
An Argon-Ion-Induced Pale Green Mutant of Arabidopsis Exhibiting Rapid Disassembly of Mesophyll Chloroplast Grana

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Isolation of Ar50-33-pg1, a Pale Green Mutant by Argon-Ion Irradiation
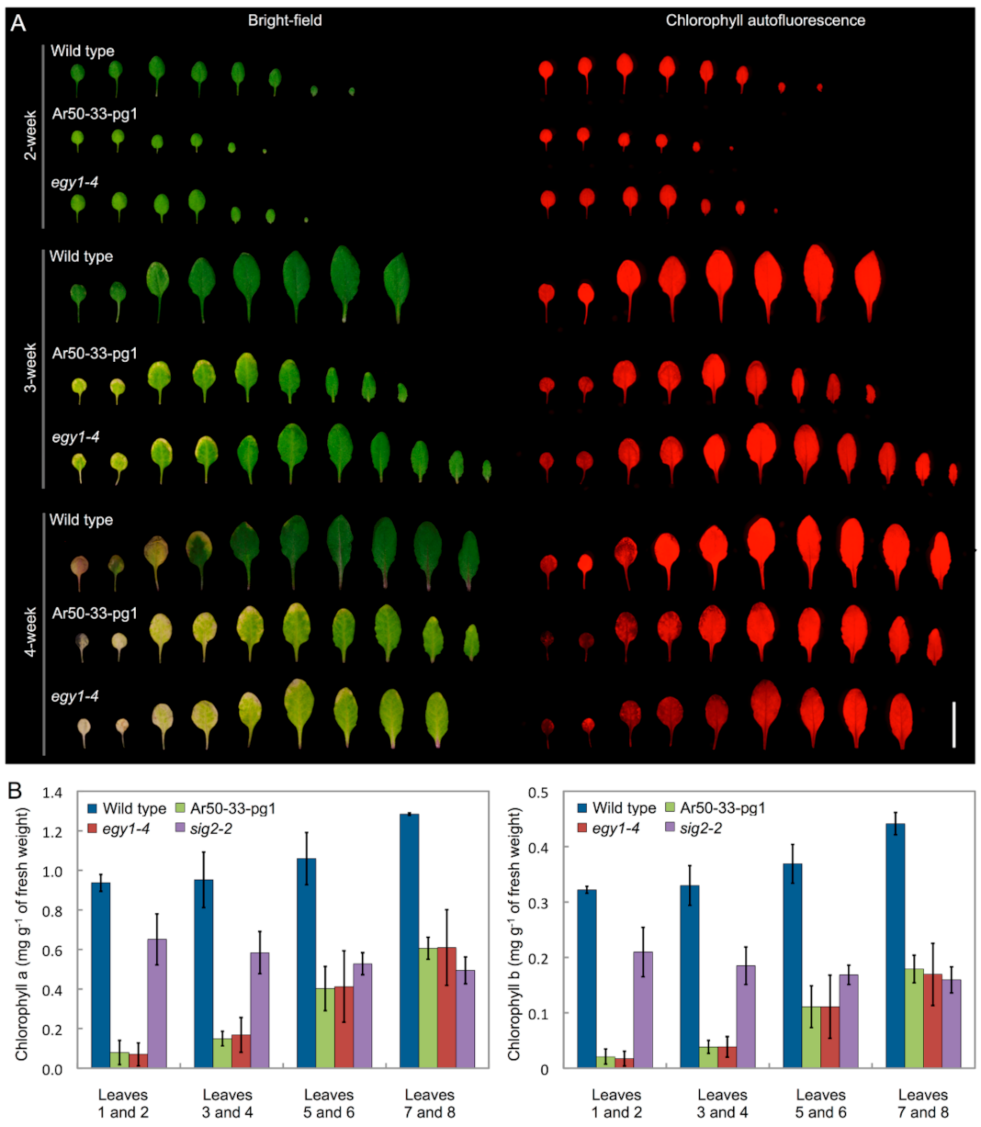
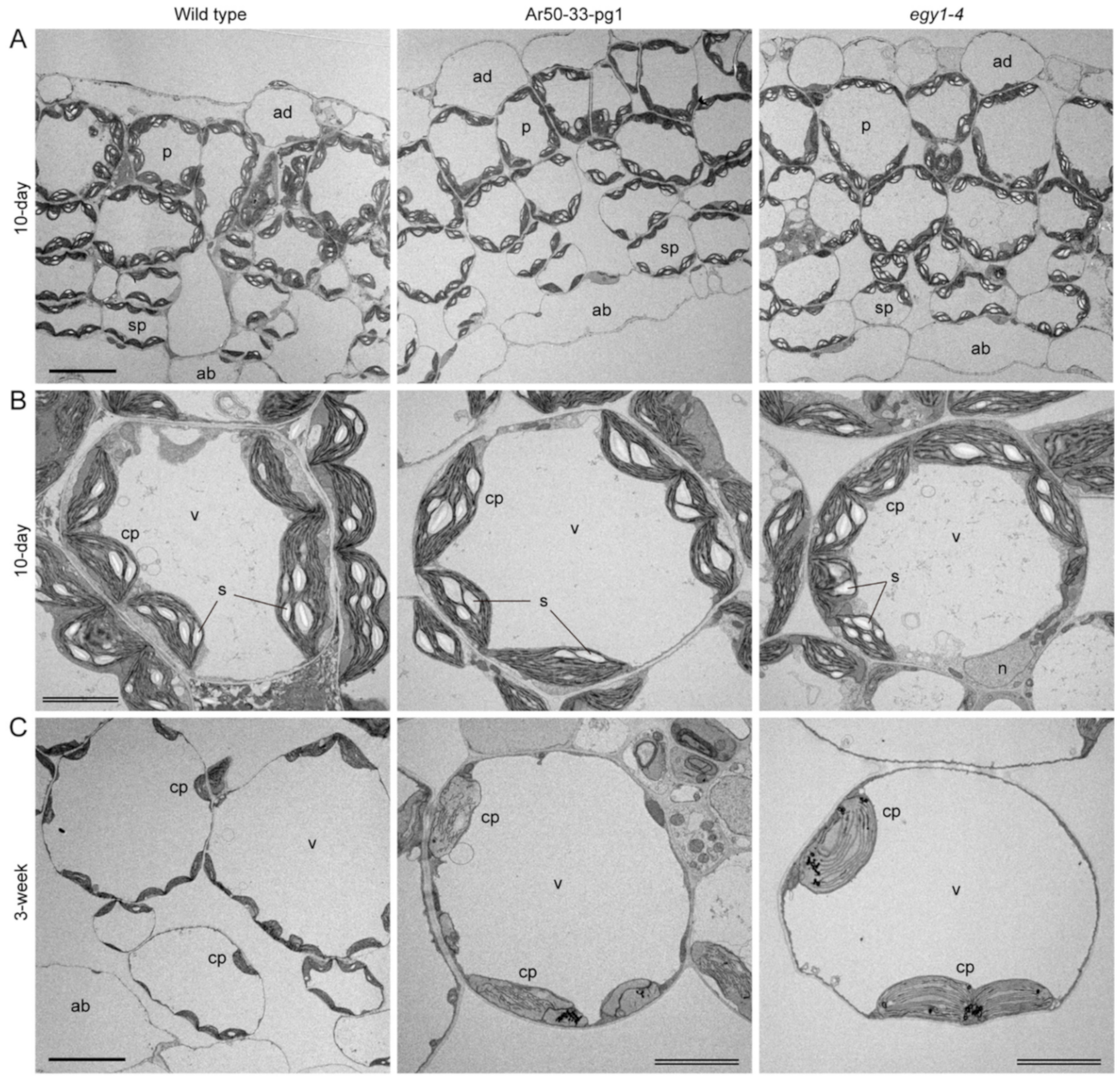
2.2. Chlorosis of Ar50-33-pg1 Leaves Is Associated with Impaired Chloroplast Morphology in Mesophyll Cells
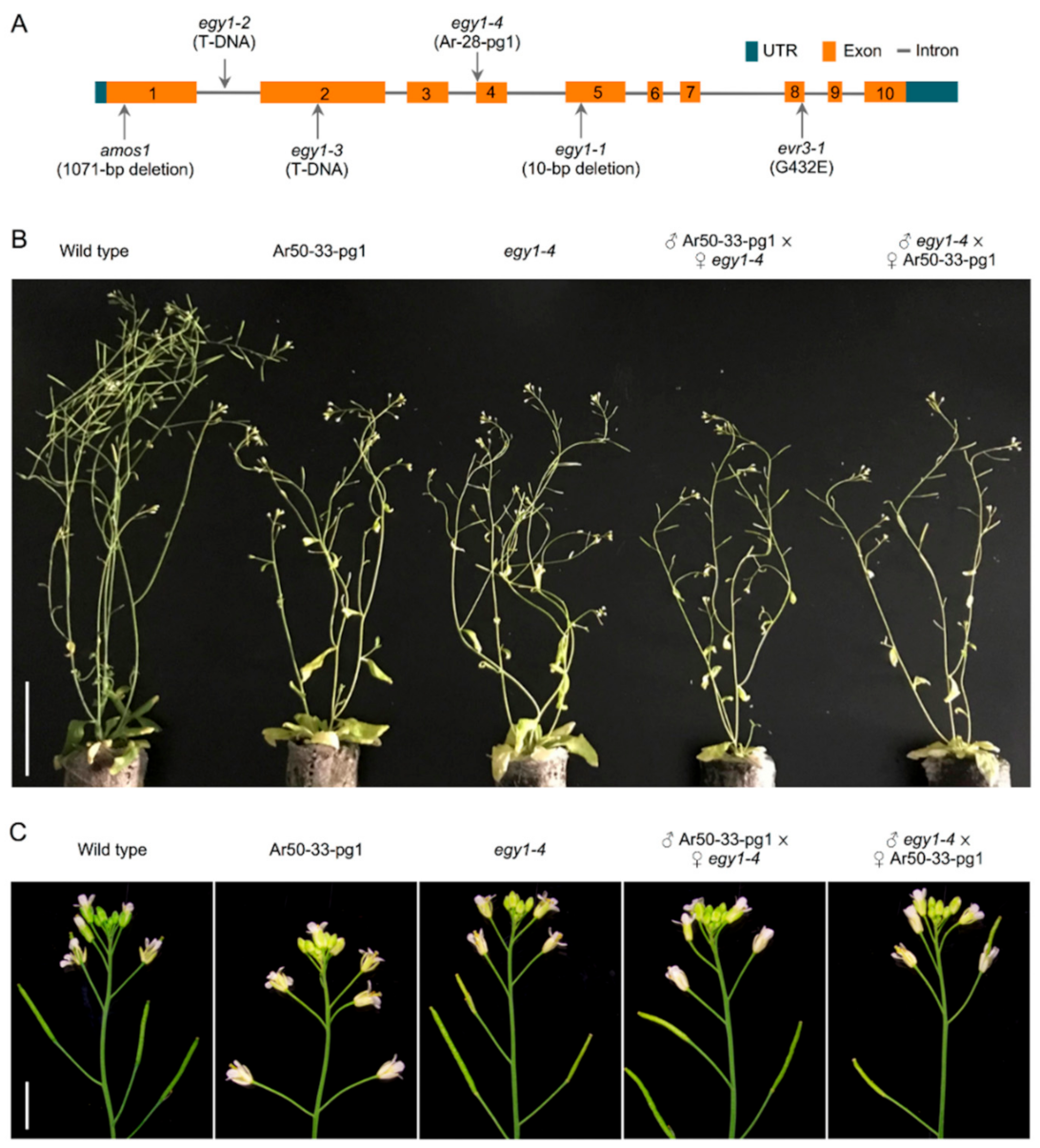
2.3. Identification of a Large Deletion Mutation in Ar50-33-pg1
2.4. EGY1 Is the Main Causal Gene for the Pale Ar50-33-pg1 Phenotype
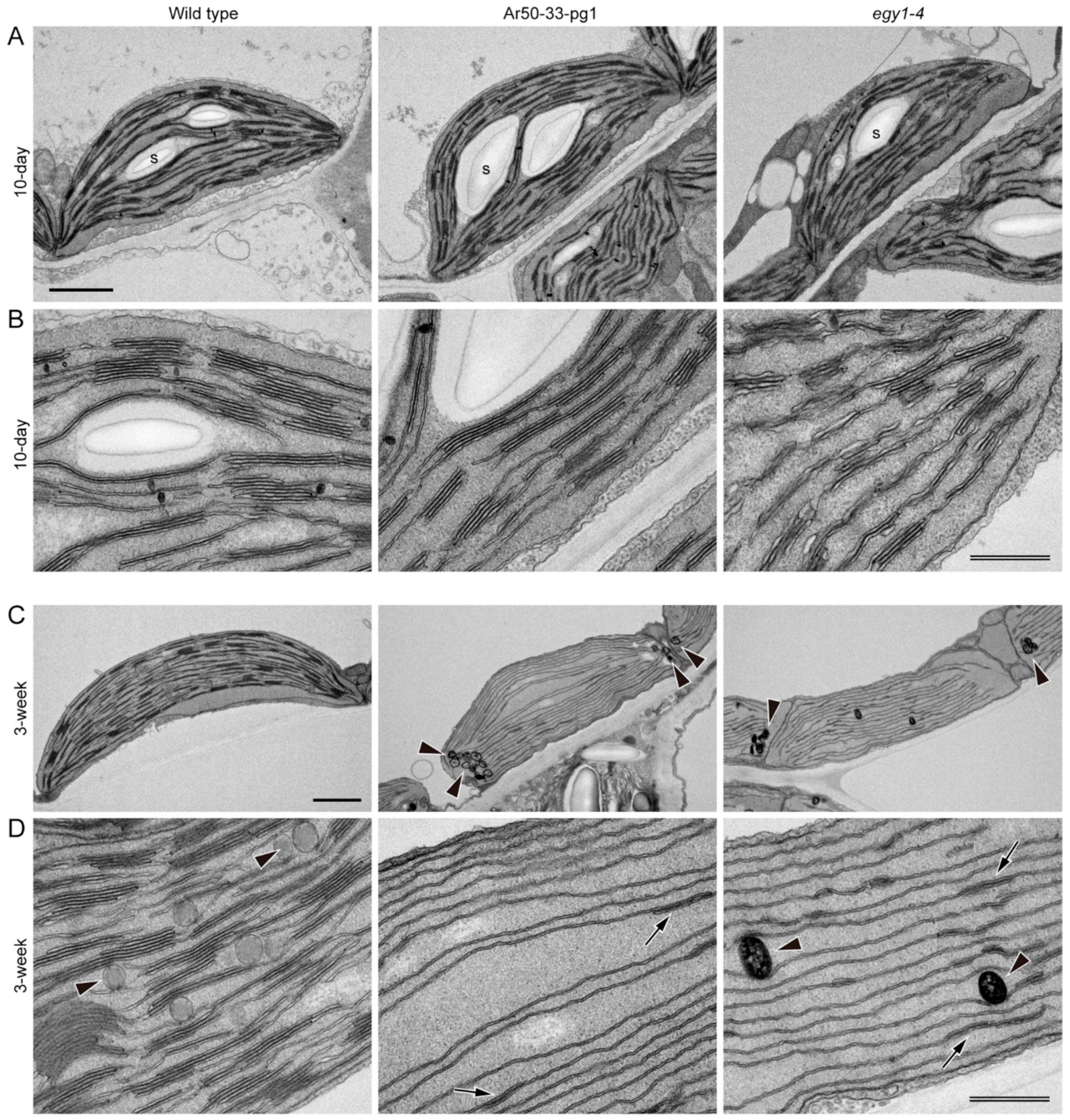
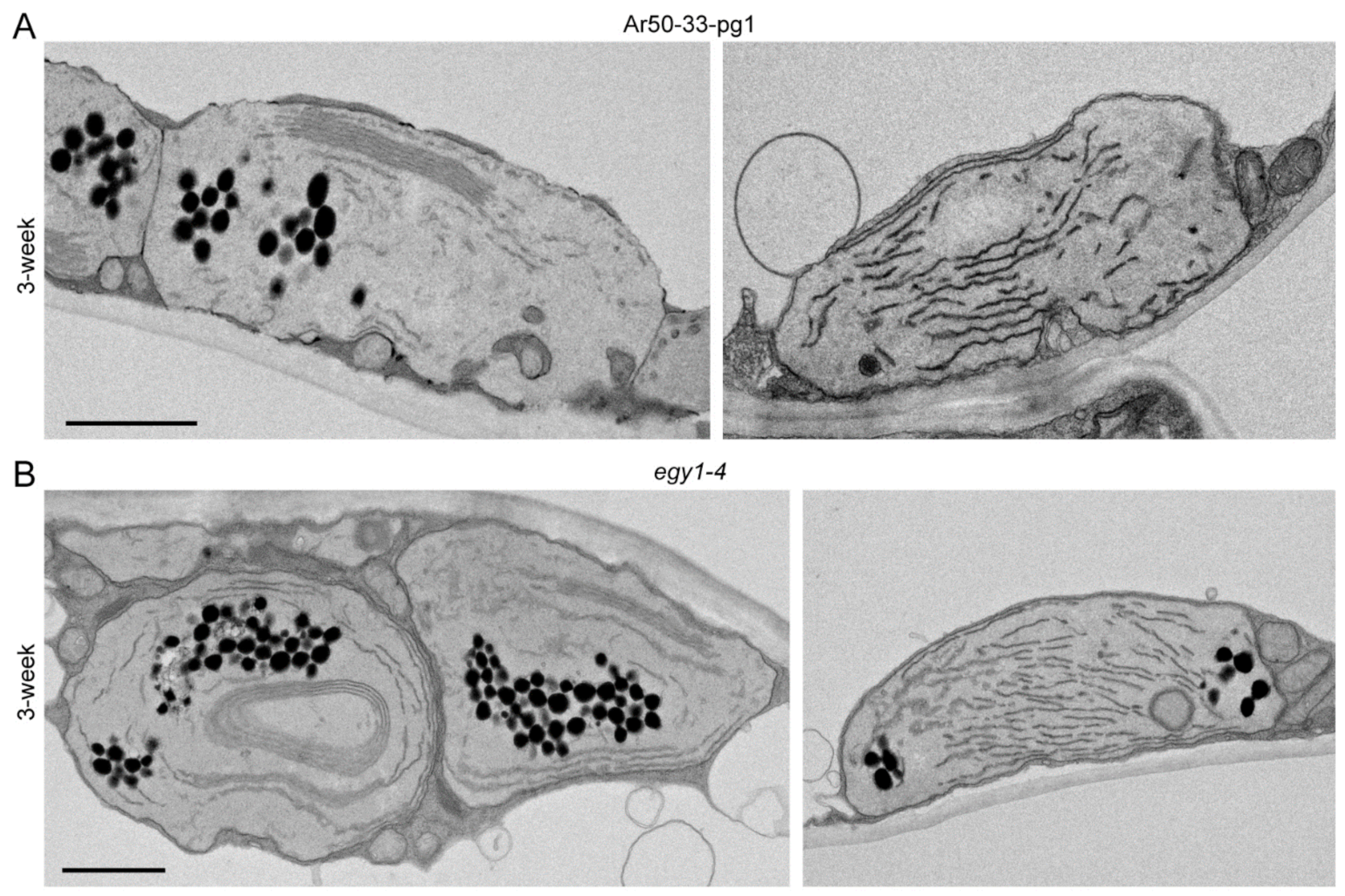
2.5. Mesophyll Chloroplasts of Ar50-33-pg1 and egy1-4 Exhibit Rapid Disassembly of Grana
2.6. Leaf Chlorosis Is Accelerated by Nitrogen Starvation in Ar50-33-pg1 and egy1-4
3. Discussion
3.1. Heavy-Ion Beam-Induced Ar50-33-pg1 as a Resource for Plant Gene Analysis
3.2. Role of EGY1 Homologs in Plants
3.3. Cell Structural Mechanism of Leaf Chlorosis Caused by Loss of EGY1
4. Materials and Methods
4.1. Plant Materials and Growth Condition
4.2. Heavy-Ion Beam Irradiation and Mutant Screening
4.3. Epifluorescence Microscopy
4.4. Fluorescence Stereomicroscopy
4.5. Protoplast Isolation and Organellar Nucleoid Staining
4.6. Measurement of Leaf Chlorophylls
4.7. Whole-Genome Resequencing Analysis and Determination of Mutation Locus
4.8. Electron Microscopy
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mullet, J.E. Chloroplast development and gene expression. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1988, 39, 475–502. [Google Scholar] [CrossRef]
- Wise, R.R. The diversity of plastid form and function. In The Structure and Function of Plastids; Wise, R.R., Hoober, J.K., Eds.; Springer: Dordrecht, The Netherlands, 2007; Volume 23, pp. 3–26. [Google Scholar] [CrossRef]
- Pyke, K. Plastid Biology, 1st ed; Cambridge University Press: Cambridge, UK, 2009; ISBN 978-0521-71197-5. [Google Scholar]
- Hörtensteiner, S. The pathway of chlorophyll degradation: Catabolites, enzymes and pathway regulation. In Plastid Development in Leaves during Growth and Senescence, Advances in Photosynthesis and Respiration; Biswal, B., Krupinska, K., Biswal, U.C., Eds.; Springer: Dordrecht, The Netherlands, 2013; Volume 36, pp. 363–392. [Google Scholar] [CrossRef]
- Tamary, E.; Nevo, R.; Naveh, L.; Levin-Zaidman, S.; Kiss, V.; Savidor, A.; Levin, Y.; Eyal, Y.; Reich, Z.; Adam, Z. Chlorophyll catabolism precedes changes in chloroplast structure and proteome during leaf senescence. Plant Direct 2019, 3, e00127. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Plastoglobuli, thylakoids, chloroplast structure and development of plastids. In Plastid Development in Leaves during Growth and Senescence, Advances in Photosynthesis and Respiration; Biswal, B., Krupinska, K., Biswal, U.C., Eds.; Springer: Dordrecht, The Netherlands, 2013; Volume 36, pp. 337–361. [Google Scholar] [CrossRef]
- Mulisch, M.; Krupinska, K. Ultrastructural analyses of senescence associated dismantling of chloroplasts revisited. In Plastid Development in Leaves during Growth and Senescence, Advances in Photosynthesis and Respiration; Biswal, B., Krupinska, K., Biswal, U.C., Eds.; Springer: Dordrecht, The Netherlands, 2013; Volume 36, pp. 307–335. [Google Scholar] [CrossRef]
- Jarvis, P.; Lopez-Juez, E. Biogenesis and homeostasis of chloroplasts and other plastids. Nat. Rev. Mol. Cell Biol. 2013, 14, 787–802. [Google Scholar] [CrossRef]
- Martin, W.; Herrmann, R.G. Gene transfer from organelles to the nucleus: How much, what happens and why? Plant Physiol. 1998, 118, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Martin, W.; Rujan, T.; Richly, E.; Hansen, A.; Cornelsen, S.; Lins, T.; Leister, D.; Stoebe, B.; Hasegawa, M.; Penny, D. Evolutionary analysis of Arabidopsis, cyanobacterial and chloroplast genomes reveals plastid phylogeny and thousands of cyanobacterial genes in the nucleus. Proc. Natl. Acad. Sci. USA 2002, 99, 12246–12251. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Savage, L.J.; Ajjawi, I.; Imre, K.M.; Yoder, D.W.; Benning, C.; DellaPenna, D.; Ohlrogge, J.B.; Osteryoung, K.W.; Weber, A.P.; et al. New connections across pathways and cellular processes: Industrialized mutant screening reveals novel associations between diverse phenotypes in Arabidopsis. Plant Physiol. 2008, 146, 1482–1500. [Google Scholar] [CrossRef] [Green Version]
- Ajjawi, I.; Lu, Y.; Savage, L.J.; Bell, S.M.; Last, R.L. Large-scale reverse genetics in Arabidopsis: Case studies from the Chloroplast 2010 project. Plant Physiol. 2010, 152, 529–540. [Google Scholar] [CrossRef] [Green Version]
- Kanamaru, K.; Sugita, M. Dynamic features of plastid genome and its transcriptional control in plastid development. In Plastid Development in Leaves during Growth and Senescence, Advances in Photosynthesis and Respiration; Biswal, B., Krupinska, K., Biswal, U.C., Eds.; Springer: Dordrecht, The Netherlands, 2013; Volume 36, pp. 189–213. [Google Scholar] [CrossRef]
- Kato, Y.; Sakamoto, W. Plastid protein degradation during leaf development and senescence: Role of proteases and chaperones. In Plastid Development in Leaves during Growth and Senescence, Advances in Photosynthesis and Respiration; Biswal, B., Krupinska, K., Biswal, U.C., Eds.; Springer: Dordrecht, The Netherlands, 2013; Volume 36, pp. 453–477. [Google Scholar] [CrossRef]
- Osteryoung, K.W.; Pyke, K.A. Division and dynamic morphology of plastids. Annu. Rev. Plant Biol. 2014, 65, 443–472. [Google Scholar] [CrossRef]
- Shimizu, T.; Masuda, T. The role of tetrapyrrole- and GUN1-dependent signaling on chloroplast biogenesis. Plants 2021, 10, 196. [Google Scholar] [CrossRef]
- Chen, G.; Bi, Y.R.; Li, N. EGY1 encodes a membrane-associated and ATP-independent metalloprotease that is required for chloroplast development. Plant J. 2005, 41, 364–375. [Google Scholar] [CrossRef]
- Guo, D.; Gao, X.; Li, H.; Zhang, T.; Chen, G.; Huang, P.; An, L.; Li, N. EGY1 plays a role in regulation of endodermal plastid size and number that are involved in ethylene-dependent gravitropism of light-grown Arabidopsis hypocotyls. Plant Mol. Biol. 2008, 66, 345–360. [Google Scholar] [CrossRef]
- Li, B.; Li, Q.; Xiong, L.; Kronzucker, H.J.; Krämer, U.; Shi, W. Arabidopsis plastid AMOS1/EGY1 integrates abscisic acid signaling to regulate global gene expression response to ammonium stress. Plant Physiol. 2012, 160, 2040–2051. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.W.; Zhu, X.F.; Li, G.J.; Kronzucker, H.J.; Shi, W.M. The chloroplast protease AMOS1/EGY1 affects phosphate homeostasis under phosphate stress. Plant Physiol. 2016, 172, 1200–1208. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Wang, J.; Zhao, X. Leaf senescence induced by EGY1 defection was partially restored by glucose in Arabidopsis thaliana. Bot. Stud. 2016, 57, 5. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.; Wang, X.; Lei, P.; Li, H.; Yan, L.; Zhao, J.; Meng, J.; Shao, J.; An, L.; Yu, F.; et al. The chloroplast metalloproteases VAR2 and EGY1 act synergistically to regulate chloroplast development in Arabidopsis. J. Biol. Chem. 2020, 295, 1036–1046. [Google Scholar] [CrossRef]
- Hara, N. Study of the variegated leaves with special reference to those caused by air spaces. Jpn. J. Bot. 1957, 16, 86–101. [Google Scholar]
- Kirk, J.T.O.; Tilney-Bassett, R.A.E. The Plastids. Their Chemistry, Structure, Growth and Inheritance; Elsevier: Amsterdam, The Netherlands, 1978. [Google Scholar]
- Kazama, Y.; Saito, H.; Fujiwara, M.; Matsuyama, T.; Hayashi, Y.; Ryuto, H.; Fukunishi, N.; Abe, T. An effective method for detection and analysis of DNA damage induced by heavy-ion beams. Biosci. Biotechnol. Biochem. 2007, 71, 2864–2869. [Google Scholar] [CrossRef] [Green Version]
- Kazama, Y.; Hirano, T.; Saito, H.; Liu, Y.; Ohbu, S.; Hayashi, Y.; Abe, T. Characterization of highly efficient heavy-ion mutagenesis in Arabidopsis thaliana. BMC Plant Biol. 2011, 11, 161. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T.; Kazama, Y.; Ohbu, S.; Shirakawa, Y.; Liu, Y.; Kambara, T.; Fukunishi, N.; Abe, T. Molecular nature of mutations induced by high-LET irradiation with argon and carbon ions in Arabidopsis thaliana. Mutat. Res. 2012, 735, 19–31. [Google Scholar] [CrossRef]
- Kazama, Y.; Ma, L.; Hirano, T.; Ohbu, S.; Shirakawa, Y.; Hatakeyama, S.; Tanaka, S.; Abe, T. Rapid evaluation of effective linear energy transfer in heavy-ion mutagenesis in Arabidopsis thaliana. Plant Biotech. 2012, 29, 441–445. [Google Scholar] [CrossRef]
- Kazama, Y.; Hirano, T.; Nishihara, K.; Ohbu, S.; Shirakawa, Y.; Abe, T. Effect of high-LET Fe-ion beam irradiation on mutation induction in Arabidopsis thaliana. Genes Genet. Syst. 2013, 88, 189–197. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T.; Kazama, Y.; Ishii, K.; Ohbu, S.; Shirakawa, Y.; Abe, T. Comprehensive identification of mutations induced by heavy-ion beam irradiation in Arabidopsis thaliana. Plant J. 2015, 82, 93–104. [Google Scholar] [CrossRef]
- Kazama, Y.; Ishii, K.; Hirano, T.; Wakana, T.; Yamada, M.; Ohbu, S.; Abe, T. Different mutational function of low- and high-linear energy transfer heavy-ion irradiation demonstrated by whole-genome resequencing of Arabidopsis mutants. Plant J. 2017, 92, 1020–1030. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, K.; Suzuki, K.I.; Iwaki, K.; Kusumi, T.; Abe, T.; Yoshida, S.; Fukui, H. Flower pigment mutations induced by heavy ion beam irradiation in an interspecific hybrid of Torenia. Plant Biotechnol. 2006, 23, 163–167. [Google Scholar] [CrossRef] [Green Version]
- Kanaya, T.; Saito, H.; Hayashi, Y.; Fukunishi, N.; Ryuto, H.; Miyazaki, K.; Kusumi, T.; Abe, T.; Suzuki, K. Heavy-ion beam-induced sterile mutants of verbena (Verbena × hybrida) with an improved flowering habit. Plant Biotechnol. 2008, 25, 91–96. [Google Scholar] [CrossRef] [Green Version]
- Boyes, D.; Zayed, A.M.; Ascenzi, R.; McCaskill, A.J.; Hoffman, N.E.; Davis, K.R.; Gorlach, J. Growth stage-based phenotypic analysis of Arabidopsis: A model for high throughput functional genomics in plants. Plant Cell 2001, 13, 1499–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noodén, L.D. Defining senescence and death in photosynthetic tissues. In Plastid Development in Leaves during Growth and Senescence, Advances in Photosynthesis and Respiration; Biswal, B., Krupinska, K., Biswal, U.C., Eds.; Springer: Dordrecht, The Netherlands, 2013; Volume 36, pp. 283–306. [Google Scholar] [CrossRef]
- Woodson, J.D.; Perez-Ruiz, J.M.; Schmitz, R.J.; Ecker, J.R.; Chory, J. Sigma factor-mediated plastid retrograde signals control nuclear gene expression. Plant J. 2013, 73, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, S.; Strand, D.D.; Kramer, D.M.; Chen, J.; Montgomery, B.L. Transcriptome and phenotyping analyses support a role for chloroplast sigma factor 2 in red-light-dependent regulation of growth, stress, and photosynthesis. Plant Direct 2018, 2, 1–17. [Google Scholar] [CrossRef]
- Shirano, Y.; Shimada, H.; Kanamaru, K.; Fujiwara, M.; Tanaka, K.; Takahashi, H.; Unno, K.; Sato, S.; Tabata, S.; Hayashi, H.; et al. Chloroplast development in Arabidopsis thaliana requires the nuclear-encoded transcription factor Sigma B. FEBS Lett. 2000, 485, 178–182. [Google Scholar] [CrossRef] [Green Version]
- Kanamaru, K.; Nagashima, A.; Fujiwara, M.; Shimada, H.; Shirano, Y.; Nakabayashi, K.; Shibata, D.; Tanaka, K.; Takahashi, H. An arabidopsis sigma factor (SIG2)-dependent expression of plastid-encoded tRNAs in chloroplasts. Plant Cell Physiol. 2001, 42, 1034–1043. [Google Scholar] [CrossRef] [Green Version]
- Hanaoka, M.; Kanamaru, K.; Takahashi, H.; Tanaka, K. Molecular genetic analysis of chloroplast gene promoters dependent on SIG2, a nucleus-encoded sigma factor for the plastid-encoded RNA polymerase, in Arabidopsis thaliana. Nucleic Acids Res. 2003, 31, 7090–7098. [Google Scholar] [CrossRef] [Green Version]
- Shiina, T.; Ishizaki, Y.; Yagi, Y.; Nakahira, Y. Function and evolution of plastid sigma factors. Plant Biotechnol. 2009, 26, 57–66. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development, 6th ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2015. [Google Scholar]
- Porra, R.J.; Thompson, W.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta 1989, 975, 383–394. [Google Scholar] [CrossRef]
- Sato, N.; Terasawa, K.; Miyajima, K.; Kabeya, Y. Organization, developmental dynamics, and evolution of plastid nucleoids. Int. Rev. Cytol. 2003, 232, 217–262. [Google Scholar] [CrossRef]
- Sakai, A.; Takano, H.; Kuroiwa, T. Organelle nuclei in higher plants: Structure, composition, function, and evolution. Int. Rev. Cytol. 2004, 238, 59–118. [Google Scholar] [CrossRef]
- Fujie, M.; Kuroiwa, H.; Kawano, S.; Mutoh, S.; Kuroiwa, T. Behavior of organelles and their nucleoids in the shoot apical meristem during leaf development in Arabidopsis thaliana L. Planta 1994, 194, 395–405. [Google Scholar] [CrossRef]
- Majeran, W.; Friso, G.; Asakura, Y.; Qu, X.; Huang, M.; Ponnala, L.; Watkins, K.P.; Barkan, A.; van Wijk, K.J. Nucleoid-enriched proteomes in developing plastids and chloroplasts from maize leaves: A new conceptual framework for nucleoid functions. Plant Physiol. 2012, 158, 156–189. [Google Scholar] [CrossRef] [Green Version]
- Golczyk, H.; Greiner, S.; Wanner, G.; Weihe, A.; Bock, R.; Börner, T.; Herrmann, R.G. Chloroplast DNA in mature and senescing leaves: A reappraisal. Plant Cell 2014, 26, 847–854. [Google Scholar] [CrossRef] [Green Version]
- Oldenburg, D.J.; Rowan, B.A.; Kumar, R.A.; Bendich, A.J. Commentary. Plant Cell 2014, 26, 855–861. [Google Scholar] [CrossRef] [Green Version]
- Takami, T.; Ohnishi, N.; Kurita, Y.; Iwamura, S.; Ohnishi, M.; Kusaba, M.; Mimura, T.; Sakamoto, W. Organelle DNA degradation contributes to the efficient use of phosphate in seed plants. Nat. Plants 2018, 4, 1044–1055. [Google Scholar] [CrossRef]
- Greiner, S.; Golczyk, H.; Malinova, I.; Pellizzer, T.; Bock, R.; Börner, T.; Herrmann, R. Chloroplast nucleoids are highly dynamic in ploidy, number, and structure during angiosperm leaf development. Plant J. 2020, 102, 730–746. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, H.; Hirano, M.; Tanaka, H.; Tanaka, T.; Kitagawa-Yogo, R.; Muramoto, N.; Mitsukawa, N. Plastid-targeted forms of restriction endonucleases enhance the plastid genome rearrangement rate and trigger the reorganization of its genomic architecture. Plant J. 2020, 102, 1042–1057. [Google Scholar] [CrossRef]
- Rippert, P.; Puyaubert, J.; Grisollet, D.; Derrier, L.; Matringe, M. Tyrosine and phenylalanine are synthesized within the plastids in Arabidopsis. Plant Physiol. 2009, 149, 1251–1260. [Google Scholar] [CrossRef] [Green Version]
- Romano, P.G.N.; Horton, P.; Gray, J.E. The Arabidopsis cyclophilin gene family. Plant Physiol. 2004, 134, 1268–1282. [Google Scholar] [CrossRef] [Green Version]
- Lange, P.R.; Geserick, C.; Tischendorf, G.; Zrenner, R. Functions of chloroplastic adenylate kinases in Arabidopsis. Plant Physiol. 2008, 146, 492–504. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Feng, P.; Xu, X.; Guo, H.; Ma, J.; Chi, W.; Lin, R.; Lu, C.; Zhang, L. A chloroplast envelope-bound PHD transcription factor mediates chloroplast signals to the nucleus. Nat. Commun. 2011, 2. [Google Scholar] [CrossRef] [Green Version]
- Ottenhof, H.H.; Ashurst, J.L.; Whitney, H.M.; Saldanha, S.A.; Schmitzberger, F.; Gweon, H.S.; Blundell, T.L.; Abell, C.; Smith, A.G. Organisation of the pantothenate (vitamin B5) biosynthesis pathway in higher plants. Plant J. 2004, 37, 61–72. [Google Scholar] [CrossRef] [Green Version]
- Hurley, B.A.; Tran, H.T.; Marty, N.J.; Park, J.; Snedden, W.A.; Mullen, R.T.; Plaxton, W.C. The dual-targeted purple acid phosphatase isozyme AtPAP26 is essential for efficient acclimation of Arabidopsis to nutritional phosphate deprivation. Plant Physiol. 2010, 153, 1112–1122. [Google Scholar] [CrossRef] [Green Version]
- Steffen, J.G.; Kang, I.H.; Macfarlane, J.; Drews, G.N. Identification of genes expressed in the Arabidopsis female gametophyte. Plant J. 2007, 51, 281–292. [Google Scholar] [CrossRef]
- De Oliveira, M.V.V.; Jin, X.; Chen, X.; Griffith, D.; Batchu, S.; Maeda, H.A. Imbalance of tyrosine by modulating TyrA arogenate dehydrogenases impacts growth and development of Arabidopsis thaliana. Plant J. 2019, 97, 901–922. [Google Scholar] [CrossRef]
- Woo, H.H.; Jeong, B.R.; Hirsch, A.M.; Hawes, M.C. Characterization of Arabidopsis AtUGT85A and AtGUS gene families and their expression in rapidly dividing tissues. Genomics 2007, 90, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Su, W.; Liu, Y.; Xia, Y.; Hong, Z.; Li, J. The Arabidopsis homolog of the mammalian os-9 protein plays a key role in the endoplasmic reticulum-associated degradation of misfolded receptor-like kinases. Mol. Plant 2012, 5, 929–940. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Ade, J.; Frye, C.A.; Innes, R.W. Regulation of plant defense responses in Arabidopsis by EDR2, a PH and START domain-containing protein. Plant J. 2005, 44, 245–257. [Google Scholar] [CrossRef] [Green Version]
- Zemach, A.; Grafi, G. Characterization of Arabidopsis thaliana methyl-CpG-binding domain (MBD) proteins. Plant J. 2003, 34, 565–572. [Google Scholar] [CrossRef]
- Ito, M.; Koike, A.; Koizumi, N.; Sano, H. Methylated DNA-binding proteins from Arabidopsis. Plant Physiol. 2003, 133, 1747–1754. [Google Scholar] [CrossRef] [Green Version]
- Hanaoka, H.; Noda, T.; Shirano, Y.; Kato, T.; Hayashi, H.; Shibata, D.; Tabata, S.; Ohsumi, Y. Leaf senescence and starvation-induced chlorosis are accelerated by the disruption of an Arabidopsis autophagy gene. Plant Physiol. 2002, 129, 1181–1193. [Google Scholar] [CrossRef] [Green Version]
- Joshi, P.; Nayak, L. Response of mature, developing and senescing chloroplasts to environmental stress. In Plastid Development in Leaves during Growth and Senescence, Advances in Photosynthesis and Respiration; Biswal, B., Krupinska, K., Biswal, U.C., Eds.; Springer: Dordrecht, The Netherlands, 2013; Volume 36, pp. 307–335. [Google Scholar] [CrossRef]
- Naito, K.; Kusaba, M.; Shikazono, N.; Takano, T.; Tanaka, A.; Tanisaka, T.; Nishimura, M. Transmissible and nontransmissible mutations induced by irradiating Arabidopsis thaliana pollen with g-rays and carbon ions. Genetics 2005, 169, 881–889. [Google Scholar] [CrossRef] [Green Version]
- García, V.J. Characterization of Thylakoid Immunophilins in Arabidopsis. Ph.D. Thesis, University of California Berkeley, Berkeley, CA, USA, 2013. [Google Scholar]
- Barry, C.S.; Aldridge, G.M.; Herzog, G.; Ma, Q.; McQuinn, R.P.; Hirschberg, J.; Giovannoni, J.J. Altered chloroplast development and delayed fruit ripening caused by mutations in a zinc metalloprotease at the lutescent2 locus of tomato. Plant Physiol. 2012, 159, 1086–1098. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Gong, D.; Xia, F.; Dai, C.; Zhang, X.; Gao, X.; Wang, S.; Qu, X.; Sun, Y.; Liu, G. A two-step mutation process in the double WS1 homologs drives the evolution of burley tobacco, a special chlorophyll-deficient mutant with abnormal chloroplast development. Planta 2020, 251, 10. [Google Scholar] [CrossRef]
- Zhang, S.; Zhi, H.; Li, W.; Shan, J.; Tang, C.; Jia, G.; Tang, S.; Diao, X. SiYGL2 is involved in the regulation of leaf senescence and photosystem II efficiency in Setaria italica (L.) P. Beauv. Front. Plant Sci. 2018, 9, 1–15. [Google Scholar] [CrossRef]
- Peltier, J.B.; Ytterberg, A.J.; Sun, Q.; van Wijk, K.J. New functions of the thylakoid membrane proteome of Arabidopsis thaliana revealed by a simple, fast, and versatile fractionation strategy. J. Biol. Chem. 2004, 279, 49367–49383. [Google Scholar] [CrossRef] [Green Version]
- Rudella, A.; Friso, G.; Alonso, J.M.; Ecker, J.R.; van Wijk, K.J. Downregulation of ClpR2 leads to reduced accumulation of the ClpRPS protease complex and defects in chloroplast biogenesis in Arabidopsis. Plant Cell 2006, 18, 1704–1721. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Law, K.; Ho, P.; Zhang, X.; Li, N. EGY2, a chloroplast membrane metalloprotease, plays a role in hypocotyl elongation in Arabidopsis. Mol. Biol. Rep. 2012, 39, 2147–2155. [Google Scholar] [CrossRef] [PubMed]
- Adamiec, M.; Misztal, L.; Kasprowicz-Maluśki, A.; Luciński, R. EGY3: Homologue of S2P protease located in chloroplasts. Plant Biol. 2020, 22, 735–743. [Google Scholar] [CrossRef]
- Klepikova, A.V.; Kasianov, A.S.; Gerasimov, E.S.; Logacheva, M.D.; Penin, A.A. A high resolution map of the Arabidopsis thaliana developmental transcriptome based on RNA-seq profiling. Plant J. 2016, 88, 1058–1070. [Google Scholar] [CrossRef]
- Chen, M.; Choi, Y.; Voytas, D.F.; Rodermel, S. Mutations in the Arabidopsis VAR2 locus cause leaf variegation due to the loss of a chloroplast FtsH protease. Plant J. 2000, 22, 303–313. [Google Scholar] [CrossRef]
- Takechi, K.; Sodmergen Murata, M.; Motoyoshi, F.; Sakamoto, W. The YELLOW VARIEGATED (VAR2) locus encodes a homologue of FtsH, an ATP-dependent protease in Arabidopsis. Plant Cell Physiol. 2000, 41, 1334–1346. [Google Scholar] [CrossRef]
- Evans, I.M.; Rus, A.M.; Belanger, E.M.; Kimoto, M.; Brusslan, J.A. Dismantling of Arabidopsis thaliana mesophyll cell chloroplasts during natural leaf senescence. Plant Biol. 2010, 12, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Nozue, H.; Oono, K.; Ichikawa, Y.; Tanimura, S.; Shirai, K.; Sonoike, K.; Nozue, M.; Hayashida, N. Significance of structural variation in thylakoid membranes in maintaining functional photosystems during reproductive growth. Physiol. Plant. 2017, 160, 111–123. [Google Scholar] [CrossRef]
- Poudyal, R.S.; Rodionova, M.V.; Kim, H.; Lee, S.; Do, E.; Allakhverdiev, S.I.; Nam, H.G.; Hwang, D.; Kim, Y. Combinatory actions of CP29 phosphorylation by STN7 and stability regulate leaf age-dependent disassembly of photosynthetic complexes. Sci. Rep. 2020, 10, 10267. [Google Scholar] [CrossRef] [PubMed]
- Sundberg, E.; Slagter, J.G.; Fridborg, I.; Cleary, S.P.; Robinson, C.; Coupland, G. ALBINO3, an Arabidopsis nuclear gene essential for chloroplast differentiation, encodes a chloroplast protein that shows homology to proteins present in bacterial membranes and yeast mitochondria. Plant Cell 1997, 9, 717–730. [Google Scholar] [CrossRef]
- Gao, H.; Sage, T.L.; Osteryoung, K.W. FZL, an FZO-like protein in plants, is a determinant of thylakoid and chloroplast morphology. Proc. Natl. Acad. Sci. USA 2006, 103, 6759–6764. [Google Scholar] [CrossRef] [Green Version]
- Liang, Z.; Zhu, N.; Mai, K.K.; Liu, Z.; Tzeng, D.; Osteryoung, K.W.; Zhong, S.; Staehelin, L.A.; Kang, B.-H. Thylakoid-bound polysomes and a dynamin related protein, FZL, mediate critical stages of the linear chloroplast biogenesis program in greening Arabidopsis cotyledons. Plant Cell 2018, 30, 1476–1495. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.-L.; Jia, Q.-S.; Yin, Q.-Q.; Lin, G.-N.; Kong, M.-M.; Yang, Z.-N. The GDC1 gene encodes a novel ankyrin domain-containing protein that is essential for grana formation in Arabidopsis. Plant Physiol. 2011, 155, 130–141. [Google Scholar] [CrossRef] [Green Version]
- Armbruster, U.; Labs, M.; Pribil, M.; Viola, S.; Xu, W.; Scharfenberg, M.; Hertle, A.P.; Rojahn, U.; Jensen, P.E.; Rappaport, F.; et al. Arabidopsis CURVATURE THYLAKOID1 proteins modify thylakoid architecture by inducing membrane curvature. Plant Cell 2013, 25, 2661–2678. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, R.; Yamamoto, H.; Kondo, M.; Takeda, S.; Ifuku, K.; Fukao, Y.; Kamei, Y.; Nishimura, M.; Shikanai, T. Grana-localized proteins, RIQ1 and RIQ2, affect the organization of light-harvesting complex II and grana stacking in Arabidopsis. Plant Cell 2016, 28, 2261–2275. [Google Scholar] [CrossRef] [Green Version]
- Pribil, M.; Labs, M.; Leister, D. Structure and dynamics of thylakoids in land plants. J. Exp. Bot. 2014, 65, 1955–1972. [Google Scholar] [CrossRef] [Green Version]
- Svozil, J.; Gruissem, W.; Baerenfaller, K. Proteasome targeting of proteins in Arabidopsis leaf mesophyll, epidermal and vascular tissues. Front. Plant Sci. 2015, 6, 376. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, H.; Nozue, H.; Suzuki, K.; Kaneko, Y.; Taguchi, G.; Hayashida, N. Characterization of the Arabidopsis thaliana mutant pcb2 which accumulates divinyl chlorophylls. Plant Cell Physiol. 2005, 46, 457–473. [Google Scholar] [CrossRef] [Green Version]
- Alonso, J.M.; Stepanova, A.N.; Leisse, T.J.; Kim, C.J.; Chen, H.; Shinn, P.; Stevenson, D.K.; Zimmerman, J.; Barajas, P.; Cheuk, R.; et al. Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 2003, 301, 653–657. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Kazama, Y.; Saito, H.; Yamamoto, Y.Y.; Hayashi, Y.; Ichida, H.; Ryuto, H.; Fukunishi, N.; Abe, T. LET-dependent effects of heavy-ion beam irradiation in Arabidopsis thaliana. Plant Biotechnol. 2008, 25, 113–117. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, H.; Yasuzawa, M.; Koike, N.; Sanjaya, A.; Moriyama, S.; Nishizawa, A.; Matsuoka, K.; Sasaki, S.; Kazama, Y.; Hayashi, Y.; et al. Arabidopsis PARC6 is critical for plastid morphogenesis in pavement, trichome, and guard cells in leaf epidermis. Front. Plant Sci. 2020, 10, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.-H.; Shen, S.-C.; Lee, L.-Y.; Lee, S.-H.; Chan, M.-T.; Lin, C.-S. Tape-Arabidopsis Sandwich—A simpler Arabidopsis protoplast isolation method. Plant Methods 2009, 5, 16. [Google Scholar] [CrossRef] [Green Version]
- Ishii, K.; Kazama, Y.; Hirano, T.; Hamada, M.; Ono, Y.; Yamada, M.; Abe, T. AMAP: A pipeline for whole-genome mutation detection in Arabidopsis thaliana. Genes Genet. Syst. 2016, 91, 229–233. [Google Scholar] [CrossRef] [Green Version]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cross | Generation | Number of Plants | Expected Ratio (Normal: Pale Green) | Chi-Square Value | ||
|---|---|---|---|---|---|---|
| Total | Normal | Pale Green | ||||
| ♂Ar50-33-pg1 * × ♀Col | F1 | 51 | 51 | 0 | 1:0 | 0.00 |
| ♂Ar50-33-pg1 * × ♀Col | F2 | 185 | 145 | 40 | 3:1 | 1.13 |
| ♂Col × ♀Ar50-33-pg1 ** | F1 | 40 | 40 | 0 | 1:0 | 0.00 |
| ♂Col × ♀Ar50-33-pg1 ** | F2 | 477 | 374 | 103 | 3:1 | 2.95 |
| AGI Code | Feature/Localization | Description/Function | Reference |
|---|---|---|---|
| AT5G33390 | Other (1.0) | glycine-rich protein | |
| AT5G33393 | Other (1.0) | myosin heavy chain-like protein | |
| AT5G33406 | Mitochondrion (0.9) | hAT dimerization domain-containing protein/transposase-like protein | |
| AT5G33806 | Other (1.0) | hypothetical protein | |
| AT5G33898 | Other (1.0) | hypothetical protein | |
| AT5G34581 | Other (0.9) | hydroxyproline-rich glycoprotein family protein | |
| AT5G34780 | Other (1.0) | putative ketopantoate reductase (KPR) | [57] |
| AT5G34828 | Signal peptide (1.0) | a Plant thionin family protein | |
| AT5G34829 | Other (0.8) | transmembrane protein | |
| AT5G34830 | Other (1.0) | hypothetical protein | |
| AT5G34850 | Lytic vacuole | AtPAP26; a root-secreted purple acid phosphatase precursor involved in extracellular phosphate-scavenging | [58] |
| AT5G34869 | Other (0.8) | hypothetical protein | |
| AT5G34870 | Other (1.0) | zinc knuckle (CCHC-type) family protein | |
| AT5G34882 | Signal peptide (1.0) | a ECA1 gametogenesis related family protein | |
| AT5G34883 | Signal peptide (1.0) | inhibitor/lipid-transfer protein/seed storage 2S albumin superfamily protein | |
| AT5G34885 | Signal peptide (1.0) | DD17; inhibitor/lipid-transfer protein/seed storage 2S albumin superfamily protein | [59] |
| AT5G34887 | Signal peptide (1.0) | inhibitor/lipid-transfer protein/seed storage 2S albumin superfamily protein | |
| AT5G34905 | Signal peptide (1.0) | ECA1 gametogenesis family protein | |
| AT5G34908 | Signal peptide (1.0) | a ECA1 gametogenesis related family protein | |
| AT5G34930 | Chloroplast stroma | TYRAAt1/TyrA1; arogenate dehydrogenase | [53,60] |
| AT5G34940 | Signal peptide (1.0) | ATGUS3/GUS3; glucuronidase 3 | [61] |
| AT5G35067 | Other (1.0) | hypothetical protein | |
| AT5G35069 | Other (0.6), Mitochondrion (0.3) | a small protein and has either evidence of transcription or purifying selection | |
| AT5G35080 | Endoplasmic reticulum | ATOS9/OS9; a protein involved in the endoplasmic reticulum-associated degradation of glycoproteins | [62] |
| AT5G35090 | Other (1.0) | hypothetical protein | |
| AT5G35100 | Chloroplast thylakoid lumen | AtCYP28; Cyclophilin-like peptidyl-prolyl cis-trans isomerase family protein | [54] |
| AT5G35110 | Other (1.0) | hypothetical protein | |
| AT5G35120 | Other (1.0) | MADS-box family protein | |
| AT5G35160 | Signal peptide (1.0) | TMN11; Endomembrane protein 70 protein family | |
| AT5G35170 | Chloroplast envelope and thylakoid membranes | AMK5; adenylate kinase family protein | [55] |
| AT5G35180 | Mitochondrion | EDR2; ENHANCED DISEASE RESISTANCE protein | [63] |
| AT5G35190 | Signal peptide (1.0) | EXT13; proline-rich extensin-like family protein | |
| AT5G35195 | Other (0.9) | a defensin-like (DEFL) family protein | |
| AT5G35200 | Other (1.0) | PICALM3; ENTH/ANTH/VHS superfamily protein | |
| AT5G35210 | Chloroplast envelope membrane | PTM/DDP1; PHD type transcription factor with transmembrane domains/DDT-PHD protein1 | [56] |
| AT5G35220 | Chloroplast thylakoid membrane | EGY1/AMOS1/EVR3; ethylene-dependent gravitropism-deficient and yellow-green 1/ammonium overly sensitive 1/enchancer of variegation 3 | [17,18,19,20,21,22] |
| AT5G35230 | Other (1.0) | hypothetical protein | |
| AT5G35300 | Other (0.9) | hypothetical protein | |
| AT5G35320 | Other (1.0) | DBH-like monooxygenase | |
| AT5G35330 | Other (1.0) | ATMBD2/MBD2; methyl-cpg-binding domain protein 02 | [64,65] |
| AT5G35338 | Other (1.0) | MBD12; methyl-cpg-binding domain protein 12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanjaya, A.; Kazama, Y.; Ishii, K.; Muramatsu, R.; Kanamaru, K.; Ohbu, S.; Abe, T.; Fujiwara, M.T. An Argon-Ion-Induced Pale Green Mutant of Arabidopsis Exhibiting Rapid Disassembly of Mesophyll Chloroplast Grana. Plants 2021, 10, 848. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10050848
Sanjaya A, Kazama Y, Ishii K, Muramatsu R, Kanamaru K, Ohbu S, Abe T, Fujiwara MT. An Argon-Ion-Induced Pale Green Mutant of Arabidopsis Exhibiting Rapid Disassembly of Mesophyll Chloroplast Grana. Plants. 2021; 10(5):848. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10050848
Chicago/Turabian StyleSanjaya, Alvin, Yusuke Kazama, Kotaro Ishii, Ryohsuke Muramatsu, Kengo Kanamaru, Sumie Ohbu, Tomoko Abe, and Makoto T. Fujiwara. 2021. "An Argon-Ion-Induced Pale Green Mutant of Arabidopsis Exhibiting Rapid Disassembly of Mesophyll Chloroplast Grana" Plants 10, no. 5: 848. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10050848