
Improving Restoration Efficiency by Modeling Optimal Sowing Periods: A Case Study of Two Native Plants and Restoration of a Site

Abstract
:1. Introduction
2. Results
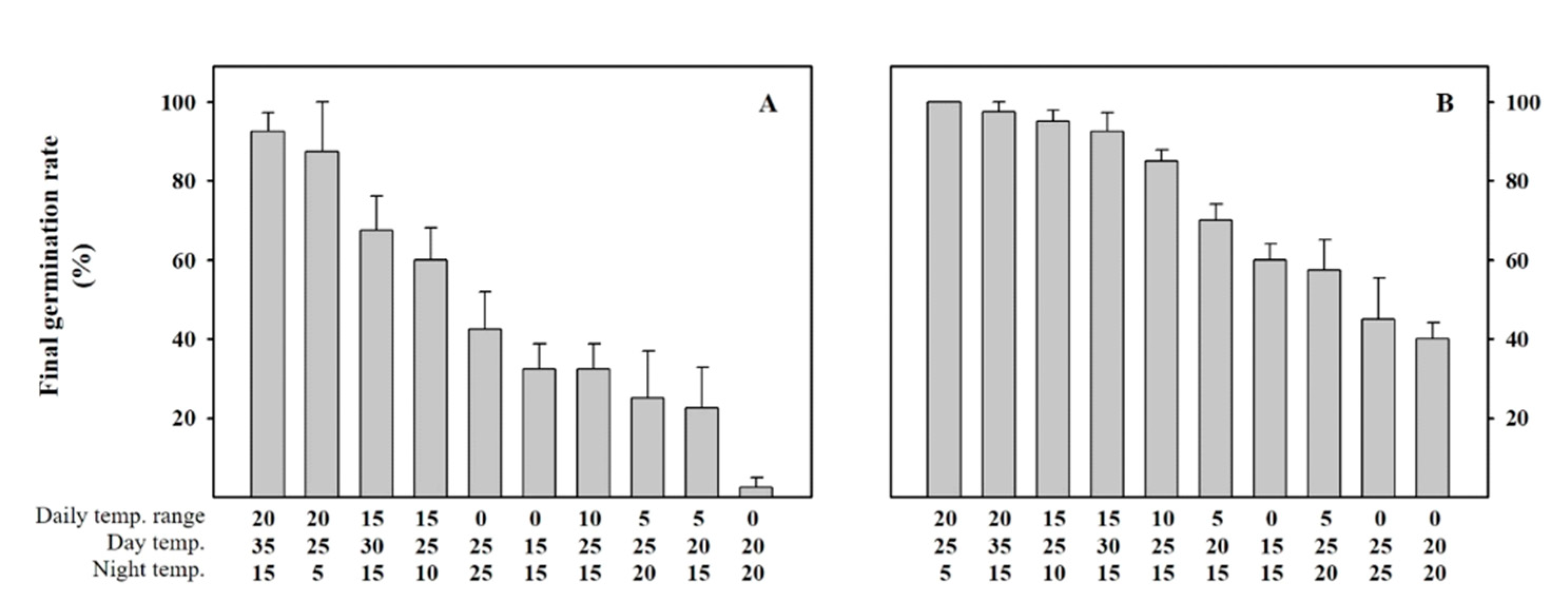
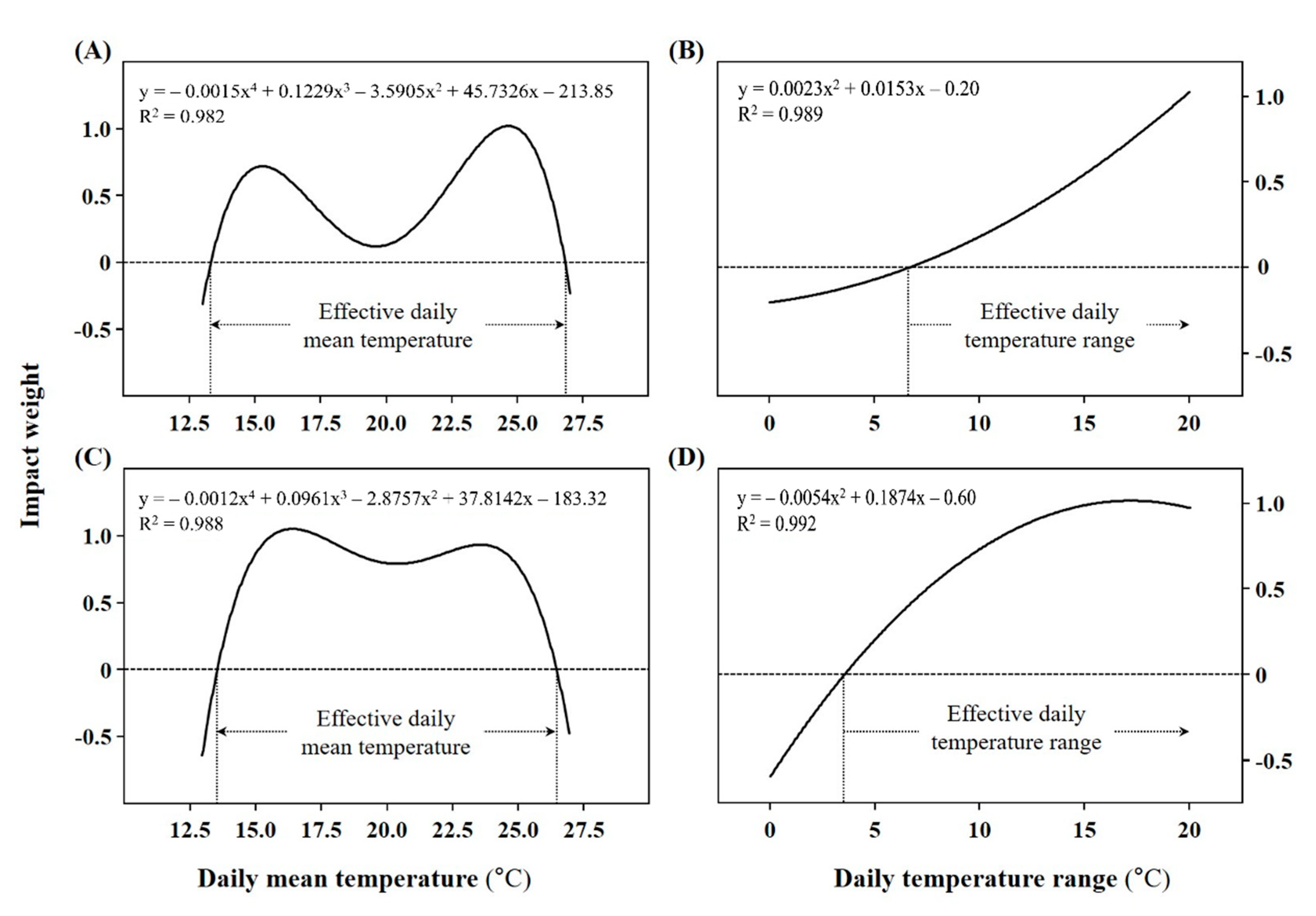
2.1. Customized Germination Model for A. rugosa and A. rubra Seeds
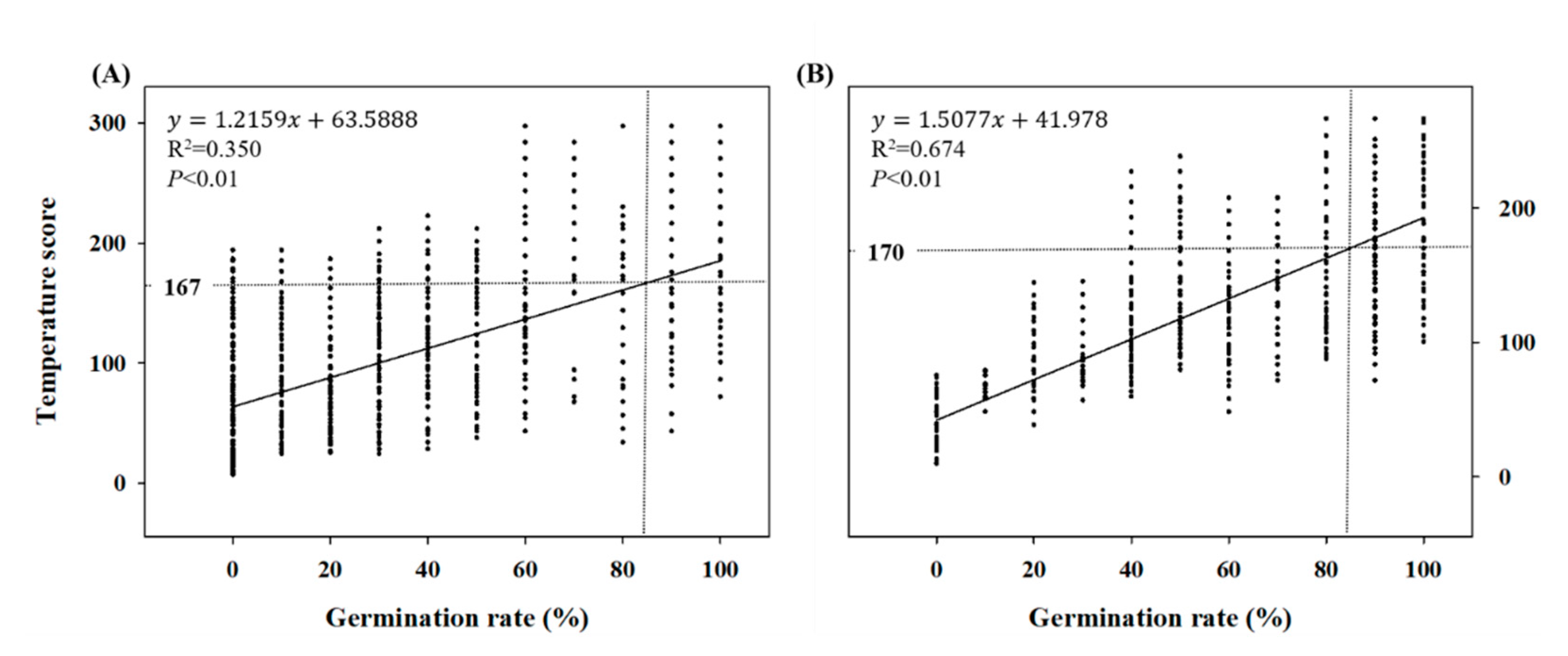
2.2. Temperature Requirement for a Final Germination Percentage of 85%
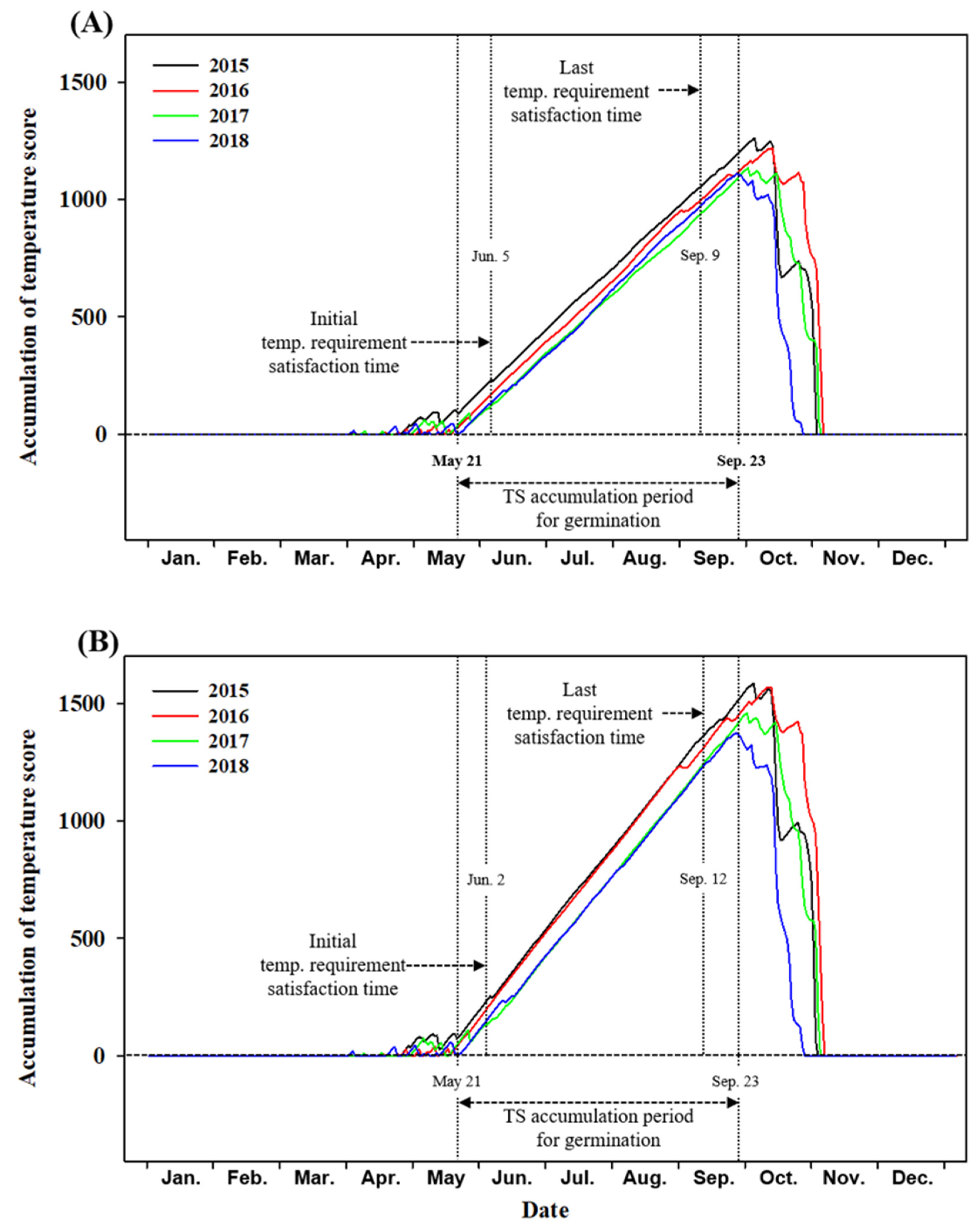
2.3. Prediction of the Adaptation of A. rugosa and A. rubra to the Mt. Gariwang Site
3. Discussion
4. Materials and Methods
4.1. Seeds and Experimental Conditions
4.2. Species-Specific Seed Vigor Test
4.3. Application of Species-Specific Seed Vigor Test for Regional Climate Data
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Paik, W.K.; Park, W.G.; Lee, W.T. Flora and vegetation of resources plants in the Mt. Kariwang (Kangwon-do). Korean J. Plant Resour. 1998, 11, 217–243. [Google Scholar]
- Forest Service. Preservation and Restoration Methods of Genetic Resources Conservation Zone in Mt. Gariwangsan; Forest Service: Daejeon, Korea, 2012. [Google Scholar]
- Park, J.K. Necessity to reflect notification of topsoil erosion investigation and monitoring using high-resolution image data. Geoenvironmental. Eng. 2018, 19, 3–7. [Google Scholar]
- Gibson, A.L.; Espeland, E.K.; Wagner, V.; Nelson, C.R. Can local adaptation research in plants inform selection of native plant materials? An analysis of experimental methodologies. Evol. Appl. 2016, 9, 1219–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Abascal, I.; Tárrega, R.; Luis-Calabuig, E.; Marcos, E. Effects of sowing native herbaceous species on the post-fire recovery in a heathland. Acta Oecol. 2003, 24, 131–138. [Google Scholar] [CrossRef]
- Keeley, J.E. Ecological impacts of wheat seeding after a Sierra Nevada wildfire. Int. J. Wildland Fire 2004, 13, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Lee, J.; Jeon, K.; Kim, H.; Choi, Y.; Jung, D.; Song, H. Native plants selection for ecological replantation in forest road slope. J. Korean Soc. Environ. Restor. Technol. 2003, 6, 24–32. [Google Scholar]
- Kim, N.C.; Nam, U.J.; Shin, K.J. A study on the slope ecological restoration models for the Baekdu-Mountain Range. J. Korean Soc. Environ. Restor. Technol. 2008, 11, 72–84. [Google Scholar]
- Reza Yousefi, A.; Rashidi, S.; Moradi, P.; Mastinu, A. Germination and seedling growth responses of Zygophyllum fabago, Salsola kali L. and Atriplex canescens to PEG-induced drought stress. Environments 2020, 7, 107. [Google Scholar] [CrossRef]
- Aghajanlou, F.; Mirdavoudi, H.; Shojaee, M.; Mac Sweeney, E.; Mastinu, A.; Moradi, P. Rangeland management and ecological adaptation analysis model for Astragalus curvirostris Boiss. Horticulturae 2021, 7, 67. [Google Scholar] [CrossRef]
- Bellairs, S.; Bell, D.T. Temperature effects on the seed-germination of 10 Kwongan species from Eneabba, Western-Australia. Aust. J. Bot. 1990, 38, 451–458. [Google Scholar] [CrossRef]
- Cony, M.; Trione, S.O. Germination with respect to temperature of two Argentinian Prosopis species. J. Arid Environ. 1996, 33, 225–236. [Google Scholar] [CrossRef]
- Dahal, P.; Bradford, K.J.; Jones, R.A. Effects of priming and endosperm integrity on seed germination rates of tomato genotypes: I. Germination at suboptimal temperature. J. Exp. Bot. 1990, 41, 1431–1439. [Google Scholar] [CrossRef]
- Steinmaus, S.J.; Prather, T.S.; Holt, J.S. Estimation of base temperatures for nine weed species. J. Exp. Bot. 2000, 51, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Tan, M.; Liao, F.; Hou, L.; Wang, J.; Wei, L.; Jian, H.; Xu, X.; Li, J.; Liu, L. Genome-wide association analysis of seed germination percentage and germination index in Brassica napus L. under salt and drought stresses. Euphytica 2017, 213, 40. [Google Scholar] [CrossRef]
- Roby, K.B. Watershed Response and Recovery from the Will Fire: Ten Years of Observations. In Proceedings of the Symposium on Fire and Watershed Management, Sacramento, CA, USA, 26–28 October 1988; US Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: Berkeley, CA, USA, 1989; Volume 109, pp. 131–136. [Google Scholar]
- Schoennagel, T.L.; Waller, D.M. Understory responses to fire and artificial seeding in an eastern Cascades Abies grandis forest, USA. Can. J. For. Res. 1999, 29, 1393–1401. [Google Scholar] [CrossRef]
- Na, C.S.; Baek, S.G.; Yang, S.Y.; Park, C.Y.; Kim, J.H.; Lee, M.H.; Park, Y.S. Species-specific seed vigor test of aging chive for restoration and regional adaptation under climate change. Appl. Ecol. Environ. Res. 2013, 18, 683–695. [Google Scholar] [CrossRef]
- Kebreab, E.; Murdoch, A.J. A model of the effects of a wide range of constant and alternating temperatures on seed germination of four Orobanche species. Ann. Bot. 1999, 84, 549–557. [Google Scholar] [CrossRef] [Green Version]
- Ryu, J.H.; Baek, W.J.; Choi, S.Y.; Lee, K.S. Studies on the cultivation and use of Agastache rugosa Kuntze. Bull. Agric. Coll. Chonbuk Natl. Univ. 1998, 29, 70–79. [Google Scholar]
- Jang, B.K.; Cho, J.S.; Lee, C.H. Effect of environmental conditions and chemical treatments on seed germination of Astilbe koreana (Kom.) Nakai. Korean J. Plant Resour. 2016, 29, 235–240. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.W.; Lee, I.Y. Germination characteristic of Cat’s-Ear (Hypochaeris radicata). Weed Turfgrass Sci. 2019, 8, 179–183. [Google Scholar]
- Lee, Y.H.; Sohn, S.I.; Hong, S.H.; Kim, C.S.; Na, C.S.; Oh, Y.J. Ecology and control of speen amaranth (Amaranths patulus) in soybean. Weed Turfgrass Sci. 2019, 8, 377–384. [Google Scholar]
- Diniz, F.O.; Chamma, L.; Novembre, A.D.L.C. Germination of Physalis peruviana L. seeds under varying conditions of temperature, light, and substrate. Rev. Cien. Agron. 2020, 51, 1–9. [Google Scholar] [CrossRef]
- Nilsson, C.; Aradottir, A.L.; Hagen, D.; Halldórsson, G.; Høegh, K.; Mitchell, R.J.; Raulund-Rasmussen, K.; Svavarsdóttir, K.; Tolvanen, A.; Wilson, S.D. Evaluating the process of ecological restoration. Ecol. Soc. 2016, 21, 41. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.S.; An, T.J.; Kim, Y.I.; Lee, J.H.; Choi, E.J.; Kim, Y.G.; Chang, J.K. Growth characteristics and dry matter productivity of Saffower by sowing time. Korean J. Med. Crop. Sci. 2021, 29, 124–134. [Google Scholar] [CrossRef]
- Lee, S.W.; Kim, J.B.; Kim, K.S.; Kim, M.S. Changes of growth characteristics, rosmarinic acid and essential oil contents according to harvest time in Agastache rugosa O. Kuntze. Korean J. Med. Crop. Sci. 1999, 7, 83–88. [Google Scholar]
- Hong, M.J.; Kim, J.H.; Kim, H.Y.; Kim, M.J.; Kim, S.M. Chemical composition and biological activity of essential oil of Agastache rugosa (Fisch. & C. A. Mey.) O. Kuntze. Korean J. Med. Crop. Sci. 2020, 28, 95–110. [Google Scholar]
- Korea National Arboretum. Korea Biodiversity Information System. Available online: http://www.nature.go.kr/kbi/plant/pilbk/selectPlantPilbkDtl.do (accessed on 10 January 2020).
- Newton, R.; Hay, F.; Probert, R. Protocol for Comparative Seed Longevity Testing. In Technical Information Sheet 01 2009; Royal Botanic Gardens: Kew, London, UK, 2009. [Google Scholar]
- Korea Meteorological Administration. KMA Weather Data Service. Available online: https://data.kma.go.kr/resources/html/en/aowdp.html (accessed on 10 January 2020).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature (°C) | Agastache rugosa | Astilbe rubra | |||
|---|---|---|---|---|---|
| Standardized Regression Coefficient | Impact Weight (Ratio) | Standardized Regression Coefficient | Impact Weight (Ratio) | ||
| Daily mean temperature (DMT) | 15.0 | 0.488 ** | 0.71 | 0.685 ** | 0.86 |
| 17.5 | 0.257 ** | 0.37 | 0.799 ** | 1.00 | |
| 20.0 | 0.089 * | 0.13 | 0.640 ** | 0.80 | |
| 22.5 | 0.421 ** | 0.61 | 0.712 ** | 0.89 | |
| 25.0 | 0.689 ** | 1.00 | 0.629 ** | 0.79 | |
| Daily temperature range (DTR) | 0 | −0.157 ** | −0.17 | −0.525 ** | −0.60 |
| 5 | −0.140 ** | −0.15 | 0.179 * | 0.20 | |
| 10 | 0.199 ** | 0.21 | 0.635 ** | 0.72 | |
| 15 | 0.538 ** | 0.58 | −0.880 ** | 1.00 | |
| 20 | 0.929 ** | 1.00 | −0.850 ** | 0.96 | |
| Species | Variable | Standardized Regression Coefficient | Equation | p-Value | R2 | |
|---|---|---|---|---|---|---|
| Agastache rugosa | Impact weight of | daily mean temperature (x1) | 0.409 | 2.948·x1 + 4.041·x2 + 7.299 | <0.001 | 0.841 |
| daily temperature range (x2) | 0.678 | |||||
| Astilbe rubra | Impact weight of | daily mean temperature (x3) | 0.613 | 3.584·x3 + 1.679·x4 + 7.816 | <0.001 | 0.751 |
| daily temperature range (x4) | 0.466 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, Y.-H.; Kim, J.; Park, C.-Y.; Park, H.-S.; Park, Y. Improving Restoration Efficiency by Modeling Optimal Sowing Periods: A Case Study of Two Native Plants and Restoration of a Site. Plants 2021, 10, 1506. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10081506
Jung Y-H, Kim J, Park C-Y, Park H-S, Park Y. Improving Restoration Efficiency by Modeling Optimal Sowing Periods: A Case Study of Two Native Plants and Restoration of a Site. Plants. 2021; 10(8):1506. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10081506
Chicago/Turabian StyleJung, Young-Ho, JunHyeok Kim, Chung-Youl Park, Hee-Seung Park, and YoSup Park. 2021. "Improving Restoration Efficiency by Modeling Optimal Sowing Periods: A Case Study of Two Native Plants and Restoration of a Site" Plants 10, no. 8: 1506. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10081506