
Spatial Distribution and Climate Warming Impact on Abies kawakamii Forest on a Subtropical Island


Abstract
:1. Introduction
2. Results
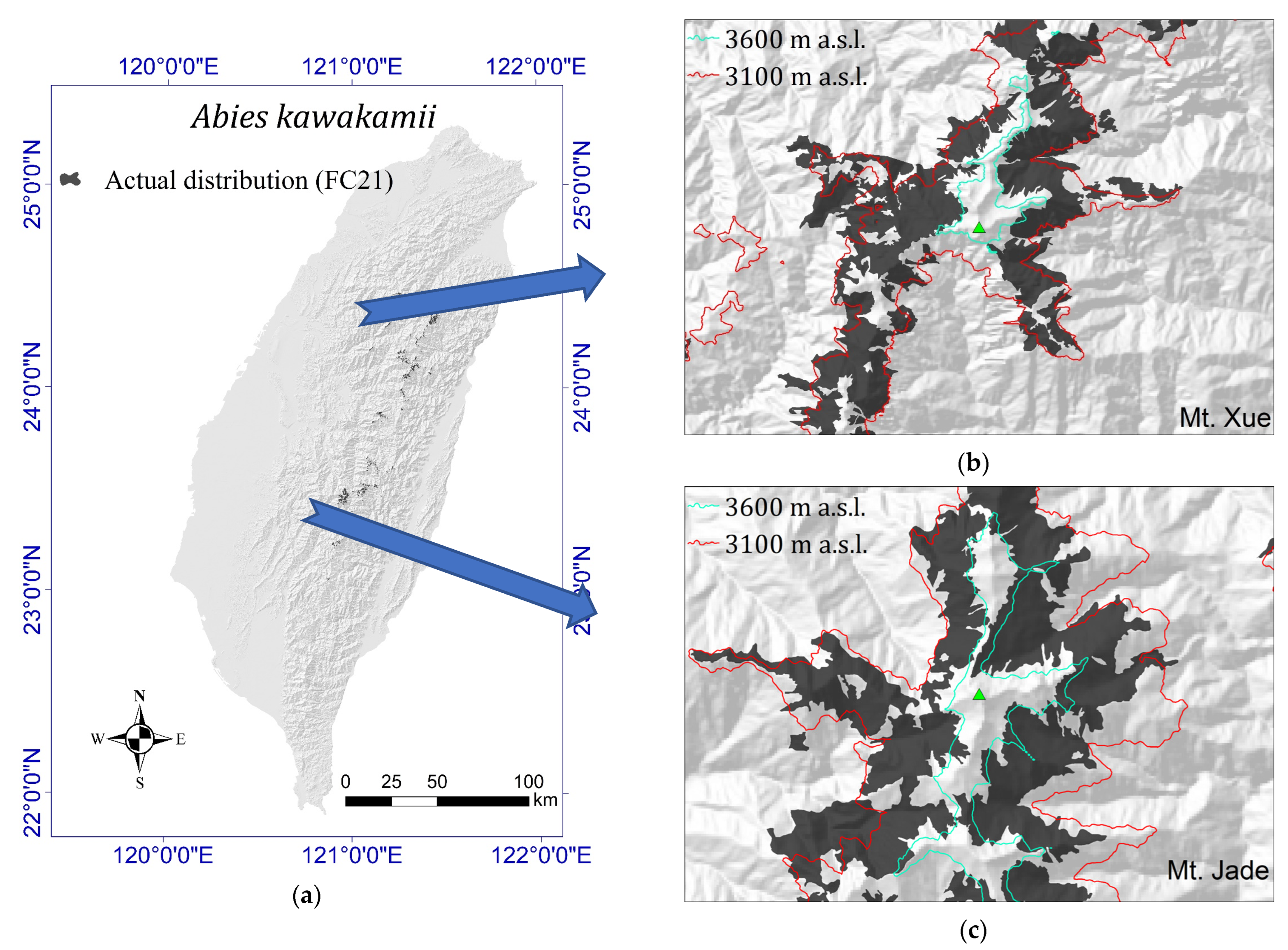
2.1. Current Actual Distribution of A. kawakamii
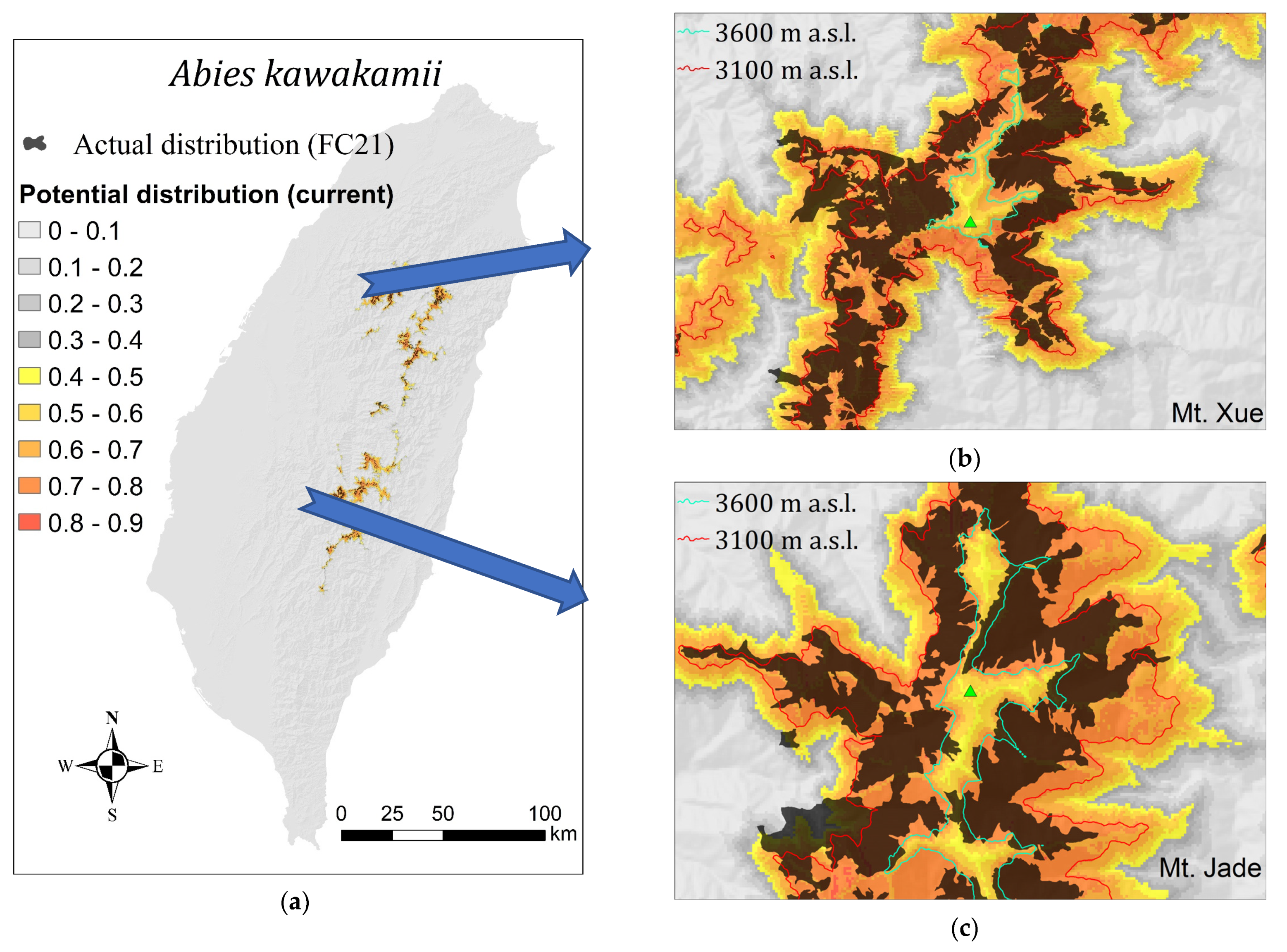
2.2. Potential Distribution Modeling Using MaxEnt

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Description and Reference | Percent Contribution | Permutation Importance |
|---|---|---|---|
| EWI | Effective warmth index [46] | 97.6 | 99.6 |
| PER | Potential evapotranspiration ratio [47] | 1.0 | 0.0 |
| bio15 | Precipitation seasonality [48] | 0.4 | 0.0 |
| WLS | Whole light sky space [49] | 0.4 | 0.1 |
| PWR | Ratio of winter half-year precipitation to Bio12 [23] | 0.2 | 0.1 |
| bio12 | Annual precipitation [48] | 0.2 | 0.0 |
| bio4 | Precipitation of driest month [48] | 0.1 | 0.0 |
| bio9 | Precipitation of coldest quarter [48] | 0.0 | 0.1 |
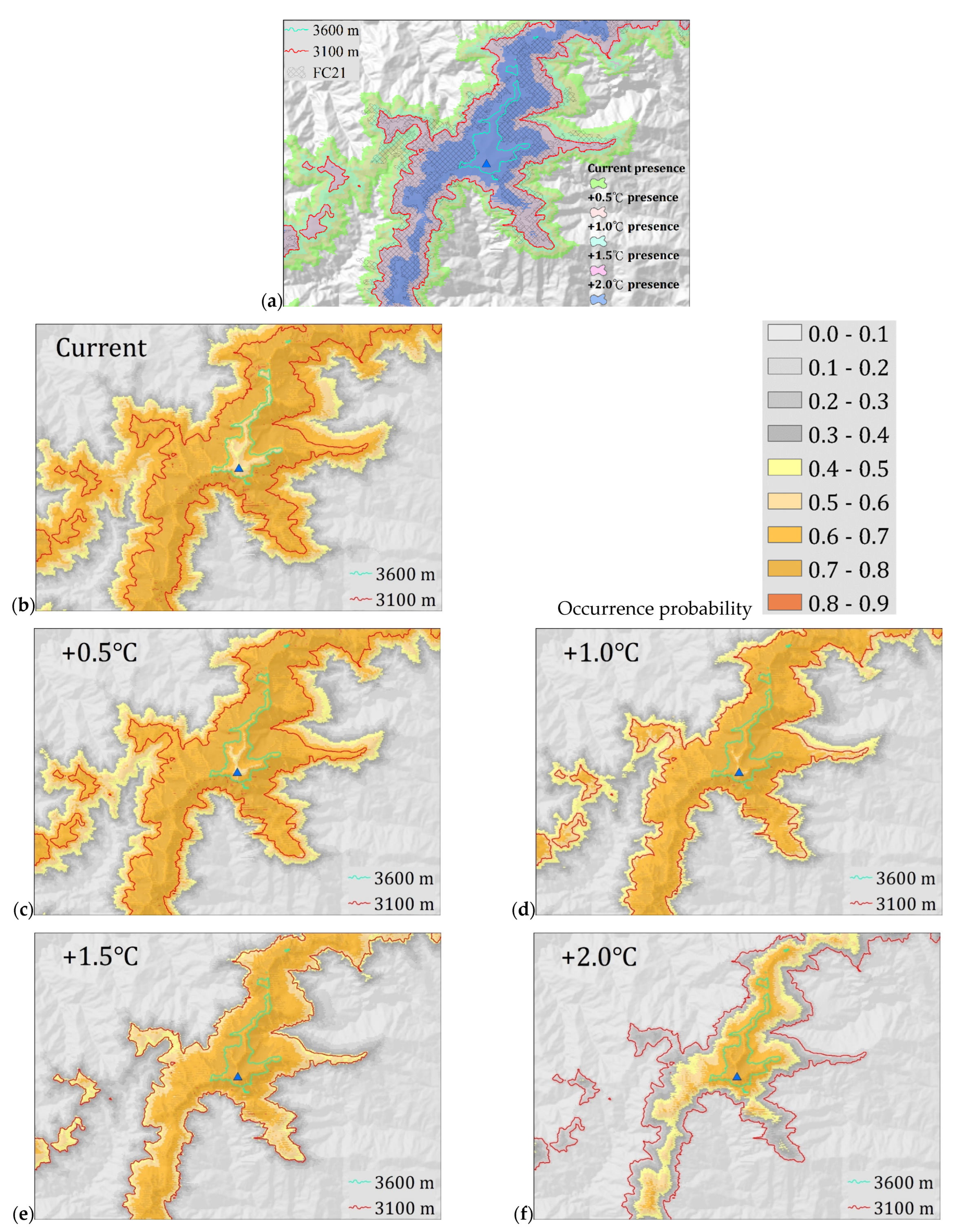
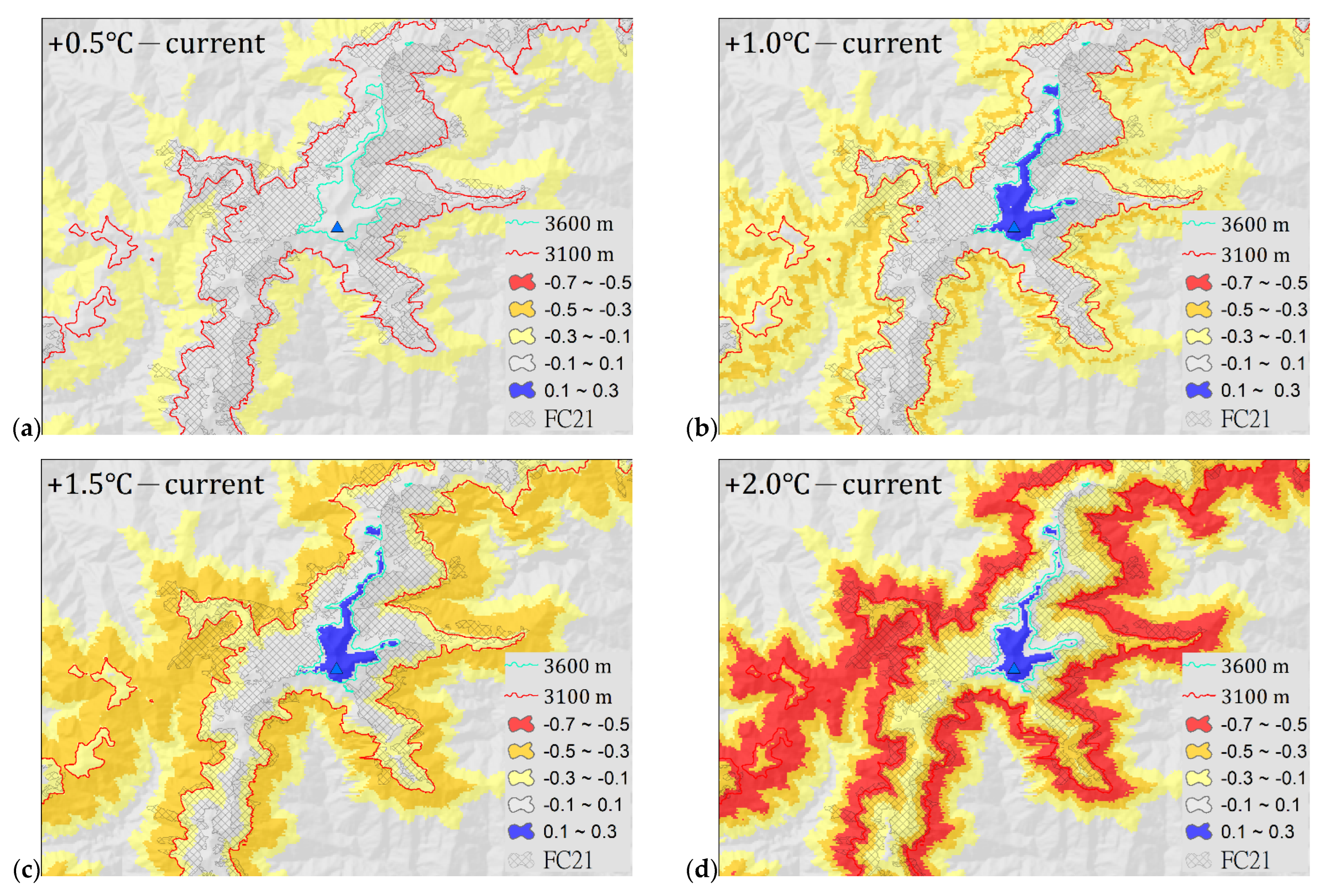
2.3. Potential Distribution under Climate Warming Scenarios
| Layer | Area (ha) | Habitat | Meaning and Reference |
|---|---|---|---|
| Actual distribution | 16,857 | Actual | Combining the ground survey of existing vegetation and the interpretation of remote sensing images to map the current actual distribution polygons of A. kawakamii-dominant forests [41] |
| Range of 3100–3600 m contours | 25,571 | Potential | The altitude range of A. kawakamii forests observed in the field [24,50] |
| Range of EWI (effective warmth index) = 12–32 | 20,922 | Potential | Use of the more ecologically significant thermal index (relative to the altitude) to estimate the temperature niche of the A. kawakamii forest belt [46] |
| Range of WI (warmth index) = 12–36 | 20,989 | Potential | Use of the more ecologically significant thermal index (relative to the altitude) to estimate the temperature niche of the A. kawakamii forest belt [50] |
| Presence range of potential distribution for the current climate | 73,151 | Potential | A species-suitable index value [51] or predicted probability of occurrence [16] of A. kawakamii individuals is higher than the presence threshold |
| Presence range of potential distribution for the +0.5 °C scenario | 51,323 | Potential | Predicted distribution of A. kawakamii by MaxEnt under the +0.5 °C scenario [52] |
| Presence range of potential distribution for the +1.0 °C scenario | 34,476 | Potential | Predicted distribution of A. kawakamii by MaxEnt under the +1.0 °C scenario [52] |
| Presence range of potential distribution for the +1.5 °C scenario | 22,106 | Potential | Predicted distribution of A. kawakamii by MaxEnt under the +1.5 °C scenario [52] |
| Presence range of potential distribution for the +2.0 °C scenario | 7338 | Potential | Predicted distribution of A. kawakamii by MaxEnt under the +2.0 °C scenario [52] |
3. Discussion
3.1. Current Actual Distribution of A. kawakamii
3.2. Potential Distribution Modeling by MaxEnt
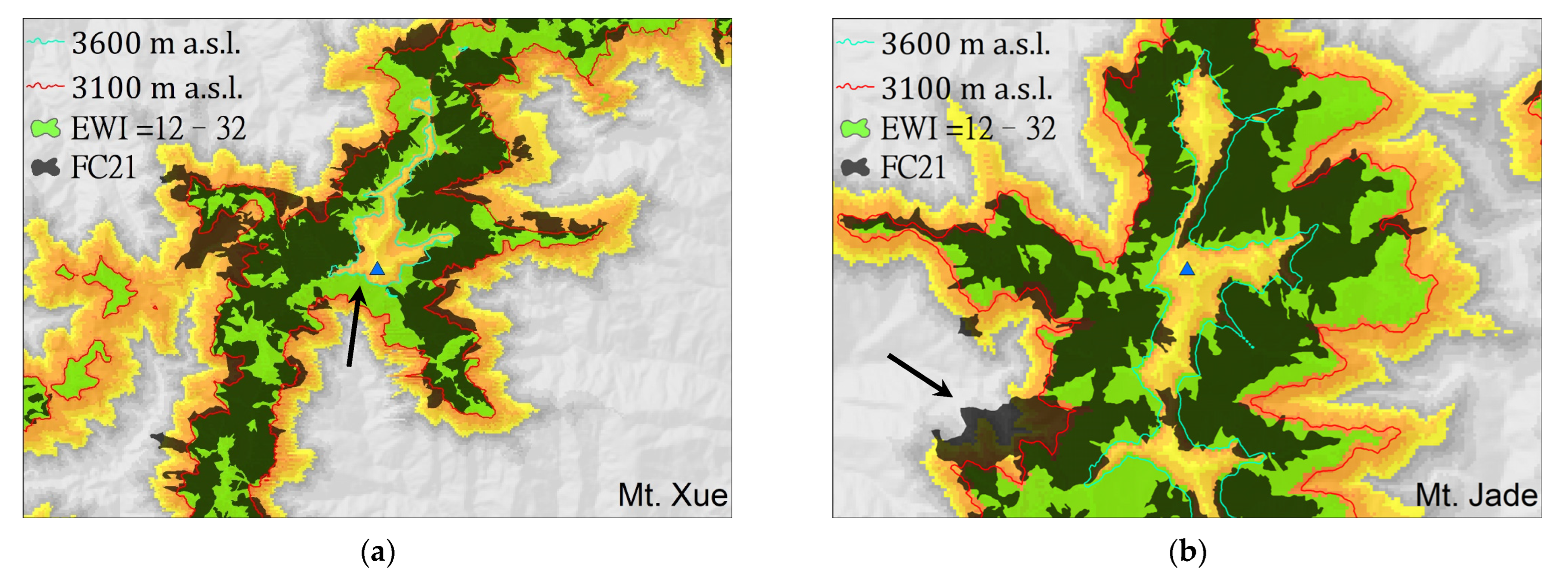
3.2.1. Range of EWI = 12–32 vs. Actual Distribution of A. kawakamii
3.2.2. Potential vs. Actual Distribution of A. kawakamii
3.2.3. Distributions of Dominant Stands vs. Individuals of A. kawakamii
3.2.4. The Influence of Non-Climatic Factors on the Distribution of A. kawakamii
3.3. Potential Distribution under Climate Warming Scenarios
3.4. Conservation and Challenges in the Future
4. Materials and Methods
4.1. Study Area and Target Species
4.2. SDM Calibration
4.3. Accuracy Evaluation
4.4. Threshold of Species Presence/Absence
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Franklin, J. Mapping Species Distributions: Spatial Inference and Prediction; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez-Meyer, E.; Nakamura, M.; Araújo, M.B. Ecological Niches and Geographic Distributions; Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Elith, J.; Franklin, J. Species Distribution Modeling. In Encyclopedia of Biodiversity, 2nd ed.; Levin, S.A., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2013; pp. 692–705. [Google Scholar]
- Franklin, J. Moving beyond static species distribution models in support of conservation biogeography: Moving beyond static species distribution models. Divers. Distrib. 2010, 16, 321–330. [Google Scholar] [CrossRef]
- Booth, T.H. Species distribution modelling tools and databases to assist managing forests under climate change. For. Ecol. Manag. 2018, 430, 196–203. [Google Scholar] [CrossRef]
- Westwood, R.; Westwood, A.R.; Hooshmandi, M.; Pearson, K.; LaFrance, K.; Murray, C. A field-validated species distribution model to support management of the critically endangered Poweshiek skipperling (Oarisma poweshiek) butterfly in Canada. Conserv. Sci. Pract. 2020, 2, e163. [Google Scholar] [CrossRef] [Green Version]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.T.; Regan, T.J.; Brotons, L.; McDonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting species distributions for conservation decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef]
- Lin, C.T.; Chiu, C.A. Comparison of predictor selection procedures in species distribution modeling: A case study of Fagus hayatae. Cerne 2020, 26, 172–182. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models: With Applications in R.; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- Mainali, K.P.; Warren, D.L.; Dhileepan, K.; McConnachie, A.; Strathie, L.; Hassan, G.; Karki, D.; Shrestha, B.B.; Parmesan, C. Projecting future expansion of invasive species: Comparing and improving methodologies for species distribution modeling. Glob. Chang. Biol. 2015, 21, 4464–4480. [Google Scholar] [CrossRef]
- Norberg, A.; Abrego, N.; Blanchet, F.G.; Adler, F.R.; Anderson, B.J.; Anttila, J.; Araújo, M.B.; Dallas, T.; Dunson, D.; Elith, J.; et al. A comprehensive evaluation of predictive performance of 33 species distribution models at species and community levels. Ecol. Monogr. 2019, 89, e01370. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Wan, J.Z.; Wang, C.J.; Tan, J.F.; Yu, F.H. Climatic niche divergence and habitat suitability of eight alien invasive weeds in China under climate change. Ecol. Evol. 2017, 7, 1541–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.T.; Gliottone, I.; Pham, M.P. Current and future predicting habitat suitability map of Cunninghamia konishii Hayata using MaxEnt model under climate change in Northern Vietnam. Eur. J. Ecol. 2021, 7, 1–17. [Google Scholar] [CrossRef]
- Kaky, E.; Nolan, V.; Alatawi, A.; Gilbert, F. A comparison between Ensemble and MaxEnt species distribution modelling approaches for conservation: A case study with Egyptian medicinal plants. Ecol. Inform. 2020, 60, 101150. [Google Scholar] [CrossRef]
- Liu, T.S. A Monograph of the Genus Abies; Department of Forestry, College of Agriculture National Taiwan University: Taipei, Taiwan, 1971. [Google Scholar]
- Xiang, X.; Cao, M.; Zhou, Z. Fossil history and modern distribution of the genus Abies (Pinaceae). Front. For. China 2007, 2, 355–365. [Google Scholar] [CrossRef]
- Xiang, Q.P. Abies fansipanensis—A new species of the genus Abies from Vietnam. J. Syst. Evol. 1997, 35, 356–359. [Google Scholar]
- Su, H.J. Studies on the climate and vegetation types of the natural forests in Taiwan (III): A scheme of geographical climatic regions. Q. J. China For. 1985, 18, 33–44. [Google Scholar]
- Chiu, C.A.; Lin, H.C.; Liao, M.C.; Tseng, Y.H.; Ou, C.H.; Lu, K.C.; Tzeng, H.Y. A physiognomic classification scheme of potential vegetation of Taiwan. Q. J. For. Res. 2008, 30, 89–112. [Google Scholar]
- Liao, M.C.; Tsai, S.T.; Wang, W.; Tzeng, H.Y.; Ou, C.H. Study on population structure of Abies kawakamii (Hayata) Ito in East Xue Trail of Xue Mountain. Q. J. For. Res. 2013, 35, 1–14. [Google Scholar]
- Li, P.H.; Liao, M.C.; Tzeng, H.Y.; Tseng, Y.H.; Yen, T.M. Applicability evaluation of tree volume equation for Abies kawakamii (Hayata) Ito based on stem analysis data in Taiwan. J. For. Res. 2021, 26, 336–343. [Google Scholar] [CrossRef]
- Wang, W.; Liao, M.C.; Tzeng, H.Y. Competition in Abies kawakamii forests at subtropical high mountain in Taiwan. PLoS ONE 2021, 16, e0254791. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Chiu, C.A.; Tsai, S.T.; Hsu, C.K.; Tzeng, H.Y.; Lu, K.C. Vegetation Research along the East Trail of Mt. Shei. Q. J. For. Res. 2010, 32, 15–34. [Google Scholar]
- Morley, P.; Donoghue, D.; Chen, J.C.; Jump, A.S. Integrating remote sensing and demography for more efficient and effective assessment of changing mountain forest distribution. Ecol. Inform. 2018, 43, 106–115. [Google Scholar] [CrossRef] [Green Version]
- Pauli, H.; Gottfried, M.; Grabherr, G. Effects of climate change on mountain–Ecosystemsupward shifting of alpine plants. World Resour. Rev. 1996, 8, 382–390. [Google Scholar]
- Winkler, D.E.; Lubetkin, K.C.; Carrell, A.A.; Jabis, M.D.; Yang, Y.; Kueppers, L.M. Responses of Alpine Plant Communities to Climate Warming. Ecosystem Consequences of Soil Warming; Academic Press: Cambridge, MA, USA, 2019; pp. 297–346. [Google Scholar]
- Randin, C.F.; Vuissoz, G.; Liston, G.E.; Vittoz, P.; Guisan, A. Introduction of snow and geomorphic disturbance variables into predictive models of alpine plant distribution in the Western Swiss Alps. Arct. Antarct. Alp. Res. 2009, 41, 347–361. [Google Scholar] [CrossRef]
- Bell, D.M.; Bradford, J.B.; Lauenroth, W.K. Mountain landscapes offer few opportunities for high-elevation tree species migration. Glob. Chang. Biol. 2014, 20, 1441–1451. [Google Scholar] [CrossRef]
- Maxwell, C.J.; Scheller, R.M. Identifying Habitat Holdouts for High Elevation Tree Species Under Climate Change. Front. For. Glob. Chang. 2020, 2, 94. [Google Scholar] [CrossRef]
- Koo, K.A.; Kong, W.S.; Park, S.U.; Lee, J.H.; Kim, J.; Jung, H. Sensitivity of Korean fir (Abies koreana Wils.), a threatened climate relict species, to increasing temperature at an island subalpine area. Ecol. Model. 2017, 353, 5–16. [Google Scholar] [CrossRef]
- Shao, Y.Z.; Zhang, X.C.; Phan, L.K.; Xiang, Q.P. Elevation shift in Abies Mill. (Pinaceae) of subtropical and temperate China and Vietnam—Corroborative evidence from cytoplasmic DNA and ecological niche modeling. Front. Plant Sci. 2017, 8, 578. [Google Scholar] [CrossRef] [Green Version]
- Altman, J.; Treydte, K.; Pejcha, V.; Cerny, T.; Petrik, P.; Srutek, M.; Song, J.S.; Trouet, V.; Dolezal, J. Tree growth response to recent warming of two endemic species in Northeast Asia. Clim. Chang. 2020, 162, 1345–1364. [Google Scholar] [CrossRef]
- Hsu, H.H.; Chen, C.T. Observed and projected climate change in Taiwan. Meteorol. Atmos. Phys. 2002, 79, 87–104. [Google Scholar] [CrossRef]
- Jump, A.S.; Huang, T.J.; Chou, C.H. Rapid altitudinal migration of mountain plants in Taiwan and its implications for high altitude biodiversity. Ecography 2012, 35, 204–210. [Google Scholar] [CrossRef]
- O’Sullivan, K.S.W.; Ruiz-Benito, P.; Chen, J.C.; Jump, A.S. Onward but not always upward: Individualistic elevational shifts of tree species in subtropical montane forests. Ecography 2021, 44, 112–123. [Google Scholar] [CrossRef]
- Chiou, C.R.; Hsieh, C.F.; Wang, J.C.; Chen, M.Y.; Liu, H.Y.; Yeh, C.L.; Yang, S.Z.; Chen, T.Y.; Hsia, Y.J.; Song, G.Z.M. The first national vegetation inventory in Taiwan. Taiwan J. For. Res. 2009, 24, 295–302. [Google Scholar]
- Parolo, G.; Rossi, G. Upward migration of vascular plants following a climate warming trend in the Alps. Basic Appl. Ecol. 2008, 9, 100–107. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting thresholds of occurrence in the prediction of species distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Cao, Y.; DeWalt, R.E.; Robinson, J.L.; Tweddale, T.; Hinz, L.; Pessino, M. Using Maxent to model the historic distributions of stonefly species in Illinois streams: The effects of regularization and threshold selections. Ecol. Model. 2013, 259, 30–39. [Google Scholar] [CrossRef]
- Trisurat, Y.; Bhumpakphan, N. Effects of land use and climate change on Siamese Eld’s Deer (Rucervus eldii siamensis) distribution in the transboundary conservation area in Thailand, Cambodia, and Lao PDR. Front. Environ. Sci. 2018, 6, 35. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.A.; Lin, P.H.; Hsu, C.K.; Shen, Z.H. A novel thermal index improves prediction of vegetation zones: Associating temperature sum with thermal seasonality. Ecol. Indic. 2012, 23, 668–674. [Google Scholar] [CrossRef]
- Holdrige, L.R. Life Zone Ecology; Tropical Science Center: San José, Costa Rica, 1967; p. 148. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Lai, Y.J.; Chou, M.D.; Lin, P.H. Parameterization of topographic effect on surface solar radiation. J. Geophys. Res. Atmos. 2010, 115, D01104. [Google Scholar] [CrossRef] [Green Version]
- Su, H.J. Studies on the climate and vegetation types of the natural forests in Taiwan (II): Altitudinal vegetation zones in relation to temperature gradient. Q. J. Chin. For. 1984, 17, 57–73. [Google Scholar]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Ngarega, B.K.; Masocha, V.F.; Schneider, H. Forecasting the effects of bioclimatic characteristics and climate change on the potential distribution of Colophospermum mopane in southern Africa using Maximum Entropy (Maxent). Ecol. Inform. 2021, 65, 101419. [Google Scholar] [CrossRef]
- Liu, Y.C.; Lu, F.Y.; Ou, C.H. Trees of Taiwan; Monographic Publication No. 7; College of Agriculture, National Chung-Shing University: Taichung, Taiwan, 1988. [Google Scholar]
- Taiwan Forest Bureau/Anonymous. The Third Forest Resources and Land Use Inventory in Taiwan; Taiwan Forest Bureau, Province Council of Agriculture: Taipei, Taiwan, 1995.
- Randin, C.F.; Engler, R.; Normand, S.; Zappa, M.; Zimmermann, N.E.; Pearman, P.B.; Vittoz, P.; Thuiller, W.; Guisan, A. Climate change and plant distribution: Local models predict high-elevation persistence. Glob. Chang. Biol. 2009, 15, 1557–1569. [Google Scholar] [CrossRef] [Green Version]
- Porfirio, L.L.; Harris, R.M.; Lefroy, E.C.; Hugh, S.; Gould, S.F.; Lee, G.; Bindoff, N.L.; Mackey, B. Improving the use of species distribution models in conservation planning and management under climate change. PLoS ONE 2014, 9, e113749. [Google Scholar] [CrossRef] [Green Version]
- Barve, N.; Barve, V.; Jiménez-Valverde, A.; Lira-Noriega, A.; Maher, S.P.; Peterson, A.T.; Soberón, J.; Villalobos, F. The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecol. Model. 2011, 222, 1810–1819. [Google Scholar] [CrossRef]
- Whittaker, R.H. Evolution and Measurement of Species Diversity. Taxon 1972, 21, 213–251. [Google Scholar] [CrossRef] [Green Version]
- Chen, I.C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Zu, K.; Wang, Z.; Zhu, X.; Lenoir, J.; Shrestha, N.; Lyu, T.; Luo, A.; Li, Y.; Ji, C.; Peng, S.; et al. Upward shift and elevational range contractions of subtropical mountain plants in response to climate change. Sci. Total Environ. 2021, 783, 146896. [Google Scholar] [CrossRef] [PubMed]
- Rabasa, S.G.; Granda, E.; Benavides, R.; Kunstler, G.; Espelta, J.M.; Ogaya, R.; Peñuelas, J.; Scherer-Lorenzen, M.; Gil, W.; Grodzki, W.; et al. Disparity in elevational shifts of E uropean trees in response to recent climate warming. Glob. Chang. Biol. 2013, 19, 2490–2499. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, A.; Meneses, R.I.; Rabatel, A.; Soruco, A.; Dangles, O.; Anthelme, F. Time lag between glacial retreat and upward migration alters tropical alpine communities. Perspect. Plant Ecol. Evol. Syst. 2018, 30, 89–102. [Google Scholar] [CrossRef]
- Schickhoff, U.; Bobrowski, M.; Böhner, J.; Bürzle, B.; Chaudhary, R.P.; Gerlitz, L.; Heyken, H.; Lange, J.; Müller, M.; Scholten, T.; et al. Do Himalayan treelines respond to recent climate change? An evaluation of sensitivity indicators. Earth Syst. Dyn. 2015, 6, 245–265. [Google Scholar] [CrossRef]
- McCain, A.H.; Scharpf, R.F. Phytophthora shoot blight and canker disease of Abies spp. Plant Dis. 1986, 70, 1036–1037. [Google Scholar] [CrossRef]
- Tsankov, G.; Mirchev, P.; Ovcharov, D. Insect pests and their role in the decline and dying of silver fir (Abies alba) in Bulgaria. Nauka Za Gorata 1994, 31, 23–33. [Google Scholar]
- Motta, R. Impact of wild ungulates on forest regeneration and tree composition of mountain forests in the Western Italian Alps. For. Ecol. Manag. 1996, 88, 93–98. [Google Scholar] [CrossRef]
- Klopcic, M.; Jerina, K.; Boncina, A. Long-term changes of structure and tree species composition in Dinaric uneven-aged forests: Are red deer an important factor? Eur. J. For. Res. 2010, 129, 277–288. [Google Scholar] [CrossRef]
- Bonner, F.T. Storage of Seeds. In The Woody Plant Seed Material. Agriculture Handbook 727; USDA, Forest Service: Washington, DC, USA, 2008; pp. 85–96. [Google Scholar]
- Krajňáková, J.; Gömöry, D.; Häggman, H. Biotechnology tools for conservation of the biodiversity of European and Mediterranean Abies species. In Biotechnology and Biodiversity. Sustainable Development and Biodiversity; Ahuja, M.R., Ramawat, K.G., Eds.; Springer International Publishing: Cham, Switzerland, 2014; Volume 14, pp. 287–310. [Google Scholar]
- Shih, F.L.; Hwang, S.Y.; Cheng, Y.P.; Lee, P.F.; Lin, T.P. Uniform genetic diversity, low differentiation, and neutral evolution characterize contemporary refuge populations of Taiwan fir (Abies kawakamii, Pinaceae). Am. J. Bot. 2007, 94, 194–202. [Google Scholar] [CrossRef] [Green Version]
- Munier, A.; Hermanutz, L.; Jacobs, J.D.; Lewis, K. The interacting effects of temperature, ground disturbance, and herbivory on seedling establishment: Implications for treeline advance with climate warming. Plant Ecol. 2010, 210, 19–30. [Google Scholar] [CrossRef]
- Greenwood, S.; Chen, J.C.; Chen, C.T.; Jump, A.S. Strong topographic sheltering effects lead to spatially complex treeline advance and increased forest density in a subtropical mountain region. Glob. Chang. Biol. 2014, 20, 3756–3766. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.A.; Lin, P.H.; Lu, K.C. GIS-based tests for quality control of meteorological data and spatial interpolation of climate data: A case study in mountainous Taiwan. Mt. Res. Dev. 2009, 29, 339–349. [Google Scholar] [CrossRef]
- Sangermano, F.; Eastman, J.R. A GIS framework for the refinement of species geographic ranges. Int. J. Geogr. Inf. Sci. 2012, 26, 39–55. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E.; Elith, J.; Graham, C.H.; Phillips, S.; Peterson, A.T. What matters for predicting the occurrences of trees: Techniques, data, or species’ characteristics? Ecol. Monogr. 2007, 77, 615–630. [Google Scholar] [CrossRef] [Green Version]
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A.; NCEAS Predicting Species Distributions Working Group. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Grimmett, L.; Whitsed, R.; Horta, A. Presence-only species distribution models are sensitive to sample prevalence: Evaluating models using spatial prediction stability and accuracy metrics. Ecol. Model. 2020, 431, 109194. [Google Scholar] [CrossRef]
- Austin, M.P.; Van Niel, K.P. Improving species distribution models for climate change studies: Variable selection and scale: Species distribution models for climate change studies: Variable selection and scale. J. Biogeogr. 2011, 38, 1–8. [Google Scholar] [CrossRef]
- Fourcade, Y.; Besnard, A.G.; Secondi, J. Paintings predict the distribution of species, or the challenge of selecting environmental predictors and evaluation statistics. Glob. Ecol. Biogeogr. 2018, 27, 245–256. [Google Scholar] [CrossRef]
- Global Biodiversity Information Facility. Available online: https://www.gbif.org/ (accessed on 5 December 2021).
- Kriticos, D.J.; Webber, B.L.; Leriche, A.; Ota, N.; Macadam, I.; Bathols, J.; Scott, J.K. CliMond: Global high resolution historical and future scenario climate surfaces for bioclimatic modelling. Methods Ecol. Evol. 2012, 3, 53–64. [Google Scholar] [CrossRef]
- Global Climatologies for Bioclimatic Modelling. Available online: https://www.climond.org (accessed on 20 December 2021).
- Global Climate and Weather Data. Available online: https://www.worldclim.org (accessed on 6 December 2021).
- Climatologies at High Resolution for the Earth’s Land Surface Areas. Available online: https://www.chelsa-climate.org (accessed on 3 January 2022).
- Steven, J.P.; Dudik, M.; Schapire, R.E. Maxent Software for Modeling Species Niches and Distributions (Version 3.4.1). Available online: https://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 20 December 2021).
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Krause, C.M.; Cobb, N.S.; Pennington, D.D. Range shifts under future scenarios of climate change: Dispersal ability matters for Colorado Plateau endemic plants. Nat. Areas J. 2015, 35, 428–438. [Google Scholar] [CrossRef]
- Lobo, J.M.; Jiménez-Valverde, A.; Real, R. AUC: A misleading measure of the performance of predictive distribution models. Glob. Ecol. Biogeogr. 2008, 17, 145–151. [Google Scholar] [CrossRef]






| Elevation Zone (m) | Area (ha) | % of Total Area |
|---|---|---|
| 2200–2299 | 0 | 0.0% |
| 2300–2399 | 8 | 0.0% |
| 2400–2499 | 26 | 0.2% |
| 2500–2599 | 118 | 0.7% |
| 2600–2699 | 284 | 1.7% |
| 2700–2799 | 501 | 3.0% |
| 2800–2899 | 986 | 5.9% |
| 2900–2999 | 1874 | 11.1% |
| 3000–3099 | 3262 | 19.4% |
| 3100–3199 | 3736 | 22.2% |
| 3200–3299 | 2770 | 16.4% |
| 3300–3399 | 1666 | 9.9% |
| 3400–3499 | 951 | 5.6% |
| 3500–3599 | 489 | 2.9% |
| 3600–3699 | 168 | 1.0% |
| 3700–3799 | 15 | 0.1% |
| 3800–3899 | 0 | 0.0% |
| Total | 16,857 | 100.0% |
| Scenario | Actual Distribution (16,857 ha) | Potential Distribution (73,151 ha) |
|---|---|---|
| +0.5 °C | Low loss: 3991 ha Medium loss: 0 ha High loss: 0 ha The total area of loss: 3991 ha (23.7%) | Low loss: 35,368 ha Medium loss: 0 ha High loss: 0 ha The total area of loss: 35,368 ha (48.3%) Low gain: 276 ha |
| +1.0 °C | Low loss: 7704 ha Medium loss: 2153 ha High loss: 0 ha Total area of loss: 9857 ha (58.5%) | Low loss: 33,644 ha Medium loss: 22,571 ha High loss: 0 ha Total area of loss: 56,216 ha (76.8%) Low gain: 580 ha |
| +1.5 °C | Low loss: 5925 ha Medium loss: 7102 ha High loss: 59 ha Total area of loss: 13,086 ha (77.6%) | Low loss: 13,238 ha Medium loss: 50,624 ha High loss: 185 ha Total area of loss: 64,047 ha (87.6%) Low gain: 545 ha |
| +2.0 °C | Low loss: 2923 ha Medium loss: 4588 ha High loss: 8709 ha Total area of loss: 16,220 ha (96.2%) | Low loss: 4992 ha Medium loss: 25,904 ha High loss: 40,282 ha Total area of loss: 71,178 ha (97.3%) Low gain: 475 ha |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiu, C.-A.; Tzeng, H.-Y.; Lin, C.-T.; Chang, K.-C.; Liao, M.-C. Spatial Distribution and Climate Warming Impact on Abies kawakamii Forest on a Subtropical Island. Plants 2022, 11, 1346. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101346
Chiu C-A, Tzeng H-Y, Lin C-T, Chang K-C, Liao M-C. Spatial Distribution and Climate Warming Impact on Abies kawakamii Forest on a Subtropical Island. Plants. 2022; 11(10):1346. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101346
Chicago/Turabian StyleChiu, Ching-An, Hsy-Yu Tzeng, Cheng-Tao Lin, Kun-Cheng Chang, and Min-Chun Liao. 2022. "Spatial Distribution and Climate Warming Impact on Abies kawakamii Forest on a Subtropical Island" Plants 11, no. 10: 1346. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101346