
Putrescine Treatment Delayed the Softening of Postharvest Blueberry Fruit by Inhibiting the Expression of Cell Wall Metabolism Key Gene VcPG1


Abstract
:1. Introduction
2. Results
2.1. Effect of Put Treatment on Decay Incidence and Firmness of Blueberry Fruit during Softening
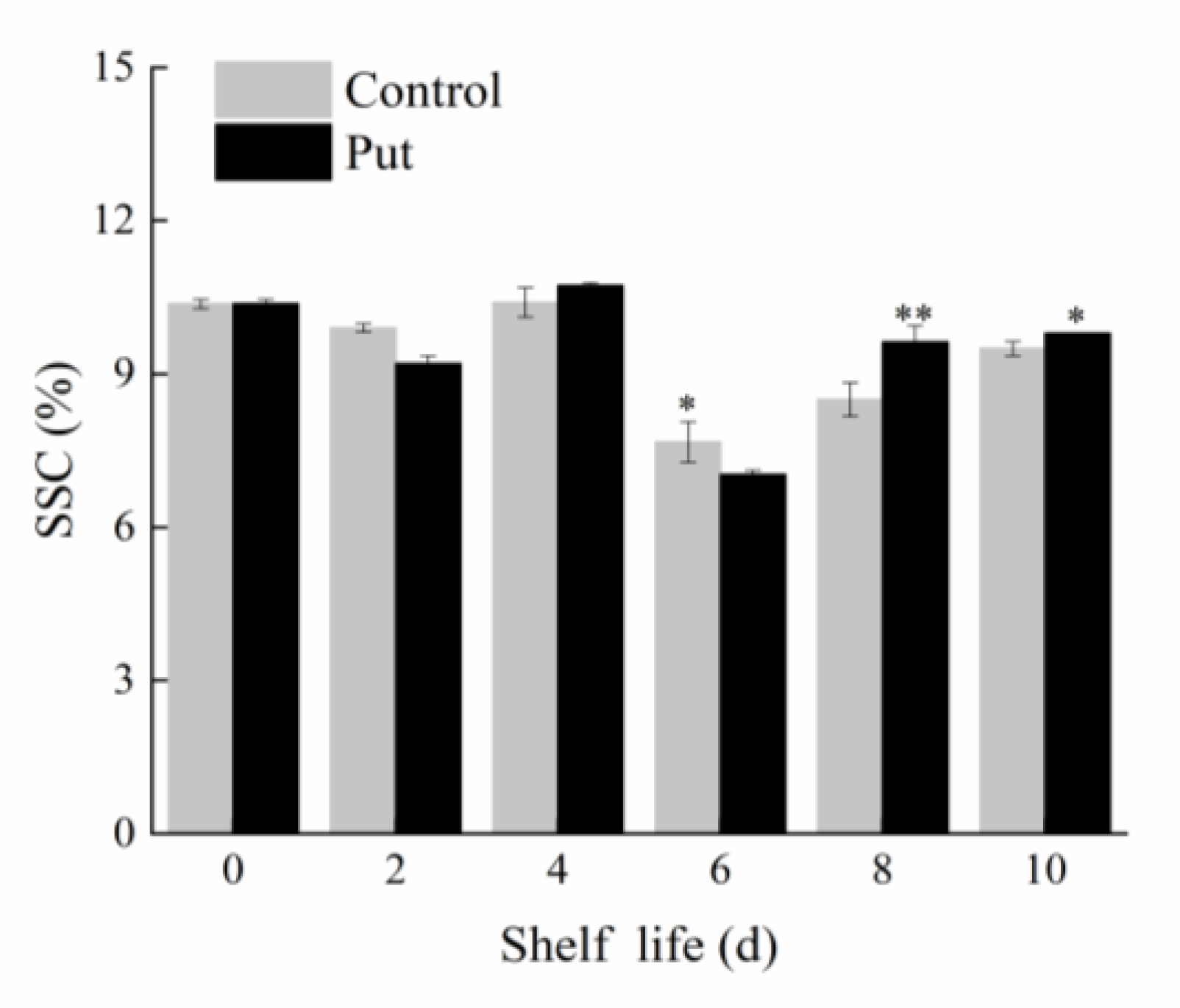
2.2. Effect of Put Treatment on Soluble Solid Content of Blueberry Fruit during Softening
2.3. Effect of Put Treatment on PP and WSP Content of Blueberry Fruit during Softening
2.4. Effects of Put Treatment on PG, β−Gal, and β−Glu Activities and Related Gene Expression Levels in Blueberry Fruit during Softening
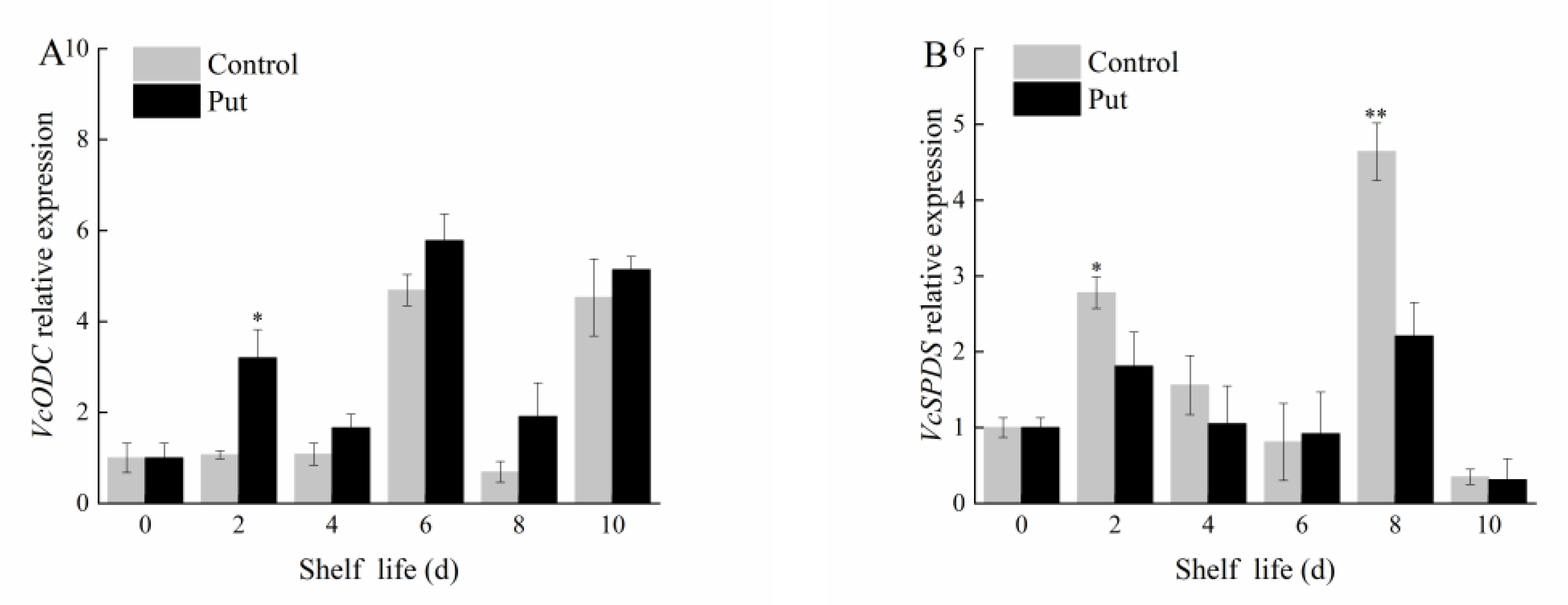
2.5. Effect of Put Treatment on the Expression of Put Anabolism−Related Genes in Blueberry Fruit during Softening
2.6. Cloning and Analysis of Coding Region VcPG1 Gene
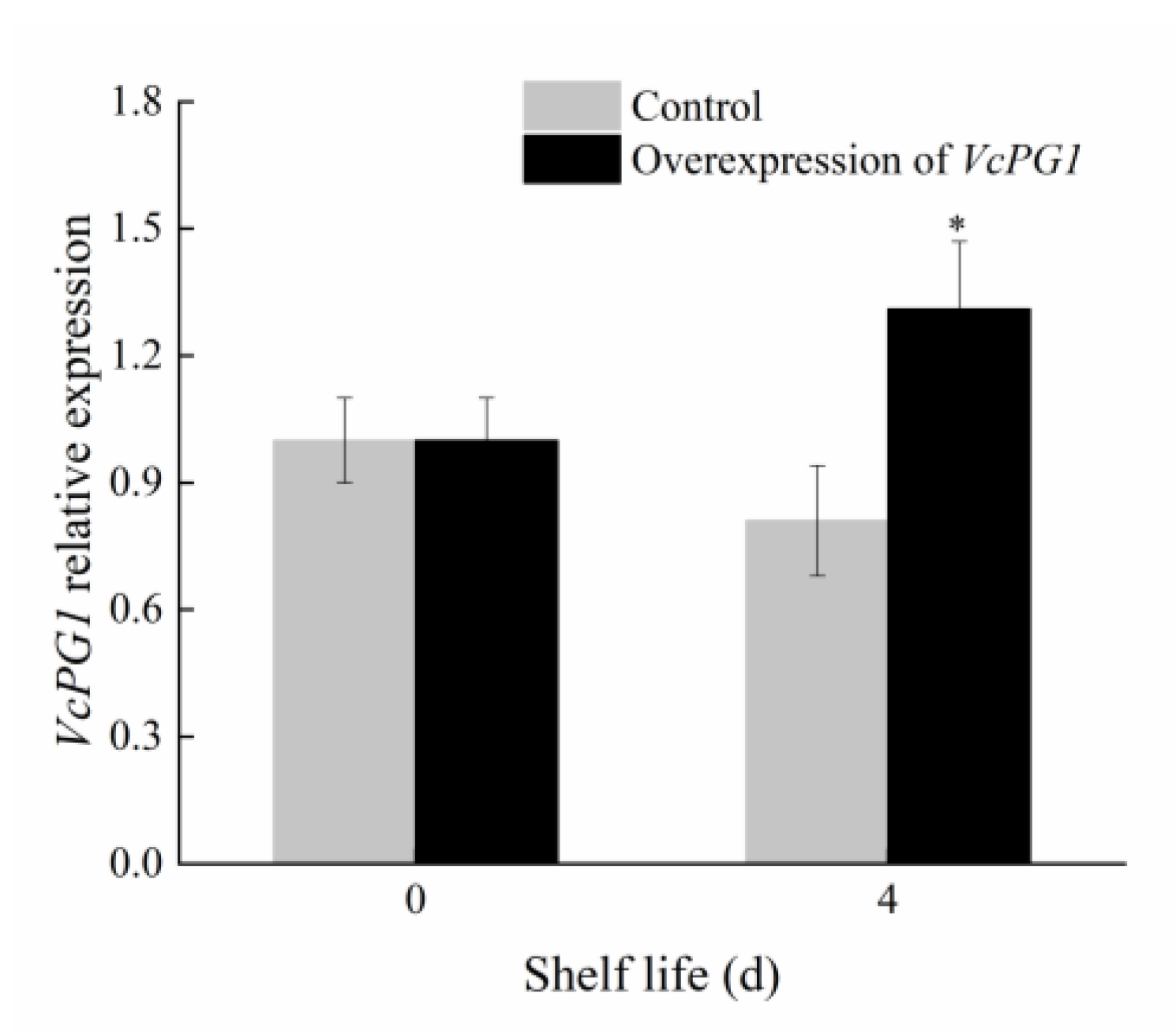
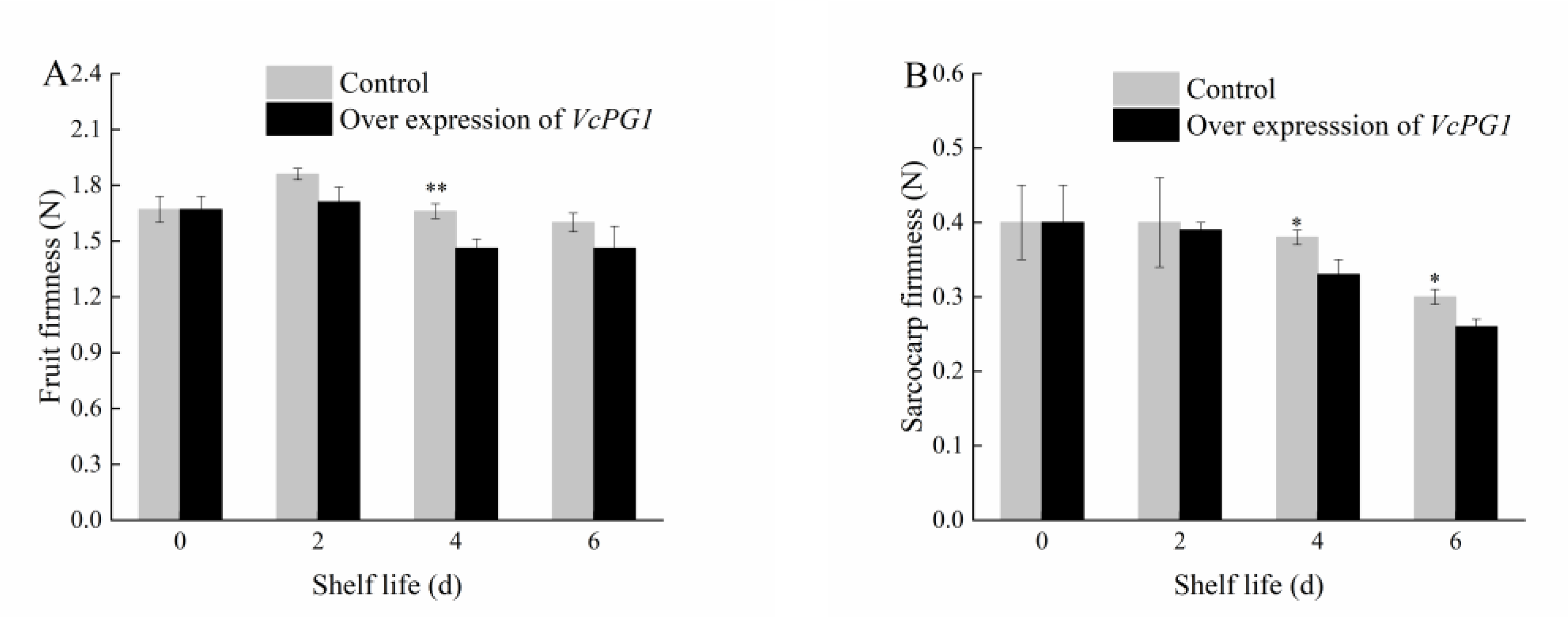
2.7. Changes in Morphology and Firmness of Blueberry Fruit Overexpressing VcPG1
2.8. Prediction of Related Transcription Factor Loci
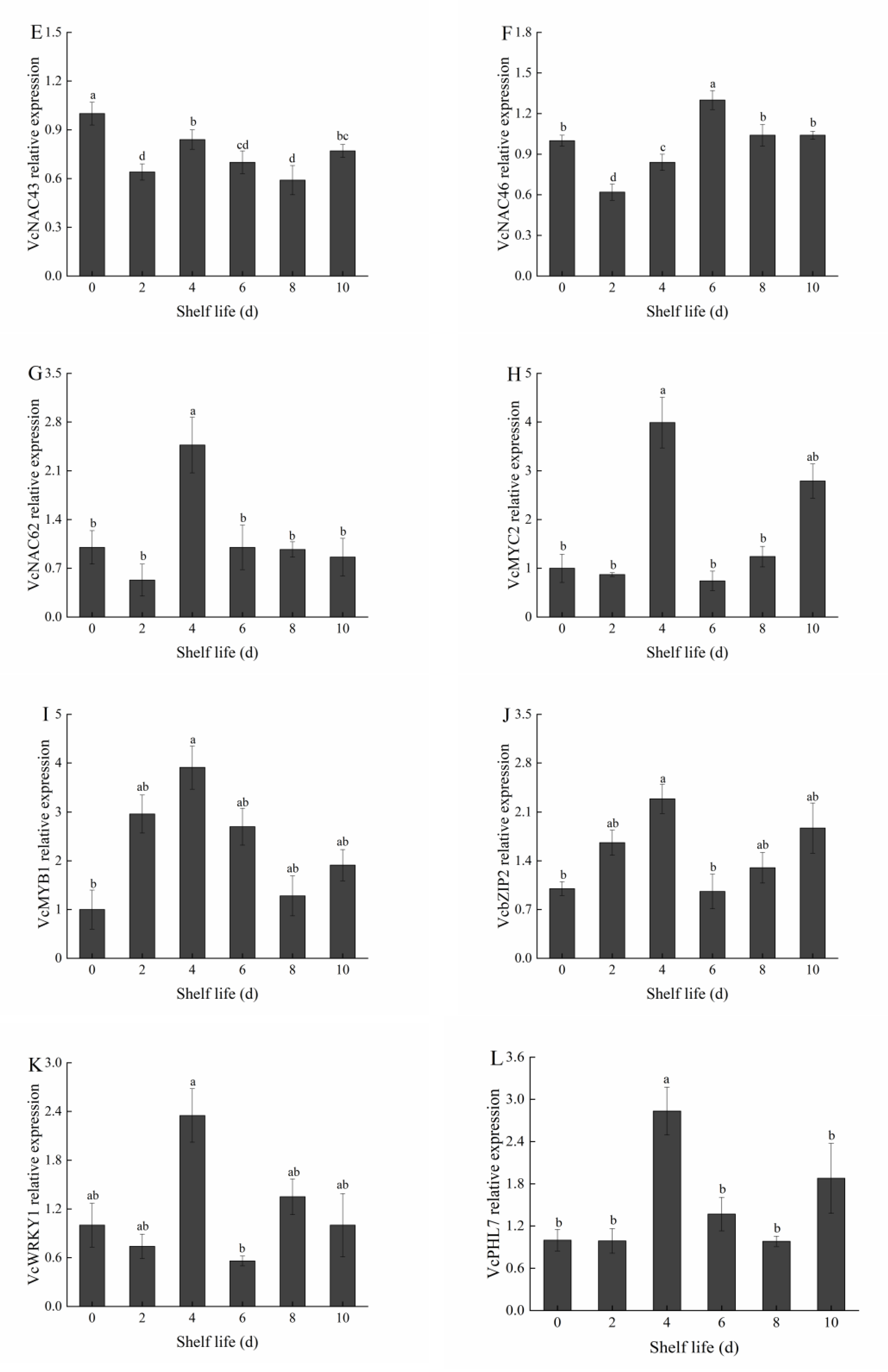
2.9. Analysis of the Expression of Predictive Transcription Factors
3. Discussion
4. Material and Methods
4.1. Fruit Materials and Treatment
4.2. Decay Incidence
4.3. Firmness
4.4. SSC
4.5. Pectin Content
4.6. PG, β−Gal, and β−Glu Activity
4.7. RNA Isolation and cDNA Synthesis
4.8. Gene Expression Analysis
4.9. Cloning of the Full−Length Sequence of VcPG1 CDS
4.10. Overexpression of VcPG1 in Blueberries
4.11. Predictive Analysis of Cis−Acting Elements of Key Transcription Factors
4.12. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Domazetovic, V.; Marcucci, G.; Falsetti, I.; Bilia, A.R.; Vincenzini, M.T.; Brandi, M.L.; Iantomasi, T. Blueberry Juice Antioxidants Protect Osteogenic Activity against Oxidative Stress and Improve Long-Term Activation of the Mineralization Process in Human Osteoblast-Like SaOS-2 Cells: Involvement of SIRT1. Antioxidants 2020, 9, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.R.; Li, J.; Zhu, L.; Chang, P.; Li, L.L.; Zhang, L.Y. Identification and Characterization of MYB-bHLH-WD40 Regulatory Complex Members Controlling Anthocyanidin Biosynthesis in Blueberry Fruits Development. Genes 2019, 10, 496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krikorian, R.; Kalt, W.; McDonald, J.E.; Shidler, M.D.; Summer, S.S.; Stein, A.L. Cognitive performance in relation to urinary anthocyanins and their flavonoid-based products following blueberry supplementation in older adults at risk for dementia. J. Funct. Foods 2020, 64, 103667. [Google Scholar] [CrossRef]
- Malin, D.H.; Lee, D.R.; Goyarzu, P.; Chang, Y.; Ennis, L.; Becontrolett, E.; Shukitt, B.; Joseph, J. Short-term blueberry-enriched diet prevents and reverses object recognition memory loss in aging rats. Nutrition 2011, 27, 338–342. [Google Scholar] [CrossRef]
- Ge, Y.H.; Zhang, J.H.; Li, C.Y.; Xue, W.J.; Zhang, S.Y.; Lv, J.Y. Trisodium phosphate delays softening of jujube fruit by inhibiting cell wall-degrading enzyme activities during ambient storage. Sci. Hortic. 2020, 262, 109059. [Google Scholar] [CrossRef]
- Wang, H.L.; Guo, X.B.; Hu, X.D.; Li, T.; Fu, X.; Liu, R. Comparison of phytochemical profiles, antioxidant and cellular antioxidant activities of different varieties of blueberry (Vaccinium spp.). Food Chem. 2017, 217, 773–781. [Google Scholar] [CrossRef]
- Wang, D.D.; Yeats, T.H.; Uluisik, S.; Rose, J.K.C.; Seymour, G.B. Fruit Softening: Revisiting the Role of Pectin. Trends Plant Sci. 2018, 23, 302–310. [Google Scholar] [CrossRef]
- Li, C.Y.; Zhang, J.H.; Ge, Y.H.; Li, X.Y.; Wei, M.L.; Hou, J.B.; Cheng, Y.; Lv, J.Y. Postharvest acibenzolar-S-methyl treatment maintains storage quality and retards softening of apple fruit. J. Food Biochem. 2020, 44, e13141. [Google Scholar] [CrossRef]
- Goulao, L.F.; Oliveira, C.M. Cell wall modifications during fruit ripening: When a fruit is not the fruit. Trends Food Sci. Technol. 2007, 19, 4–25. [Google Scholar] [CrossRef] [Green Version]
- Dhall, R.K. Advances in Edible Coatings for Fresh Fruits and Vegetables: A Review. Crit. Rev. Food Sci. Nutr. 2013, 53, 435–450. [Google Scholar] [CrossRef]
- Rose, J.K.C.; Bennett, A.B. Cooperative disassembly of the cellulose–xyloglucan network of plant cell walls: Parallels between cell expansion and fruit ripening. Trends Plant Sci. 1999, 4, 176–183. [Google Scholar] [CrossRef]
- Guo, S.L.; Song, J.; Zhang, B.B.; Jiang, H.; Ma, R.J.; Yu, M.L. Genome-wide identification and expression analysis of beta-galactosidase family members during fruit softening of peach. Postharvest Biol. Technol. 2018, 136, 111–123. [Google Scholar] [CrossRef]
- Wei, J.M.; Ma, F.W.; Shi, S.G.; Qi, X.D.; Zhu, X.Q.; Yuan, J.W. Changes and postharvest regulation of activity and gene expression of enzymes related to cell wall degradation in ripening apple fruit. Postharvest Biol. Technol. 2010, 56, 147–154. [Google Scholar] [CrossRef]
- Win, N.M.; Yoo, J.; Kwon, S.; Watkins, C.B.; Kang, I. Characterization of Fruit Quality Attributes and Cell Wall Metabolism in 1-Methylcyclopropene (1-MCP)-Treated ‘Summer King’ and ‘Green Ball’ Apples during Cold Storage. Front. Plant Sci. 2019, 10, 1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.L.; Abbott, J.A.; Gross, K.C. Down-Regulation of Tomato β-Galactosidase 4 Results in Decreased Fruit Softening. Plant Physiol. 2002, 129, 1755–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Xu, C.J.; Korban, S.S.; Chen, K.S. Regulatory mechanisms of textural changes in ripening fruits. Crit. Rev. Plant Sci. 2010, 29, 222–243. [Google Scholar] [CrossRef]
- Yi, J.W.; Wang, Y.; Ma, X.S.; Zhang, J.Q.; Zhao, M.L.; Huang, X.M.; Li, J.G.; Hu, G.B.; Wang, H.C. LcERF2 modulates cell wall metabolism by directly targeting a UDP-glucose-4-epimerase gene to regulate pedicel development and fruit abscission of litchi. Plant J. 2021, 106, 801–816. [Google Scholar] [CrossRef]
- Zhai, Z.F.; Feng, C.; Wang, Y.Y.; Sun, Y.T.; Peng, X.; Xiao, Y.Q.; Zhang, X.; Zhou, X.; Jiao, J.L.; Wang, W.L.; et al. Genome-Wide Identification of the Xyloglucan endotransglucosylase/Hydrolase (XTH) and Polygalacturonase (PG) Genes and Characterization of Their Role in Fruit Softening of Sweet Cherry. Int. J. Mol. Sci. 2021, 22, 12331. [Google Scholar] [CrossRef]
- Barman, K.; Asrey, R.; Pal, R.K. Putrescine and carnauba wax pretreatments alleviate chilling injury, enhance shelf life and preserve pomegranate fruit quality during cold storage. Sci. Hortic. 2011, 130, 795–800. [Google Scholar] [CrossRef]
- Qian, C.L.; Ji, Z.J.; Zhu, Q.; Qi, X.H.; Li, Q.Q.; Yin, J.D.; Liu, J.; Kan, J.; Zhang, M.; Jin, C.H.; et al. Effects of 1-MCP on proline, polyamine, and nitric oxide metabolism in postharvest peach fruit under chilling stress. Hortic. Plant J. 2021, 7, 188–196. [Google Scholar] [CrossRef]
- Mirdehghan, S.H.; Rahimi, S. Pre-harvest application of polyamines enhances antioxidants and table grape (Vitis vinifera L.) quality during postharvest period. Food Chem. 2016, 196, 1040–1047. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, S.M.; Hosseini, M.S.; Karimi, M.; Ebrahimzadeh, A. Effects of postharvest polyamine application and edible coating on maintaining quality of mango ( Mangifera indica L.) cv. Langra during cold storage. Food Sci. Nutr. 2019, 7, 433–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palma, F.; Carvajal, F.; Jamilena, M.; Garrido, D. Contribution of polyamines and other related metabolites to the maintenance of zucchini fruit quality during cold storage. Plant Physiol. Biochem. 2014, 82, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Li, J.X.; Luo, M.L.; Zhou, X.; Zhou, Q.; Wei, B.D.; Cheng, S.C.; Ji, S.J. Polyamine treatment ameliorates pericarp browning in cold-stored ‘Nanguo’ pears by protecting mitochondrial structure and function. Postharvest Biol. Technol. 2021, 178. [Google Scholar] [CrossRef]
- Ghosh, A.; Saha, I.; Debnath, S.C.; Hasanuzzaman, M.; Adak, M.K. Chitosan and putrescine modulate reactive oxygen species metabolism and physiological responses during chili fruit ripening. Plant Physiol. Biochem. 2021, 163, 55–67. [Google Scholar] [CrossRef]
- Singh, V.; Jawandha, S.K.; Gill, P.P.S. Putrescine application reduces softening and maintains the quality of pear fruit during cold storage. Acta Physiol. Plant. 2020, 42, 932–936. [Google Scholar] [CrossRef]
- Razzaq, K.; Khan, A.S.; Malik, A.U.; Shahid, M.; Ullah, S. Role of putrescine in regulating fruit softening and antioxidative enzyme systems in ‘Samar Bahisht Chaunsa’ mango. Postharvest Biol. Technol. 2014, 96, 23–32. [Google Scholar] [CrossRef]
- Chea, S.; Yu, D.J.; Park, J.; Oh, H.D.; Chung, S.W.; Lee, H.J. Fruit softening correlates with enzymatic and compositional changes in fruit cell wall during ripening in ‘Bluecrop’ highbush blueberries. Sci. Hortic. 2019, 245, 163–170. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhang, F.; Ji, S.J.; Dai, H.Y.; Zhou, X.; Wei, B.D.; Cheng, S.C.; Wang, A. Abscisic acid accelerates postharvest blueberry fruit softening by promoting cell wall metabolism. Sci. Hortic. 2021, 288, 110325. [Google Scholar] [CrossRef]
- Chen, J.; Li, Y.X.; Li, F.F.; Hong, K.Q.; Yuan, D.B. Effects of procyanidin treatment on the ripening and softening of banana fruit during storage. Sci. Hortic. 2021, 292, 110644. [Google Scholar] [CrossRef]
- Chen, Y.P.; Zhang, X.C.; Luo, Z.S.; Sun, J.; Li, L.; Yin, X.R.; Li, J.K.; Xu, Y.Q. Effects of inside-out heat-shock via microwave on the fruit softening and quality of persimmon during postharvest storage. Food Chem. 2021, 349, 129161. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Li, T.T.; Chen, S.Q.; Li, Y.L.; Rashid, A. The biochemical and molecular mechanisms of softening inhibition by chitosan coating in strawberry fruit (Fragaria × ananassa) during cold storage. Sci. Hortic. 2020, 271, 109483. [Google Scholar] [CrossRef]
- Wang, S.Y.; Zhou, Q.; Zhou, X.; Wei, B.D.; Ji, S.J. The effect of ethylene absorbent treatment on the softening of blueberry fruit. Food Chem. 2018, 246, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.B.; Cheng, X.; Wu, C.; Fan, G.J.; Li, T.T.; Dong, C. Retardation of postharvest softening of blueberry fruit by methyl jasmonate is correlated with altered cell wall modification and energy metabolism. Sci. Hortic. 2021, 276, 109752. [Google Scholar] [CrossRef]
- Petkou, T.; Pritsa, T.S.; Sfakiotakis, E.M. Effects of polyamines on ethylene production, respiration and ripening of kiwifruit. J. Hortic. Sci. Biotechnol. 2004, 79, 977–980. [Google Scholar] [CrossRef]
- Meng, L.; Zhi, H.H.; Dong, Y. The influence of pre- and postharvest 1-MCP application and oxygen regimes on textural properties, cell wall metabolism, and physiological disorders of late-harvest ‘Bartlett’ pears. Postharvest Biol. Technol. 2021, 173, 111429. [Google Scholar] [CrossRef]
- Pan, H.X.; Wang, L.M.; Wang, R.; Xie, F.; Cao, J.K. Modifications of cell wall pectin in chilling-injured ‘Friar’ plum fruit subjected to intermediate storage temperatures. Food Chem. 2018, 242, 538–547. [Google Scholar] [CrossRef]
- Brummell, D.A. Cell wall disassembly in ripening fruit. Funct. Plant Biol. 2006, 33, 103–119. [Google Scholar] [CrossRef]
- Chen, H.J.; Cao, S.F.; Fang, X.J.; Mu, H.L.; Yang, H.L.; Wang, X.; Xu, Q.Q.; Gao, H.Y. Changes in fruit firmness, cell wall composition and cell wall degrading enzymes in postharvest blueberries during storage. Sci. Hortic. 2015, 188, 44–48. [Google Scholar] [CrossRef]
- Zhao, Y.T.; Zhu, X.; Hou, Y.Y.; Wang, X.Y.; Li, X.H. Effects of nitric oxide fumigation treatment on retarding cell wall degradation and delaying softening of winter jujube (Ziziphus jujube Mill. cv. Dongzao) fruit during storage. Postharvest Biol. Technol. 2019, 156, 110954. [Google Scholar] [CrossRef]
- Brummell, D.A.; Harpster, M.H. Cell wall metabolism in fruit softening and quality and its manipulation in transgenic plants. Plant Mol. Biol. 2001, 47, 311–340. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chen, Y.H.; Lin, H.T.; Lin, M.S.; Chen, Y.H.; Lin, Y.F. 1-Methylcyclopropene containing-papers suppress the disassembly of cell wall polysaccharides in Anxi persimmon fruit during storage. Int. J. Biol. Macromol. 2020, 151, 723–729. [Google Scholar] [CrossRef]
- Bu, J.W.; Yu, Y.C.; Aisikaer, G.; Ying, T.J. Postharvest UV-C irradiation inhibits the production of ethylene and the activity of cell wall-degrading enzymes during softening of tomato (Lycopersicon esculentum L.) fruit. Postharvest Biol. Technol. 2013, 86, 337–345. [Google Scholar] [CrossRef]
- Ahmad, S.K.; Zora, S.; Nadeem, A.A. Pre-storage putrescine application suppresses ethylene biosynthesis and retards fruit softening during low temperature storage in ‘Angelino’ plum. Postharvest Biol. Technol. 2007, 46, 36–46. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.Y.; Zhou, Q.; Zhou, X.; Zhang, F.; Ji, S.J. Ethylene plays an important role in the softening and sucrose metabolism of blueberries postharvest. Food Chem. 2019, 310, 125965. [Google Scholar] [CrossRef]
- Ahmad, A.; Ali, A. Improvement of postharvest quality, regulation of antioxidants capacity and softening enzymes activity of cold-stored carambola in response to polyamines application. Postharvest Biol. Technol. 2018, 148, 208–217. [Google Scholar] [CrossRef]
- Duan, B.; Ge, Y.H.; Li, C.Y.; Gao, X.N.; Tang, Q.; Li, X.; Wei, M.L.; Chen, Y.R. Effect of exogenous ATP treatment on sucrose metabolism and quality of Nanguo pear fruit. Sci. Hortic. 2019, 249, 71–76. [Google Scholar] [CrossRef]
- Noussaire, E.F.; Khalil, E.M.; Mia, E.; Henk, A.S.; Filali-Zegzouti, Y.; Hassan, H. Physicochemical and functional characterization of pectin extracted from Moroccan citrus peels. LWT 2022, 162, 113508. [Google Scholar] [CrossRef]
- Ren, Y.Y.; Sun, P.P.; Wang, X.X.; Zhu, Z.Y. Degradation of cell wall polysaccharides and change of related enzyme activities with fruit softening in Annona squamosa during storage. Postharvest Biol. Technol. 2020, 166, 111203. [Google Scholar] [CrossRef]
- Zhang, C.H.; Xiong, Z.H.; Yang, H.Y.; Wu, W.L. Changes in pericarp morphology, physiology and cell wall composition account for flesh firmness during the ripening of blackberry ( Rubus spp.) fruit. Sci. Hortic. 2019, 250, 59–68. [Google Scholar] [CrossRef]
- Ji, Y.R.; Hu, W.Z.; Liao, J.; Xiu, Z.L.; Jiang, A.L.; Guan, Y.G.; Yang, X.Z.; Feng, K. Ethanol vapor delays softening of postharvest blueberry by retarding cell wall degradation during cold storage and shelf life. Postharvest Biol. Technol. 2021, 177, 111538. [Google Scholar] [CrossRef]
- Laura, M.P.; Francisco, J.M.H.; Maaike, B.; Robert, C.S.; Irene, L.V.; Roberto, S.; Jose-Manuel, F.Z.; Jose, L.C.; Rosario, B.P.; Juan, M.B. An R2R3-MYB transcription factor (FaEOBII) regulates eugenol production in ripe strawberry (Fragaria × ananassa) fruit receptacles. Plant Physiol. 2015, 168, 598–614. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.L.; Zhou, X.; Hao, Y.; Sun, H.J.; Zhou, Q.; Sun, Y.Y.; Ji, S.J. Methyl jasmonate pretreatment improves aroma quality of cold-stored ‘Nanguo’ pears by promoting ester biosynthesis. Food Chem. 2021, 338, 127846. [Google Scholar] [CrossRef] [PubMed]
- Colle, M.; Leisner, C.P.; Wai, C.M.; Ou, S.; Bird, K.A.; Wang, J.; Wisecaver, J.H.; Yocca, A.E.; Alger, E.I.; Tang, H.B.; et al. Haplotype-phased genome and evolution of phytonutrient pathways of tetraploid blueberry. GigaScience 2019, 8, giz012. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.F.; Li, W.B.; Tian, N.F.; Wang, X.; Wu, W.Z.; Zheng, S. Cloning and functional identification of setaria italica somatic embryogenesis receptor-like kinase1 gene (SiSERK1). Gene 2021, 813, 146119. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward Primer | Reverse Primer | |
|---|---|---|
| Actin | 5′−ACTACCATCCACTCTATCACCG−3′ | 5′−AACACCTTACCAACAGCCTTG−3′ |
| VcPG1 | 5′−ACCACCAACCGCATTA−3′ | 5′−AAGCGTCAGGTGAGTAAG−3′ |
| VcPG2 | 5′−ACGGTTCAGGGTGTCTGGAT−3′ | 5′−GGTTGGGTGGTGTGTTTGCT−3′ |
| VcPG3 | 5′−CAGGGTCATGTGGCTGGTA−3′ | 5′−AGACGGGCGGACGCTTAA−3′ |
| Vcβ−Gal | 5′−CTTCTCTCTCTTCTCGCCGC−3′ | 5′−CGAATGCCTTTGCCCTCAAC−3′ |
| Vcβ−Glu | 5′−TCGACCGAAGCGTCGCTACT−3′ | 5′−GTCCACCATGTCCGCCCAAT−3′ |
| VcSPDS | 5′−TTCGTCGTCTCCACCTCATC−3′ | 5′−CCAGTTTCCTTGCCCAATTC−3′ |
| VcODC | 5′−GCGAACCCTAACAGCCACCA−3′ | 5′−TACCACGCCCAAGTCCAGCA−3′ |
| VcNAC26 | 5′−GATGGCATCCGGTGCTTCTCC−3′ | 5′−GAAGTGGACGACAAGCTCCTCATC−3′ |
| VcNAC23 | 5′−TTCATTGGTGGTTCCGCGAGAAG−3′ | 5′−GTCGATGGAAGGAACATCCTGCTC−3′ |
| VcNAC33 | 5′−GCGAACGAACAGATTGGGTAATGC−3′ | 5′−CCAGGTCCGCTCTTCTTGAACAC−3′ |
| VcNAC35 | 5′−CCAGTGTTCACAAGTGATGGCAAG−3′ | 5′−GGAGCCTGAACTCCTGCATTATCC−3′ |
| VcNAC43 | 5′−TCCAGAGCGAGCCGACGAAG−3′ | 5′−GCAACGGCGAGGTGAAGGTG−3′ |
| VcNAC46 | 5′−TCCAGAGCGAGCCGACGAAG−3′ | 5′−CGAGGTGAAGGTGCTGTCCATG−3′ |
| VcNAC62 | 5′−TGCTTCTCCAGCCACAGT−3′ | 5′−ACAACCGATTGACCTCCT−3′ |
| VcbZIP2 | 5′−ATGGCGATGGCAATGGGAAA−3′ | 5′−AGGGCTCGAAGCCGAGAGAA−3′ |
| VcMYC2 | 5′−GTAAAGGACCACCCCCACCT−3′ | 5′−CGTTCACGACCGACACACTC−3′ |
| VcWRKY1 | 5′−AGAAAAGAGGATGGGGAGGA−3′ | 5′−ATGATGATGATGGTGGTGGC−3′ |
| VcMYB1 | 5′−CTGCTCATCCTTACCCACA−3′ | 5′−TTCCACCGCCATCAATAC−3′ |
| VcPHL7 | 5′−AGGGATACTGCTCCAACT−3′ | 5′−GCAAGGCGATACTTCTGT−3′ |
| Forward Primer | Reverse Primer | |
|---|---|---|
| VcPG1 | 5′−ATCCTCCTCCCCATCCTCCT−3′ | 5′−TACAAATCATTACAAGTTTATCTAAGCACA−3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, X.; Dai, H.; Wang, S.; Ji, S.; Zhou, X.; Li, J.; Zhou, Q. Putrescine Treatment Delayed the Softening of Postharvest Blueberry Fruit by Inhibiting the Expression of Cell Wall Metabolism Key Gene VcPG1. Plants 2022, 11, 1356. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101356
Song X, Dai H, Wang S, Ji S, Zhou X, Li J, Zhou Q. Putrescine Treatment Delayed the Softening of Postharvest Blueberry Fruit by Inhibiting the Expression of Cell Wall Metabolism Key Gene VcPG1. Plants. 2022; 11(10):1356. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101356
Chicago/Turabian StyleSong, Xiangchong, Hongyu Dai, Siyao Wang, Shujuan Ji, Xin Zhou, Jianan Li, and Qian Zhou. 2022. "Putrescine Treatment Delayed the Softening of Postharvest Blueberry Fruit by Inhibiting the Expression of Cell Wall Metabolism Key Gene VcPG1" Plants 11, no. 10: 1356. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101356