
Induction of Somatic Embryogenesis in Tamarillo (Solanum betaceum Cav.) Involves Increases in the Endogenous Auxin Indole-3-Acetic Acid



Abstract
:1. Introduction
2. Results
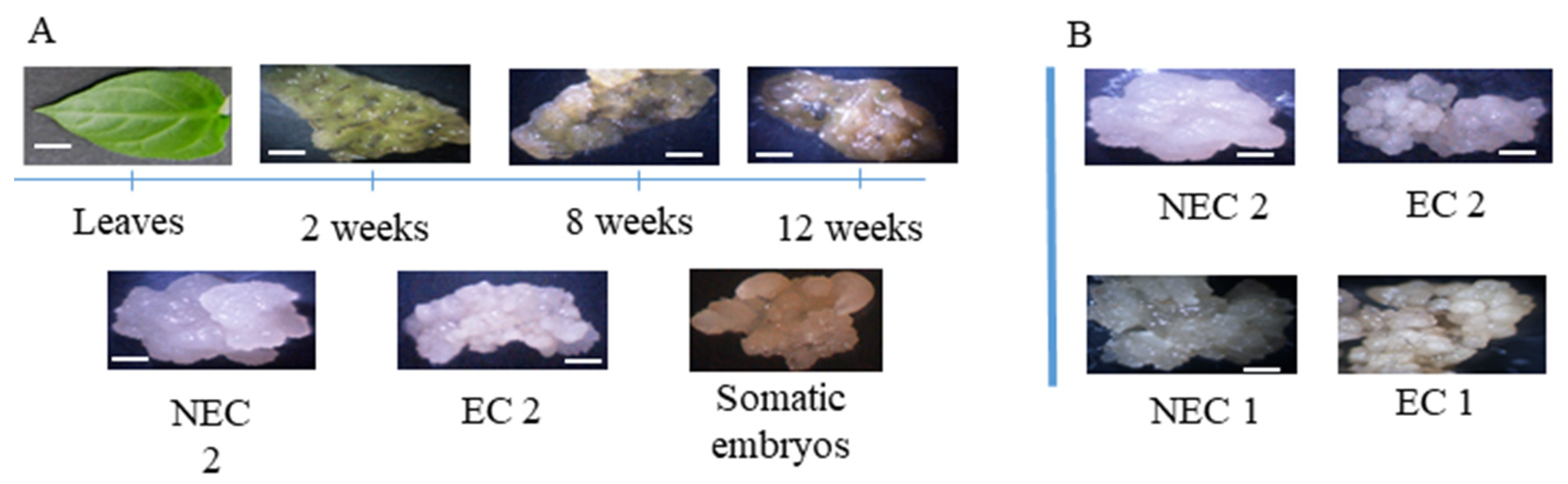
2.1. Somatic Embryogenesis Induction and Callus Proliferation
2.2. IAA Location on the Induction of SE
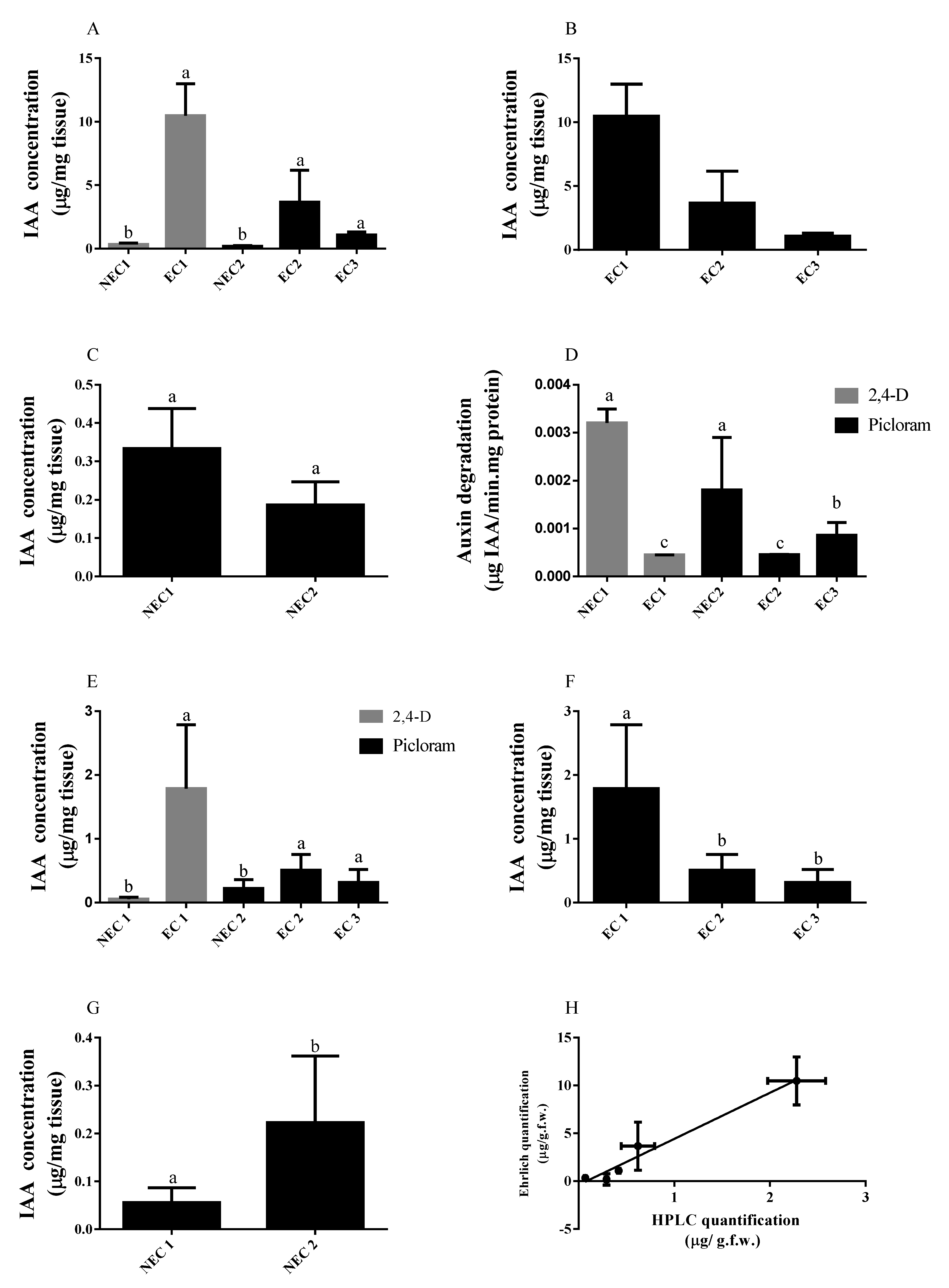
2.3. IAA Levels Increment in EC
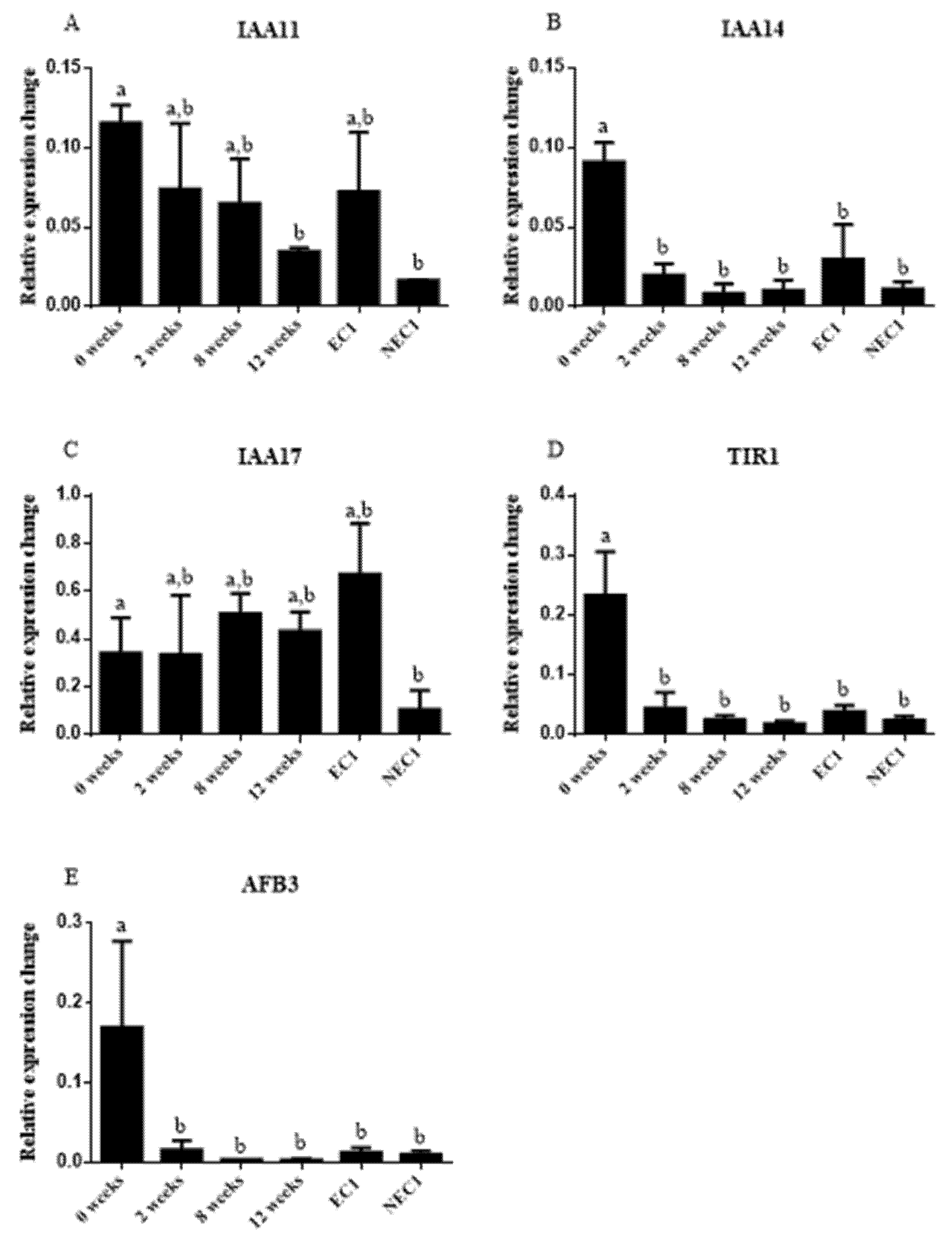
2.4. Auxin-Related Gene Expression Relates with IAA Levels during SE Induction
3. Discussion
3.1. IAA Distribution Is Important for Somatic Embryo Conversion
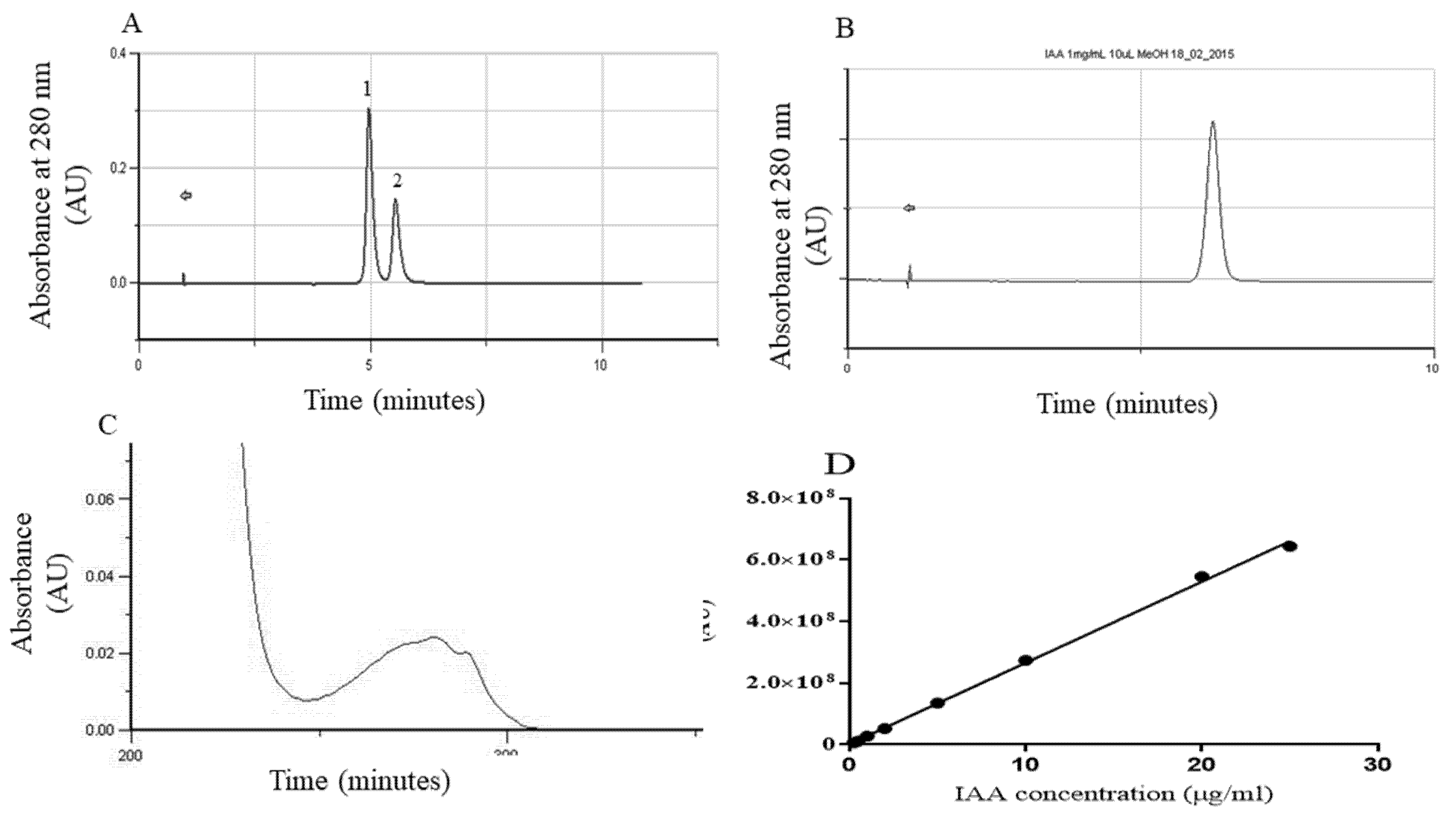
3.2. IAA Quantification through SE Induction
3.3. Immunolocalization of IAA during SE Induction
3.4. Auxin-Related Gene Differential Expression Is Influenced by IAA Endogenous Levels
4. Materials and Methods
4.1. Somatic Embryogenesis Induction from Leaf Segments of In Vitro Propagated Shoots
4.2. Embryogenic and Non-Embryogenic Calli Subcultures and Maintenance
4.3. Quantification of IAA
4.3.1. Ehrlich Reaction
4.3.2. HPLC
4.4. Immunohistochemistry IAA Localization
4.5. Expression of Auxin-Related Genes
4.6. Statistical and Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kahia, J.; Sallah, P.K.; Diby, L.; Kouame, C.; Kirika, M.; Niyitegeka, S.; Asiimwe, T. A novel regeneration system for tamarillo (Cyphomandra betacea) via organogenesis from hypocotyl, leaf, and root explants. HortScience 2015, 50, 1375–1378. [Google Scholar] [CrossRef] [Green Version]
- Correia, S.; Lopes, M.L.; Canhoto, J.M. Somatic embryogenesis induction system for cloning an adult Cyphomandra betacea (Cav.) Sendt. (tamarillo). Trees 2011, 25, 1009–1020. [Google Scholar] [CrossRef]
- Diep, T.T.; Rush, E.C.; Yoo, M.J.Y. Tamarillo (Solanum betaceum Cav.): A Review of Physicochemical and Bioactive Properties and Potential Applications. Food Rev. Int. 2020, 1–25. [Google Scholar] [CrossRef]
- Prohens, J.; Nuez, F. The tamarillo (Cyphomandra betacea): A review of a promising small fruit crop. Small Fruits Rev. 2001, 1, 43–68. [Google Scholar] [CrossRef]
- Kou, M.-C.; Yen, J.-H.; Hong, J.-T.; Wang, C.-L.; Lin, C.-W.; Wu, M.-J. Cyphomandra betacea Sendt. phenolics protect LDL from oxidation and PC12 cells from oxidative stress. LWT-Food Sci. Technol. 2009, 42, 458–463. [Google Scholar] [CrossRef]
- Correia, S.; Canhoto, J. Biotechnology of tamarillo (Cyphomandra betacea): From in vitro cloning to genetic transformation. Sci. Hortic. (Amst.) 2012, 148, 61–168. [Google Scholar] [CrossRef]
- Mossop, D.W. Isolation, purification, and properties of tamarillo mosaic virus, a member of the potato virus Y group. N. Z. J. Agric. Res. 1977, 20, 535–541. [Google Scholar] [CrossRef] [Green Version]
- Correia, S.; Canhoto, J. Somatic embryogenesis of tamarillo (Solanum betaceum Cav.). In Vegetative Propagation of Forest Trees; Jain, S., Gupta, P., Eds.; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Cohen, D.; Elliott, D. Micropropagation methods for blueberries and tamarillos [Cyphomandra betacea, tree-tomato]. In Combined Proceedings International Plant Propagators’ Society; International Plant Propagators’ Society: Monroe, CT, USA, 1979. [Google Scholar]
- Guimarães, M.L.; Tomé, M.C.; Cruz, G.S. Cyphomandra betacea (Cav.) Sendtn. (Tamarillo) BT-Trees IV; Bajaj, Y.P.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 120–137. [Google Scholar]
- Obando, M.; Goreux, A.; Jordan, M. In vitro regeneration of Cyphomandra betacea (Tamarillo) and Andean fruit species. Cienc. Investig. Agrar. 1992, 19, 125–130. [Google Scholar] [CrossRef]
- Correia, S.; Canhoto, J. Somatic Embryogenesis of Tamarillo (Solanum betaceum Cav.). In Step Wise Protocols for Somatic Embryogenesis on Important Woody Plants; Jain, S.M., Gupta, P., Eds.; Springer International Publishing: New York, NY, USA, 2018; Volume 2, pp. 171–179. [Google Scholar]
- Méndez-Hernández, H.A.; Ledezma-Rodriguez, M.; Avilez-Montalvo, R.N.; Juarez-Gomez, Y.L.; Skeete, A.; Avilez-Montalvo, J.; De-la-Pena, C.; Loyola-Vargas, V.M. Signaling Overview of Plant Somatic Embryogenesis. Front. Plant Sci. 2019, 10, 77. [Google Scholar] [CrossRef] [Green Version]
- Stasolla, C.; Yeung, E.C. Recent advances in conifer somatic embryogenesis: Improving somatic embryo quality. Plant Cell. Tissue Organ Cult. 2003, 74, 15–35. [Google Scholar] [CrossRef]
- Jiménez, V.M. Regulation of in vitro somatic embryogenesis with emphasis on the role of endogenous hormones. Rev. Bras. Fisiol. Veg. 2001, 13, 196–223. [Google Scholar] [CrossRef] [Green Version]
- Guimarães, M.L.S.; Cruz, G.S.; Montezuma-De-Carvalho, J.M. Somatic embryogenesis and plant regeneration in Cyphomandra betacea (Cav.) Sendt. Plant Cell. Tissue Organ Cult. 1988, 15, 161–167. [Google Scholar] [CrossRef]
- Lopes, M.L.; Ferreira, M.R.; Carloto, J.M.; Cruz, G.S.; Canhoto, J.M. Somatic Embryogenesis Induction in Tamarillo (Cyphomandra betacea). In Somatic Embryogenesis in Woody Plants; Springer: Dordrecht, The Netherlands, 2000; pp. 433–455. [Google Scholar]
- Canhoto, J.M.; Lopes, M.L.; Cruz, G.S. Protocol for Somatic Embryogenesis: Tamarillo (Cyphomandra betacea (Cav.) Sendtn.)). In Protocol for Somatic Embryogenesis in Woody Plants; Jain, P., Gupta, S.M., Eds.; Springer: Dordrecht, The Netherlands, 2005. [Google Scholar]
- Raghavan, V. Role of 2,4-dichlorophenoxyacetic acid (2,4-D) in somatic embryogenesis on cultured zygotic embryos of Arabidopsis: Cell expansion, cell cycling, and morphogenesis during continuous exposure of embryos to 2,4-D. Am. J. Bot. 2004, 91, 1743–1756. [Google Scholar] [CrossRef] [PubMed]
- Winnicki, K. The Winner Takes It All: Auxin-The Main Player during Plant Embryogenesis. Cells 2020, 9, 606. [Google Scholar] [CrossRef] [Green Version]
- Gaj, M.D. Direct somatic embryogenesis as a rapid and efficient system for in vitro regeneration of Arabidopsis thaliana. Plant Cell. Tissue Organ Cult. 2001, 64, 39–46. [Google Scholar] [CrossRef]
- Hazubska-Przybył, T.; Ratajczak, E.; Obarska, A.; Pers-Kamczyc, E. Different roles of auxins in somatic embryogenesis efficiency in two picea species. Int. J. Mol. Sci. 2020, 21, 3394. [Google Scholar] [CrossRef]
- Wójcik, A.M.; Wójcikowska, B.; Gaj, M.D. Current perspectives on the auxin-mediated genetic network that controls the induction of somatic embryogenesis in plants. Int. J. Mol. Sci. 2020, 21, 1333. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Zhang, X.; Yuan, D.; Jin, F.; Zhang, Y.; Xu, J. Transcript profiling reveals complex auxin signalling pathway and transcription regulation involved in dedifferentiation and redifferentiation during somatic embryogenesis in cotton. BMC Plant Biol. 2012, 12, 110. [Google Scholar] [CrossRef] [Green Version]
- Fehér, A. Somatic embryogenesis—Stress-induced remodeling of plant cell fate. Biochim. Biophys. Acta-Gene Regul. Mech. 2015, 1849, 385–402. [Google Scholar] [CrossRef]
- Liu, C.M.; Xu, Z.H.; Chua, N.H. Auxin polar transport is essential for the establishment of bilateral symmetry during early plant embryogenesis. Plant Cell 1993, 5, 621–630. [Google Scholar] [CrossRef]
- Vondrakova, Z.; Dobrev, P.I.; Pesek, B.; Fischerova, L.; Vagner, M.; Motyka, V. Profiles of endogenous phytohormones over the course of norway spruce somatic embryogenesis. Front. Plant Sci. 2018, 9, 1283. [Google Scholar] [CrossRef] [PubMed]
- Möller, B.; Weijers, D. Auxin control of embryo patterning. Cold Spring Harb. Perspect. Biol. 2009, 1, a001545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, S.K.; Das, A.K.; Gantait, S.; Gurel, S.; Gurel, E. Influence of auxin and its polar transport inhibitor on the development of somatic embryos in Digitalis trojana. 3 Biotech 2018, 8, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teale, W.; Palme, K. Naphthylphthalamic acid and the mechanism of polar auxin transport. J. Exp. Bot. 2018, 69, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Jenik, P.D.; Gillmor, C.S.; Lukowitz, W. Embryonic patterning in Arabidopsis thaliana. Annu. Rev. Cell Dev. Biol. 2007, 23, 207–236. [Google Scholar] [CrossRef]
- Jiang, K.; Asami, T. Chemical regulators of plant hormones and their applications in basic research and agriculture. Biosci. Biotechnol. Biochem. 2018, 82, 1265–1300. [Google Scholar] [CrossRef] [Green Version]
- Stuepp, C.A.; Wendling, I.; Trueman, S.J.; Koehler, H.S.; Zuffellato-Ribas, K.C. The use of auxin quantification for understanding clonal tree propagation. Forests 2017, 8, 27. [Google Scholar] [CrossRef]
- Ayil-Gutiérrez, B.; Galaz-Ávalos, R.M.; Peña-Cabrera, E.; Loyola-Vargas, V.M. Dynamics of the concentration of IAA and some of its conjugates during the induction of somatic embryogenesis in Coffea canephora. Plant Signal. Behav. 2013, 8, e26998. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Zheng, R.; Liu, G.; Xu, Y.; Zhou, Y.; Laux, T.; Zhen, Y.; Harding, S.A.; Shi, J.; Chen, J. Desiccation treatment and endogenous IAA levels are key factors influencing high frequency somatic embryogenesis in Cunninghamia lanceolata (Lamb.) hook. Front. Plant Sci. 2017, 8, 2054. [Google Scholar] [CrossRef] [Green Version]
- Ehmann, A. The van URK-Salkowski reagent-a sensitive and specific chromogenic reagent for silica gel thin-layer chromatographic detection and identification of indole derivatives. J. Chromatogr. A 1977, 132, 267–276. [Google Scholar] [CrossRef]
- Anthony, A.; Street, H. A colorimetric method for the estimation of certain indoles. New Phytol. 1969, 69, 47–50. [Google Scholar] [CrossRef]
- Guo, J.M.; Xin, Y.Y.; Yin, X.B. Selective differentiation of indoleacetic acid and indolebutyric acid using colorimetric recognition after ehrlich reaction. J. Agric. Food Chem. 2010, 58, 6556–6561. [Google Scholar] [CrossRef] [PubMed]
- Pujari, D.S.; Chanda, S.V. Effect of salinity stress on growth, peroxidase and IAA oxidase activities in Vigna seedlings. Acta Physiol. Plant. 2002, 24, 435–439. [Google Scholar] [CrossRef]
- Simon, S.; Petrášek, J. Why plants need more than one type of auxin. Plant Sci. 2011, 180, 454–460. [Google Scholar] [CrossRef] [Green Version]
- Paciorek, T.; Friml, J. Auxin signaling. J. Cell Sci. 2006, 119, 1199–1202. [Google Scholar] [CrossRef] [Green Version]
- Peer, W.A.; Cheng, Y.; Murphy, A.S. Evidence of oxidative attenuation of auxin signalling. J. Exp. Bot. 2013, 64, 2629–2639. [Google Scholar] [CrossRef]
- Michalczuk, L.; Cooke, T.J.; Cohen, J.D. Auxin levels at different stages of carrot somatic embryogenesis. Phytochemistry 1992, 31, 1097–1103. [Google Scholar] [CrossRef]
- Pasternak, T.P.; Prinsen, E.; Ayaydin, F.; Miskolczi, P.; Potters, G.; Asard, H.; Van Onckelen, H.A.; Dudits, D.; Fehér, A. The Role of auxin, pH, and stress in the activation of embryogenic cell division in leaf protoplast-derived cells of alfalfa. Plant Physiol. 2002, 129, 1807–1819. [Google Scholar] [CrossRef] [Green Version]
- Grossmann, K. Auxin Herbicide Action. Plant Signal. Behav. 2007, 2, 421–423. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Zhang, X. Regulation of Somatic Embryogenesis in Higher Plants. CRC Crit. Rev. Plant Sci. 2010, 29, 36–57. [Google Scholar] [CrossRef]
- Correia, S.; Vinhas, R.; Manadas, B.; Lourenço, A.S.; Veríssimo, P.; Canhoto, J.M. Comparative Proteomic Analysis of Auxin-Induced Embryogenic and Nonembryogenic Tissues of the Solanaceous Tree Cyphomandra betacea (Tamarillo). J. Proteome Res. 2012, 11, 1666–1675. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.H.; Liu, Y.B.; Bai, B.; Zhang, X.S. Establishment of embryonic shoot-root axis is involved in auxin and cytokinin response during Arabidopsis somatic embryogenesis. Front. Plant Sci. 2015, 5, 792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Sanz, H.; Manzanera, J.A.; Solís, M.T.; Gómez-Garay, A.; Pintos, B.; Risueño, M.C.; Testillano, P.S. Early markers are present in both embryogenesis pathways from microspores and immature zygotic embryos in cork oak, Quercus suber L. BMC Plant Biol. 2014, 14, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, B.; Su, Y.H.; Yuan, J.; Zhang, X.S. Induction of Somatic Embryos in Arabidopsis Requires Local YUCCA Expression Mediated by the Down-Regulation of Ethylene Biosynthesis. Mol. Plant 2013, 6, 1247–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, I.R.; Maximiano, M.R.; Almeida, R.F.; da Cunha, R.N.V.; Lopes, R.; Scherwinski-Pereira, J.E.; Mehta, A. Genotype-dependent changes of gene expression during somatic embryogenesis in oil palm hybrids (Elaeis oleifera x E. guineensis). PLoS ONE 2019, 13, e0209445. [Google Scholar] [CrossRef]
- Luo, J.; Zhou, J.J.; Zhang, J.Z. Aux/IAA gene family in plants: Molecular structure, regulation, and function. Int. J. Mol. Sci. 2018, 19, 259. [Google Scholar] [CrossRef] [Green Version]
- Elhiti, M.; Stasolla, C. Transduction of Signals during Somatic Embryogenesis. Plants 2022, 11, 178. [Google Scholar] [CrossRef]
- Kulaeva, O.N.; Prokoptseva, O.S. Recent Advances in the Study of Mechanisms of Action of Phytohormones. Biochemical 2004, 69, 233–247. [Google Scholar] [CrossRef]
- Gulzar, B.; Mujib, A.; Malik, M.Q.; Sayeed, R.; Mamgain, J.; Ejaz, B. Genes, proteins and other networks regulating somatic embryogenesis in plants. J. Genet. Eng. Biotechnol. 2020, 18, 31. [Google Scholar] [CrossRef]
- Sun, R.; Wang, S.; Ma, D.; Li, Y.; Liu, C. Genome-Wide Analysis of Cotton Auxin Early Somatic Embryogenesis. Genes 2019, 10, 730. [Google Scholar] [CrossRef] [Green Version]
- Dharmasiri, N.; Dharmasiri, S.; Estelle, M. The F-box protein TIR1 is an auxin receptor. Nature 2005, 435, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 474–497. [Google Scholar] [CrossRef]
- Kim, Y.; Oh, Y.J.; Park, W.J. HPLC-based quantification of indole-3-acetic acid in the primary root tip of maize. J. Nanobiotechnol. 2006, 3, 40–45. [Google Scholar]
- Nakurte, I.; Keisa, A.; Rostoks, N. Development and validation of a reversed-phase liquid chromatography method for the simultaneous determination of indole-3-acetic acid, indole-3-pyruvic acid, and abscisic acid in Barley (Hordeum vulgare L.). J. Anal. Methods Chem. 2012, 2012, 103575. [Google Scholar] [CrossRef] [Green Version]
- Cordeiro, D.; Rito, M.; Borges, F.; Canhoto, J.; Correia, S. Selection and validation of reference genes for qPCR analysis of miRNAs and their targets during somatic embryogenesis in tamarillo (Solanum betaceum Cav.). Plant Cell Tissue Organ Cult. 2020, 143, 109–120. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TIBA (µM) | Mass Increment (g) | Embryo Formation (Number/g of Tissues) | |
|---|---|---|---|
| Normal | Abnormal | ||
| 0 | 1.057 ± 0.140 a,b | 22.16 ± 9.58 a | 9.08 ± 8.02 b |
| 0.5 | 1.140 ± 0.201 a | 17.42 ± 1.99 a | 3.86 ± 1.18 b,c |
| 1 | 1.132 ± 0.102 a | 14.4 ± 3.45 a | 1.71 ± 0.20 c |
| 5 | 1.045 ± 0.217 b | 7.77 ± 2.90 b | 10.13 ± 2.62 a,b |
| 10 | 0.695 ± 0.190 b | 2.08 ± 1.50 b | 1.51 ± 0.744 c |
| Tissue | Protein Concentration (mg/mL) ± SD |
|---|---|
| NEC1 | 0.112 ± 0.029 c |
| NEC2 | 0.290 ± 0.067 c |
| EC1 | 0.693 ± 0.075 a |
| EC2 | 0.625 ± 0.071 a |
| EC3 | 0.388 ± 0.290 a,b |
| Gene | Forward Primer | Reverse Primer | Amplicon Length (bp) |
|---|---|---|---|
| Ef1α | ACAAGCGTGTCATCGAGAGG | TGTGTCCAGGGGCATCAATC | 183 |
| FeSOD | TCACCATCGACGTTTGGG AG | GACTGCTTCCCATGACACCA | 114 |
| IAA11 | AGGAAGGGTGCCTAGTTAGC | TGACACCCCTCGAGTAAGGA | 631 |
| IAA14 | AGTTTTCCGACGAAGAGGGT | GTTGGCCACCAGTGAGATCAT | 332 |
| IAA17 | TTGATGAAGAGCTCGGAGGC | CCCCGTGGCCTTATTTACGA | 335 |
| TIR1 | AGATGGCTGTCCAAAGCTCC | GAGCCTTGTCTCCAAACGGA | 389 |
| AFB3 | CTGTACGGAAATGGGGTGCT | GCAGAGTACGGGGAACCAAA | 284 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caeiro, A.; Caeiro, S.; Correia, S.; Canhoto, J. Induction of Somatic Embryogenesis in Tamarillo (Solanum betaceum Cav.) Involves Increases in the Endogenous Auxin Indole-3-Acetic Acid. Plants 2022, 11, 1347. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101347
Caeiro A, Caeiro S, Correia S, Canhoto J. Induction of Somatic Embryogenesis in Tamarillo (Solanum betaceum Cav.) Involves Increases in the Endogenous Auxin Indole-3-Acetic Acid. Plants. 2022; 11(10):1347. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101347
Chicago/Turabian StyleCaeiro, André, Sandra Caeiro, Sandra Correia, and Jorge Canhoto. 2022. "Induction of Somatic Embryogenesis in Tamarillo (Solanum betaceum Cav.) Involves Increases in the Endogenous Auxin Indole-3-Acetic Acid" Plants 11, no. 10: 1347. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101347