
Identification of Plant Growth Promoting Rhizobacteria That Improve the Performance of Greenhouse-Grown Petunias under Low Fertility Conditions

Abstract
:1. Introduction
2. Results
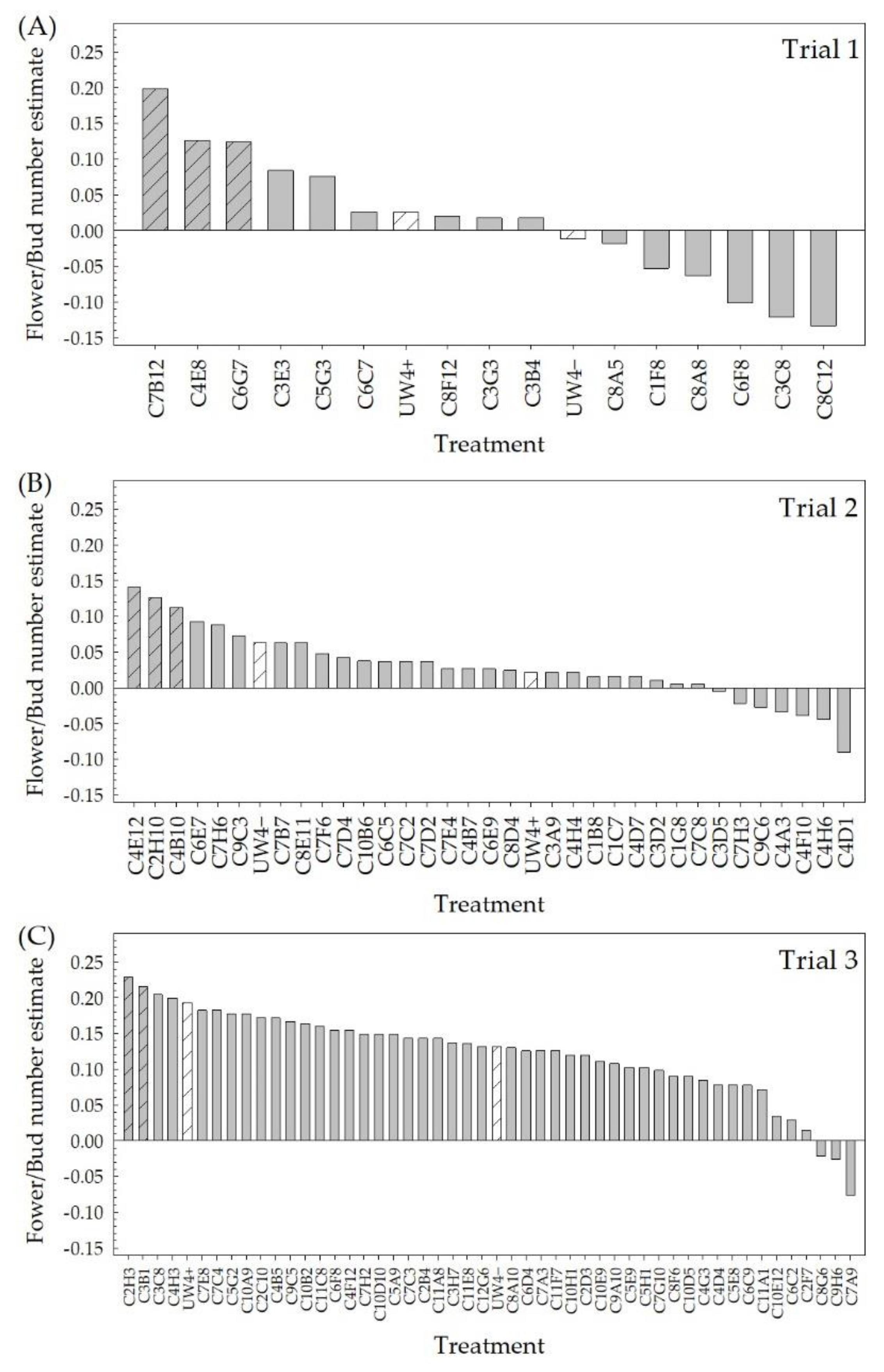
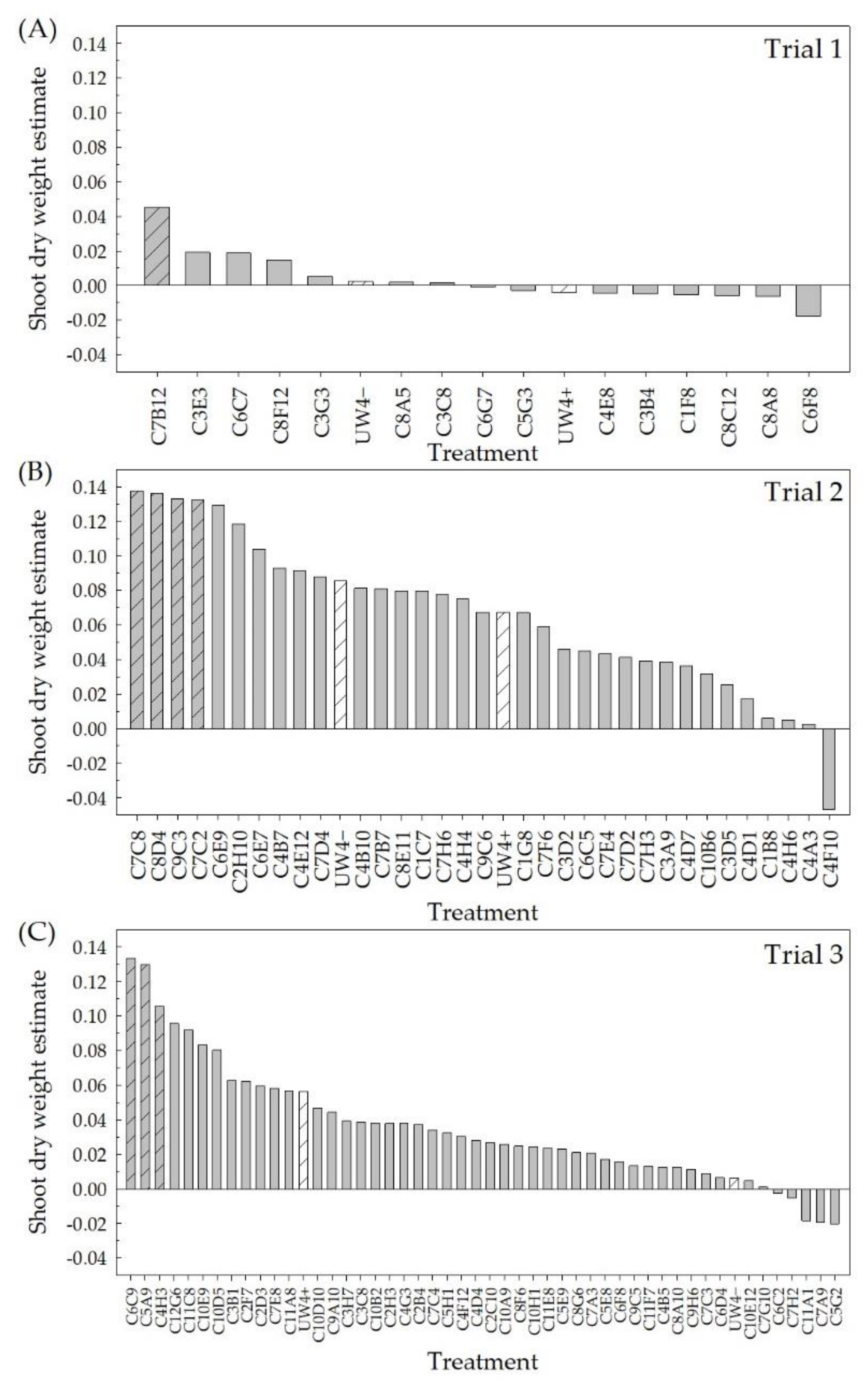
2.1. High-Throughput Greenhouse Trials
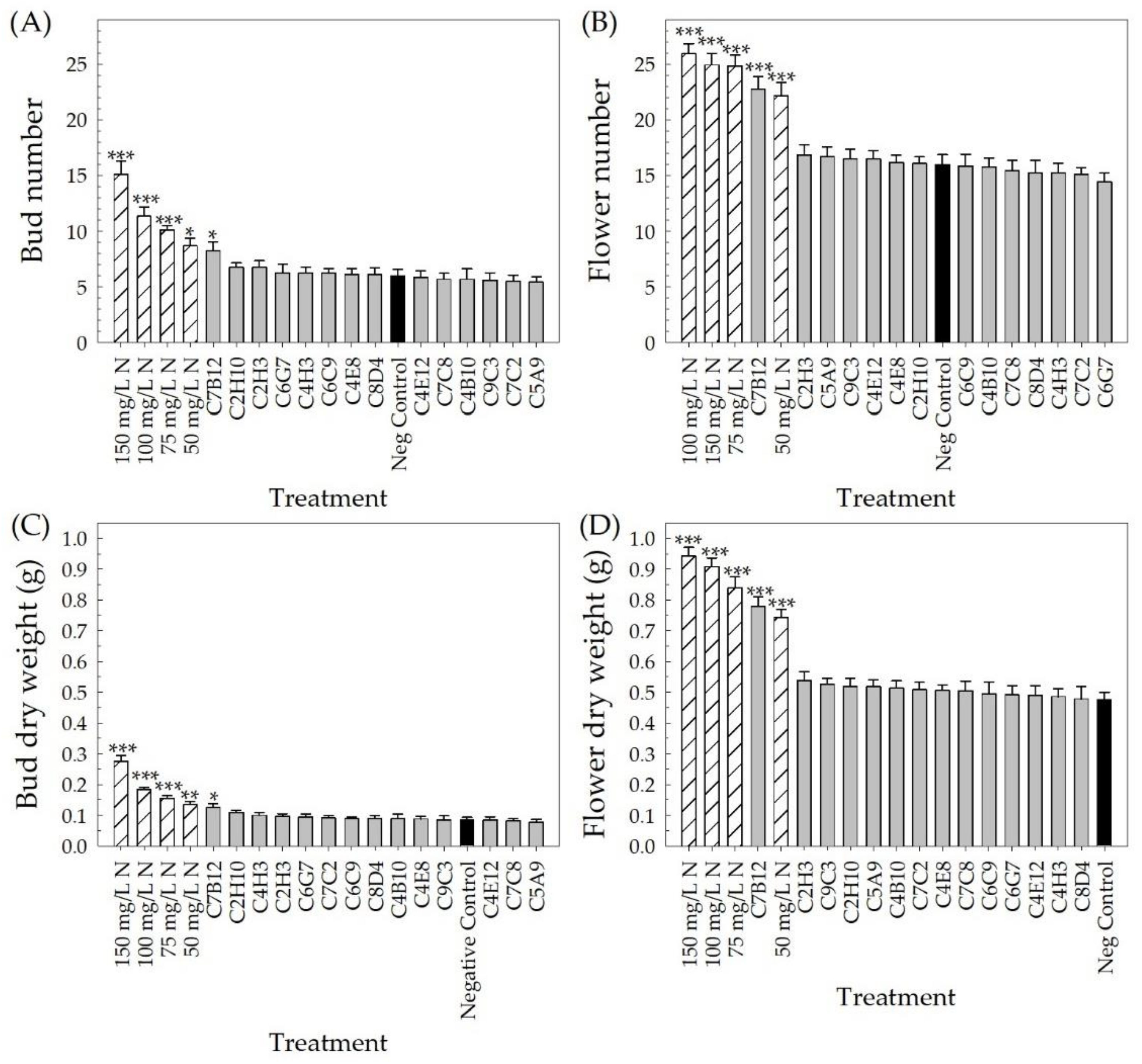
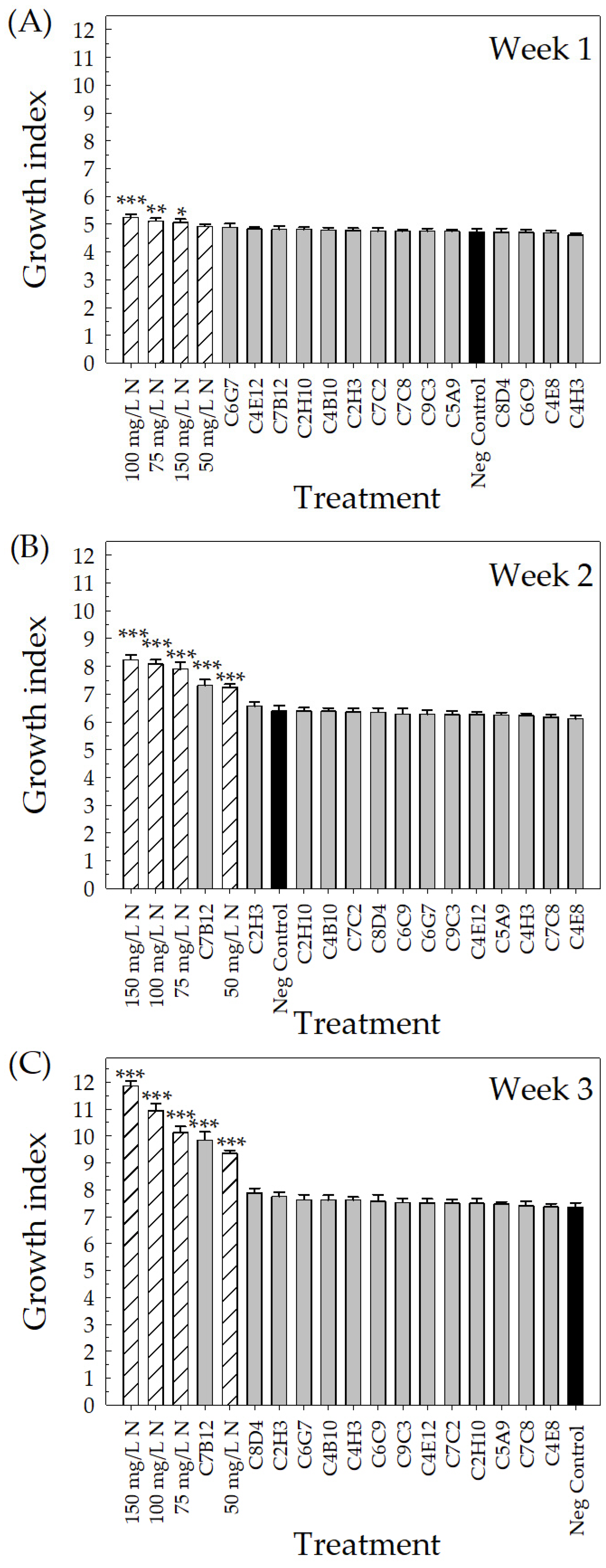
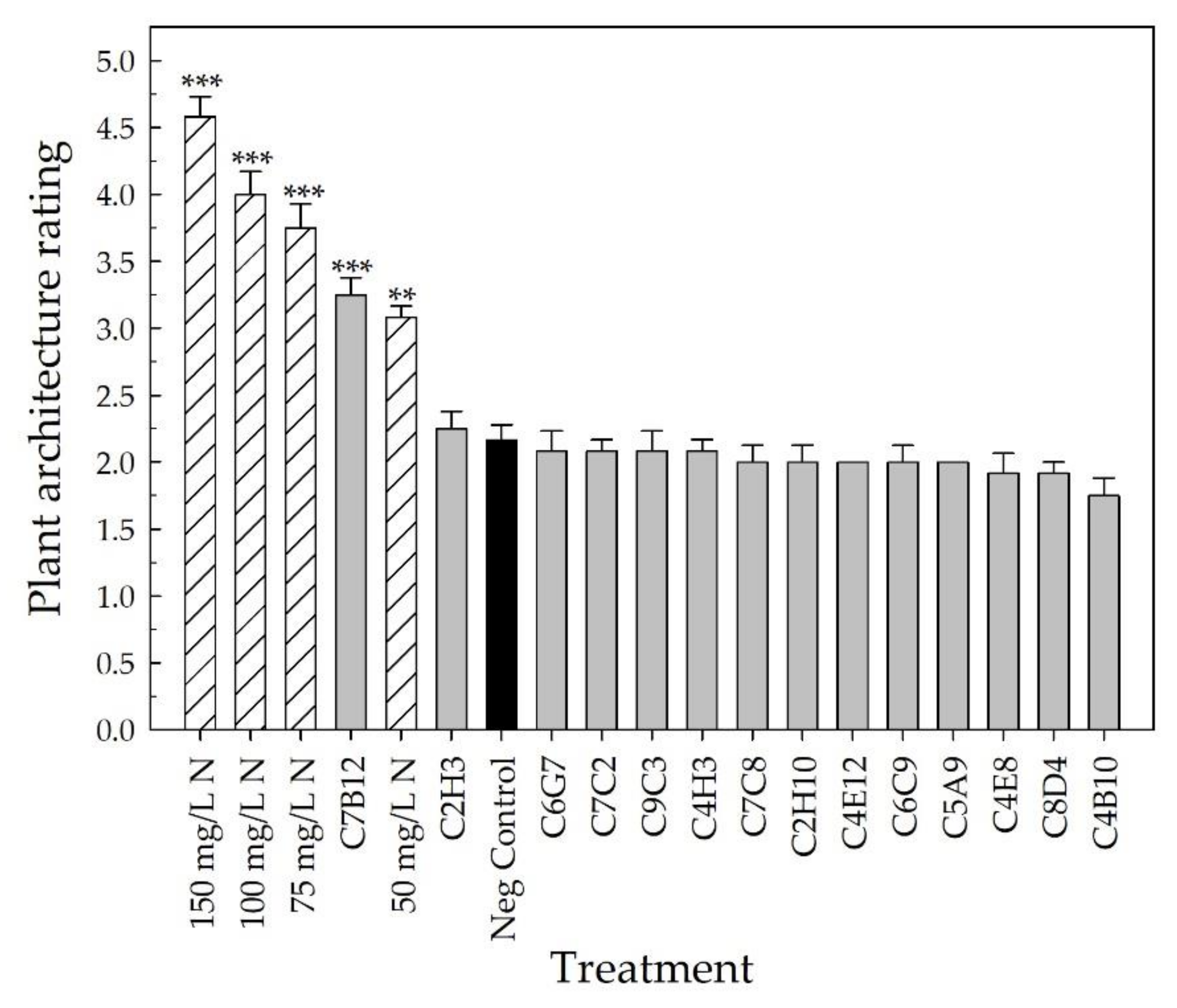
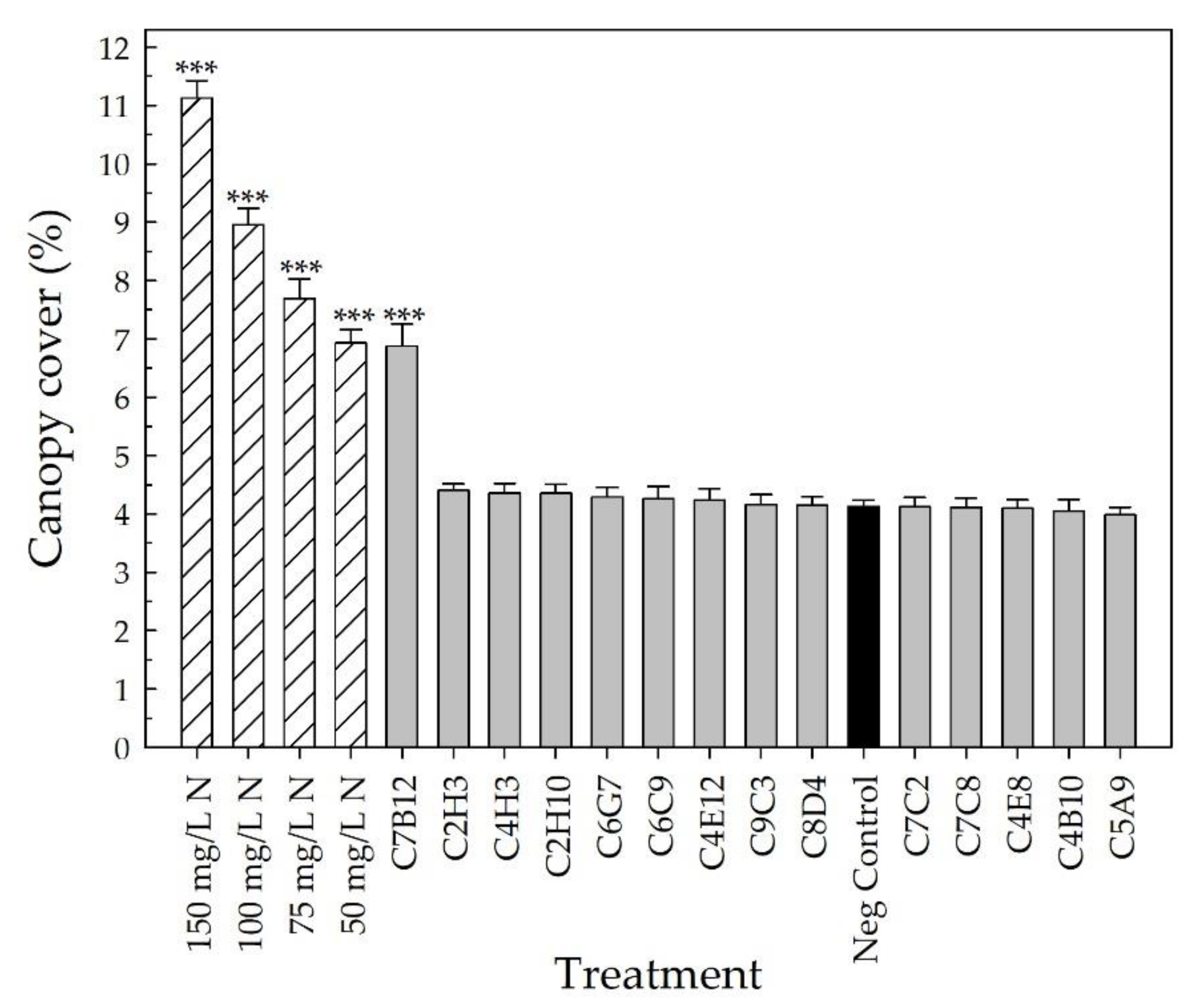
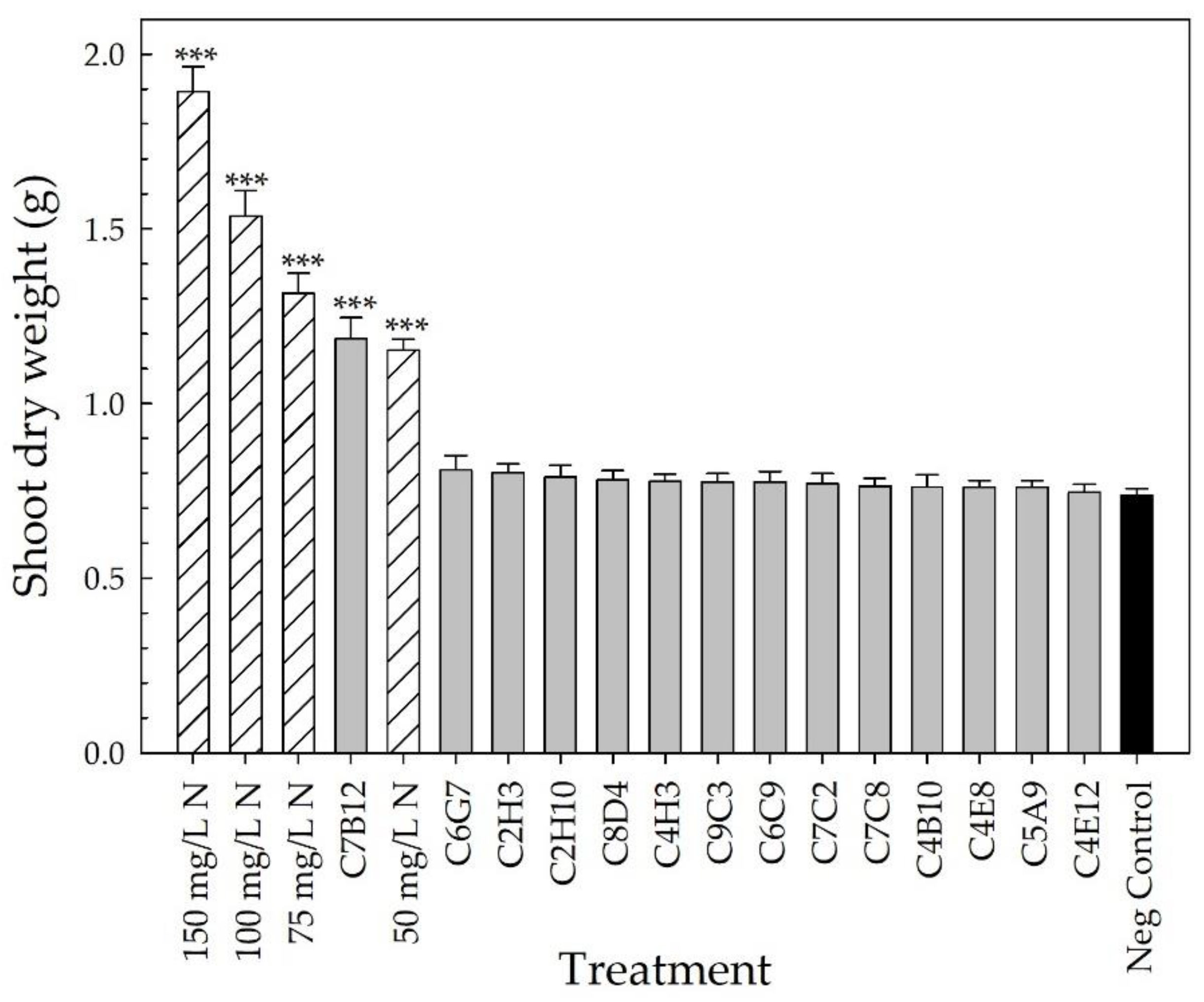
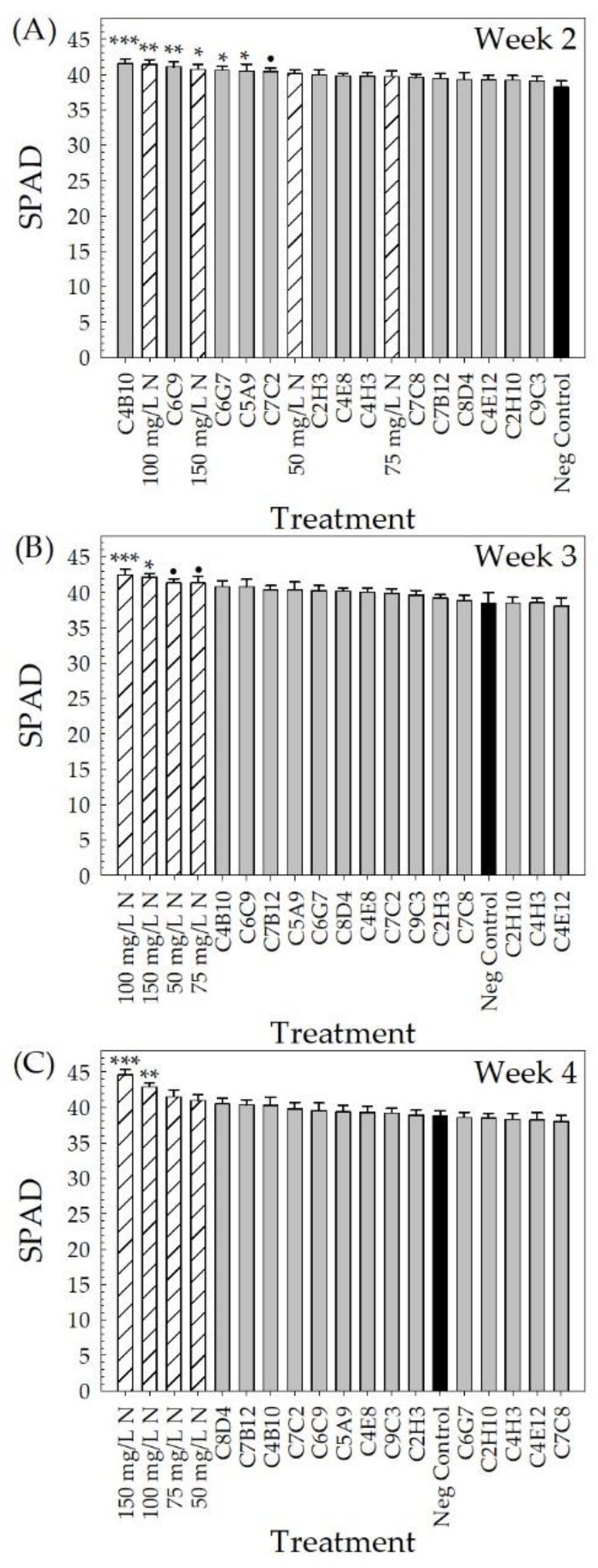
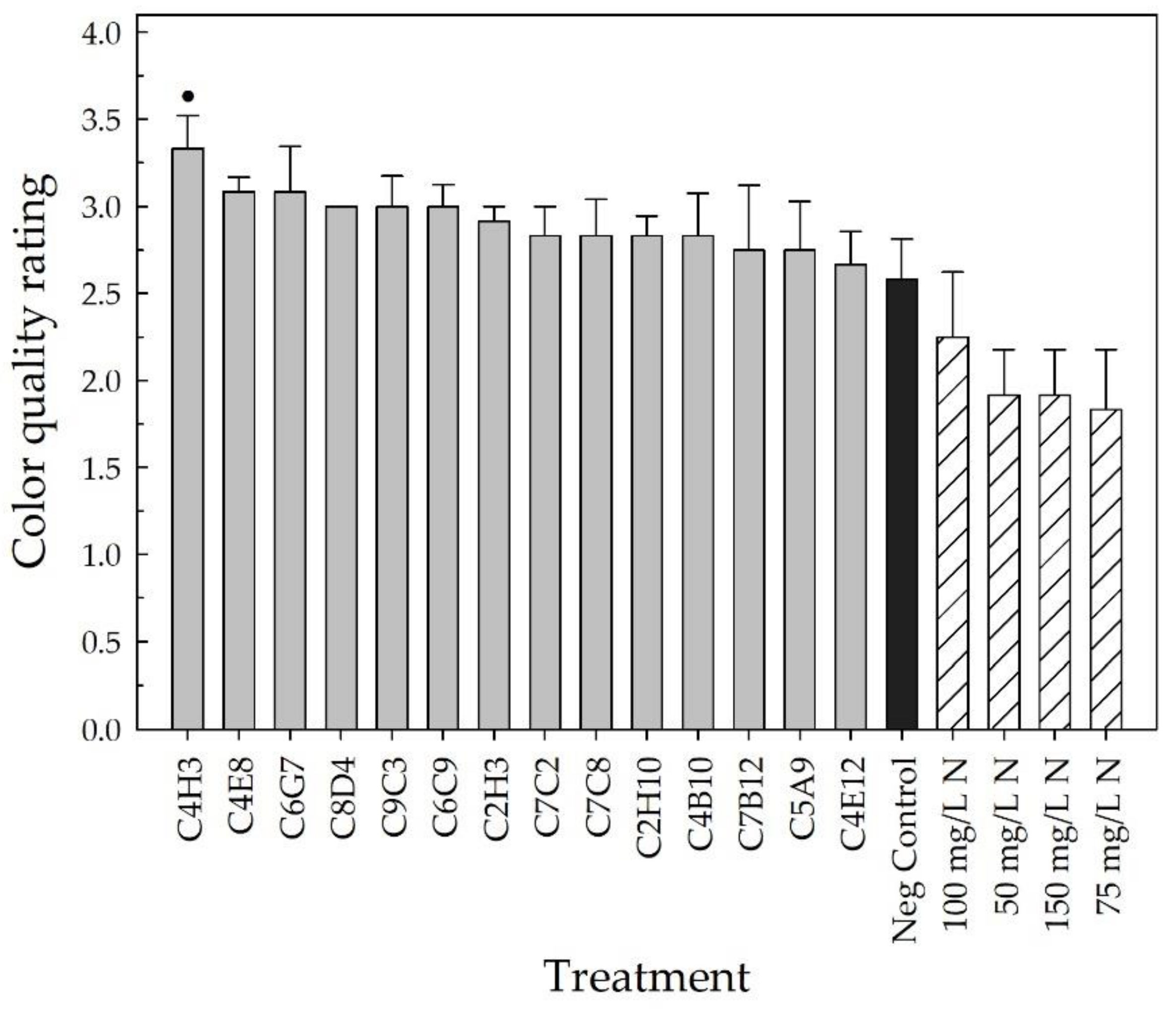
2.2. Greenhouse Validation Trial
2.3. Putative Plant-Growth-Promoting Mechanisms
3. Discussion
4. Materials and Methods
4.1. Bacteria Collections
4.2. High-Throughput Greenhouse Trials
4.2.1. Plant Material and Greenhouse Conditions
4.2.2. Experimental Design and Bacterial Application
4.2.3. Data Collection
4.3. Taxonomic Classification of Top-Performing Bacterial Isolates
4.4. Greenhouse Validation Trial with Low-Fertility Conditions
4.4.1. Plant Material and Greenhouse Conditions
4.4.2. Experimental Design and Bacterial Application
4.4.3. Measuring Plant Performance
4.5. Statistical Analysis
4.5.1. High-Throughput Greenhouse Trials
4.5.2. Greenhouse Validation Trial
4.6. Identification of Genes Involved in Putative Plant-Growth-Promoting Mechanisms
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, J.; Huang, Y.; Caldwell, R.D. Best management practices for minimizing nitrate leaching from container-grown nurseries. Sci. World J. 2001, 1, 96–102. [Google Scholar] [CrossRef] [Green Version]
- Paparozzi, E.T. Nutrition of floricultural crops: How far have we come? HortScience 2003, 38, 1031–1035. [Google Scholar] [CrossRef] [Green Version]
- Zaidi, A.; Khan, M.S.; Ahmad, E.; Saif, S.; Rizvi, A.; Shahid, M. Growth stimulation and management of diseases of ornamental plants using phosphate solubilizing microorganisms: Current perspective. Acta Physiol. Plant. 2016, 38, 1–21. [Google Scholar] [CrossRef]
- Daughtrey, M.L.; Benson, D.M. Principles of plant health management for ornamental plants. Annu. Rev. Phytopathol. 2005, 43, 141–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noordegraaf, C.V. Production and marketing of high qauality plants. Acta Hortic. 1994, 353, 134–148. [Google Scholar] [CrossRef]
- Wallach, R. Chapter 3. Physical characteristics of soilless media. In Soilless Culture: Theory and Practice; Elsevier: Amsterdam, The Netherlands, 2019; pp. 33–112. ISBN 9780444636966. [Google Scholar]
- Silber, A.; Bar-tal, A. Chapter 6. Nutrition of substrate-grown plants. In Soilless Culture: Theory and Practice; Elsevier: Amsterdam, The Netherlands, 2019; pp. 197–257. ISBN 9780444636966. [Google Scholar]
- Alsanius, B.W.; Wohanka, W. Chapter 5. Root zone microbiology of soilless cropping systems. In Soilless Culture: Theory and Practice; Elsevier: Amsterdam, The Netherlands, 2019; pp. 149–194. ISBN 9780444636966. [Google Scholar]
- Silber, A. Chapter 4. Chemical characteristics of soilless media. In Soilless Culture: Theory and Practice; Elsevier: Amsterdam, The Netherlands, 2019; pp. 113–148. ISBN 9780444636966. [Google Scholar]
- Lieth, J.H.; Oki, L.R. Chapter 9. Irrigation in Soilless Production. In Soilless Culture: Theory and Practice; Elsevier: Amsterdam, The Netherlands, 2019; pp. 379–424. ISBN 9780444636966. [Google Scholar]
- Jez, J.M.; Lee, S.G.; Sherp, A.M. The next green movement: Plant biology for the environment and sustainability. Science 2016, 353, 1241–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vox, G.; Teitel, M.; Pardossi, A.; Minuto, A.; Tinivella, F.; Schettini, E. Chapter 1: Sustainable greenhouse systems. In Sustainable Agriculture: Technology, Planning and Management; Salazar, A., Rios, I., Eds.; Nova Science Publishers, Inc.: New York, NY, USA, 2010; pp. 1–79. ISBN 978-1-60876-269-9. [Google Scholar]
- Barrett, G.E.; Alexander, P.D.; Robinson, J.S.; Bragg, N.C. Achieving environmentally sustainable growing media for soilless plant cultivation systems—A review. Sci. Hortic. 2016, 212, 220–234. [Google Scholar] [CrossRef] [Green Version]
- du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants application in horticultural crops under abiotic stress conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Hamid, B.; Zaman, M.; Farooq, S.; Fatima, S.; Sayyed, R.Z.; Baba, Z.A.; Sheikh, T.A.; Reddy, M.S.; Enshasy, H.E.; Gafur, A.; et al. Bacterial plant biostimulants: A sustainable way towards improving growth, productivity, and health of crops sustainability bacterial plant biostimulants: A sustainable way towards improving growth, productivity, and health of crops. Sustainability 2021, 13, 2856. [Google Scholar] [CrossRef]
- Adesemoye, A.O.; Torbert, H.A.; Kloepper, J.W. Plant growth-promoting rhizobacteria allow reduced application rates of chemical fertilizers. Microb. Ecol. 2009, 58, 921–929. [Google Scholar] [CrossRef]
- Yang, J.; Kloepper, J.W.; Ryu, C. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef]
- Dodd, I.C.; Ruiz-Lozano, J.M. Microbial enhancement of crop resource use efficiency. Curr. Opin. Biotechnol. 2012, 23, 236–242. [Google Scholar] [CrossRef]
- Sarki, M.S.; Panhwar, Q.A.; Ali, A.; Jamali, M.S.; Rajpar, I.; Depar, N. Assessment of potential bacterial isolates for enhancing plant nutrient uptake and growth of wheat. Malaysian J. Soil Sci. 2021, 25, 107–124. [Google Scholar]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orozco-Mosqueda, M.d.C.; Rocha-Granados, M.d.C.; Glick, B.R.; Santoyo, G. Microbiome engineering to improve biocontrol and plant growth-promoting mechanisms. Microbiol. Res. 2018, 208, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth-promoting rhizobacteria: Context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakim, S.; Naqqash, T.; Nawaz, M.S.; Laraib, I.; Siddique, M.J.; Zia, R.; Mirza, M.S.; Imran, A. Rhizosphere engineering with plant growth-promoting microorganisms for agricu; lture and ecological sustainability. Front. Sustain. Food Syst. 2021, 5. [Google Scholar] [CrossRef]
- Raviv, M.; Lieth, J.H.; Bar-tal, A. Chapter 14. Growing plants in soilless culture: Operational conclusions. In Soilless Culture: Theory and Practice; Elsevier: Amsterdam, The Netherlands, 2019; pp. 637–670. ISBN 9780444636966. [Google Scholar]
- Adesemoye, A.O.; Kloepper, J.W. Plant-microbes interactions in enhanced fertilizer-use efficiency. Appl. Microbiol. Biotechnol. 2009, 85, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Gadagi, R.S.; Krishnaraj, P.U.; Kulkarni, J.H.; Sa, T. The effect of combined Azospirillum inoculation and nitrogen fertilizer on plant growth promotion and yield response of the blanket flower Gaillardia pulchella. Sci. Hortic. 2004, 100, 323–332. [Google Scholar] [CrossRef]
- Chen, Y.P.; Rekha, P.D.; Arun, A.B.; Shen, F.T.; Lai, W.A.; Young, C.C. Phosphate solubilizing bacteria from subtropical soil and their tricalcium phosphate solubilizing abilities. Appl. Soil Ecol. 2006, 34, 33–41. [Google Scholar] [CrossRef]
- Susilowati, L.E.; Syekhfani, S. Characterization of phosphate solubilizing bacteria isolated from Pb contaminated soils and their potential for dissolving tricalcium phosphate. J. Degrad. Min. Lands Manag. 2014, 1, 57–62. [Google Scholar] [CrossRef]
- Gahan, J.; Schmalenberger, A. The role of bacteria and mycorrhiza in plant sulfur supply. Front. Plant Sci. 2014, 5, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Yang, Q.; Ge, K.; Hu, X.; Qi, G.; Du, B.; Liu, K.; Ding, Y. Promotion of iron nutrition and growth on peanut by Paenibacillus illinoisensis and Bacillus sp. strains in calcareous soil. Brazilian J. Microbiol. 2017, 48, 656–670. [Google Scholar] [CrossRef] [PubMed]
- Hafeez, B.; Khanif, Y.M.; Saleem, M. Role of zinc in plant nutrition- A review. Am. J. Exp. Agric. 2013, 3, 374–391. [Google Scholar] [CrossRef]
- Saravanan, V.S.; Kalaiarasan, P.; Madhaiyan, M.; Thangaraju, M. Solubilization of insoluble zinc compounds by Gluconacetobacter diazotrophicus and the detrimental action of zinc ion (Zn2+) and zinc chelates on root knot nematode Meloidogyne incognita. Lett. Appl. Microbiol. 2007, 44, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Goteti, P.K.; Emmanuel, L.D.A.; Desai, S.; Shaik, M.H.A. Prospective zinc solubilising bacteria for enhanced nutrient uptake and growth promotion in maize (Zea mays L.). Int. J. Microbiol. 2013, 2013, 869697. [Google Scholar] [CrossRef] [Green Version]
- Sirohi, G.; Upadhyay, A.; Shankar Srivastava, P.; Srivastava, S. PGPR mediated zinc biofertilization of soil and its impact on growth and productivity of wheat. J. Soil Sci. Plant Nutr. 2015, 15, 202–216. [Google Scholar] [CrossRef] [Green Version]
- Raiz, U.; Mehdi, S.M.; Iqbal, S.; Khalid, H.I.; Qadir, A.A.; Anum, W.; Ahmad, M.; Murtaza, G. Chapter 9: Bio-fertilizers: Eco-friendly approach for plant and soil environment. In Bioremediation and Biotechnology; Hakeem, K.R., Bhat, R.A., Qadri, H., Eds.; Springer Nature Switzerland AG: Cham, Switzerland, 2020; pp. 189–213. ISBN 9783030356903. [Google Scholar]
- Park, H.-G.; Lee, Y.-S.; Kim, K.-Y.; Park, Y.-S.; Park, K.-H.; Han, T.-H.; Park, C.-M.; Ahn, Y.-S. Inoculation with Bacillus licheniformis MH48 promotes nutrient uptake in seedlings of the ornamental plant Camellia Japonica grown in Korean reclaimed coastal lands. Korean J. Hortic. Sci. Technol. 2017, 35, 11–20. [Google Scholar] [CrossRef]
- Patten, C.L.; Glick, B.R. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl. Environ. Microbiol. 2002, 68, 3795–3801. [Google Scholar] [CrossRef] [Green Version]
- Glick, B.R.; Karaturovic, D.M.; Newell, P.C. A novel procedure for rapid isolation of plant growth promoting pseudomonads. Can. J. Microbiol. 1995, 41, 533–536. [Google Scholar] [CrossRef]
- Belimov, A.A.; Safronova, V.I.; Sergeyeva, T.A.; Egorova, T.N.; Matveyeva, V.A.; Tsyganov, V.E.; Borisov, A.Y.; Tikhonovich, I.A.; Kluge, C.; Preisfeld, A.; et al. Characterization of plant growth promoting rhizobacteria isolated from polluted soils and containing 1-aminocyclopropane-1-carboxylate deaminase. Can. J. Microbiol. 2001, 47, 642–652. [Google Scholar] [CrossRef]
- Nordstedt, N.P.; Chapin, L.J.; Taylor, C.G.; Jones, M.L. Identification of Pseudomonas spp. that increase ornamental crop quality during abiotic stress. Front. Plant Sci. 2020, 10, 1754. [Google Scholar] [CrossRef] [PubMed]
- Ghazi, A.; Attia, E. Effect of chemical, bio fertilization and compost tea spraying on growth and productivity of Zantedeschia bulbs. Sci. J. Flowers Ornam. Plants 2016, 3, 147–166. [Google Scholar] [CrossRef] [Green Version]
- Hoda, E.E.-M.; Mona, S. Effect of bio and chemical fertilizers on growth and flowering of Petunia hybrida plants. Am. J. Plant Physiol. 2014, 9, 68–77. [Google Scholar]
- Yasmin, S.; Hafeez, F.Y.; Schmid, M.; Hartmann, A. Plant-beneficial rhizobacteria for sustainable increased yield of cotton with reduced level of chemical fertilizers. Pakistan J. Bot. 2013, 45, 655–662. [Google Scholar]
- Nordstedt, N.P.; Jones, M.L. Isolation of rhizosphere bacteria that improve quality and water stress tolerance in greenhouse ornamentals. Front. Plant Sci. 2020, 11, 826. [Google Scholar] [CrossRef]
- Paulitz, T.C.; Bélanger, R.R. Biological control in greenhouse systems. Annu. Rev. Phytopathol. 2001, 39, 103–133. [Google Scholar] [CrossRef]
- Garima, G.; Nath, J.P. Screening of potential PGPR candidates as future biofertilizers-A strategic approach from lab to field. Res. J. Biotechnol. 2015, 10, 48–62. [Google Scholar]
- Egamberdiyeva, D. The effect of plant growth promoting bacteria on growth and nutrient uptake of maize in two different soils. Appl. Soil Ecol. 2007, 36, 184–189. [Google Scholar] [CrossRef]
- Koukounaras, A. Advanced greenhouse horticulture: New technologies and cultivation practices. Horticulturae 2021, 7, 1. [Google Scholar] [CrossRef]
- De Muynck, C.; Pereira, C.S.S.; Naessens, M.; Parmentier, S.; Soetaert, W.; Vandamme, E.J. The genus Gluconobacter oxydans: Comprehensive overview of biochemistry and biotechnological applications. Crit. Rev. Biotechnol. 2007, 27, 147–171. [Google Scholar] [CrossRef]
- Eida, A.A.; Bougouffa, S.; L’Haridon, F.; Alam, I.; Weisskopf, L.; Bajic, V.B.; Saad, M.M.; Hirt, H. Genome insights of the plant-growth promoting bacterium Cronobacter muytjensii JZ38 with volatile-mediated antagonistic activity against Phytophthora infestans. Front. Microbiol. 2020, 11, 369. [Google Scholar] [CrossRef] [Green Version]
- Uroz, S.; Oger, P. Caballeronia mineralivorans sp. nov., isolated from oak-Scleroderma citrinum mycorrhizosphere. Syst. Appl. Microbiol. 2017, 40, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Gyaneshwar, P.; Hirsch, A.M.; Moulin, L.; Chen, W.M.; Elliott, G.N.; Bontemps, C.; Santos, P.E.-D.L.; Gross, E.; Bueno dos Reis, F., Jr.; Sprent, J.I.; et al. Legume-nodulating betaproteobacteria: Diversity, host range, and future prospects. Mol. Plant-Microbe Interact. 2011, 24, 1276–1288. [Google Scholar] [CrossRef] [Green Version]
- Dobritsa, A.P.; Samadpour, M. Transfer of eleven species of the genus Burkholderia to the genus Paraburkholderia and proposal of Caballeronia gen. nov. to accommodate twelve species of the genera Burkholderia and Paraburkholderia. Int. J. Syst. Evol. Microbiol. 2016, 66, 2836–2846. [Google Scholar] [CrossRef] [PubMed]
- Puri, A.; Padda, K.P.; Chanway, C.P. In vitro and in vivo analyses of plant-growth-promoting potential of bacteria naturally associated with spruce trees growing on nutrient-poor soils. Appl. Soil Ecol. 2020, 149, 103538. [Google Scholar] [CrossRef]
- Lu, P.; Zheng, L.Q.; Sun, J.J.; Liu, H.M.; Li, S.P.; Hong, Q.; Li, W.-J. Burkholderia zhejiangensis sp. nov., a methyl-parathion-degrading bacterium isolated from a wastewater-treatment system. Int. J. Syst. Evol. Microbiol. 2012, 62, 1337–1341. [Google Scholar] [CrossRef]
- Popoca-Ursino, E.C.; Martínez-Ocampo, F.; Dantán-González, E.; Sánchez-Salinas, E.; Ortiz-Hernández, M.L. Characterization of methyl parathion degradation by a Burkholderia zhejiangensis strain, CEIB S4-3, isolated from agricultural soils. Biodegradation 2017, 28, 351–367. [Google Scholar] [CrossRef]
- Hernández-Mendoza, A.; Martínez-Ocampo, F.; Beltrán, L.F.L.A.; Popoca-Ursino, E.C.; Ortiz-Hernández, L.; Sánchez-Salinas, E.; Dantán-González, E. Draft genome sequence of the organophosphorus compound-degrading Burkholderia zhejiangensis strain CEIB S4-3. Genome Announc. 2014, 2, e01323-14. [Google Scholar] [CrossRef] [Green Version]
- Kamran, S.; Shahid, I.; Baig, D.N.; Rizwan, M.; Malik, K.A.; Mehnaz, S. Contribution of zinc solubilizing bacteria in growth promotion and zinc content of wheat. Front. Microbiol. 2017, 8, 2593. [Google Scholar] [CrossRef] [Green Version]
- Patel, P.; Trivedi, G.; Saraf, M. Iron biofortification in mungbean using siderophore producing plant growth promoting bacteria. Environ. Sustain. 2018, 1, 357–365. [Google Scholar] [CrossRef]
- Turan, M.; Gulluce, M.; Şahin, F. Effects of plant-prowth-promoting rhizobacteria on yield, growth, and some physiological characteristics of wheat and barley plants. Commun. Soil Sci. Plant Anal. 2012, 43, 1658–1673. [Google Scholar] [CrossRef]
- Hmaeid, N.; Wali, M.; Mahmoud, O.M.-B.; Pueyo, J.J.; Ghnaya, T.; Abdelly, C. Efficient rhizobacteria promote growth and alleviate NaCl-induced stress in the plant species Sulla carnosa. Appl. Soil Ecol. 2019, 133, 104–113. [Google Scholar] [CrossRef]
- Mishra, S.K.; Khan, M.H.; Misra, S.; Dixit, V.K.; Gupta, S.; Tiwari, S.; Gupta, S.C.; Chauhan, P.S. Drought tolerant Ochrobactrum sp. inoculation performs multiple roles in maintaining the homeostasis in Zea mays L. subjected to deficit water stress. Plant Physiol. Biochem. 2020, 150, 1–14. [Google Scholar] [CrossRef]
- Diez-Mendez, A.; Rivas, R. Improvement of saffron production using Curtobacterium herbarum as a bioinoculant under greenhouse conditions. AIMS Microbiol. 2017, 3, 354–364. [Google Scholar] [CrossRef]
- Saharan, B.S.; Nehra, V. Plant growth promoting rhizobacteria: A critical review. Life Sci. Med. Res. 2011, 21, 1–30. [Google Scholar]
- Kraiser, T.; Gras, D.E.; Gutiérrez, A.G.; González, B.; Gutiérrez, R.A. A holistic view of nitrogen acquisition in plants. J. Exp. Bot. 2011, 62, 1455–1466. [Google Scholar] [CrossRef] [PubMed]
- Selvakumar, G.; Kundu, S.; Joshi, P.; Nazim, S.; Gupta, A.D.; Mishra, P.K.; Gupta, H.S. Characterization of a cold-tolerant plant growth-promoting bacterium Pantoea dispersa 1A isolated from a sub-alpine soil in the North Western Indian Himalayas. World J. Microbiol. Biotechnol. 2008, 24, 955–960. [Google Scholar] [CrossRef]
- Imran, A.; Saadalla, M.J.A.; Khan, S.-U.; Mirza, M.S.; Malik, K.A.; Hafeez, F.Y. Ochrobactrum sp. Pv2Z2 exhibits multiple traits of plant growth promotion, biodegradation and N-acyl-homoserine-lactone quorum sensing. Ann. Microbiol. 2014, 64, 1797–1806. [Google Scholar] [CrossRef]
- Goldstein, A.H.; Liu, S.T. Molecular cloning and regulation of a mineral phosphate solubilizing gene from Erwinia herbicola. Biotechnology 1987, 5, 72–74. [Google Scholar] [CrossRef]
- Gyaneshwar, P.; Parekh, L.J.; Archana, G.; Poole, P.S.; Collins, M.D.; Hutson, R.A.; Kumar, G.N. Involvement of a phosphate starvation inducible glucose dehydrogenase in soil phosphate solubilization by Enterobacter asburiae. FEMS Microbiol. Lett. 1999, 171, 223–229. [Google Scholar] [CrossRef]
- Rodríguez, H.; Fraga, R.; Gonzalez, T.; Bashan, Y. Genetics of phosphate solubilization and its potential applications for improving plant growth-promoting bacteria. Plant Soil 2006, 287, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Wanner, B.L. Signal transduction in the control of phosphate-regulated genes of Escherichia coli. Kidney Int. 1996, 49, 964–967. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, Y.J.; Wanner, B.L. Global regulation by the seven-component Pi signaling system. Curr. Opin. Microbiol. 2010, 13, 198–203. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Fan, J.B.; Du, L.; Xu, H.; Zhang, Q.H.; He, Y.Q. The application of phosphate solubilizing endophyte Pantoea dispersa triggers the microbial community in red acidic soil. Appl. Soil Ecol. 2014, 84, 235–244. [Google Scholar] [CrossRef]
- Singh, N.P.; Singh, R.K.; Meena, V.S.; Meena, R.K. Can we use maize (Zea mays) rhizobacteria as plant growth promoter? Vegetos 2015, 28, 86–99. [Google Scholar] [CrossRef]
- Wan, W.; Qin, Y.; Wu, H.; Zuo, W.; He, H.; Tan, J.; Wang, Y.; He, D. Isolation and characterization of phosphorus solubilizing bacteria with multiple phosphorus sources utilizing capability and their potential for lead immobilization in soil. Front. Microbiol. 2020, 11, 752. [Google Scholar] [CrossRef] [Green Version]
- Grayston, S.; Germida, J. Influence of crop rhizospheres on populations and activity of heterotrophic sulfur-oxidizing microorganisms. Soil Biol. Biochem. 1990, 22, 457–463. [Google Scholar] [CrossRef]
- Grayston, S.J.; Germida, J.J. Sulfur-oxidizing bacteria as plant growth promoting rhizobacteria for canola. Can. J. Microbiol. 1991, 37, 521–529. [Google Scholar] [CrossRef]
- Mahmood, Q.; Hu, B.; Cai, J.; Zheng, P.; Azim, M.R.; Jilani, G.; Islam, E. Isolation of Ochrobactrum sp. QZ2 from sulfide and nitrite treatment system. J. Hazard. Mater. 2009, 165, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Du, Y.; Hu, L.; Xu, J.; Long, Y.; Shen, D. Effects of sulfur-metabolizing bacterial community diversity on H2S emission behavior in landfills with different operation modes. Biodegradation 2016, 27, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Boyce, A.N. Role of plant growth promoting rhizobacteria in agricultural sustainability-A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef] [PubMed]
- Vacheron, J.; Desbrosses, G.; Bouffaud, M.L.; Touraine, B.; Moënne-Loccoz, Y.; Muller, D.; Legendre, L.; Wisniewski-Dyé, F.; Prigent-Combaret, C. Plant growth-promoting rhizobacteria and root system functioning. Front. Plant Sci. 2013, 4, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Panwar, M.; Tewari, R.; Gulati, A.; Nayyar, H. Indigenous salt-tolerant rhizobacterium Pantoea dispersa (PSB3) reduces sodium uptake and mitigates the effects of salt stress on growth and yield of chickpea. Acta Physiol. Plant. 2016, 38, 1–12. [Google Scholar] [CrossRef]
- Kulkarni, G.B.; Nayak, A.S.; Sajjan, S.S.; Oblesha, A.; Karegoudar, T.B. Indole-3-acetic acid biosynthetic pathway and aromatic amino acid aminotransferase activities in Pantoea dispersa strain GPK. Lett. Appl. Microbiol. 2013, 56, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Saleh-Lakha, S.; Glick, B.R. The effect of native and ACC deaminase-containing Azospirillum brasilense Cd1843 on the rooting of carnation cuttings. Can. J. Microbiol. 2005, 51, 511–514. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Penrose, D.M.; Glick, B.R. Strategies used by rhizobia to lower plant ethylene levels and increase nodulation. Can. J. Microbiol. 2002, 48, 947–954. [Google Scholar] [CrossRef]
- Shah, S.; Li, J.; Moffatt, B.A.; Glick, B.R. Isolation and characterization of ACC deaminase genes from two different plant growth-promoting rhizobacteria. Can. J. Microbiol. 1998, 44, 833–843. [Google Scholar] [CrossRef]
- Jaya, D.K.; Giyanto Nurhidayat, N.; Antonius, S. Isolation, identification, and detection of ACC deaminase gene-encoding rhizobacteria from rhizosphere of stressed pineapple. Indones. J. Biotechnol. 2019, 24, 17–25. [Google Scholar] [CrossRef]
- Bullock, D.G.; Anderson, D.S. Evaluation of the Minolta SPAD-502 chlorophyll meter for nitrogen management in corn. J. Plant Nutr. 1998, 21, 741–755. [Google Scholar] [CrossRef]
- Choi, S.T.; Park, D.S.; Kang, S.M.; Park, S.J. Use of a chlorophyll meter to diagnose nitrogen status of “Fuyu” persimmon leaves. HortScience 2011, 46, 821–824. [Google Scholar] [CrossRef] [Green Version]
- Zhalnina, K.; Louie, K.; Hao, Z.; Mansoori, N.; da Rocha, U.; Shi, S.; Cho, H.; Karaoz, U.; Loque, D.; Bowen, B.; et al. Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly. Nat. Microbiol. 2018, 3, 470–480. [Google Scholar] [CrossRef] [Green Version]
- de Freitas, J.R. Yield and N assimilation of winter wheat (Triticum aestivum L., var. Norstar) inoculated with rhizobacteria. Pedobiologia 2000, 44, 97–104. [Google Scholar] [CrossRef]
- Morel, M.A.; Cagide, C.; Minteguiaga, M.A.; Dardanelli, M.S.; Castro-Sowinski, S. The pattern of secreted molecules during the co-inoculation of alfalfa plants with Sinorhizobium meliloti and Delftia sp. strain JD2: An interaction that improves plant yield. Mol. Plant-Microbe Interact. 2015, 28, 134–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Güneş, A.; Turan, M.; Güllüce, M.; Şahin, F. Nutritional content analysis of plant growth-promoting rhizobacteria species. Eur. J. Soil Biol. 2014, 60, 88–97. [Google Scholar] [CrossRef]
- Monteiro, R.A.; Balsanelli, E.; Wassem, R.; Marin, A.M.; Brusamarello-Santos, L.C.C.; Schmidt, M.A.; Tadra-Sfeir, M.Z.; Pankievicz, V.C.S.; Cruz, L.M.; Chubatsu, L.S.; et al. Herbaspirillum-plant interactions: Microscopical, histological and molecular aspects. Plant Soil 2012, 356, 175–196. [Google Scholar] [CrossRef]
- Elbeltagy, A.; Nishioka, K.; Sato, T.; Suzuki, H.; Ye, B.; Hamada, T.; Isawa, T.; Mitsui, H.; Minamisawa, K. Endophytic colonization and in planta nitrogen fixation by a Herbaspirillum sp.isolated from wild rice species. Appl. Environ. Microbiol. 2001, 67, 5285–5293. [Google Scholar] [CrossRef] [Green Version]
- Estrada, G.A.; Baldani, V.L.D.; de Oliveira, D.M.; Urquiaga, S.; Baldani, J.I. Selection of phosphate-solubilizing diazotrophic Herbaspirillum and Burkholderia strains and their effect on rice crop yield and nutrient uptake. Plant Soil 2013, 369, 115–129. [Google Scholar] [CrossRef]
- Subhashini, D.V.; Anuradha, M.; Damodar Reddy, D.; Vasanthi, J. Development of bioconsortia for optimizing nutrient supplementation through microbes for sustainable tobacco production. Int. J. Plant Prod. 2016, 10, 479–490. [Google Scholar] [CrossRef]
- Park, J.; Faust, J.E. Fertilization strategy affects production and postproduction performance of petunia. Horttechnology 2021, 31, 217–224. [Google Scholar] [CrossRef]
- Ball, V. Ball Redbook, 16th ed.; Ball, V., Ed.; Ball Publishing: Batavia, IL, USA, 1998; ISBN 1-883052-15-7. [Google Scholar]
- Li, J.; Ovakim, D.H.; Charles, T.C.; Glick, B.R. An ACC deaminase minus mutant of Enterobacter cloacae UW4 no longer promotes root elongation. Curr. Microbiol. 2000, 41, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Park, E.; Glick, B.R. 1-Aminocyclopropane-1-carboxylate deaminase from Pseudomonas putida UW4 facilitates the growth of canola in the presence of salt. Can. J. Microbiol. 2007, 53, 912–918. [Google Scholar] [CrossRef]
- Hontzeas, N.; Zoidakis, J.; Glick, B.R.; Abu-Omar, M.M. Expression and characterization of 1-aminocyclopropane-1-carboxylate deaminase from the rhizobacterium Pseudomonas putida UW4: A key enzyme in bacterial plant growth promotion. Biochim. Biophys. Acta 2004, 1703, 11–19. [Google Scholar] [CrossRef]
- Hontzeas, N.; Saleh, S.S.; Glick, B.R. Changes in gene expression in canola roots induced by ACC-deaminase- containing plant-growth-promoting bacteria. Mol. Plant-Microbe Interact. 2004, 17, 865–871. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-R, L.M.; Gunturu, S.; Harvey, W.T.; Rosselló-Mora, R.; Tiedje, J.M.; Cole, J.R.; Konstantinidis, K.T. The Microbial Genomes Atlas (MiGA) webserver: Taxonomic and gene diversity analysis of Archaea and Bacteria at the whole genome level. Nucleic Acids Res. 2018, 46, W282–W288. [Google Scholar] [CrossRef]
- Konstantinidis, K.T.; Tiedje, J.M. Genomic insights that advance the species definition for prokaryotes. Proc. Natl. Acad. Sci. USA 2005, 102, 2567–2572. [Google Scholar] [CrossRef] [Green Version]
- Niu, G.; Rodriguez, D.S.; Starman, T. Response of bedding plants to saline water irrigation. HortScience 2010, 45, 628–636. [Google Scholar] [CrossRef] [Green Version]
- Patrignani, A.; Ochsner, T.E. Canopeo: A powerful new tool for measuring fractional green canopy cover. Agron. J. 2015, 107, 2312–2320. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. 2019. Available online: https://www.R-project.org/ (accessed on 17 June 2021).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 3, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogle, D.H.; Wheeler, P.; Dinno, A. FSA: Fisheries stock analysis. R Packag. Version 2017, 17, 636. [Google Scholar]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter Selected for | ||||||
|---|---|---|---|---|---|---|
| Bacterial Strain | Taxonomic Classification | Accession No. 1 | Plant Origin 2 | GH Facility Origin 3 | Flower/Bud Number 4 | Shoot DW 4 |
| C7B12 | Caballeronia zhejiangensis | SAMN14930932 | Plectranthus scutellarioides ‘Electric Lime’ | 1 | X | X |
| C4E8 | Pseudarthrobacter equi | SAMN14930920 | Petunia × hybrida ‘Peppy Blue’ | 2 | X | |
| C6G7 | Raoultella terrigena | SAMN14930926 | Plectranthus scutellarioides ‘Vino’ | 3 | X | |
| C4E12 | Pseudomonas putida | SAMN14930924 | Plectranthus scutellarioides ‘Vino’ | 4 | X | |
| C2H10 | Curtobacterium sp. | SAMN14930927 | Plectranthus scutellarioides ‘Kingswood Torch’ | 5 | X | |
| C4B10 | Pseudomonas putida | SAMN14930922 | Catharanthus roseus ‘Titan Lilac’ | 2 | X | |
| C7C8 | Herbaspirillum sp. | SAMN14930928 | Plectranthus scutellarioides ‘Dark Star’ | 1 | X | |
| C8D4 | Pseudomonas corrugata | SAMN14930921 | Plectranthus scutellarioides ‘Fishnet Stockings’ | 6 | X | |
| C9C3 | Herbaspirillum sp. | SAMN14854762 | Plectranthus scutellarioides ‘Colorblaze Marooned’ | 7 | X | |
| C7C2 | Herbaspirillum sp. | SAMN14930931 | Zinnia elegans ‘Magellan’ | 8 | X | |
| C2H3 | Pantoea dispersa | SAMN14930933 | Zinnia elegans ‘Dreamland Coral’ | 9 | X | |
| C3B1 | Unknown | NA | Plectranthus scutellarioides ‘Wild Lime’ | 5 | X | |
| C6C9 | Ochrobactrum sp. | SAMN14930923 | Plectranthus scutellarioides ‘Vino’ | 3 | X | |
| C5A9 | Microbacterium sp. | SAMN14930925 | Pelargonium × hortorum ‘Calliope Dark Red’ | 9 | X | |
| C4H3 | Pseudomonas oryzihabitans | SAMN14930929 | Plectranthus scutellarioides ‘Colorblaze Apple Brandy’ | 2 | X | |
| Flowering 1 | Vegetative Growth | Vegetative Quality | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strain | Bud | Flower | Bud DW (g) | Flower DW (g) | GI 2 Week 2 | GI Week 3 | Canopy Cover 3 | Plant Archt. 4 | Shoot DW 1 (g) | SPAD 5 Week 2 | SPAD Week 3 | SPAD Week 4 | Color Rating 6 | Overall Score 7 |
| C7B12 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 11 | ||
| C2H3 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 9 | ||||
| C4H3 | 1 | 1 | 1 | 1 | 1 | 5 | ||||||||
| C2H10 | 1 | 1 | 1 | 1 | 1 | 5 | ||||||||
| C6G7 | 1 | 1 | 1 | 1 | 1 | 5 | ||||||||
| C6C9 | 1 | 1 | 1 | 1 | 4 | |||||||||
| C4B10 | 1 | 1 | 1 | 3 | ||||||||||
| C5A9 | 1 | 1 | 1 | 3 | ||||||||||
| C8D4 | 1 | 1 | 1 | 3 | ||||||||||
| C9C3 | 1 | 1 | 1 | 3 | ||||||||||
| C7C2 | 1 | 1 | 2 | |||||||||||
| C4E12 | 1 | 1 | ||||||||||||
| C4E8 | 1 | 1 | ||||||||||||
| C7C8 | 0 | |||||||||||||
| Bacterial Strains 1 | ||||||||
|---|---|---|---|---|---|---|---|---|
| Putative Plant Growth Promotion Property | Gene Function | Gene Name | C7B12 | C2H3 | C4H3 | C2H10 | C6G7 | C6C9 |
| Nitrogen metabolism and transport | Ammonium transport | amtB-glnK | x | x | x | x | ||
| Nitrate and nitrite transport | nrtABC | |||||||
| Nitrate and nitrite transport | narK | x | x | |||||
| Nitrogen metabolism | glnGL | x | x | x | ||||
| Nitrite reduction | nasDEF | |||||||
| Nitrogenase enzyme | nifDHK | |||||||
| Phosphate solubilization and transport | Pyrroloquinoline quinone synthase | pqq | x | x | x | x | ||
| Glucose dehydrogenase | gdh | x | x | x | x | |||
| High affinity phosphate transporter system | pstABCS | x | x | x | x | |||
| Sulfur metabolism and transport | Sulfate ABC transporter | cysPWAT-sbp | x | x | x | |||
| Sulfur metabolism | cysND | x | x | x | x | x | ||
| Sulfur metabolism | cysC | x | x | |||||
| Sulfur metabolism | cysHIJ | x | x | |||||
| Siderophore production (BGCs 2) | Enterobactin synthesis | x | ||||||
| Ornibactin synthesis | x | |||||||
| Turnerbactin synthesis | x | x | ||||||
| Desferrioxamine synthesis | x | |||||||
| Pyoverdin synthesis | x | |||||||
| Zinc solubilization and transport | Glyoxylate/hydroxypyruvate reductase B | ghrB | x | x | x | x | x | |
| High affinity ABC zinc transporter system | znuABC | x | x | x | ||||
| Zinc export | zitB | x | x | x | x | |||
| Zinc export | zntABR | x | x | x | ||||
| IAA 3 synthesis | Trp 4 biosynthesis operon | trpABCDE | x | x | x | x | ||
| Trp-specific importer | mtr | x | x | |||||
| Indole-3-pyruvate decarboxylase | ipdC | x | x | |||||
| Conversion of indole-3-acetaldehyde to IAA | aldAB | |||||||
| Conversion of Trp to IAA | iaaHM | |||||||
| ACC 5 degradation | ACC deaminase structural gene | acdS | x | x | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
South, K.A.; Nordstedt, N.P.; Jones, M.L. Identification of Plant Growth Promoting Rhizobacteria That Improve the Performance of Greenhouse-Grown Petunias under Low Fertility Conditions. Plants 2021, 10, 1410. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10071410
South KA, Nordstedt NP, Jones ML. Identification of Plant Growth Promoting Rhizobacteria That Improve the Performance of Greenhouse-Grown Petunias under Low Fertility Conditions. Plants. 2021; 10(7):1410. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10071410
Chicago/Turabian StyleSouth, Kaylee A., Nathan P. Nordstedt, and Michelle L. Jones. 2021. "Identification of Plant Growth Promoting Rhizobacteria That Improve the Performance of Greenhouse-Grown Petunias under Low Fertility Conditions" Plants 10, no. 7: 1410. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10071410