
Application of CRISPR/Cas9 System for Efficient Gene Editing in Peanut

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
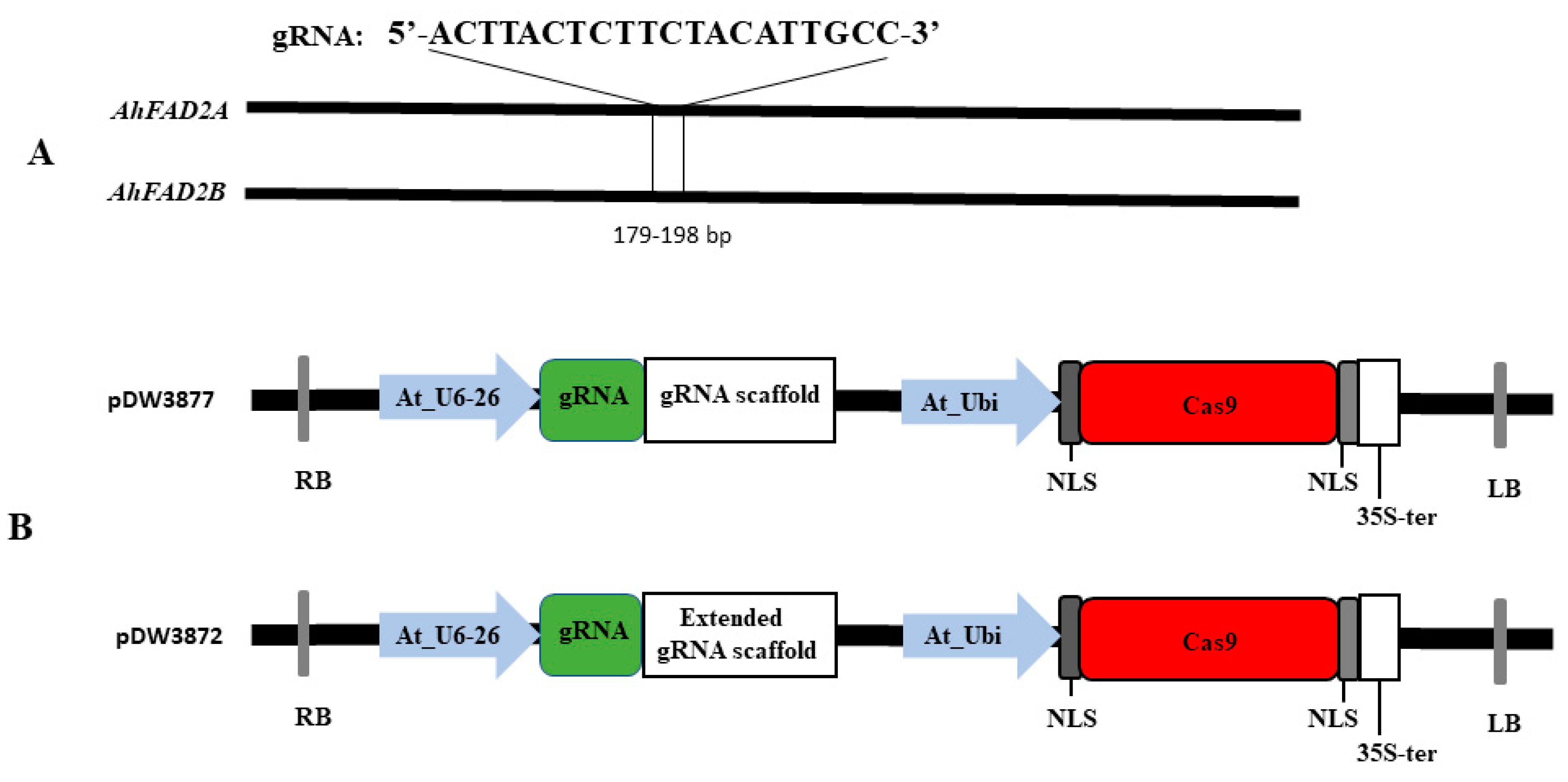
2.2. Vector Construction
2.3. Hairy Root Transformation
2.4. Validation of Mutagenesis
2.5. Evaluation of the Editing Efficiency
3. Results
3.1. Designing and Cloning of the gRNAs into the Constructs
3.2. Types of Edits
3.3. Editing Efficiency of Two Constructs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Chu, Y.; Chee, P.; Culbreath, A.; Isleib, T.G.; Holbrook, C.C.; Ozias-Akins, P. Major QTLs for Resistance to Early and Late Leaf Spot Diseases Are Identified on Chromosomes 3 and 5 in Peanut (Arachis hypogaea). Front. Plant Sci. 2019, 10, 883. [Google Scholar] [CrossRef] [PubMed]
- Andersen, P.C.; Gorbet, D.W. Influence of Year and Planting Date on Fatty Acid Chemistry of High Oleic Acid and Normal Peanut Genotypes. J. Agric. Food Chem. 2002, 50, 1298–1305. [Google Scholar] [CrossRef] [PubMed]
- Zahran, H.A.; Tawfeuk, H.Z. Physicochemical properties of new peanut (Arachis hypogaea L.) varieties. OCL 2019, 26, 19. [Google Scholar] [CrossRef] [Green Version]
- Al-Muhsen, S.; Clarke, A.E.; Kagan, R.S. Peanut allergy: An overview. CMAJ Can. Med. Assoc. J. 2003, 168, 1279–1285. [Google Scholar]
- Pandey, P.K.; Quilichini, T.; Vaid, N.; Gao, P.; Xiang, D.; Datla, R. Versatile and multifaceted CRISPR/Cas gene editing tool for plant research. Semin. Cell Dev. Biol. 2019, 96, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, M.; Trivedi, P.K.; Pandey, A. Emerging tools and paradigm shift of gene editing in cereals, fruits, and horticultural crops for enhancing nutritional value and food security. Food Energy Secur. 2021, 10, e258. [Google Scholar] [CrossRef]
- Wang, F.; Wang, C.; Liu, P.; Lei, C.; Hao, W.; Gao, Y.; Liu, Y.G.; Zhao, K. Enhanced rice blast resistance by CRISPR/Cas9-targeted muta-genesis of the ERF transcription factor gene OsERF922. PLoS ONE 2016, 11, e0154027. [Google Scholar]
- Nekrasov, V.; Wang, C.; Win, J.; Lanz, C.; Weigel, D.; Kamoun, S. Rapid generation of a transgene-free powdery mildew resistant tomato by genome deletion. Sci. Rep. 2017, 7, 482. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Bai, Y.; Wu, G.; Zou, S.; Chen, Y.; Gao, C.; Tang, D. Simultaneous modification of three homoeologs of TaEDR1 by genome editing enhances powdery mildew resistance in wheat. Plant J. 2017, 91, 714. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, D.; Zhang, D.; Zhao, X.; Cao, X.; Dong, L.; Liu, J.; Chen, K.; Zhang, H.; Gao, C.; et al. Analysis of the functions of TaGW2 homoeologs in wheat grain weight and protein content traits. Plant J. 2018, 94, 857–866. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.; Turesson, H.; Nicolia, A.; Fält, A.-S.; Samuelsson, M.; Hofvander, P. Efficient targeted multiallelic mutagenesis in tetraploid potato (Solanum tuberosum) by transient CRISPR-Cas9 expression in protoplasts. Plant Cell Rep. 2016, 36, 117–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, M.; Zhu, J.; Gong, L.; He, L.; Lee, C.; Han, S.; Chen, C.; He, G. Mutagenesis of FAD2 genes in peanut with CRISPR/Cas9 based gene editing. BMC Biotechnol. 2019, 19, 24. [Google Scholar] [CrossRef] [PubMed]
- Nadakuduti, S.S.; Enciso-Rodríguez, F. Advances in Genome Editing With CRISPR Systems and Transformation Technologies for Plant DNA Manipulation. Front. Plant Sci. 2021, 11, 2267. [Google Scholar] [CrossRef] [PubMed]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimatani, Z.; Kashojiya, S.; Takayama, M.; Terada, R.; Arazoe, T.; Ishii, H.; Teramura, H.; Yamamoto, T.; Komatsu, H.; Miura, K.; et al. Targeted base editing in rice and tomato using a CRISPR-Cas9 cytidine deaminase fusion. Nat. Biotechnol. 2017, 35, 441–443. [Google Scholar] [CrossRef]
- Zong, Y.; Wang, Y.; Li, C.; Zhang, R.; Chen, K.; Ran, Y.; Qiu, J.-L.; Wang, D.; Gao, C. Precise base editing in rice, wheat and maize with a Cas9-cytidine deaminase fusion. Nat. Biotechnol. 2017, 35, 438–440. [Google Scholar] [CrossRef]
- Shan, Q.; Voytas, D.F. Editing plant genes one base at a time. Nat. Plants 2018, 4, 412–413. [Google Scholar] [CrossRef]
- Li, C.; Zhang, R.; Meng, X.; Chen, S.; Zong, Y.; Lu, C.; Qiu, J.-L.; Chen, Y.-H.; Li, J.; Gao, C. Targeted, random mutagenesis of plant genes with dual cytosine and adenine base editors. Nat. Biotechnol. 2020, 38, 875–882. [Google Scholar] [CrossRef]
- Lowder, L.G.; Zhou, J.; Zhang, Y.; Malzahn, A.; Zhong, Z.; Hsieh, T.-F.; Voytas, D.; Zhang, Y.; Qi, Y. Robust Transcriptional Activation in Plants Using Multiplexed CRISPR-Act2.0 and mTALE-Act Systems. Mol. Plant 2018, 11, 245–256. [Google Scholar] [CrossRef] [Green Version]
- Moradpour, M.; Abdulah, S.N.A. CRISPR/dC as9 platforms in plants: Strategies and applications beyond genome editing. Plant Biotechnol. J. 2020, 18, 32–44. [Google Scholar] [CrossRef] [Green Version]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; van der Oost, J.; Regev, A.; et al. Cpf1 Is a Single RNA-Guided Endonuclease of a Class 2 CRISPR-Cas System. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Malzahn, A.A.; Sretenovic, S.; Qi, Y. The emerging and uncultivated potential of CRISPR technology in plant science. Nat. Plants 2019, 5, 778–794. [Google Scholar] [CrossRef] [PubMed]
- Kleinstiver, B.P.; Sousa, A.A.; Walton, R.; Tak, Y.E.; Hsu, J.Y.; Clement, K.; Welch, M.M.; Horng, J.E.; Malagon-Lopez, J.; Scarfò, I.; et al. Engineered CRISPR–Cas12a variants with increased activities and improved targeting ranges for gene, epigenetic and base editing. Nat. Biotechnol. 2019, 37, 276–282. [Google Scholar] [CrossRef]
- Sun, X.; Hu, Z.; Chen, R.; Jiang, Q.; Song, G.; Zhang, H.; Xi, Y. Targeted mutagenesis in soybean using the CRISPR-Cas9 system. Sci. Rep. 2015, 5, 10342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Z.; Zhang, Z.; Hua, K.; Gao, X.; Mao, Y.; Botella, J.R.; Zhu, J.-K. A Highly Efficient Cell Division-Specific CRISPR/Cas9 System Generates Homozygous Mutants for Multiple Genes in Arabidopsis. Int. J. Mol. Sci. 2018, 19, 3925. [Google Scholar] [CrossRef] [Green Version]
- Dang, Y.; Jia, G.; Choi, J.; Ma, H.; Anaya, E.; Ye, C.; Shankar, P.; Wu, H. Optimizing sgRNA structure to improve CRISPR-Cas9 knockout efficiency. Genome Biol. 2015, 16, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Cornejo, M.-J.; Luth, D.; Blankenship, K.M.; Anderson, O.D.; Blechl, A.E. Activity of a maize ubiquitin promoter in transgenic rice. Plant Mol. Biol. 1993, 23, 567–581. [Google Scholar] [CrossRef]
- Miao, J.; Guo, D.; Zhang, J.; Huang, Q.; Qin, G.; Zhang, X.; Wan, J.; Gu, H.; Qu, L.-J. Targeted mutagenesis in rice using CRISPR-Cas system. Cell Res. 2013, 23, 1233–1236. [Google Scholar] [CrossRef] [Green Version]
- Nandy, S.; Pathak, B.; Zhao, S.; Srivastava, V. Heat-shock-inducible CRISPR/Cas9 system generates heritable mutations in rice. Plant Direct 2019, 3, e00145. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, J.; Wei, P.; Zhang, B.; Gou, F.; Feng, Z.; Mao, Y.; Yang, L.; Zhang, H.; Xu, N.; et al. The CRISPR/C as9 system produces specific and homozygous targeted gene editing in rice in one generation. Plant Biotechnol. J. 2014, 12, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Mao, Y.; Xu, N.; Zhang, B.; Wei, P.; Yang, D.L.; Wang, Z.; Zhang, Z.; Zheng, R.; Yang, L.; et al. Multigeneration analysis reveals the inheritance, specificity, and patterns of CRISPR/Cas-induced gene modifications in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 4632–4637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Wang, G.; Ma, S.; Xie, X.; Wu, X.; Zhang, X.; Wu, Y.; Zhao, P.; Xia, Q. CRISPR/Cas9-mediated targeted mutagenesis in Nicotiana tabacum. Plant Mol. Biol. 2014, 87, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Zheng, A.; Jiang, M.; Xue, S.; Yuan, Q.; Jiang, L.; Chen, Q.; Li, M.; Wang, Y.; Zhang, Y.; et al. CRISPR/Cas9-mediated mutagenesis of homologous genes in Chinese kale. Sci. Rep. 2018, 8, 16786. [Google Scholar] [CrossRef] [PubMed]
- Arndell, T.; Sharma, N.; Langridge, P.; Baumann, U.; Watson-Haigh, N.S.; Whitford, R. gRNA validation for wheat genome editing with the CRISPR-Cas9 system. BMC Biotechnol. 2019, 19, 71. [Google Scholar] [CrossRef]
- Zhang, H.; Cao, Y.; Zhang, H.; Xu, Y.; Zhou, C.; Liu, W.; Zhu, R.; Shang, C.; Li, J.; Shen, Z.; et al. Efficient generation of CRISPR/Cas9-mediated homozy-gous/biallelic Medicago truncatula mutants using a hairy root system. Front. Plant Sci. 2020, 11, 294. [Google Scholar] [CrossRef]
- Butler, N.M.; Jansky, S.H.; Jiang, J. First-generation genome editing in potato using hairy root transformation. Plant Biotechnol. J. 2020, 18, 2201–2209. [Google Scholar] [CrossRef]
- Shu, H.; Luo, Z.; Peng, Z.; Wang, J. The application of CRISPR/Cas9 in hairy roots to explore the functions of AhNFR1 and AhNFR5 genes during peanut nodulation. BMC Plant Biol. 2020, 20, 417. [Google Scholar] [CrossRef]
- Jacobs, T.B.; Lafayette, P.R.; Schmitz, R.J.; Parrott, W.A. Targeted genome modifications in soybean with CRISPR/Cas9. BMC Biotechnol. 2015, 15, 16. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.; Powell, G.; Moore, K.; Abbott, A. The high oleate trait in the cultivated peanut (Arachis hypogaea L.) II. Molecular bais and genetics of the trait. Mol. Gen. Genet. 2000, 263, 806–811. [Google Scholar] [CrossRef]
- Lopez, Y.; Nadaf, H.L.; Smith, O.D.; Connell, J.P.; Reddy, A.S.; Fritz, A.K. Isolation and characterization of the Delta (12) fatty acid desaturase in peanut (Arachis hypogaea L.) and search for polymorphisms for the high oleate trait in Spanish market-type lines. Theor. Appl. Genet. 2000, 101, 1131–1138. [Google Scholar] [CrossRef]
- Chu, Y.; Holbrook, C.C.; Ozias-Akins, P. Two Alleles of ahFAD2B Control the High Oleic Acid Trait in Cultivated Peanut. Crop Sci. 2009, 49, 2029–2036. [Google Scholar] [CrossRef] [Green Version]
- Nawade, B.; Bosamia, T.C.; Thankappan, R.; Rathnakumar, A.L.; Kumar, A.; Dobaria, J.R.; Kundu, R.; Mishra, G.P. Insights into the Indian Peanut Genotypes for ahFAD2 Gene Polymorphism Regulating Its Oleic and Linoleic Acid Fluxes. Front. Plant Sci. 2016, 7, 1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Construct | No of Independent Roots Screened | Edited Roots | Editing Efficiency (EE) | EE in FAD2A | EE in FAD2B | Biallelic Efficiency |
|---|---|---|---|---|---|---|
| pDW3872/Cas9-extended scaffold | 25 | 8 | 32.0% | 20.8% | 30.4% | 16.0% |
| pDW3877/Cas9-regular scaffold | 34 | 8 | 23.5% | 17.7% | 17.7% | 11.8% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neelakandan, A.K.; Wright, D.A.; Traore, S.M.; Ma, X.; Subedi, B.; Veeramasu, S.; Spalding, M.H.; He, G. Application of CRISPR/Cas9 System for Efficient Gene Editing in Peanut. Plants 2022, 11, 1361. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101361
Neelakandan AK, Wright DA, Traore SM, Ma X, Subedi B, Veeramasu S, Spalding MH, He G. Application of CRISPR/Cas9 System for Efficient Gene Editing in Peanut. Plants. 2022; 11(10):1361. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101361
Chicago/Turabian StyleNeelakandan, Anjanasree K., David A. Wright, Sy M. Traore, Xingli Ma, Binita Subedi, Suman Veeramasu, Martin H. Spalding, and Guohao He. 2022. "Application of CRISPR/Cas9 System for Efficient Gene Editing in Peanut" Plants 11, no. 10: 1361. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101361