
Supporting Physiological Trait for Indirect Selection for Grain Yield in Drought-Stressed Popcorn

 ,
,  ,
,  ,
,  , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
2.1. Effects of Different Water Conditions on Dependent (GY and PE) and Morphological Variables and on SPAD and NDVI Values
2.2. Multiple Regressions Based on Morphological Characters and Dependent Variables GY and PE
2.3. Multiple Regressions Based on the SPAD Index of Different Measurement Dates and the Dependent Variables GY and PE
2.4. Multiple Regressions Based on NDVI Values of Different Measurement Dates and on the Dependent Variables GY and PE
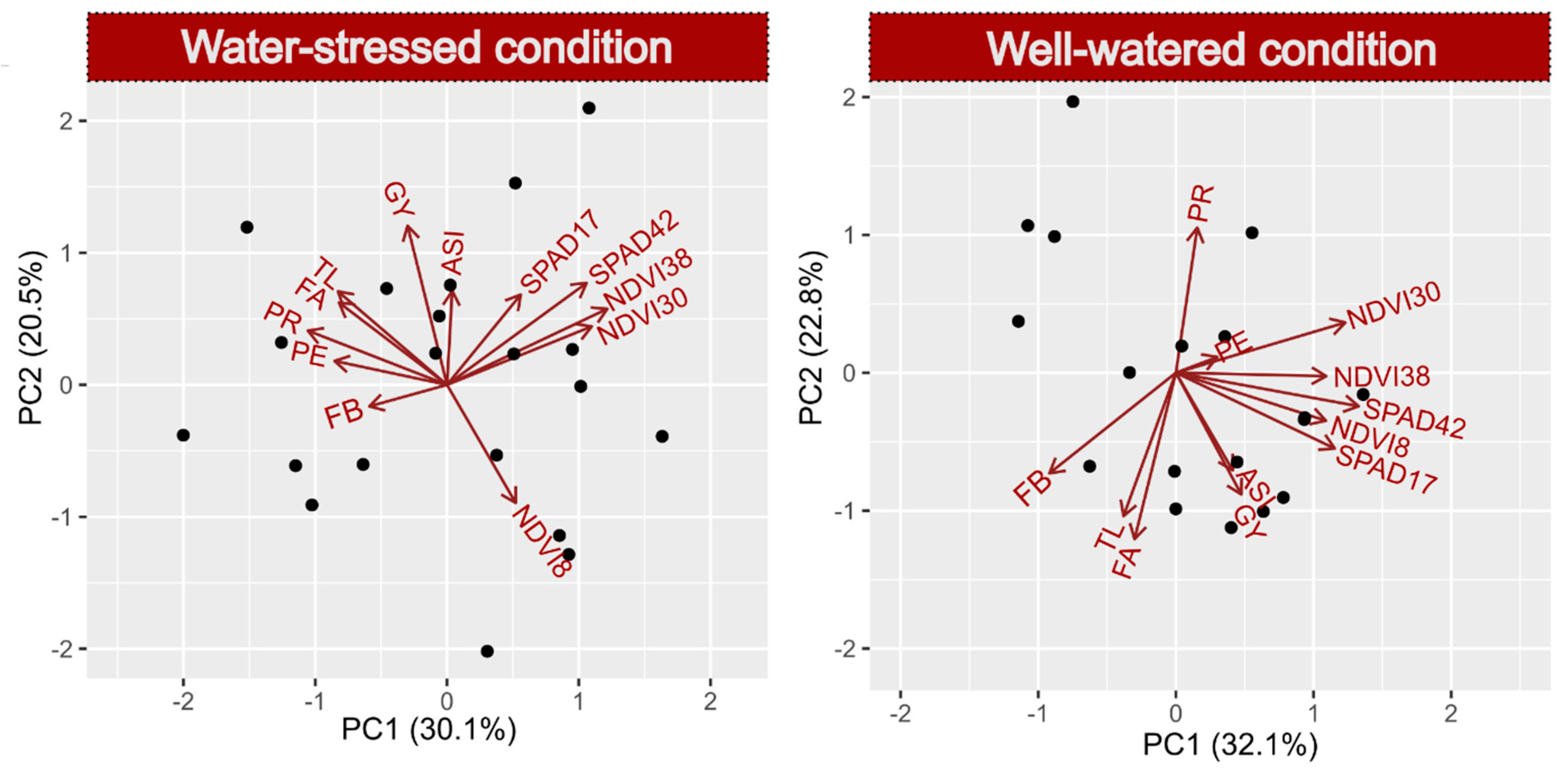
2.5. Biplot Using Significant Traits on Response Variables GY and PE
2.6. Selection of Predictors by MLR Based on Pre-Selected Variables
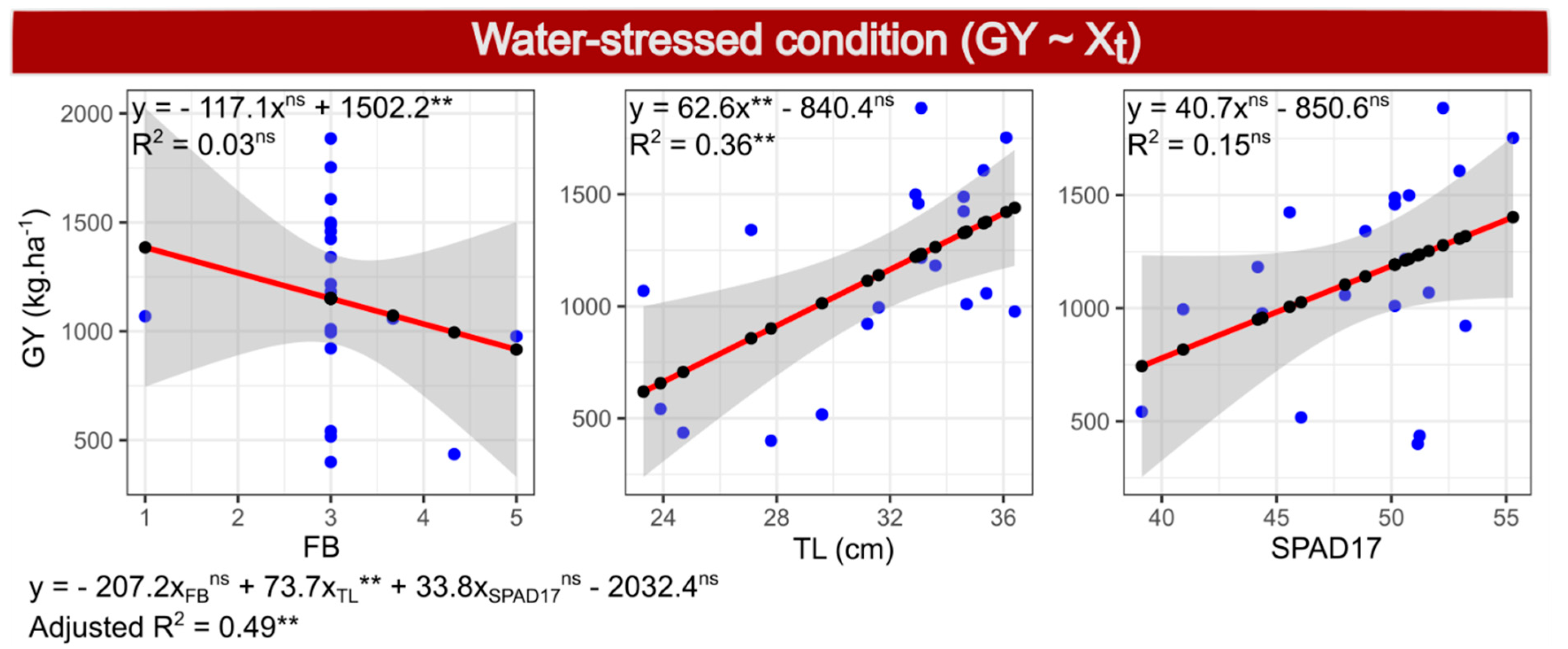
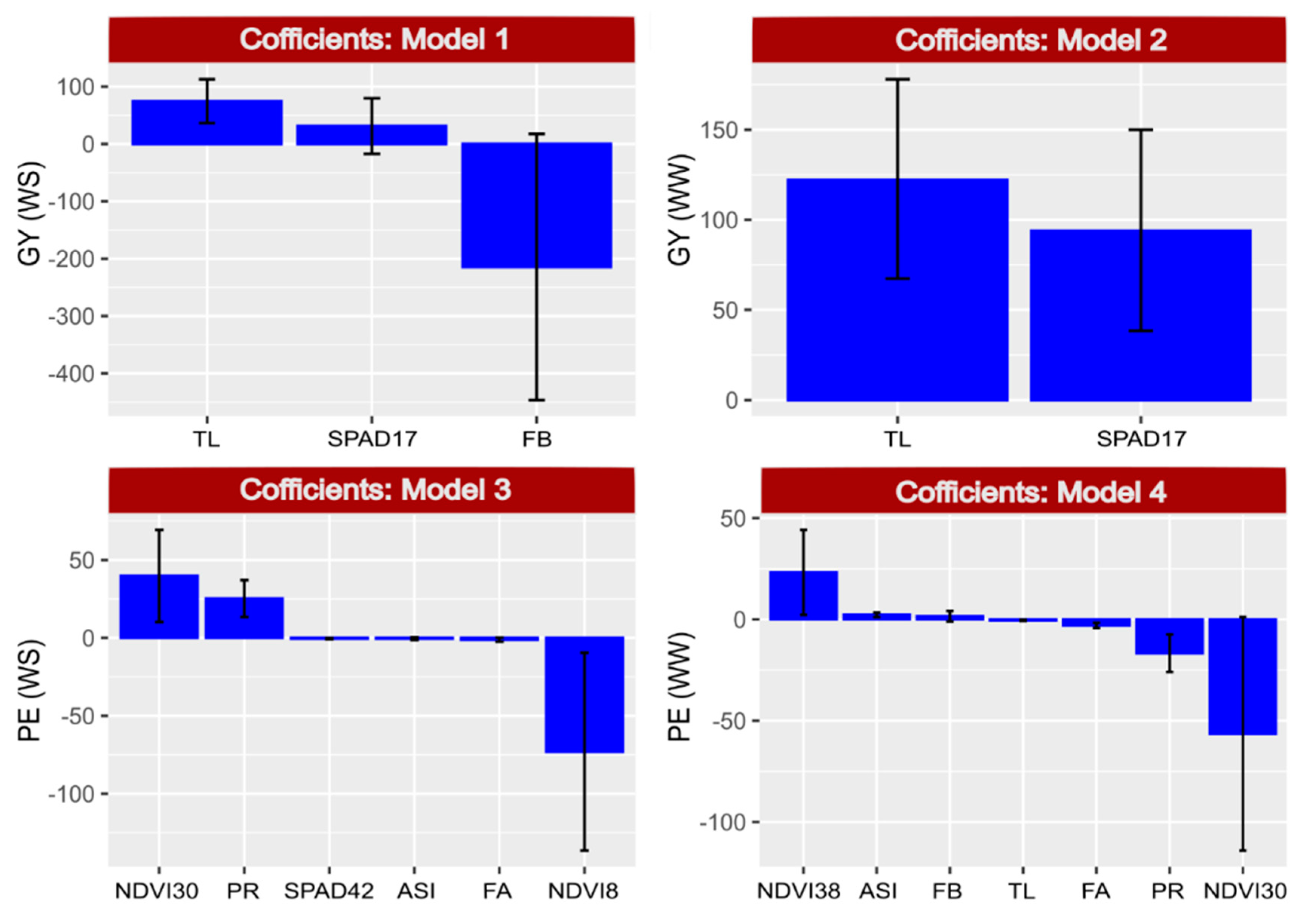
2.7. Selection of Model 1—GY under WS
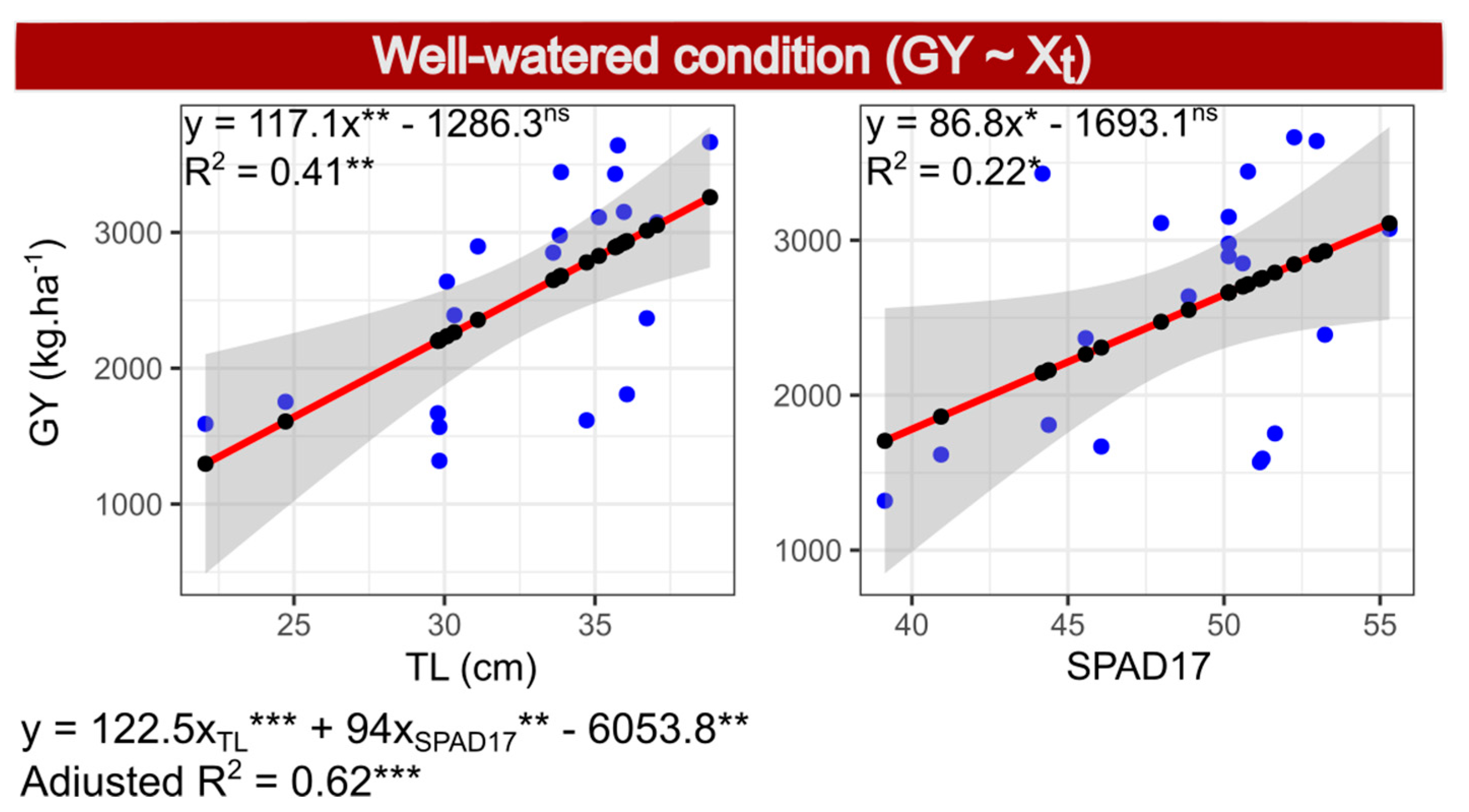
2.8. Selection of Model 2—GY under WW Conditions
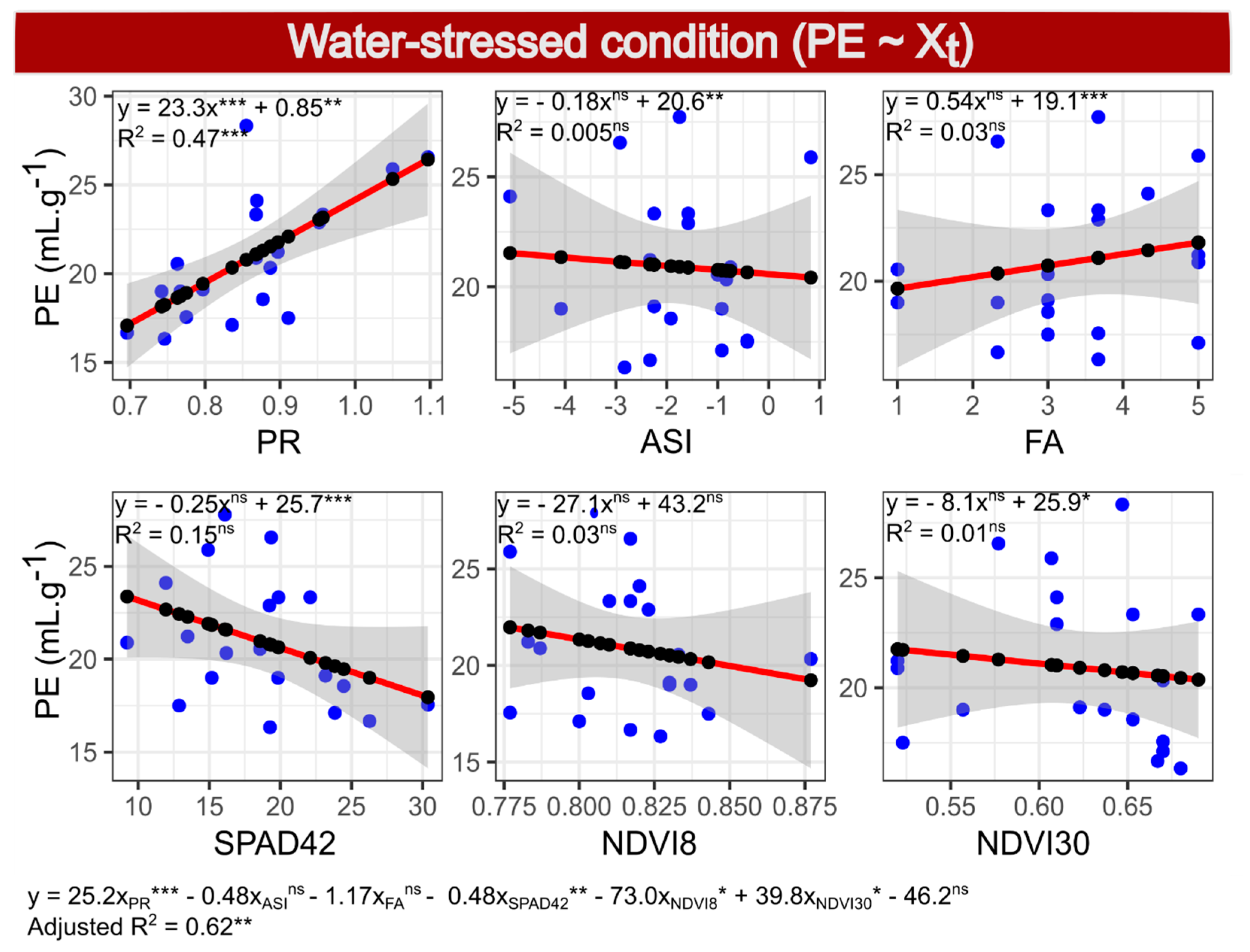
2.9. Selection of Model 3—PE under WS
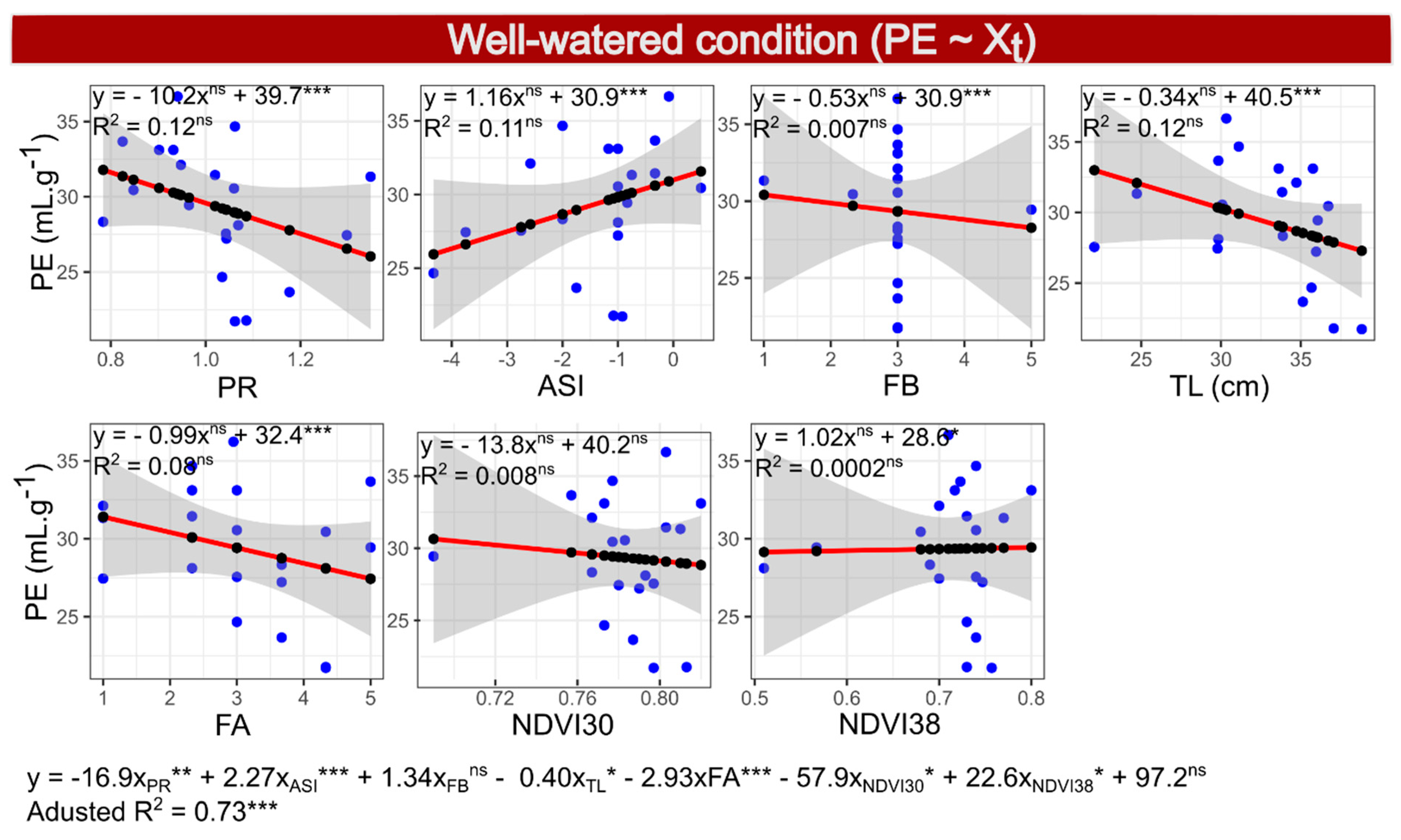
2.10. Selection of Model 4—PE under WW Conditions
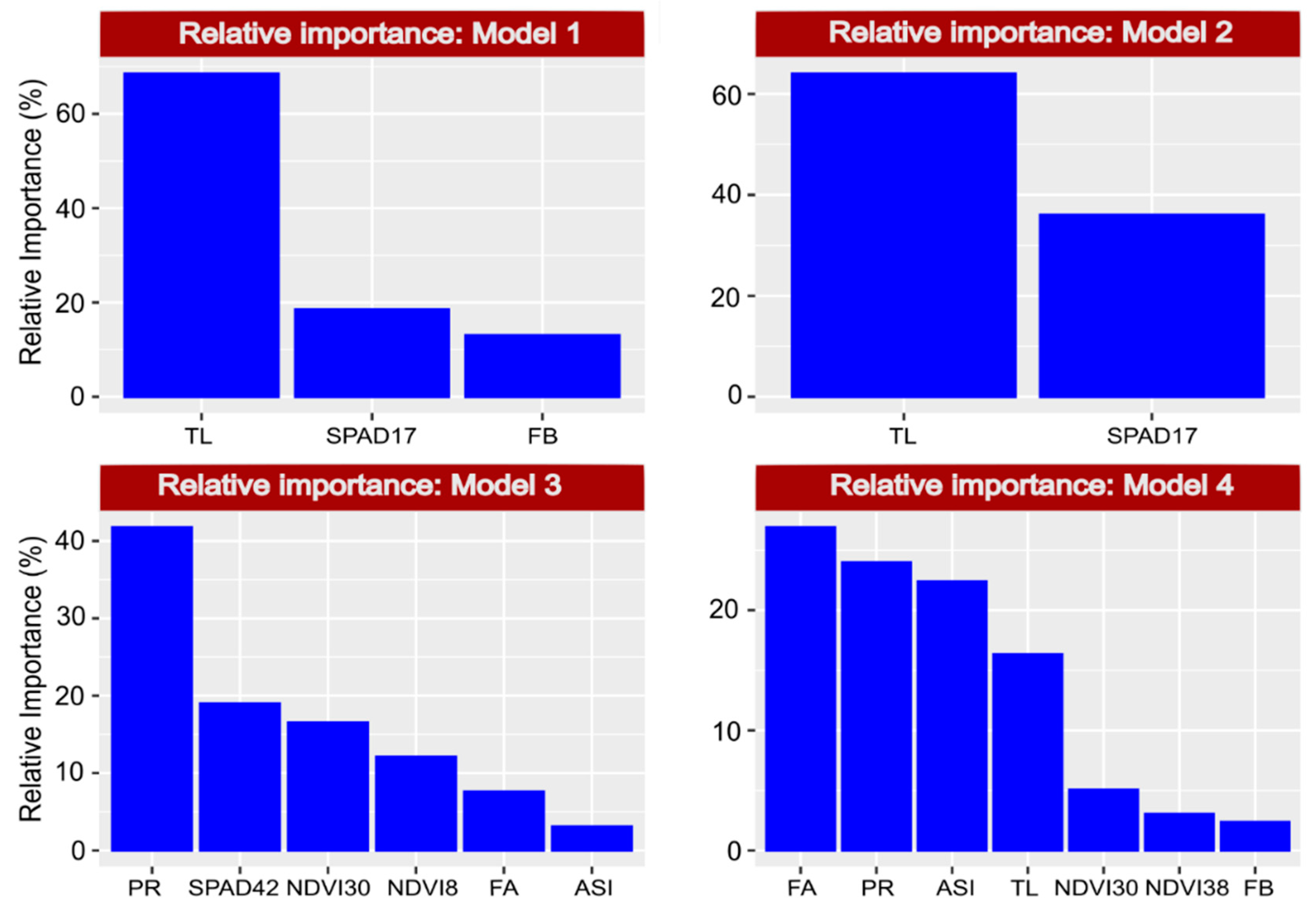
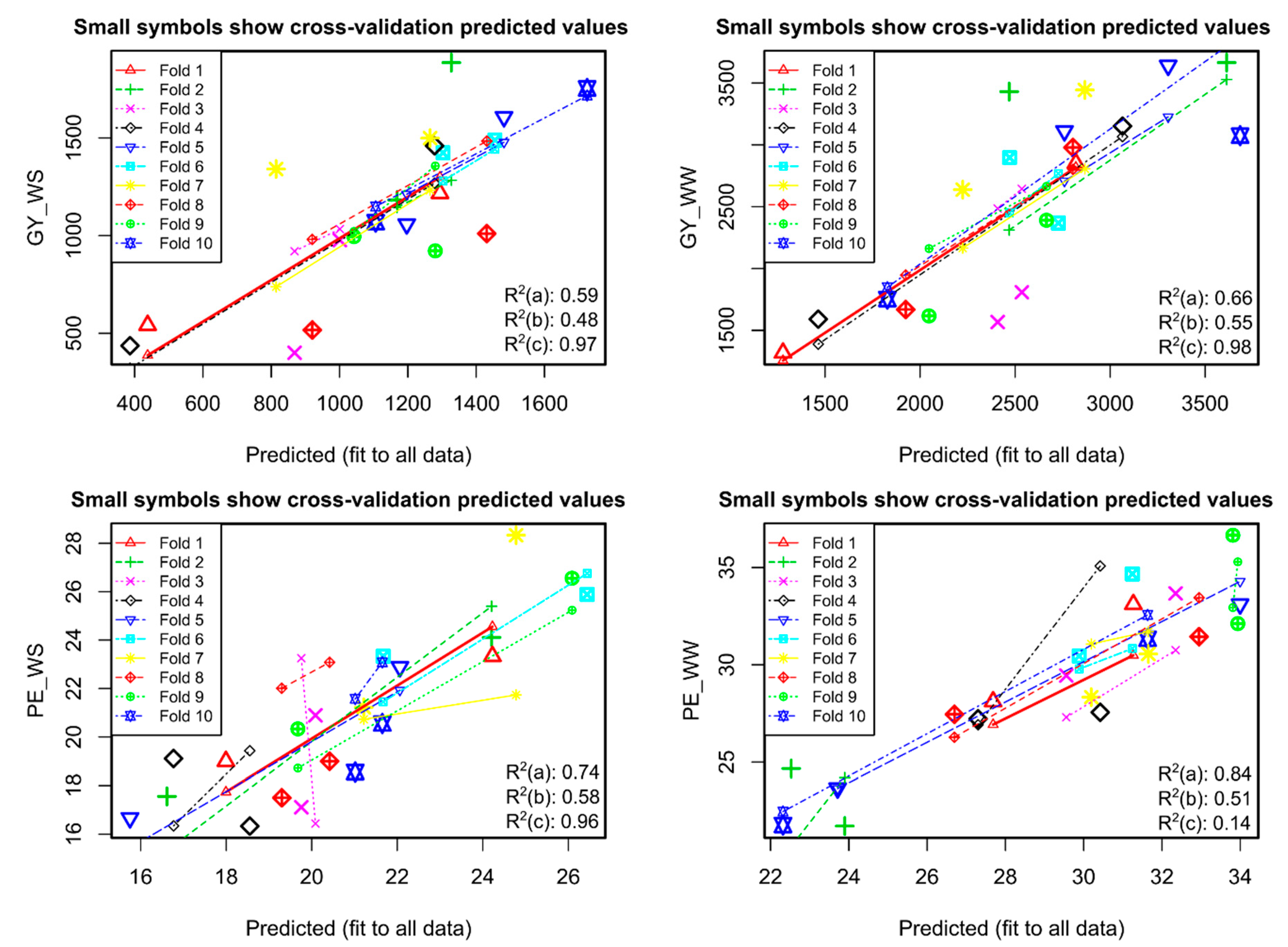
2.11. Predictive Effect on Dependent Variables (GY and PE) and Relative Importance of Independent Variables
3. Discussion
3.1. Effect of Water Restriction on GY, PE, Morpho-Agronomic Traits and on the Greenness Index
3.2. Multiple Regressions
4. Materials and Methods
4.1. Traits Evaluated
4.2. Analysis of Variance for Each Water Condition (WC) and Combined Analysis
4.3. Multiple Linear Regressions (MLR)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, S.; Qin, F. Genetic dissection of maize drought tolerance for trait improvement. Mol. Breed. 2021, 41, 8. [Google Scholar] [CrossRef]
- Mir, R.R.; Zaman-Allah, M.; Sreenivasulu, N.; Trethowan, R.; Varshney, R.K. Integrated genomics, physiology and breeding approaches for improving drought tolerance in crops. Theor. Appl. Genet. 2012, 125, 625–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araus, J.L.; Kefauver, S.C.; Zaman-Allah, M.; Olsen, M.S.; Cairns, J.E. Translating High-Throughput Phenotyping into Genetic Gain. Trends Plant. Sci. 2018, 23, 451–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrestha, R.K.; Lei, P.; Shi, D.; Hashimi, M.H.; Wang, S.; Xie, D.; Ni, J.; Ni, C. Response of maize (Zea mays L.) towards vapor pressure deficit. Environ. Exp. Bot. 2021, 181, 104293. [Google Scholar] [CrossRef]
- Awange, J.L.; Mpelasoka, F.; Goncalves, R.M. When every drop counts: Analysis of Droughts in Brazil for the 1901–2013 period. Sci. Total Environ. 2016, 566–567, 1472–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Loon, A.F.; Gleeson, T.; Clark, J.; Van Dijk, A.I.J.M.; Stahl, K.; Hannaford, J.; Di Baldassarre, G.; Teuling, A.J.; Tallaksen, L.M.; Uijlenhoet, R.; et al. Drought in the Anthropocene. Nat. Geosci. 2016, 9, 89–91. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Zhou, G.; He, Q.; Zhou, H. Stomatal limitations to photosynthesis and their critical Water conditions in different growth stages of maize under water stress. Agric. Water Manag. 2020, 241, 106330. [Google Scholar] [CrossRef]
- Kamphorst, S.H.; Gonçalves, G.M.B.; do Amaral Júnior, A.T.; de Lima, V.J.; Leite, J.T.; Schmitt, K.F.M.; dos Santos Junior, D.R.; Santos, J.S.; de Oliveira, F.T.; Corrêa, C.C.G.; et al. Screening of Popcorn Genotypes for Drought Tolerance Using Canonical Correlations. Agronomy 2020, 10, 1519. [Google Scholar] [CrossRef]
- Chapman, S.C.; Edmeades, G.O. Selection Improves Drought Tolerance in Tropical Maize Populations: II. Direct and correlated responses among secondary traits. Crop Sci. 1999, 39, 1315–1324. [Google Scholar] [CrossRef]
- Monneveux, P.; Sanchez, C.; Tiessen, A. Future progress in drought tolerance in maize needs new secondary traits and cross combinations. J. Agric. Sci. 2008, 146, 287–300. [Google Scholar] [CrossRef] [Green Version]
- Dias, K.O.D.G.; Gezan, S.A.; Guimarães, C.T.; Parentoni, S.N.; de Oliveira Guimarães, P.E.; Carneiro, N.P.; Portugal, A.F.; Bastos, E.A.; Cardoso, M.J.; de Oliveira Anoni, C.; et al. Estimating Genotype × Environment Interaction for and Genetic Correlations among Drought Tolerance Traits in Maize via Factor Analytic Multiplicative Mixed Models. Crop Sci. 2018, 58, 72–83. [Google Scholar] [CrossRef]
- Bolaños, J.; Edmeades, G.O. Eight cycles of selection for drought tolerance in lowland tropical maize. I. Responses in grain yield, biomass, and radiation utilization. Field Crops Res. 1993, 31, 233–252. [Google Scholar] [CrossRef]
- Câmara, T.M.M.; Bento, D.A.V.; Alves, G.F.; Santos, M.F.; Moreira, J.U.V.; de Souza Júnior, C.L. Parâmetros genéticos de caracteres relacionados à tolerância à deficiência hídrica em milho tropical. Bragantia 2007, 66, 595–603. [Google Scholar] [CrossRef]
- Santos, M.X.; Andrade, C.L.T.; Oliveira, A.C.; Leite, C.E.P.; Carvalho, H.W.L.; Gama, E.E.G.; Pacheco, C.A.P.; Gumarães, P.E.O.; Parentoni, S.N. Comportamento de Híbridos de Milho Selecionados e não Selecionados para Asi sob Estresse de Água no Florescimento e no Enchimento de Grãos. Rev. Bras. Milho E Sorgo 2003, 2, 71–81. [Google Scholar] [CrossRef]
- Adebayo, M.A.; Menkir, A.; Blay, E.; Gracen, V.; Danquah, E.; Hearne, S. Genetic analysis of drought tolerance in adapted × exotic crosses of maize inbred lines under managed stress conditions. Euphytica 2014, 196, 261–270. [Google Scholar] [CrossRef]
- Al-Naggar, A.; Atta, M.; Ahmed, M.; Younis, A. Genetic Variance, Heritability and Selection Gain of Maize (Zea mays L.) Adaptive Traits to High Plant Density Combined with Water Stress. J. Appl. Life Sci. Int. 2016, 7, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Cairns, J.E.; Sanchez, C.; Vargas, M.; Ordoñez, R.; Araus, J.L. Dissecting Maize Productivity: Ideotypes Associated with Grain Yield under Drought Stress and Well-watered Conditions. J. Integr. Plant. Biol. 2012, 54, 1007–1020. [Google Scholar] [CrossRef] [PubMed]
- Ramzan, J.; Aslam, M.; Ahsan, M.; Awan, F.S. Selection of screening criteria against drought stress at early growth stages in maize (Zea mays L.). Pak. J. Agric. Sci. 2019, 56, 633–643. [Google Scholar]
- Zia, S.; Romano, G.; Spreer, W.; Sanchez, C.; Cairns, J.; Araus, J.L.; Müller, J. Infrared Thermal Imaging as a Rapid Tool for Identifying Water-Stress Tolerant Maize Genotypes of Different Phenology. J. Agron. Crop Sci. 2013, 199, 75–84. [Google Scholar] [CrossRef]
- Kamphorst, S.H.; do Amaral Júnior, A.T.; de Lima, V.J.; Santos, P.H.A.D.; Rodrigues, W.P.; Vivas, J.M.S.; Gonçalves, G.M.B.; Schmitt, K.F.M.; Leite, J.T.; Vivas, M.; et al. Comparison of Selection Traits for Effective Popcorn (Zea mays L. var. Everta) Breeding Under Water Limiting Conditions. Front. Plant. Sci. 2020, 11, 1289. [Google Scholar] [CrossRef] [PubMed]
- de Lima, V.J.; do Amaral Júnior, A.T.; Kamphorst, S.H.; Bispo, R.B.; Leite, J.T.; de Oliveira Santos, T.; Schmitt, K.F.M.; Chaves, M.M.; de Oliveira, U.A.; Santos, P.H.A.D.; et al. Combined Dominance and Additive Gene Effects in Trait Inheritance of Drought-Stressed and Full Irrigated Popcorn. Agronomy 2019, 9, 782. [Google Scholar] [CrossRef] [Green Version]
- Kamphorst, S.H.; do Amaral Júnior, A.T.; de Lima, V.J.; Guimarães, L.J.M.; Schmitt, K.F.M.; Leite, J.T.; Santos, P.H.A.D.; Chaves, M.M.; Mafra, G.S.; dos Santos Junior, D.R.; et al. Can Genetic Progress for Drought Tolerance in Popcorn Be Achieved by Indirect Selection? Agronomy 2019, 9, 792. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, F.F.; Gomide, R.L.; de Albuquerque, P.E.P.; de Andrade, C.L.T.; Leite, C.E.P.; Parentoni, S.N.; de Oliveira Guimarães, P.E.; Guimarães, L.J.M.; Silva, A.R.; Bastos, E.A.; et al. Evaluation of maize core collection for drought tolerance. Crop Breed. Appl. Biotechnol. 2010, 10, 312–320. [Google Scholar] [CrossRef]
- Costa, E.F.N.; Santos, M.F.; Moro, G.V.; Alves, G.F.; de Souza Júnior, C.L. Herança da senescência retardada em milho. Pesqui. Agropecuária Bras. 2008, 43, 207–213. [Google Scholar] [CrossRef]
- Zaidi, P.H.; Srinivasan, G.; Cordova, H.; Sanchez, C. Gains from improvement for mid-season drought tolerance in tropical maize (Zea mays L.). Field Crops Res. 2004, 89, 135–152. [Google Scholar] [CrossRef]
- Li, X.-H.; Liu, X.-D.; Li, M.-S.; Zhang, S.-H. Identification of quantitative trait loci for anthesis-silking interval and yield components under drought stress in maize. J. Integr. Plant. Biol. 2003, 45, 852–857. [Google Scholar]
- Durães, F.O.M.; dos Santos, M.X.; Gama, E.E.G.e.; Magalhães, P.C.; Albuquerque, P.E.P.; Guimarães, C.T. Fenotipagem Associada a Tolerância a Seca em Milho para Uso em Melhoramento, Estudos Genômicos e Seleção Assistida por Marcadores. Embrapa Milho E Sorgo-Circ. Técnica (INFOTECA-E) 2004, 39, 18. [Google Scholar]
- Da Silva, W.J.; Vidal, B.C.; Martins, M.E.Q.; Vargas, H.; Pereira, C.; Zerbetto, M.; Miranda, L.C.M. What makes popcorn pop. Nature 1993, 362, 417. [Google Scholar] [CrossRef]
- Araus, J.L.; Sánchez, C.; Cabrera-Bosquet, L. Is heterosis in maize mediated through better water use? New Phytol. 2010, 187, 392–406. [Google Scholar] [CrossRef] [PubMed]
- Joshi, A.K.; Kumari, M.; Singh, V.P.; Reddy, C.M.; Kumar, S.; Rane, J.; Chand, R. Stay green trait: Variation, inheritance and its association with spot blotch resistance in spring wheat (Triticum aestivum L.). Euphytica 2006, 153, 59–71. [Google Scholar] [CrossRef]
- Tollenaar, M.; Ahmadzadeh, A.; Lee, E.A. Physiological Basis of Heterosis for Grain Yield in Maize. Crop Sci. 2004, 44, 2086. [Google Scholar] [CrossRef]
- Araus, J.L.; Serret, M.D.; Edmeades, G.O. Phenotyping maize for adaptation to drought. Front. Physiol. 2012, 3, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamphorst, S.H.; de Lima, V.J.; do Amaral Júnior, A.T.; Schmitt, K.F.M.; Leite, J.T.; Carvalho, C.M.; Silva, R.M.R.; Xavier, K.B.; Fereira, F.R.A.; Santos, P.H.A.; et al. Popcorn breeding for water-stress tolerance or for agronomic water-use efficiency? Genet. Mol. Res. 2018, 17. [Google Scholar] [CrossRef]
- Kamphorst, S.H.; de Lima, V.J.; Schimitt, K.F.M.; Leite, J.T.; Azeredo, V.C.; Pena, G.F.; Santos, P.H.A.D.; Júnior, D.R.S.; da Silva Júnior, S.B.; Bispo, R.B.; et al. Water stress adaptation of popcorn roots and association with agronomic traits. Genet. Mol. Res. 2018, 17, 1–14. [Google Scholar] [CrossRef]
- Wattoo, F.M.; Rana, R.M.; Fiaz, S.; Zafar, S.A.; Noor, M.A.; Hassan, H.M.; Bhatti, M.H.; Ur Rehman, S.; Anis, G.B.; Amir, R.M. Identification of drought tolerant maize genotypes and seedling based morpho-physiological selection indices for crop improvement. Sains Malays. 2018, 47, 295–302. [Google Scholar]
- Gazal, A.; Ahmed Dar, Z.; Ahmad Lone, A.; Yousuf, N.; Gulzar, S. Studies on Maize Yield under Drought Using Correlation and Path Coefficient Analysis. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 516–521. [Google Scholar] [CrossRef] [Green Version]
- James, G.; Witten, D.; Hastie, T.; Tibshirani, R.A. Introduction to Statistical Learning; Springer Texts in Statistics; Springer: New York, NY, USA, 2013; Volume 103, ISBN 978-1-4614-7137-0. [Google Scholar]
- Bruce, W.B.; Edmeades, G.O.; Barker, T.C. Molecular and physiological approaches to maize improvement for drought tolerance. J. Exp. Bot. 2002, 53, 13–25. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Santos, T.; do Amaral Junior, A.T.; Bispo, R.B.; de Lima, V.J.; Kamphorst, S.H.; Leite, J.T.; dos Santos Júnior, D.R.; Santos, P.H.A.D.; de Oliveira, U.A.; Schmitt, K.F.M.; et al. Phenotyping Latin American Open-Pollinated Varieties of Popcorn for Environments with Low Water Availability. Plants 2021, 10, 1211. [Google Scholar] [CrossRef]
- Leite, J.T.; Teixeira, A.; Kamphorst, S.H.; de Lima, V.J.; dos Santos Junior, D.R.; Schmitt, K.F.M.; de Souza, Y.P.; de Oliveira Santos, T.; Bispo, R.B.; Mafra, G.S.; et al. Water Use Efficiency in Popcorn (Zea mays L. var. everta): Which Physiological Traits Would Be Useful for Breeding? Plants 2021, 10, 1450. [Google Scholar] [CrossRef]
- Bolaños, J.; Edmeades, G.O. The importance of the anthesis-silking interval in breeding for drought tolerance in tropical maize. Field Crops Res. 1996, 48, 65–80. [Google Scholar] [CrossRef]
- Levitt, J. Water, Radiation, Salt, and Other Stresses; Academic Press: New York, NY, USA; Elsevier: New York, NY, USA, 1980; Volume 2. [Google Scholar]
- Baret, F.; Madec, S.; Irfan, K.; Lopez, J.; Comar, A.; Hemmerlé, M.; Dutartre, D.; Praud, S.; Tixier, M.H. Leaf-rolling in maize crops: From leaf scoring to canopy-level measurements for phenotyping. J. Exp. Bot. 2018, 69, 2705–2716. [Google Scholar] [CrossRef]
- Combe, L.; Escobar-Gutiérrez, A.J. Sénescence d’un pied de maïs: Évolution de la floraison à la récolte. Botany 2009, 87, 1036–1053. [Google Scholar] [CrossRef]
- Adebayo, M.A.; Menkir, A.; Hearne, S.; Kolawole, A.O. Gene action controlling normalized difference vegetation index in crosses of elite maize (Zea mays L.) inbred lines. Cereal Res. Commun. 2017, 45, 675–686. [Google Scholar] [CrossRef] [Green Version]
- De Castro, F.A.; Campostrini, E.; Netto, A.T.; De Menezes De Assis Gomes, M.; Ferraz, T.M.; Glenn, D.M. Portable chlorophyll meter (PCM-502) values are related to total chlorophyll concentration and photosynthetic capacity in papaya (Carica papaya L.). Theor. Exp. Plant. Physiol. 2014, 26, 201–210. [Google Scholar] [CrossRef]
- Hallauer, A.R.; Carena, M.J.; Miranda Filho, J.B. Quantitative Genetics in Maize Breeding; Springer: Ames, IA, USA, 2010. [Google Scholar]
- Olive, D.J. Linear Regression; Springer International Publishing: Cham, Switzerland, 2017; ISBN 978-3-319-55250-7. [Google Scholar]
- Edmeades, G.O.; Bolaños, J.; Chapman, S.C.; Lafitte, H.R.; Banziger, M.; Edmeades, G.O. Selection Improves Drought Tolerance in Tropical Maize Populations: I. Gains in biomass, grain yield, and harvest index. Crop Sci. 1999, 39, 1306–1315. [Google Scholar] [CrossRef]
- Lima Neto, F.P.; Souza Júnior, C.L. de Number of recombinations and genetic properties of a maize population undergoing recurrent selection. Sci. Agric. 2009, 66, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Vittorazzi, C.; Júnior, A.T.A.; Guimarães, A.G.; Silva, F.H.L.; Pena, G.F.; Daher, R.F.; Gerhardt, I.F.S.; Oliveira, G.H.F.; Santos, P.H.A.D.; Souza, Y.P.; et al. Evaluation of genetic variability to form heterotic groups in popcorn. Genet. Mol. Res. 2018, 17, 18083. [Google Scholar] [CrossRef]
- van Raij, B.; Cantarella, H.; Quaggio, J.A.; Furlani, A.M.C. Recomendações de Adubação e Calagem Para o Estado de São Paulo; Instituto Agronômico/Fundação IAC Campinas: Campinas, Brazil, 1997; Volume 285. [Google Scholar]
- IPGRI-International Plant Genetic Resources Institute. Descritores Para o Milho; International Maize and Wheat Improvement Center, Mexico City/International Plant Genetic Resources Institute: Rome, Italy, 2000. [Google Scholar]
- Lindeman, R.H.; Merenda, P.F.; Gold, R.Z. Introduction to Bivariate and Multivariate Analysis; Scott. Foresman Co.: Glenview, UK, 1980. [Google Scholar]
- Kuhn, M.; Johnson, K. Applied Predictive Modeling; Springer New York: New York, NY, USA, 2013; ISBN 978-1-4614-6848-6. [Google Scholar]
- R Core Team R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | WC | Mean ± Standard Deviations | Proportional Reductions (%) | Interaction G*WC | ||
|---|---|---|---|---|---|---|
| Dependent variables | ||||||
| GY | WS | 1139.11 | ± | 477.24 | 55.30 | * |
| WW | 2548.07 | ± | 910.89 | |||
| PE | WS | 20.91 | ± | 3.74 | 28.75 | ** |
| WW | 29.35 | ± | 5.24 | |||
| Morphological traits | ||||||
| GW | WS | 9.70 | ± | 1.47 | 23.49 | ns |
| WW | 12.67 | ± | 1.71 | |||
| PR | WS | 0.86 | ± | 0.14 | 15.84 | ns |
| WW | 1.02 | ± | 0.18 | |||
| TL | WS | 31.60 | ± | 4.56 | 3.50 | ns |
| WW | 32.75 | ± | 4.62 | |||
| TB | WS | 14.18 | ± | 3.48 | 7.01 | ns |
| WW | 15.25 | ± | 4.61 | |||
| ASI | WS | −1.77 | ± | 1.41 | −25.44 | ns |
| WW | −1.41 | ± | 1.29 | |||
| FA | WS | 3.33 | ± | 1.34 | −8.70 | ns |
| WW | 3.07 | ± | 1.38 | |||
| FB | WS | 3.10 | ± | 0.77 | −4.49 | ns |
| WW | 2.97 | ± | 0.69 | |||
| SPAD index | ||||||
| SPAD7 | WS | 46.23 | ± | 4.03 | 5.82 | ns |
| WW | 49.08 | ± | 3.47 | |||
| SPAD12 | WS | 45.47 | ± | 4.47 | 4.48 | ns |
| WW | 47.61 | ± | 4.12 | |||
| SPAD17 | WS | 43.55 | ± | 4.17 | 10.82 | ns |
| WW | 48.84 | ± | 4.55 | |||
| SPAD22 | WS | 40.33 | ± | 5.15 | 17.64 | ns |
| WW | 48.97 | ± | 4.15 | |||
| SPAD28 | WS | 39.70 | ± | 5.15 | 17.56 | ns |
| WW | 48.16 | ± | 3.97 | |||
| SPAD35 | WS | 32.51 | ± | 6.91 | 29.32 | ns |
| WW | 45.99 | ± | 5.17 | |||
| SPAD42 | WS | 18.81 | ± | 6.60 | 52.20 | ns |
| WW | 39.36 | ± | 6.73 | |||
| Normalized Difference Vegetation Index (NVDI) | ||||||
| NDVI8 | WS | 0.82 | ± | 0.03 | −0.20 | ns |
| WW | 0.81 | ± | 0.04 | |||
| NDVI19 | WS | 0.73 | ± | 0.04 | 9.51 | ns |
| WW | 0.81 | ± | 0.04 | |||
| NDVI24 | WS | 0.67 | ± | 0.05 | 15.28 | ns |
| WW | 0.79 | ± | 0.04 | |||
| NDVI30 | WS | 0.62 | ± | 0.07 | 20.41 | ns |
| WW | 0.78 | ± | 0.04 | |||
| NDVI38 | WS | 0.47 | ± | 0.10 | 33.26 | ns |
| WW | 0.71 | ± | 0.10 | |||
| Variables | GY~MORPH | PE~MORPH | ||
|---|---|---|---|---|
| WS | WW | WS | WW | |
| Intercept | −2105.89 | −3035.85 | −3.32 | 66.02 *** |
| GW | 100.82 | 215.02 | 0.06 | −0.03 |
| PR | 604.33 | −1143.68 | 33.17 ** | −17.33 * |
| TL | 68.61 * | 135.02 * | −0.32 * | −0.37 |
| TR | 23.99 | −2.505 | 0.38 | −0.029 |
| ASI | 25.93 | −98.29 | −0.42 | 1.81 * |
| FA | 95.55 | −1.66 | 0.51 | −2.55 ** |
| FB | −333.45 * | −164.81 | −0.79 * | 1.55 |
| Adjusted R2 | 0.52 * | 0.31 ns | 0.39 ns | 0.57 ** |
| Variables | GY~SPAD Index | PE~SPAD Index | ||
|---|---|---|---|---|
| WS | WW | WS | WW | |
| Intecept | 778.45 | 6334.46 | 43.52626 ** | 15.88745 |
| SPAD7 | 27.12 | −190.49 | −0.56 | 0.2992 |
| SPAD12 | −45.5 | −119.97 | −0.26 | 0.118 |
| SPAD17 | −37.47 | 280.58 ** | 0.42 | −0.59819 |
| SPAD22 | 81.88 | 37.61 | −0.03 | −0.37379 |
| SPAD28 | −57.05 | 23.85 | −0.13 | 0.65234 |
| SPAD35 | 78.86 | −216.18 | 0.49 | 0.18314 |
| SPAD42 | −42.14 | 114.95 | −0.61582 * | 0.02108 |
| Adjusted R2 | 0.2257 ns | 0.425 * | 0.1937 ns | −0.4308 ns |
| Variable | GY~NDVI | PE~NDVI | ||
|---|---|---|---|---|
| WS | WW | WS | WW | |
| Intecept | 3667 | −4247 | 66.72 * | −13.99 |
| NDVI8 | −6149 | 1950 | −18.86 | 100.31 * |
| NDVI19 | 8744 | −8872 | 3.32 | 116.93 |
| NDVI24 | −7108 | 15358 | −84.78 | −26.73 |
| NDVI30 | 1965 | −4620 | 64.31 * | −154.04 * |
| NDVI38 | −894 | 5550 | −34.50 * | 11.75 |
| Adjusted R2 | 0.233 ns | 0.076 ns | 0.268 ns | 0.268 ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamphorst, S.H.; Gonçalves, G.M.B.; Amaral Júnior, A.T.d.; Lima, V.J.d.; Schmitt, K.F.M.; Leite, J.T.; Azeredo, V.C.; Gomes, L.P.; Silva, J.G.d.S.; Carvalho, C.M.; et al. Supporting Physiological Trait for Indirect Selection for Grain Yield in Drought-Stressed Popcorn. Plants 2021, 10, 1510. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10081510
Kamphorst SH, Gonçalves GMB, Amaral Júnior ATd, Lima VJd, Schmitt KFM, Leite JT, Azeredo VC, Gomes LP, Silva JGdS, Carvalho CM, et al. Supporting Physiological Trait for Indirect Selection for Grain Yield in Drought-Stressed Popcorn. Plants. 2021; 10(8):1510. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10081510
Chicago/Turabian StyleKamphorst, Samuel Henrique, Gabriel Moreno Bernardo Gonçalves, Antônio Teixeira do Amaral Júnior, Valter Jário de Lima, Kátia Fabiane Medeiros Schmitt, Jhean Torres Leite, Valdinei Cruz Azeredo, Letícia Peixoto Gomes, José Gabriel de Souza Silva, Carolina Macedo Carvalho, and et al. 2021. "Supporting Physiological Trait for Indirect Selection for Grain Yield in Drought-Stressed Popcorn" Plants 10, no. 8: 1510. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10081510