
Alterations of Content and Composition of Individual Sulfolipids, and Change of Fatty Acids Profile of Galactolipids in Lettuce Plants (Lactuca sativa L.) Grown under Sulfur Nutrition

 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Impact of S Application on Different Lipids of the Chloroplast Membrane
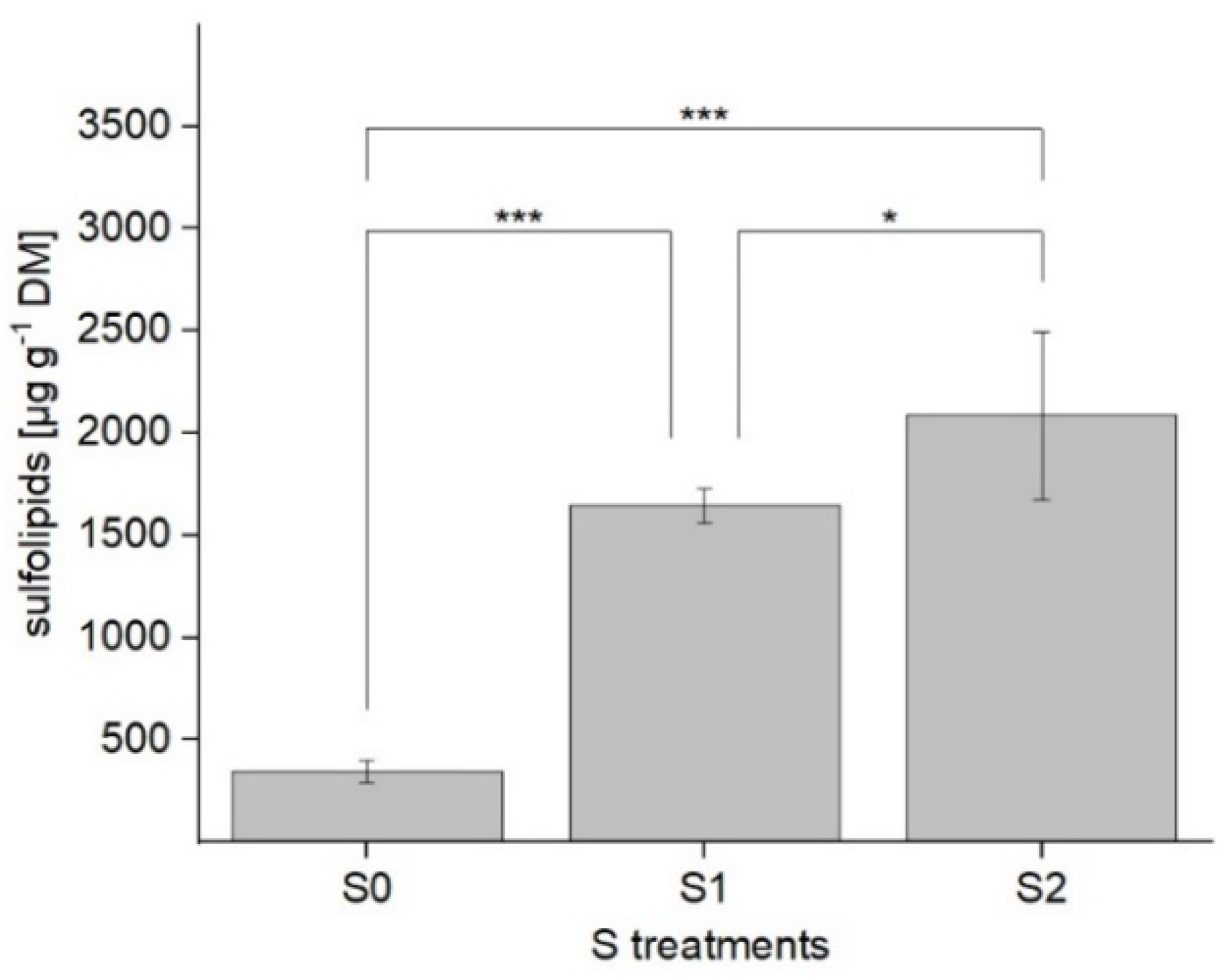
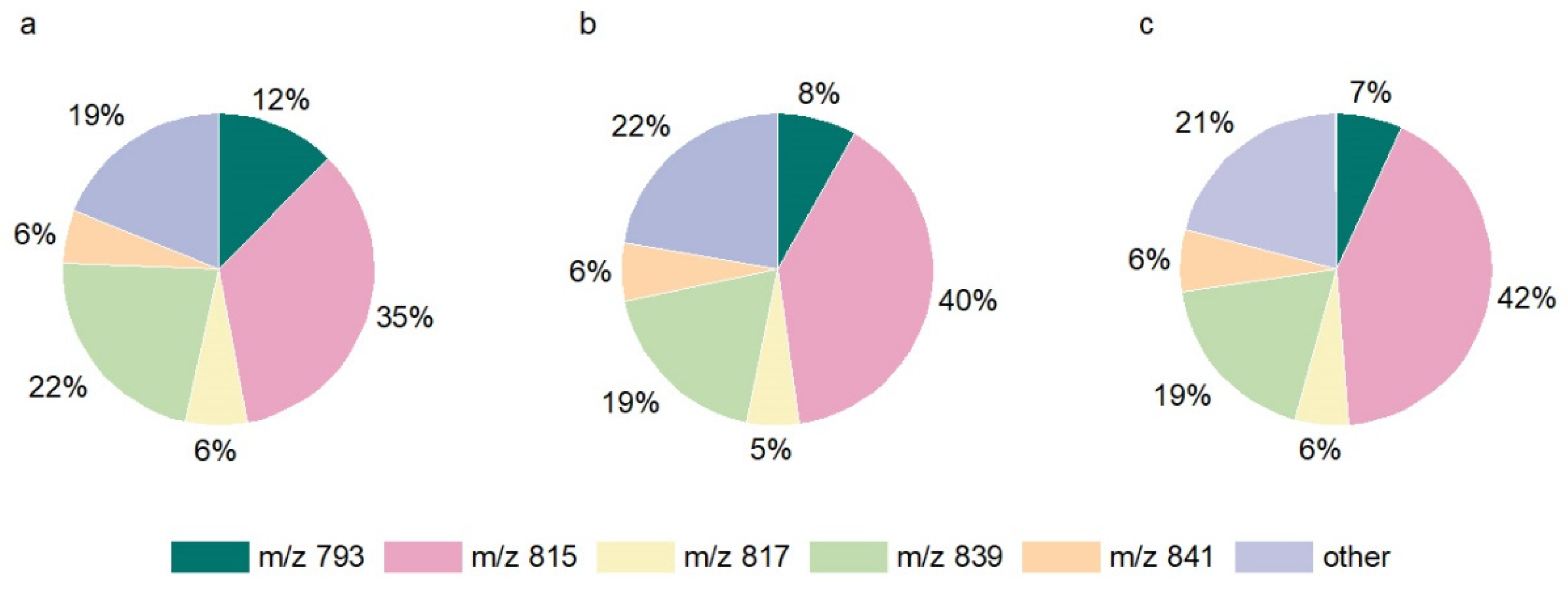
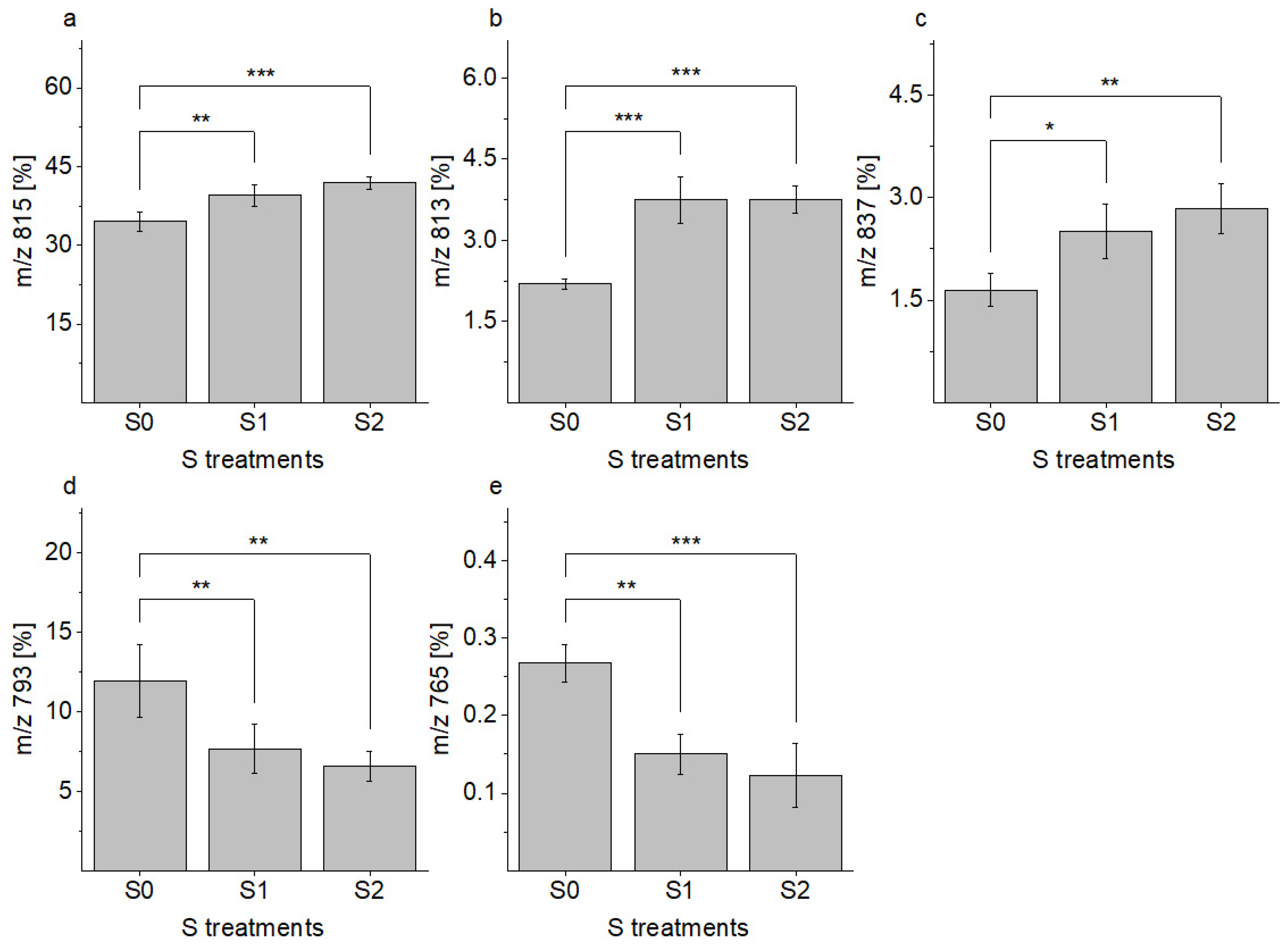
2.1.1. Determination and Composition of SQDG
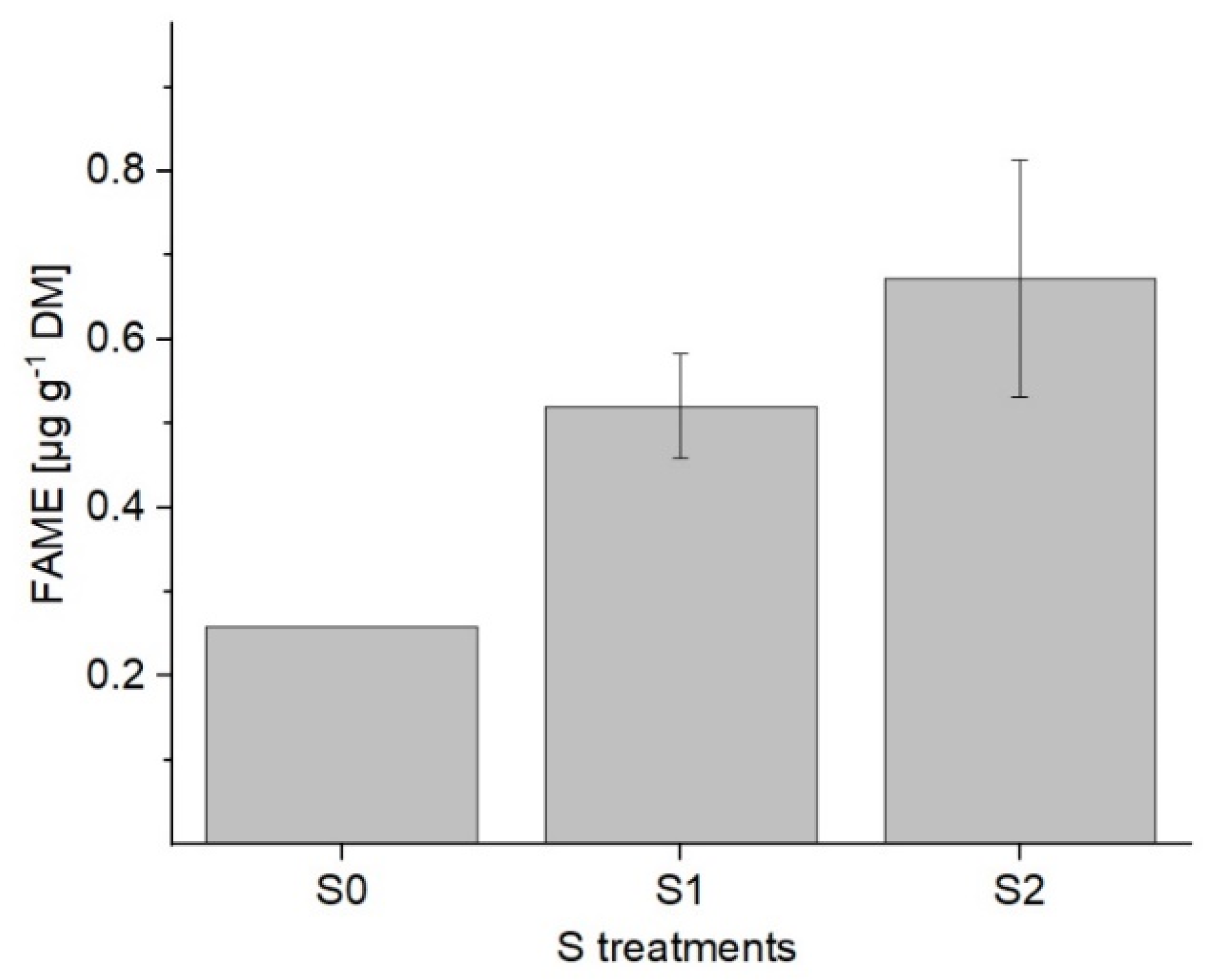
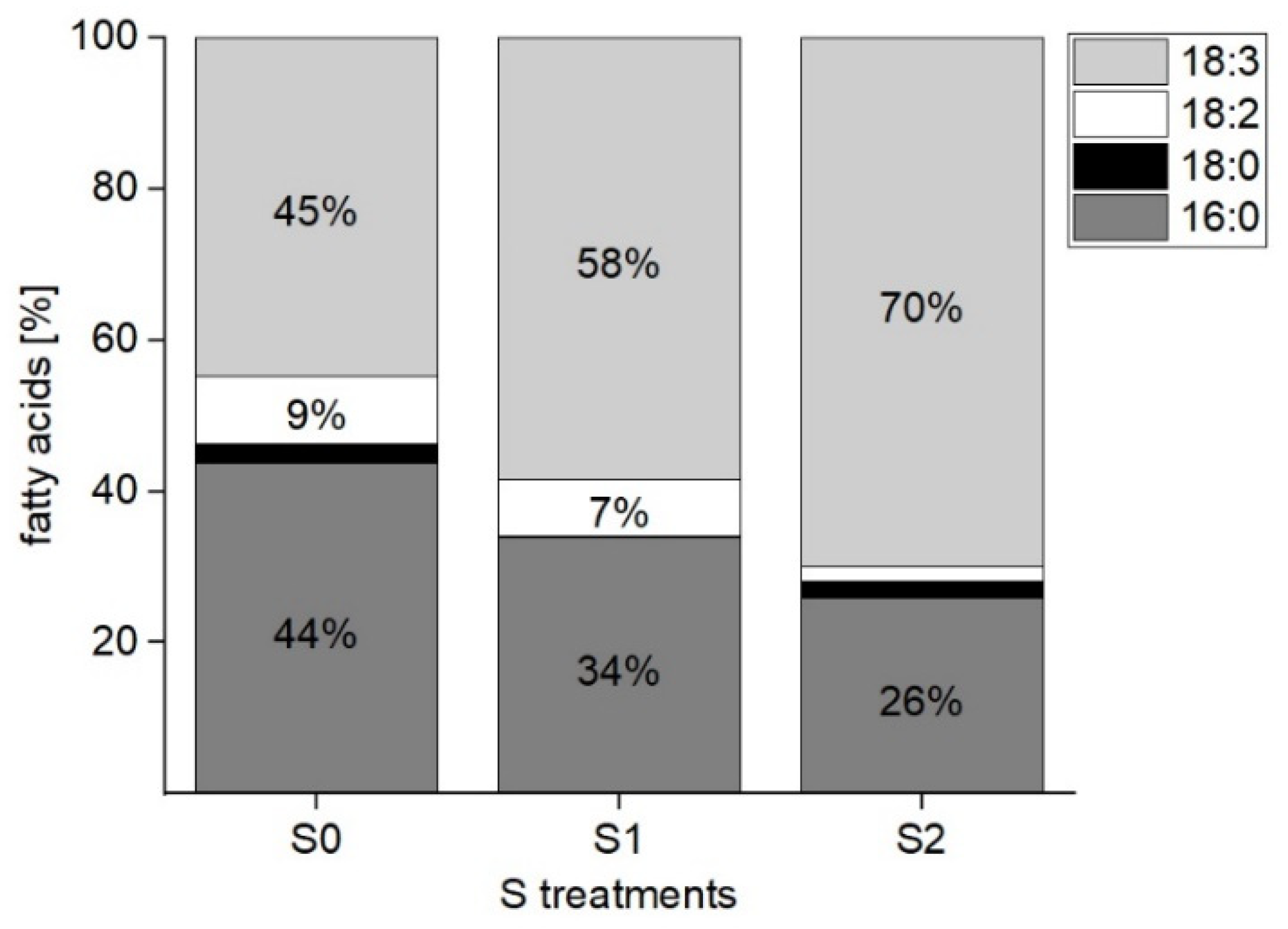
2.1.2. Determination of Fatty Acids in the Galactolipids
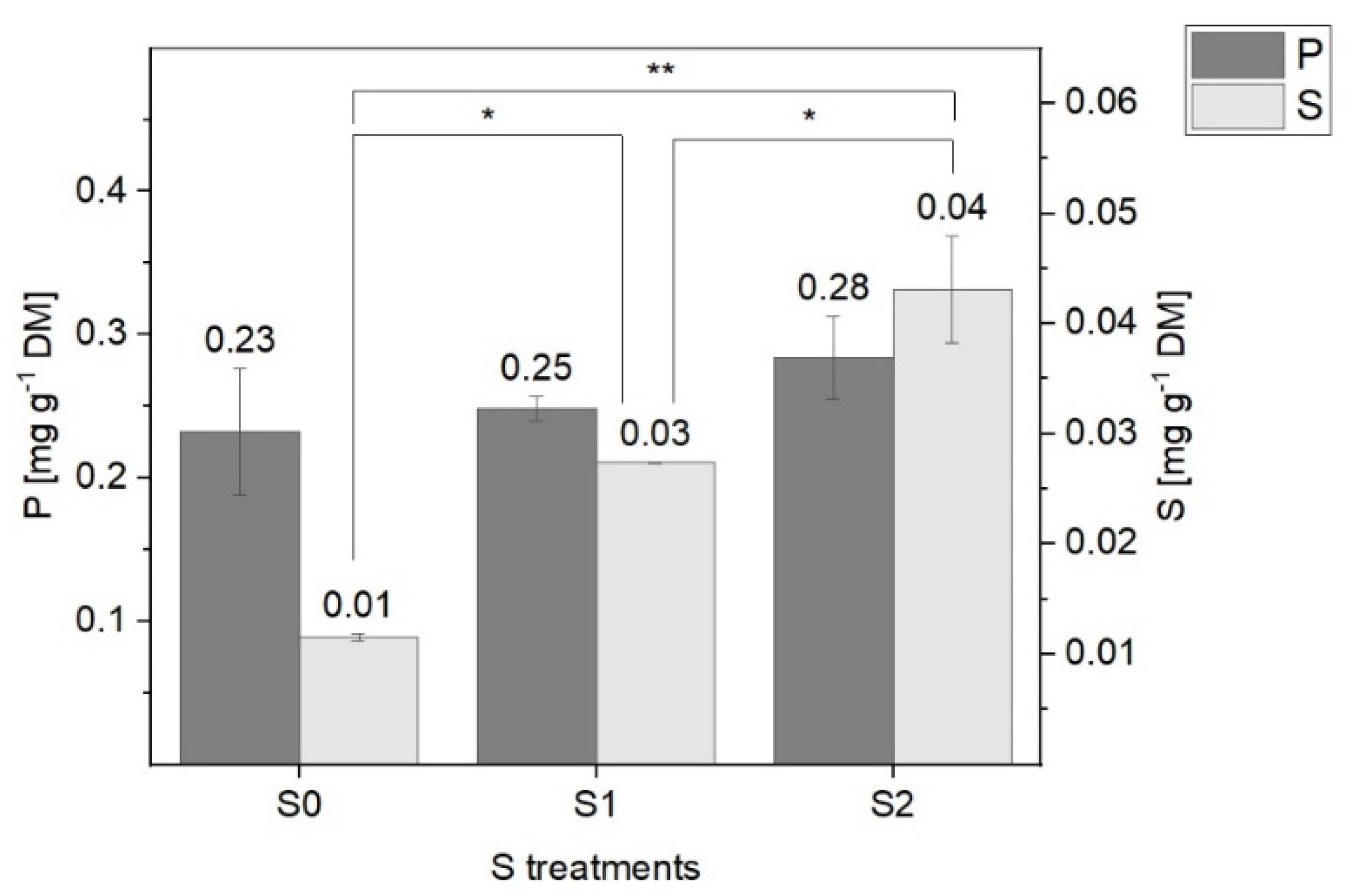
2.2. Total P and S Contents
2.3. Impact of Various S Levels on Pigment Concentration
3. Discussion
4. Materials and Methods
4.1. Chemicals and Materials
4.2. Plant Material and Growth Conditions
4.3. Lipid Extraction Procedure
4.4. Clean-Up and Separation of the Lipids by Solid Phase Extraction
4.5. Determination of Sulfolipids Using LC-ESI-MS/MS
4.6. GC-MSD Determination of Galactolipids on the Basis of their Fatty Acids Profile
4.7. Determination of the Total P and S Content in the SQDG Extracts
4.8. Determination of Total Chlorophyll a, b and Carotenoids
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maathuis, F.J.M.; Diatloff, E. Roles and Functions of Plant Mineral Nutrients. In Plant Mineral Nutrients: Methods and Protocols; Maathuis, F.J.M., Ed.; Humana Press: Totowa, NJ, USA, 2013; pp. 1–21. [Google Scholar] [CrossRef]
- Kopriva, S.; Malagoli, M.; Takahashi, H. Sulfur nutrition: Impacts on plant development, metabolism, and stress responses. J. Exp. Bot. 2019, 70, 4069–4073. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, M.A.; Mühling, K.H. Plant-derived sulfur containing natural products produced as a response to biotic and abiotic stresses: A review of their structural diversity and medicinal importance. J. Appl. Bot. Food Qual. 2019, 92, 204–215. [Google Scholar] [CrossRef]
- Gustafson, K.R.; Cardellina, J.H.; Fuller, R.W.; Weislow, O.S.; Kiser, R.F.; Snader, K.M.; Patterson, G.M.; Boyd, M.R. AIDS-antiviral sulfolipids from cyanobacteria (blue-green algae). J. Natl. Cancer Inst. 1989, 81, 1254–1258. [Google Scholar] [CrossRef]
- Burrichter, A.; Denger, K.; Franchini, P.; Huhn, T.; Müller, N.; Spiteller, D.; Schleheck, D. Anaerobic degradation of the plant sugar sulfoquinovose concomitant with H2S production: Escherichia coli K-12 and Desulfovibrio sp. strain DF1 as co-culture model. Front. Microbiol. 2018, 9, 2792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haange, S.-B.; Groeger, N.; Froment, J.; Rausch, T.; Burkhardt, W.; Gonnermann, S.; Braune, A.; Blaut, M.; von Bergen, M.; Rolle-Kampczyk, U. Multiplexed Quantitative Assessment of the Fate of Taurine and Sulfoquinovose in the Intestinal Microbiome. Metabolites 2020, 10, 430. [Google Scholar] [CrossRef]
- Peck, S.C.; Denger, K.; Burrichter, A.; Irwin, S.M.; Balskus, E.P.; Schleheck, D. A glycyl radical enzyme enables hydrogen sulfide production by the human intestinal bacterium Bilophila wadsworthia. Proc. Natl. Acad. Sci. USA 2019, 116, 3171–3176. [Google Scholar] [CrossRef] [Green Version]
- Fischer, J.; Treblin, M.; Sitz, T.; Rohn, S. Development of a targeted HPLC-ESI-QqQ-MS/MS method for the quantification of sulfolipids from a cyanobacterium, selected leafy vegetables, and a microalgae species. Anal. Bioanal. Chem. 2021, 413, 1941–1954. [Google Scholar] [CrossRef]
- Block, M.A.; Dorne, A.J.; Joyard, J.; Douce, R. Preparation and characterization of membrane fractions enriched in outer and inner envelope membranes from spinach chloroplasts. II. Biochemical characterization. J. Biol. Chem. 1983, 258, 13281–13286. [Google Scholar] [CrossRef]
- Leech, R.; Murphy, D. Cooperative function of chloroplasts in the biosynthesis of small molecules. Top. Photosynth. 1976, 1, 365–401. [Google Scholar]
- Lunde, C.; Zygadlo, A.; Simonsen, H.T.; Nielsen, P.L.; Blennow, A.; Haldrup, A. Sulfur starvation in rice: The effect on photosynthesis, carbohydrate metabolism, and oxidative stress protective pathways. Physiol. Plant. 2008, 134, 508–521. [Google Scholar] [CrossRef]
- Lin, Y.-T.; Chen, L.-J.; Herrfurth, C.; Feussner, I.; Li, H.-m. Reduced biosynthesis of digalactosyldiacylglycerol, a major chloroplast membrane lipid, leads to oxylipin overproduction and phloem cap lignification in Arabidopsis. Plant Cell 2016, 28, 219–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Testerink, C.; Munnik, T. Molecular, cellular, and physiological responses to phosphatidic acid formation in plants. J. Exp. Bot. 2011, 62, 2349–2361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djanaguiraman, M.; Prasad, P.; Kumari, J.; Rengel, Z. Root length and root lipid composition contribute to drought tolerance of winter and spring wheat. Plant Soil 2019, 439, 57–73. [Google Scholar] [CrossRef] [Green Version]
- Sui, N.; Han, G. Salt-induced photoinhibition of PSII is alleviated in halophyte Thellungiella halophila by increases of unsaturated fatty acids in membrane lipids. Acta Physiol. Plant. 2014, 36, 983–992. [Google Scholar] [CrossRef]
- Liu, X.; Ma, D.; Zhang, Z.; Wang, S.; Du, S.; Deng, X.; Yin, L. Plant lipid remodeling in response to abiotic stresses. Environ. Exp. Bot. 2019, 165, 174–184. [Google Scholar] [CrossRef]
- Lambers, H.; Cawthray, G.R.; Giavalisco, P.; Kuo, J.; Laliberté, E.; Pearse, S.J.; Scheible, W.-R.; Stitt, M.; Teste, F.; Turner, B.L. Proteaceae from severely phosphorus-impoverished soils extensively replace phospholipids with galactolipids and sulfolipids during leaf development to achieve a high photosynthetic phosphorus-use-efficiency. New Phytol. 2012, 196, 1098–1108. [Google Scholar] [CrossRef]
- Sugimoto, K.; Sato, N.; Tsuzuki, M. Utilization of a chloroplast membrane sulfolipid as a major internal sulfur source for protein synthesis in the early phase of sulfur starvation in Chlamydomonas reinhardtii. FEBS Lett. 2007, 581, 4519–4522. [Google Scholar] [CrossRef] [Green Version]
- Murata, N.; Ishizaki-Nishizawa, O.; Higashi, S.; Hayashi, H.; Tasaka, Y.; Nishida, I. Genetically engineered alteration in the chilling sensitivity of plants. Nature 1992, 356, 710–713. [Google Scholar] [CrossRef]
- Wolter, F.; Schmidt, R.; Heinz, E. Chilling sensitivity of Arabidopsis thaliana with genetically engineered membrane lipids. EMBO J. 1992, 11, 4685–4692. [Google Scholar] [CrossRef]
- Murakami, Y.; Tsuyama, M.; Kobayashi, Y.; Kodama, H.; Iba, K. Trienoic fatty acids and plant tolerance of high temperature. Science 2000, 287, 476–479. [Google Scholar] [CrossRef]
- Bybordi, A.; Tabatabaei, S.J.; Ahmadev, A. Effects of salinity on fatty acid composition of canola (Brassica napus L.). J. Food Agric. Environ. 2010, 8, 113–115. [Google Scholar]
- Kim, M.J.; Moon, Y.; Tou, J.C.; Mou, B.; Waterland, N.L. Nutritional value, bioactive compounds and health benefits of lettuce (Lactuca sativa L.). J. Food Compos. Anal. 2016, 49, 19–34. [Google Scholar] [CrossRef]
- Petrazzini, L.L.; Souza, G.A.; Rodas, C.L.; Emrich, E.B.; Carvalho, J.G.; Souza, R.J. Nutritional deficiency in crisphead lettuce grown in hydroponics. Hortic. Bras. 2014, 32, 310–313. [Google Scholar] [CrossRef] [Green Version]
- Abdalla, M.A.; Li, F.; Wenzel-Storjohann, A.; Sulieman, S.; Tasdemir, D.; Mühling, K.H. Comparative Metabolite Profile, Biological Activity and Overall Quality of Three Lettuce (Lactuca sativa L., Asteraceae) Cultivars in Response to Sulfur Nutrition. Pharmaceutics 2021, 13, 713. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, P.; Das, P.; Mukherjee, R.; Banik, S.; Karmakar, S.; Chatterjee, S. Extraction and quantification of pigments from Indian traditional medicinal plants: A comparative study between tree, shrub, and herb. Int. J. Pharm. Sci. Res. 2018, 9, 3052–3059. [Google Scholar] [CrossRef]
- Roosta, H.R.; Estaji, A.; Niknam, F. Effect of iron, zinc and manganese shortage-induced change on photosynthetic pigments, some osmoregulators and chlorophyll fluorescence parameters in lettuce. Photosynthetica 2018, 56, 606–615. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Oukarroum, A.; Alexandrov, V.; Kouzmanova, M.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Allakhverdiev, S.I.; Goltsev, V. Identification of nutrient deficiency in maize and tomato plants by in vivo chlorophyll a fluorescence measurements. Plant Physiol. Biochem. 2014, 81, 16–25. [Google Scholar] [CrossRef]
- Fu, C.; Li, M.; Zhang, Y.; Zhang, Y.; Yan, Y.; Wang, Y.A. Morphology, photosynthesis, and internal structure alterations in field apple leaves under hidden and acute zinc deficiency. Sci. Hortic. 2015, 193, 47–54. [Google Scholar] [CrossRef]
- Solovchenko, A.; Khozin-Goldberg, I.; Selyakh, I.; Semenova, L.; Ismagulova, T.; Lukyanov, A.; Mamedov, I.; Vinogradova, E.; Karpova, O.; Konyukhov, I.; et al. Phosphorus starvation and luxury uptake in green microalgae revisited. Algal Res. 2019, 43, 101651. [Google Scholar] [CrossRef]
- Benning, C. Biosynthesis and function of the sulfolipid sulfoquinovosyl diacylglycerol. Annu. Rev. Plant Biol. 1998, 49, 53–75. [Google Scholar] [CrossRef]
- Sato, N.; Aoki, M.; Maru, Y.; Sonoike, K.; Minoda, A.; Tsuzuki, M. Involvement of sulfoquinovosyl diacylglycerol in the structural integrity and heat-tolerance of photosystem II. Planta 2003, 217, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Sato, N.; Sonoike, K.; Tsuzuk, M.; Kawaguchi, A. Impaired photosystem II in a mutant of Chlamydomonas reinhardtii defective in sulfoquinovosyl diacylglycerol. Eur. J. Biochem. 1995, 234, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, Y.; Umena, Y.; Nagao, R.; Endo, K.; Kobayashi, K.; Akita, F.; Suga, M.; Wada, H.; Noguchi, T.; Shen, J.R. Thylakoid membrane lipid sulfoquinovosyl-diacylglycerol (SQDG) is required for full functioning of photosystem II in Thermosynechococcus elongatus. J. Biol. Chem. 2018, 293, 14786–14797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minoda, A.; Sato, N.; Nozaki, H.; Okada, K.; Takahashi, H.; Sonoike, K.; Tsuzuki, M. Role of sulfoquinovosyl diacylglycerol for the maintenance of photosystem II in Chlamydomonas reinhardtii. Eur. J. Biochem. 2002, 269, 2353–2358. [Google Scholar] [CrossRef] [PubMed]
- Minoda, A.; Sonoike, K.; Okada, K.; Sato, N.; Tsuzuki, M. Decrease in the efficiency of the electron donation to tyrosine Z of photosystem II in an SQDG-deficient mutant of Chlamydomonas. FEBS Lett. 2003, 553, 109–112. [Google Scholar] [CrossRef] [Green Version]
- Horton, P.; Ruban, A. Molecular design of the photosystem II light-harvesting antenna: Photosynthesis and photoprotection. J. Exp. Bot. 2005, 56, 365–373. [Google Scholar] [CrossRef]
- Schaller, S.; Latowski, D.; Jemiola-Rzeminska, M.; Dawood, A.; Wilhelm, C.; Strzalka, K.; Goss, R. Regulation of LHCII aggregation by different thylakoid membrane lipids. Biochim. Biophys. Acta Bioenerg. 2011, 1807, 326–335. [Google Scholar] [CrossRef] [Green Version]
- Kirchhoff, H.; Hinz, H.-J.; Rösgen, J. Aggregation and fluorescence quenching of chlorophyll a of the light-harvesting complex II from spinach in vitro. Biochim. Biophys. Acta Bioenerg. 2003, 1606, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, K.; Tsuzuki, M.; Sato, N. Regulation of synthesis and degradation of a sulfolipid under sulfur-starved conditions and its physiological significance in Chlamydomonas reinhardtii. New Phytol. 2010, 185, 676–686. [Google Scholar] [CrossRef]
- Belkhadi, A.; Hediji, H.; Abbes, Z.; Nouairi, I.; Barhoumi, Z.; Zarrouk, M.; Chaibi, W.; Djebali, W. Effects of exogenous salicylic acid pre-treatment on cadmium toxicity and leaf lipid content in Linum usitatissimum L. Ecotoxicol. Environ. Saf. 2010, 73, 1004–1011. [Google Scholar] [CrossRef]
- Routaboul, J.-M.; Fischer, S.F.; Browse, J. Trienoic Fatty Acids Are Required to Maintain Chloroplast Function at Low Temperatures. Plant Physiol. 2000, 124, 1697–1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iba, K. Acclimative response to temperature stress in higher plants: Approaches of gene engineering for temperature tolerance. Annu. Rev. Plant Biol. 2002, 53, 225–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham Thi, A.T.; Vieira Da Silva, J.; Mazliak, P. The role of membrane lipids in drought resistance of plants. Actual. Bot. 1990, 137, 99–114. [Google Scholar] [CrossRef] [Green Version]
- Ben Hamed, K.; Ben Youssef, N.; Ranieri, A.; Zarrouk, M.; Abdelly, C. Changes in content and fatty acid profiles of total lipids and sulfolipids in the halophyte Crithmum maritimum under salt stress. J. Plant Physiol. 2005, 162, 599–602. [Google Scholar] [CrossRef]
- Zhang, M.; Barg, R.; Yin, M.; Gueta-Dahan, Y.; Leikin-Frenkel, A.; Salts, Y.; Shabtai, S.; Ben-Hayyim, G. Modulated fatty acid desaturation via overexpression of two distinct omega-3 desaturases differentially alters tolerance to various abiotic stresses in transgenic tobacco cells and plants. Plant J. 2005, 44, 361–371. [Google Scholar] [CrossRef]
- Liu, S.; Wang, W.; Li, M.; Wan, S.; Sui, N. Antioxidants and unsaturated fatty acids are involved in salt tolerance in peanut. Acta Physiol. Plant 2017, 39, 207. [Google Scholar] [CrossRef]
- He, M.; Ding, N.-Z. Plant Unsaturated Fatty Acids: Multiple Roles in Stress Response. Front. Plant Sci. 2020, 11, 562785. [Google Scholar] [CrossRef]
- Mikami, K.; Murata, N. Membrane fluidity and the perception of environmental signals in cyanobacteria and plants. Prog. Lipid Res. 2003, 42, 527–543. [Google Scholar] [CrossRef]
- Jemal, F.; Zarrouk, M.; Ghorbal, M.H. Effect of cadmium on lipid composition of pepper. Biochem. Soc. Trans. 2000, 28, 907–910. [Google Scholar] [CrossRef]
- Howlett, N.G.; Avery, S.V. Induction of lipid peroxidation during heavy metal stress in Saccharomyces cerevisiae and influence of plasma membrane fatty acid unsaturation. Appl. Environ. Microbiol. 1997, 63, 2971–2976. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.-n.; Qu, D.; Shao, L.-l.; Yi, W.-j. Effects of sulfur fertilization on the contents of photosynthetic pigments and MDA under drought stress. Acta Bot. Sin. 2005, 25, 1579. [Google Scholar]
- Fatma, M.; Asgher, M.; Masood, A.; Khan, N.A. Excess sulfur supplementation improves photosynthesis and growth in mustard under salt stress through increased production of glutathione. Environ. Exp. Bot. 2014, 107, 55–63. [Google Scholar] [CrossRef]
- Terry, N. Effects of sulfur on the photosynthesis of intact leaves and isolated chloroplasts of sugar beets. Plant Physiol. 1976, 57, 477–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Individual SQDG m/z | SQDG Content at Varied S Treatments (mM) | Enrichment Factor (c1.5mM/c0mM) | ||
|---|---|---|---|---|
| S0 (%) 1 | S1 (%) 2 | S2 (%) 3 | ||
| 765 | 0.3 a,b | 0.2 a | 0.1 b | 3.1 |
| 787 | 0.2 | 0.3 | 0.3 | 7.9 |
| 789 | 0.2 | 0.3 | 0.3 | 9.2 |
| 791 | 0.6 | 0.6 | 0.5 | 5.0 |
| 792 | 1.1 | 1.2 | 1.1 | 6.0 |
| 793 | 12.4 a | 8.3 b | 6.9 b | 3.4 |
| 813 | 2.2 a | 3.7 b | 3.8 b | 10.4 |
| 815 | 34.5 a | 39.5 b | 41.9 b | 7.3 |
| 817 | 6.4 | 5.4 | 5.5 | 5.2 |
| 819 | 1.5 | 1.9 | 1.8 | 7.3 |
| 821 | 0.2 | 0.3 | 0.3 | 7.8 |
| 837 | 1.6 a,b | 2.5 a | 2.8 b | 10.4 |
| 839 | 22.2 | 18.5 | 18.5 | 5.0 |
| 841 | 5.6 | 6.0 | 6.4 | 6.9 |
| 843 | 2.6 | 2.7 | 2.4 | 5.4 |
| 845 | 1.4 | 1.2 | 1.1 | 4.8 |
| 847 | 0.9 | 0.9 | 0.6 | 4.1 |
| 849 | 0.3 | 0.3 | 0.2 | 2.9 |
| 855 | 2.1 | 2.4 | 2.0 | 5.9 |
| 555 | 3.5 | 3.6 | 3.5 | 5.9 |
| S Treatment Level (mM) | Chl a (mg g−1 DM) 1 | Chl b (mg g−1 DM) 2 | Chl a/b Ratio | Car (mg g−1 DM) 3 |
|---|---|---|---|---|
| S0 | 1.355 ± 0.094 a | 0.559 ± 0.051 b | 2.424 | 0.842 ± 0.038 c |
| S1 | 2.277 ± 0.083 a | 1.019 ± 0.049 b | 2.235 | 1.298 ± 0.013 b |
| S2 | 2.781 ± 0.28 a | 1.131 ± 0.108 b | 2.459 | 1.659 ± 0.047 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Körber, T.T.; Frantz, N.; Sitz, T.; Abdalla, M.A.; Mühling, K.H.; Rohn, S. Alterations of Content and Composition of Individual Sulfolipids, and Change of Fatty Acids Profile of Galactolipids in Lettuce Plants (Lactuca sativa L.) Grown under Sulfur Nutrition. Plants 2022, 11, 1342. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101342
Körber TT, Frantz N, Sitz T, Abdalla MA, Mühling KH, Rohn S. Alterations of Content and Composition of Individual Sulfolipids, and Change of Fatty Acids Profile of Galactolipids in Lettuce Plants (Lactuca sativa L.) Grown under Sulfur Nutrition. Plants. 2022; 11(10):1342. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101342
Chicago/Turabian StyleKörber, Tania T., Noah Frantz, Tobias Sitz, Muna A. Abdalla, Karl H. Mühling, and Sascha Rohn. 2022. "Alterations of Content and Composition of Individual Sulfolipids, and Change of Fatty Acids Profile of Galactolipids in Lettuce Plants (Lactuca sativa L.) Grown under Sulfur Nutrition" Plants 11, no. 10: 1342. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101342