 10, 1454. [Google Scholar] [CrossRef] [PubMed]
- Kutter, C.; Schöb, H.; Stadler, M.; Meins, F., Jr.; Si-Ammour, A. MicroRNA-mediated regulation of stomatal development in Arabidopsis. Plant Cell 2007, 19, 2417–2429. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-E.; Ma, J.; Wu, N.; Su, Y.-Q.; Zhang, Z.-W.; Yuan, M.; Zhang, H.-Y.; Zeng, X.-Y.; Yuan, S. The roles of Arabidopsis proteins of Lhcb4, Lhcb5 and Lhcb6 in oxidative stress under natural light conditions. Plant Physiol. Biochem. 2018, 130, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Yabuta, S.; Ifuku, K.; Takabayashi, A.; Ishihara, S.; Ido, K.; Ishikawa, N.; Endo, T.; Sato, F. Three PsbQ-like proteins are required for the function of the chloroplast NAD(P)H dehydrogenase complex in Arabidopsis. Plant Cell Physiol. 2010, 51, 866–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, L.; Gray, J.E. The signaling peptide EPF2 controls asymmetric cell divisions during stomatal development. Curr. Biol. 2009, 19, 864–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geisler, M.; Nadeau, J.; Sack, F.D. Oriented asymmetric divisions that generate the stomatal spacing pattern in Arabidopsis are disrupted by the too many mouths mutation. Plant Cell 2000, 12, 2075–2086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boccalandro, H.E.; Giordano, C.V.; Ploschuk, E.L.; Piccoli, P.N.; Bottini, R.; Casal, J.J. Phototropins but not cryptochromes mediate the blue light-specific promotion of stomatal conductance, while both enhance photosynthesis and transpiration under full sunlight. Plant Physiol. 2012, 158, 1475–1484. [Google Scholar] [CrossRef] [Green Version]
- Kanno, Y.; Hanada, A.; Chiba, Y.; Ichikawa, T.; Nakazawa, M.; Matsui, M.; Koshiba, T.; Kamiya, Y.; Seo, M. Identification of an abscisic acid transporter by functional screening using the receptor complex as a sensor. PNAS 2012, 109, 9653–9658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schilbach, S.; Hantsche, M.; Tegunov, D.; Dienemann, C.; Wigge, C.; Urlaub, H.; Cramer, P. Structures of transcription pre-initiation complex with TFIIH and Mediator. Nature 2017, 551, 204–209. [Google Scholar] [CrossRef]
- Hyun, T.K.; van der Graaff, E.; Albacete, A.; Eom, S.H.; Großkinsky, D.K.; Böhm, H.; Janschek, U.; Rim, Y.; Ali, W.W.; Kim, S.Y.; et al. The Arabidopsis PLAT Domain Protein1 is critically involved in abiotic stress tolerance. PLoS ONE 2014, 9, e112946. [Google Scholar] [CrossRef] [Green Version]
- Higashi, Y.; Okazaki, Y.; Takano, K.; Myouga, F.; Shinozaki, K.; Knoch, E.; Fukushima, A.; Saito, K. HEAT INDUCIBLE LIPASE1 remodels chloroplastic monogalactosyldiacylglycerol by liberating α-linolenic acid in Arabidopsis leaves under heat stress. Plant Cell 2018, 30, 1887–1905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Gügel, I.L.; Giavalisco, P.; Zeisler, V.; Schreiber, L.; Soll, J.; Philippar, K. FAX1, a novel membrane protein mediating plastid fatty acid export. PLoS Biol. 2015, 13, e1002053. [Google Scholar] [CrossRef] [Green Version]
- Aubert, Y.; Vile, D.; Pervent, M.; Aldon, D.; Ranty, B.; Simonneau, T.; Vavasseur, A.; Galaud, J.P. RD20, a stress-inducible caleosin, participates in stomatal control, transpiration and drought tolerance in Arabidopsis thaliana. Plant Cell Physiol. 2010, 12, 1975–1987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balazadeh, S.; Kwasniewski, M.; Caldana, C.; Mehrnia, M.; Zanor, M.I.; Xue, G.-P.; Mueller-Roeber, B. ORS1, an H2O2-esponsive NAC transcription factor, controls senescence in Arabidopsis thaliana. Mol. Plant 2011, 4, 346–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Mao, Y.; Duan, X.; Zhou, H.; Lai, D.; Zhang, Y.; Shen, W. Arabidopsis HY1-modulated stomatal movement: An integrative hub is functionally associated with ABI4 in dehydration-induced ABA responsiveness. Plant Physiol. 2016, 170, 1699–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, X.Y.; Xu, C.Y.; Jing, R.L.; Li, R.Z.; Mao, X.G.; Wang, J.P.; Chang, X.P. Molecular cloning and characterization of wheat calreticulin (CRT) gene involved in drought-stressed responses. J. Exp. Bot. 2008, 59, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Gao, J.; Zhu, X.; Song, Y.; Li, Z.; Ren, G.; Zhou, Z.; Benke Kuai, B. ABF2, ABF3, and ABF4 promote ABA-mediated chlorophyll degradation and leaf senescence by transcriptional activation of chlorophyll catabolic genes and senescence-associated genes in Arabidopsis. Mol. Plant. 2016, 9, 1272–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goetz, M.; Vivian-Smith, A.; Johnson, S.D.; Koltunow, A.M. AUXIN RESPONSE FACTOR8 is a negative regulator of fruit initiation in Arabidopsis. Plant Cell 2006, 18, 1873–1886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurie, D.A.; Reymondie, S. High frequencies of fertilization and haploid seeding production in crosses between commercial hexaploid wheat varieties and maize. Plant Breed. 1991, 106, 182–189. [Google Scholar] [CrossRef]
- Dospekhov, B.A. The Technique of Field Experiment (with the Basic Statistical Processing of Experimantsl Results; Agropromizdat: Moskow, Russia, 1985. (In Russian) [Google Scholar]
- Hammer, Ų.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Peleg, Z.; Saranga, Y.; Suprunova, T.; Ronin, Y.; Röder, M.S.; Kilian, A.; Korol, A.B.; Fahima, T. High-density genetic map of durum wheat wild emmer wheat based on SSR and DArT markers. Theor. Appl. Genet. 2008, 117, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mistry, J.; Finn, R.D.; Eddy, S.R.; Bateman, A.; Punta, M. Challenges in homology search: HMMER3 and convergent evolution of coiled-coil regions. Nucl. Acids Res. 2013, 41, e121. [Google Scholar] [CrossRef] [Green Version]


| Principal Components | Normal Watering | Drought | Tolerance Indexes | |||
|---|---|---|---|---|---|---|
| Contribution to the Total Variation, % | Input of Certain Trait into Principal Component | Contribution to the Total Variation, % | Input of Certain Trait into Principal Component | Contribution to the Total Variation, % | Input of Certain Trait into Principal Component | |
| PC1 | 42.8 | GS (−0.93) DT (0.19) LOX (0.21) WUE (0.11) | 37.6 | LOX (−0.93) DT (0.22) TGW (0.19) SpkN (0.12) GS (−0.12) | 39.6 | DF (0.29), SpkN (0.28), SL (0.28), DWR (0.27), E (−0.25), GS (−0.25), A (−0.25), StL (0.23), PL (0.23), GNmain (0.23), Fert (0.21), DT (0.19) SOD (0.18) |
| PC2 | 16.9 | LOX (0.67) ETR (0.57) F0 (−0.40) GS (0.19) GNmain (0.18) | 27.0 | DT (0.75) GS (−0.59) WUE (0.13) LOX (0.17) | 10.9 | ChlA (0.40), ChlB (0.40), Car (0.40), ChlA+B (0.35), ETR (0.31), Fv/Fm (0.17), F0 (−16), WUE (0.16), E (0.15), GS (0.14) |
| PC3 | 15.0 | DT (0.63) GC (0.23) GNsecond (−0.38) GNtotal (−0.34) GNmain (−0.20) SpkN (0.13) DF (0.11) SOD (0.15) F0 (0.16) ETR (−0.12) GS (0.18) | 15.8 | GS (0.45) DT (0.23) TGW (0.59) GNsecond (−0.37) GNtotal (−0.31) LOX (0.28) GNmain (−0.19) SpkN (0.13) SOD (0.10) | 7.7 | ETR (0.46), Y(II) (0.45), Fv/Fm (0.40), Car (−0.26), ChB (0.24), ChlA+B (0.24), ChlA (0.22), F0 (−17), NT (0.19), NPQ (0.15) |
| Growing Conditions | Clusters | |||
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |
| Normal watering | High gas exchange: E (164), GS (170), A (139), WUE (77) | Low gas exchange: E (40), GS (37), A (44) Chlorophyll fluorescence: F0 (133), Y(II) (73), ETR (73) Yield components: GNsecond (86), GWsecond (85), GNtotal (86) LOX activity (66) | Chlorophyll content: ChlA (112), ChlB (115), ChlA+B (114) Shoot weight (82) NPQ (113) LOX activity (200) | Low gas exchange: E (74), GS (72), A (83) |
| Drought | High gas exchange: E (134), GS (135), A (135), WUE (69) Yield components: GNsecond (85), GNtotal (84), TGW (116) LOX activity (72) | Low gas exchange: E (74), GS (72) LOX activity (61) | Low gas exchange: E (74), GS (75), WUE (123) LOX activity (146) | Yield components: GNsecond (115), GNtotal (112) LOX activity (112) |
| Trait | Env | QTL (PVE%) | Position (cM) | Linked Marker | p-Value | Add |
|---|---|---|---|---|---|---|
| Physiological and biochemical traits | ||||||
| ETR | 1N | Qetr.icg-2AL (9) | 78.2 | BS00041816_51, Ra_c34214_1320 | 0.0027 | −6.7 |
| Y (II) | 1N | Qyld-ph.icg-2AL (9) | 78.2 | BS00041816_51, Ra_c34214_1320 | 0.0036 | 0.10 |
| ChlA | 1N | QchlA.icg-2AS (10) | 0.00 | BobWhite_c13373_250 | 0.0023 | 0.24 |
| ChlB | 1D | QChlB.icg-2A (7) | 122.5 | RAC875_c27530_860 | 0.0090 | 0.13 |
| ChlA+B | 1N | QChlA_B.icg-2AS (9) | 0.00 | BobWhite_c13373_250 | 0.0031 | 0.32 |
| Car | 1D | QCar.icg-2A (7) | 76.0 | Excalibur_c47535_389, Excalibur_c28017_641 | 0.0089 | 0.12 |
| ChlA+B/Car | 1D | QChlA_B/Car 2AS (8) | 66.2 | IAAV2654 | 0.0077 | −1.4 |
| CAT | 1N | Qcat.icg-2A (6) | 37.5 | Tdurum_contig42423_2448 | 0.0083 | 0.035 |
| GR | 1N | Qgr.icg-2A (8) | 135.7 | RFL_Contig5277_888 | 0.0066 | 0.08 |
| LOX | 1N | Qlox.icg-2AL.1 (8) | 120.3 | BS00065245_51 | 0.0028 | −5.5 |
| 1D | Qlox.icg-2AS.2 (4) | 49.6 | RAC875_rep_c78744_228, Excalibur_c92241_336 | 0.0069 | 5.1 | |
| 1D | Qlox.icg-2A.3 (4) | 73.8 | BobWhite_c1049_338 | 0.0069 | −8.4 | |
| DHAR | 1D | Qdhar.icg-2AL (8) | 77.1 | Ku_c5710_312 | 0.0078 | 0.30 |
| SOD | 1D | Qsod.icg-2AS.1 (10) | 51.7 | wsnp_Ex_rep_c103167_88182254, BS00076693_51 | 0.0012 | 6.9 |
| 1D | Qsod.icg-2A.2 (8) | 76.0 | wsnp_CAP8_c2677_1394934 | 0.0063 | −10.0 | |
| SW | 1N | Qsw.icg-2AL (8) | 77.1 | Ra_c42714_1137, Ku_c5710_312 | 0.0026 | −0.68 |
| Agronomical traits | ||||||
| DT | 3N | Qdtill.icg-2A.1 (8) | 76.0 | Excalibur_c47535_389, Excalibur_c28017_641, wsnp_CAP8_c2677_1394934 | 0.0024 | −1.4 |
| 3N | Qdtill.icg-2AL.2 (7) | 115.06 | BS00063368_51 | 0.0048 | −1.5 | |
| DF | 3N | Qflw.icg-2AL.1 (14) | 77.1 | Ku_c5710_312 | 0.0020 | −1.0 |
| 2D | Qflw.icg-2A.1 (14) | 77.1 | Ra_c42714_1137 | 0.0002 | 6.4 | |
| 5 | Qflw.icg-2AL.2 (12) | 123.6 | wsnp_Ex_c3695_6740339 | 0.0006 | 0.52 | |
| DWR | 2D | Qtill.icg-2A (11) | 77.1 | Ra_c42714_1137 | 0.0008 | 10.2 |
| StL | 3N | Qstl.icg-2AL.1 (8) | 88.1 | GENE-1381_132 | 0.0080 | 9.4 |
| 4N | Qstl.icg-2AL.2 (10) | 77.1 | Ku_c5710_312 | 0.0087 | 3.3 | |
| 4D | Qstl.icg-2A.3 (8) | 49.6 | RAC875_rep_c78744_228 | 0.0084 | 5.1 | |
| 2D | Qstl.icg-2AL.4 (8) | 78.2 | BS00041816_51, Ra_c34214_1320 | 0.0046 | 10.5 | |
| 2D | Qstl.icg-2A.2 (7) | 77.1 | Ra_c42714_1137 | 0.0078 | 10.1 | |
| 5 | Qstl.icg-2AL.4 (15) | 78.2 | Ra_c34214_1320, BS00041816_51 | 0.0002 | 2.2 | |
| SpL | 3N | Qsl.icg-2AS.1 (9) | 45.2 | Tdurum_contig42153_5854, RAC875_rep_c111906_144 | 0.0028 | −0.76 |
| 4N | Qsl.icg-2AS.2 (8) | 0.00 | BobWhite_c13373_250 | 0.0041 | −3.8 | |
| GNmain | 3N | Qgrms.icg-2SA (8) | 45.2 | Tdurum_contig42153_5854, RAC875_rep_c111906_144 | 0.0039 | −0.4 |
| PL | 4N | Qped.icg-2A (10) | 73.8 | BobWhite_c1049_338 | 0.0019 | 3.5 |
| 5 | Qped.icg-2A (8) | 73.8 | BS00078116_51, BobWhite_c1049_338 | 0.0069 | −1.6 | |
| TGW | 3N | Qtgw-2AS.1 (11) | 68.4 | wsnp_Ex_c15822_24204224, BS00083146_51 | 0.0008 | 8.6 |
| 3N | Qtgw-2A.2 (7) | 77.1 | Ra_c42714_1137 | 0.0086 | −5.4 | |
| 4N | Qtgw-2A.3 (6) | 73.8 | BobWhite_c1049_338 | 0.0080 | 0.78 | |
| 3D | Qtgw.icg-2A.2 (11) | 77.1 | Ra_c42714_1137 | 0.0015 | −0.45 | |
| 4D | Qtgw.icg-2AL.2 (10) | 77.1 | Ku_c5710_312 | 0.0053 | −3.0 | |
| SpkN | 3D | Qspln.icg-2A (7) | 77.1 | Ra_c42714_1137 | 0.0064 | −1.7 |
| GWmain | 3D | Qgwms.icg-2A (11) | 76.0 | Excalibur_c47535_389, Excalibur_c28017_641 | 0.0013 | −0.29 |
| GNsecond | 4D | Qgnsec.icg-2AS (7) | 66.2 | IAAV2654, Kukri_c1728_1876 | 0.0087 | −0.68 |
| GNtotal | 3D | Qtotgn.icg-2A (11) | 77.1 | Ra_c42714_1137 | 0.0015 | −0.47 |
| GC | 3N | Qglucont.icg-2A.1 (8) | 120.3 | BS00065245_51 | 0.0045 | 1.7 |
| 3D | Qglucont.icg-2A.2 (8) | 67.2 | RAC875_c17787_274 | 0.0045 | 1.7 | |
| Tolerance indices | ||||||
| IT-A | 1 | QIT-Aphot.icg-2AL (13) | 771 | Ku_c5710_312 | 0.0002 | 171 |
| IT-E | 1 | QIT-Ephot.icg-2AL (10) | 77.1 | Ra_c42714_1137 | 0.0019 | 221 |
| IT-Gs | 1 | QIT-Gsphot.icg-2AL (10) | 77.1 | Ra_c42714_1137 | 0.0020 | 223 |
| IT-ETR | 1 | QIT-etr.icg-2AL (10) | 78.2 | Ra_c34214_1320, BS00041816_51 | 0.0017 | 30 |
| IT-Y(II) | 1 | QIT-yld-ph.icg-2AL (10) | 78.2 | Ra_c34214_1320, BS00041816_51 | 0.0018 | 30 |
| IT-DT | 3 | QIT-dtill.icg-2AL (6) | 123.6 | wsnp_Ex_c3695_6740339 | 0.0082 | 1.8 |
| IT-StL | 3 | QIT-stl.icg-2A (10) | 111.9 | TA004785-1734, Excalibur_c18514_238 | 0.0021 | 11 |
| IT-SpL | 4 | QIT-sl.icg-2AS (6) | 0.00 | BobWhite_c13373_250 | 0.0045 | −4.1 |
| IT-SpkN | 3 | QIT-spln.icg-2AS (11) | 48.5 | CAP11_c2293_200 | 0.0011 | 6.7 |
| IT-GWmain | 3 | QIT-wms.icg-2AL (10) | 77.1 | Ku_c5710_312 | 0.0096 | −20 |
| IT-TGW | 3 | QIT-tgw.icg-2AL (11) | 77.1 | Ku_c5710_312 | 0.0074 | 8.3 |
| IT-GNsecond | 3 | QIT-gnsec.icg-2A (9) | 77.1 | Ra_c42714_1137 | 0.0040 | 24 |
| IT-GWsecond | 3 | QIT-gwst.icg-2AL (11) | 120.3 | BS00065245_51, Tdurum_contig53038_714 | 0.0011 | 9.2 |
| IT-GNtotal | 3 | QIT-totgn.icg-2A (9) | 122.5 | RAC875_c27530_860 | 0.0045 | 12 |
| Chromosome Region (cM) | Gene Number | |||||
|---|---|---|---|---|---|---|
| N | Position | Length | In the Region | per cM | Candidate | Including Candidate TF |
| 1 | 45.2–51.7 | 6.5 | 132 | 20 | 4 | 1 |
| 2 | 66.2–68.3 | 2.1 | 189 | 90 | 9 | 7 |
| 3 | 73.8–78.1 | 4.3 | 523 | 122 | 17 | 5 |
| 4 | 117.6–123.0 | 5.4 | 250 | 46 | 9 | 4 |
| Total gene number | 1094 | - | 39 | 18 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pshenichnikova, T.A.; Osipova, S.V.; Smirnova, O.G.; Leonova, I.N.; Permyakova, M.D.; Permyakov, A.V.; Rudikovskaya, E.G.; Konstantinov, D.K.; Verkhoturov, V.V.; Lohwasser, U.; et al. Regions of Chromosome 2A of Bread Wheat (Triticum aestivum L.) Associated with Variation in Physiological and Agronomical Traits under Contrasting Water Regimes. Plants 2021, 10, 1023. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10051023
Pshenichnikova TA, Osipova SV, Smirnova OG, Leonova IN, Permyakova MD, Permyakov AV, Rudikovskaya EG, Konstantinov DK, Verkhoturov VV, Lohwasser U, et al. Regions of Chromosome 2A of Bread Wheat (Triticum aestivum L.) Associated with Variation in Physiological and Agronomical Traits under Contrasting Water Regimes. Plants. 2021; 10(5):1023. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10051023
Chicago/Turabian StylePshenichnikova, Tatyana A., Svetlana V. Osipova, Olga G. Smirnova, Irina N. Leonova, Marina D. Permyakova, Alexey V. Permyakov, Elena G. Rudikovskaya, Dmitrii K. Konstantinov, Vasiliy V. Verkhoturov, Ulrike Lohwasser, and et al. 2021. "Regions of Chromosome 2A of Bread Wheat (Triticum aestivum L.) Associated with Variation in Physiological and Agronomical Traits under Contrasting Water Regimes" Plants 10, no. 5: 1023. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10051023